

Abstract
We have evaluated the influence of the secretory phenotype of presynaptic boutons on the accumulation of postsynaptic glycine receptors (GlyRs), type A GABA receptors (GABAARs), and gephyrin clusters. The cellular distribution of these components was analyzed on motoneurons cultured either alone or with glycinergic and/or GABAergic neurons. In motoneurons cultured alone, we observed gephyrin clusters at nonsynaptic sites and in front of cholinergic boutons, whereas glycine and GABAA receptors formed nonsynaptic clusters. These receptors are functionally and pharmacologically similar to those found in cultures of all spinal neurons. Motoneurons receiving GABAergic innervation from dorsal root ganglia neurons displayed postsynaptic clusters of gephyrin and GABAARβ but not of GlyRα/β subunits. In motoneurons receiving glycinergic and GABAergic innervation from spinal interneurons, gephyrin, GlyRα/β, and GABAARβ formed mosaics at synaptic loci. These results indicate that (1) the transmitter phenotype of the presynaptic element determines the postsynaptic accumulation of specific receptors but not of gephyrin and (2) the postsynaptic accumulation of gephyrin alone cannot account for the formation of GlyR-rich microdomains.
Keywords: dorsal root ganglia, GABAA receptor, gephyrin, glycine receptor, motoneurons, picrotoxinin, presynaptic innervation, spinal neurons, strychnine
In the nervous system, certain ionotropic receptors accumulate at postsynaptic membrane-facing terminals enriched in the corresponding neurotransmitter. Such is the case for GlyR (Triller et al., 1985, 1987; Van den Pol and Gorcs, 1988), GABAAR α6 and GABAAR β3 subunits (Baude et al., 1992; Todd et al., 1996), and ionotropic glutamate receptors (Petralia and Wenthold, 1992; Craig et al., 1993; Aoki et al., 1994; Petralia et al., 1994). Other GABAA and glutamate receptor subtypes are also present at nonsynaptic sites (Baude et al., 1992, 1994, 1995; Sur et al., 1995a). On the cytoplasmic side, peripheral membrane proteins bind and contribute to the clustering of various types of receptors. These include gephyrin for GlyR and some GABAAR subtypes [see references in Vannier and Triller (1997) and Essrich et al. (1998)] and PDZ domain (initially identifed in PSD95, Disc Large, and Zonula Occludens 1 proteins) molecules for ionotropic and metabotropic glutamate receptors [see references in Sheng and Wyszynski (1997)]. These molecules are functionally homologous to rapsyn, which stabilizes the muscular nicotinic acetylcholine receptor (nAChR) [see references in Sanes (1997)]. Peripheral proteins anchor receptors by direct or indirect attachment to perisynaptic or subsynaptic cytoskeletal elements. An unresolved question is that of the involvement of presynaptic innervation in postsynaptic accumulation of transmitter receptors and associated proteins. We have examined this question for the postsynaptic accumulation of GlyR, GABAAR, and gephyrin in spinal motoneurons.
Glycine and GABA are involved in postsynaptic inhibition through the activation of the chloride (Cl−) channels associated with GlyR and GABAAR. Both receptors belong to the nAChR superfamily (Betz, 1990). GlyR is a pentamer composed of α and β subunits (α3β2) [see references in Vannier and Triller (1997)]. Gephyrin links GlyR through its β subunit to the subsynaptic cytoskeleton (Kirsch and Betz, 1995; Meyer et al., 1995). GABAARs are also pentamers. The α2, α3, α5, β3, and γ2 subunit mRNAs are detected in motoneurons (Persohn et al., 1991; Wisden et al., 1991). A recent study has shown that gephyrin is involved in the stabilization of GABAAR in the postsynaptic membrane (Essrich et al., 1998).
We have analyzed the distribution of gephyrin, GlyRα/β, and GABAARβ2/3 subunits on cultured motoneurons: (1) alone, establishing connections among themselves and therefore receiving cholinergic inputs; (2) with dorsal root ganglia (DRG) neurons to supply a GABAergic innervation, taking advantage of the fact that a subclass of DRG neurons expresses a GABAergic phenotype (Roy et al., 1991; Chauvet et al., 1995); or (3) with spinal interneurons to provide glycinergic and GABAergic innervations. We show that in the absence of inhibitory presynaptic innervation, motoneurons express GlyR and GABAAR that are functional and form nonsynaptic clusters. In the presence of presynaptic innervation, these receptors accumulate specifically under boutons containing the corresponding neurotransmitter. In contrast, gephyrin accumulates under all synaptic terminals whether they contain acetylcholine (ACh), GABA, or glycine. Our results demonstrate that gephyrin alone is not able to sort GlyR and GABAAR to synaptic sites and that glycinergic and GABAergic endings provide specific signals for accumulation of the corresponding receptors.
MATERIALS AND METHODS
Motoneuron purification and culture. Motoneurons were purified from ventral spinal cords of embryonic day 15 Sprague Dawley rats (Janvier, Le Genest-sur-Isle, France) by a combination of metrizamide density-gradient centrifugation and immunopanning on dishes coated with the 192 antibody as previously described (Henderson et al., 1995). This antibody recognizes the low-affinity neurotrophin receptor p75NTR, which is selectively expressed by motoneurons at this developmental stage (Chandler et al., 1984;Yan and Johnson, 1988) (Developmental Studies Hybridoma Bank, Iowa City, IA). Purified motoneurons (2 × 103) were plated on polyornithine-laminin (Sigma, St. Louis, MO) precoated 12 mm coverslips in four-well plates (Nunc, Dannstadt, Germany). Culture medium was the Neurobasal-B27 medium combination (Life Technologies, Gaithersburg, MD) supplemented with 0.5 mml-glutamine (Life Technologies), 25 μm β-mercaptoethanol (Life Technologies), and 2% horse serum (v/v; Sigma). For certain experiments, motoneurons were cultured in the glycine-free MEM medium (Life Technologies) supplemented with vitamins and essential amino acids (except glycine) (Life Technologies), 1 mm glucose (Sigma), B27, 0.5 mml-glutamine (Life Technologies), 25 μm β-mercaptoethanol (Life Technologies), and 2% horse serum (v/v; Sigma). The amount of glycine in the horse serum was found to be 229 μm (Beckman 6300 amino acid analyzer), which gave a final concentration of 4.6 μm. To keep motoneurons alive, 1 ng/ml recombinant rat ciliary neurotrophic factor (CNTF; Peprotech, London) (Arakawa et al., 1990) and 100 pg/ml recombinant human glial cell line-derived neurotrophic factor (GDNF; Peprotech) (Henderson et al., 1994) were added to the culture medium. Cultures were kept at 37°C in 7.5% CO2 for up to 11 d in vitro(DIV). The culture medium was renewed every 4 d.
Motoneuron and DRG neuron cocultures. DRG were dissected from embryonic day 15 (E15) mice, collected in PBS (120 mm, pH 7.4), and centrifuged for 5 min at 800 ×g. The supernatant was discarded and replaced by the Neurobasal-B27 medium with 0.2% horse serum. One or two whole DRG explants were added to purified motoneurons plated the day before. The culture medium was completely replaced after 4 d, and cultures were maintained for up to 7 DIV.
Motoneuron and spinal interneuron cocultures. Primary cultures of spinal cord neurons were prepared from E15 Sprague Dawley rats as described previously (Béchade et al., 1996; Lévi et al., 1998). In parallel, ventral spinal cords were dissected and dissociated, and neurons were centrifuged on a 6.5% metrizamide cushion. The interface of the metrizamide cushion enriched in large motoneurons was collected. Neurons (2 × 103) from the latter fraction were mixed with 105 spinal neurons and plated on 12 mm glass coverslips. Cultures were maintained in the Neurobasal-B27 medium with 0.2% horse serum for up to 11 DIV.
Antibodies. The mAbs used in this study were as follows: (1) mAb7a, which recognizes gephyrin (1:200; Boehringer Mannheim, Mannheim, Germany) (Pfeiffer et al., 1984). Splice variants of gephyrin transcripts are widely expressed in the CNS (Kirsch et al., 1993a). Immunocytochemistry with mAb7a showed the same widespread distribution (Kirsch and Betz, 1993), suggesting that mAb7a has a broad spectrum of recognition for gephyrin. However, some heavy forms of gephyrin may not be detected by mAb7a (Kawasaki et al., 1997); (2) mAb4a, which binds to all GlyR α and β subunit isoforms (1:100; gift from H. Betz) (Pfeiffer et al., 1984; Schröder et al., 1991); (3) mAb bd17 directed against the GABAA Rβ2/3 subunits (10 μg/ml; Boehringer Mannheim) (Richards et al., 1987); (4) anti-Islet1 homeodomain protein (clone 2D6, 1:2; Developmental Studies Hybridoma Bank) (Ericson et al., 1992; Tsuchida et al., 1994); (5) anti-155 kDa rat neurofilament protein that specifically stains axons (clone 2H3, 1:2000; Developmental Studies Hybridoma Bank) (Dodd et al., 1988); (6) AP14, which detects MAP2A (1:50; gift from B. Riederer) (Binder et al., 1984); and (7) anti-synaptophysin (1:20; Boehringer Mannheim). As secondary antibodies, we used a carboxymethyl indocyanine-3 (CY3)-coupled affinity-purified goat anti-mouse IgG (1:200; Jackson Immunoresearch Laboratories, West Grove, PA). Polyclonal antibodies (pAbs) were also used: (1) rabbit anti-synapsin I (1:2000; gift from P. de Camilli) (De Camilli et al., 1983); (2) rabbit anti-glutamate decarboxylase of 67 kDa (GAD67, 1:2000; Chemicon, Temecula, CA) (Wong et al., 1974); (3) rabbit anti-MAP2 (1:250; Sigma); (4) rabbit antiserum raised against the rat GABAARβ3 subunit (5 μg/ml) (Todd et al., 1996); (5) rabbit anti-glutamate receptor 1 (GluR1, 10 μg/ml; Chemicon) (Wenthold et al., 1992); and (6) goat anti-choline acetyltransferase (ChAT, 1:500; Chemicon). The rabbit pAbs were recognized by a fluorescein (FITC)-conjugated affinity-purified goat anti-rabbit IgG (H+L) (1:200; Jackson Immunoresearch Laboratories). For double-staining of gephyrin and ChAT, mAb7a was recognized by an FITC-conjugated affinity-purified horse anti-mouse IgG (H+L) (1:200; Vector Laboratories, Burlingame, CA), and the goat pAb anti-ChAT was recognized by a CY3-conjugated affinity-purified donkey anti-sheep IgG (H+L) (1:200; Jackson Immunoresearch Laboratories).
Immunocytochemistry. For all immunodetections except that of mAb4a, cells were fixed with 4% (w/v) paraformaldehyde for 15 min. Cells were washed in PBS and permeabilized with 0.12% Triton X-100 in PBS with 0.12% (w/v) gelatin for 5 min. The permeabilized cells were incubated with primary antibodies overnight at 4°C. The next day, cells were washed in PBS and incubated with secondary fluorescent antibodies for 45 min at room temperature. For mAb4a immunocytochemistry, a methanol/acetic acid (95:5) mixture was used as fixative (10 min at −20°C). Cells were rinsed in PBS and incubated successively in primary and secondary antibodies as described above. For multiple-labeling experiments, antibodies were incubated simultaneously. For immunoperoxidase reactions, cells were incubated successively with the Islet1 mAb and the secondary biotinylated affinity-purified horse anti-mouse IgG (H+L) (1:200; Vector Laboratories) for 1 hr, rinsed in PBS, and then incubated with the ABC complex (1:200; Vector Laboratories) for 30 min. Peroxidase staining was obtained by incubating cells in a DAB–H2O2 reactant (Sigma). Peroxidase reactions were monitored under the microscope and stopped by washing with PBS.
For sequential mAb7a and mAb4a immunodetections, cells were fixed in a methanol/acetic acid (95:5) solution (10 min at −20°C), rinsed, and incubated with mAb4a (1:100, 1 hr). After washes, cells were treated with lissamine rhodamine-conjugated affinipure Fab fragment goat anti-mouse IgG (H+L) (1:50, 45 min; Jackson Immunoresearch Laboratories). They were then rinsed and incubated with unconjugated Fab fragment goat anti-mouse IgG (H+L) (1:50, 45 min; Jackson Immunoresearch Laboratories) to saturate still unbound mAb4a. Then they were incubated with mAb7a (1:200, 1 hr), which was further recognized by an FITC-conjugated affinity-purified horse anti-mouse IgG (H+L) (1:200, 45 min; Vector Laboratories). All incubations were performed at room temperature. Control cells were exposed to the same treatment except that they were incubated with only one of the primary antibodies.
In all experiments, the specificity of immunolabeling and the absence of antibody cross-reaction in double-staining experiments were controlled by omission of the primary antibodies. Cultures were observed with a standard or a confocal (Molecular Dynamics, Sunnyvale, CA) epifluorescence microscope. Fluorescent images were acquired on a Hamamatsu CCD camera (C5985) mounted on a Leica DMR/HCS microscope (objective 40 or 63×), and dual color images were obtained using Imagespace software (Molecular Dynamics). Images were prepared for printing using Adobe Photoshop software.
Quantitative analysis. Motoneuron purity was estimated by counting under phase contrast (objective 40×) the number of cells with nuclear peroxidase staining of Islet1. The proportions of cells displaying gephyrin-, GlyRα/β-, and GABAARβ3–immunoreactivity (IR) were determined by visual inspection using a standard fluorescence microscope (objective 63×). For each experiment, 36 ± 3 fields were analyzed. For these quantifications, data are expressed as means ± SEM of three independent experiments. Colocalization of gephyrin, GlyRα/β, or GABAARβ2/3 with synapsin was determined from double-staining experiments of synapsin and mAb7a, mAb4a, or mAbbd17. The quantifications were performed on images acquired with a Hamamatsu CCD camera and specific filters for FITC and CY3 fluorescences (Leica). This procedure was chosen because it allows a minimal bleed-through between FITC and CY3 channels, which was always found to be lower than 0.3%. Confocal microscopy was used to measure the surface area of gephyrin, GlyRα/β, and GABAARβ3 clusters. To improve accuracy, the quantification was performed by means of simple staining experiments. Immunopositive cells were displayed in the center of the digitized field, and all neurites within the field were computed. Excitation was obtained with an argon ion laser set at 514 nm for CY3 excitation, and the emitted light was filtered with a long-pass filter (530 nm). Pixel size and focus steps were 0.21 and 0.3 μm, respectively (objective 63×, numerical aperture 1.4), with images of 512 × 512 pixels. Digitized series of optical sections at different planes of focus were collected using a host computer (Indy, Silicon Graphics). The background noise was reduced, and the contrast was enhanced by applying a median (3 × 3 × 3) Gaussian filter. Maximum intensity projections were derived from these sections using Imagespace (Molecular Dynamics) software. The surface area of clusters was determined with NIH 1.52 software. The threshold intensity fluorescence was set manually for each cell to insure efficient detection and to avoid coalescence of clusters. An image-object was computed if it comprised at least three pixels. This analysis was performed on motoneurons cultured alone or in the presence of spinal interneurons, as well as on interneurons. Statistical analysis was performed using StatView F.4.11 software.
Electrophysiology. The experiments were performed at room temperature (20–23°C) between 6 and 8 DIV in the whole-cell configuration of the patch-clamp technique. Before recording, the culture medium was replaced by the external solution to be used during the experiment, which contained (in mm): 150 NaCl, 2.5 KCl, 1.8 CaCl2, 1 MgCl2, 20 glucose, and 10 HEPES-NaOH, pH 7.4. The internal solutions used to fill the recording electrode contained (in mm): either 145 CsCl, 1 MgCl2, 10 EGTA, 1 CaCl2, 3 ATP-Mg, 0.3 GTP-Na, and 10 HEPES-CsOH, pH 7.2, for internal solution A or 145 Cs methanesulfonate, 15 CsCl, 1 MgCl2, 0.1 EGTA, 3 ATP-Mg, 0.3 GTP-Na, and 10 HEPES-CsOH, pH 7.2, for internal solution B. The culture dish was continuously perfused with the external solution. In addition, a fast perfusion system was used for rapid applications of glycine and modulators onto the recorded cell. All solutions applied via this system contained 0.2 μmtetrodotoxin. This system was made of glass syringes, Teflon taps, and Teflon tubing connected to two parallel glass barrels (one of them only containing glycine); lateral movements of this system were controlled by a computer-driven motor to apply the solution of the desired barrel to the cell [continuously perfused with one of the solutions of this system; for more details, see Chesnoy-Marchais (1996)]. For concentrations of glycine below 40 μm, successive responses recorded under identical conditions were quite stable when tested every 40 sec. For higher concentrations (80–160 μm), the interval between successive tests was longer (100 sec) to allow for recovery from desensitization. Stock solutions of strychnine (hemisulfate, Sigma) and picrotoxinin (Sigma) were prepared at 1 mm in water and at 100 mm in ethanol, respectively. In the experiments using picrotoxinin, the solutions applied by fast perfusion contained 1:1000 ethanol. When a modulator was applied “with preincubation,” it was applied continuously between and during the successive glycine applications, that is, in both barrels of the fast perfusion. When applied “without preincubation,” it was present only in the glycine-containing barrel.
Patch-clamp micropipettes were made from hard glass (Kimax 51); the shank of each pipette was covered with Sylgard, and the tip was fire-polished. Their resistance was close to 5 MΩ. The cells were voltage-clamped by an EPC7 List amplifier, which was controlled by a TANDON 38620 computer, via a Cambridge Electronic Design (CED) 1401 interface, using CED patch- and voltage-clamp software. The current monitor output of the amplifier was filtered at 0.3 kHz before being sampled on-line at 0.6 kHz. The bath was connected to the ground via an agar bridge. Membrane potentials were corrected for junction potentials. The series resistance was routinely measured and compensated. The zero indicated on the current traces is the absolute zero current level.
RESULTS
Gephyrin, GlyR, and GABAAR cellular distribution in motoneurons cultured alone
Motoneurons were purified by a combination of metrizamide density-gradient and immunopanning techniques (Henderson et al., 1995). As estimated by immunoperoxidase staining of the motoneuronal embryonic marker Islet1 (Ericson et al., 1992), 89.2 ± 1.3% of the cells had a motoneuronal phenotype (Fig.1A). The remaining cells corresponded to Islet1-negative but p75 low-affinity neurotrophin receptor- and L14 lectin-positive motoneurons (Henderson et al., 1993). Double-immunofluorescence experiments with antibodies against MAP2 and 155 kDa neurofilament protein were performed to label dendrites and axons, respectively. These experiments showed that at 7 DIV, motoneurons are polarized with somato-dendritic and axonal compartments (Fig. 1B). We used the mAb7a and mAb4a antibodies to stain gephyrin and GlyRα/β subunits, respectively (Pfeiffer et al., 1984), whereas GABAARβ3 subunit was recognized by a previously characterized pAb (Todd et al., 1996). The gephyrin, GlyRα/β, and GABAARβ3 formed puncta on somata and dendrites as shown in double-labeling experiments with anti-MAP2 antibodies (Fig. 1C–E). A few scattered spots of gephyrin were also detected along the length of some axons (data not shown). Clusters of GlyRα/β and GABAARβ3 were not detected in axons. However, in some neurons, clusters of GlyRα/β (Fig. 1D) or GABAARβ3 (Fig. 1E) were detected at the level of the axon-hillock. Confocal optical sections revealed small, round or ellipsoidal clusters of gephyrin at membrane areas in contact with the coverslip (Fig. 1F1exemplifies a neuron with a large number of clusters) as well as at distance from it (Fig. 1F2). Numerous bright spots of GlyRα/β (Fig. 1G1) and GABAARβ3 (data not shown) were also detected at the substrate–neuron interface as well as at a distance from it. On some cells, GlyRα/β-IR (Fig.1G2) and GABAARβ3-IR (Fig.1K1, arrow) formed large patches at the neuronal surface.
Fig. 1.

Gephyrin, GlyR, and GABAAR distribution on motoneurons cultured alone. A, Immunoperoxidase showing the nuclear staining of the embryonic motoneuronal marker Islet1. B, Double-staining of the dendritic protein MAP2 (green) and the axon-enriched 155 kDa neurofilament protein (red).C–E, Presence of clusters (arrows) of gephyrin (C, red), GlyRα/β (D, red), or GABAARβ3 (E, green) on MAP2-IR somato-dendritic compartment (green in C, D;red in E). GlyRα/β clusters are also accumulated at the level of axon-hillock (arrowhead inD). F, G, Confocal visualization of Geph-IR (F1–2) and GlyRα/β-IR (G1–2), respectively. Discontinuous Geph-IR (arrows) at the cell-to-substrate contact (F1) and at a distance from the coverslip (F2). GlyRα/β forms clusters at the cell-to-substrate contact (arrows in G1) and occasionally displays a continuous labeling (arrows in G2) on sections passing through the nucleus. H, I, Double-immunofluorescence showing that Geph-IR clusters (H1, I1) accumulate in front of synapsin-IR boutons (arrowheads in H2) displaying ChAT-IR (arrowheads in I2). Geph-IR clusters (H1, I1) are also detected at nonsynaptic sites (arrows). In contrast, GlyRα/β (J1) and GABAARβ2/3 (K1) do not concentrate at synaptic sites (arrowheads in J1–2 andK1–2, respectively). Insets inH2, I2, J2, and K2 show superimposed images of H1–H2, I1–I2, J1–J2, andK1–K2, respectively. ChAT, Choline acetyltransferase;GABAARβ2/3, GABAARβ2/3 subunits-IR;GABAARβ3, GABAARβ3 subunits-IR; Geph, gephyrin-IR;GlyRα/β, GlyRα/β subunits-IR;MAP2, dendritic marker; NF, NF155 Kda-IR;Syn, synapsin-IR. Scale bar: A,B, 50 μm; C–K2, 10 μm.
The relationships of these receptor components to presynaptic endings were analyzed in double-staining experiments in which the synaptic boutons were identified with a pAb against synapsin. For these experiments, we used the mAb bd17 directed against GABAARβ2/3 subunits (Richards et al., 1987) because the anti-synapsin Ab is a pAb. Synapsin-positive endings were detected over somata and/or dendrites of many motoneurons and could correspond to autapses or may originate from other motoneurons. On these cells, gephyrin-IR clusters were found on the soma and neurites at synaptic and nonsynaptic loci (Fig.1H1–H2). Almost all presynaptic specializations were facing gephyrin-IR clusters. These postsynaptic gephyrin-IR clusters were larger than those observed at nonsynaptic loci and extend all along the synaptic contact (Fig.1H1). To verify that presynaptic boutons in these pure cultures of motoneurons were cholinergic, we immunostained for ChAT (Fig. 1I1–2). We found that after 11 DIV, almost all boutons were ChAT-positive and that 90.2 ± 3.8% (10 cells encompassing 199 boutons) of these boutons were apposed to gephyrin-IR clusters (Fig. 1I1–2). In contrast, most GlyRα/β (Fig. 1J1–2) and GABAARβ2/3 (Fig.1K1--2) clusters were detected at the cell surface at sites that were not facing presynaptic boutons. Because glycine was present at high concentration (400 μm) in the culture medium, it may be involved in the clustering of gephyrin. To examine this issue, motoneurons were cultured in glycine-depleted medium. This medium was not completely glycine free because the added serum contributed to a final concentration of 4.6 μm (see Materials and Methods). However, this low concentration is shown to activate <2% of the GlyR channels (see Fig. 3). With this low amount of glycine, we found that motoneurons had postsynaptic and nonsynaptic clusters of gephyrin (data not shown), similar to motoneurons cultured in a glycine-rich medium.
Fig. 3.

Comparison of the functional and pharmacological properties of GlyRs in cultures of purified motoneurons and all spinal neurons. A–C, Concentration–response curves in both types of cultures (purified motoneurons: A,C, bars labeled MN; cultures of all spinal neurons: B, C, bars labeledSN). The internal solution and membrane potential were solution A and −20 mV (A–C, bars labeledMN2 and SN) or solution B and −70 mV(C, bars labeled MN1). The left traces in A or B show superimposed current recordings obtained during successive applications of glycine at increasing concentrations (10–160 μm). Theright graph in A or B(obtained from the corresponding left traces) shows for each cell the fit of the concentration dependence of the peak response by a Hill equation: y =Imax/(1 + (EC50/x)nH). The values and corresponding errors given by the computer for EC50, nH, andImax for each of these cells were, respectively, 46.7 ± 3.4 μm, 1.92 ± 0.16 and 0.744 ± 0.035 nA in A, and 47.7 ± 0.9 μm, 2.16 ± 0.06 and 0.833 ± 0.011 nA inB. C, Mean values and SD for these three parameters, derived from several such experiments performed in different cells under identical conditions. D, Voltage dependence of the responses of purified motoneurons to glycine (internal solution A). Glycine (10 μm) was applied for 2 sec at different test potentials, using long voltage jumps from the holding potential (−20 mV) toward the test potential (the voltage jump beginning 0.8 sec before each glycine application). Left graphs,Normalized I–V curves for three different motoneurons, obtained by dividing each glycine response by the response of the corresponding cell at −60 mV. Right traces, Records obtained in one of these cells during voltage jumps to −80 and +80 mV.E, Strychnine sensitivity of glycine responses (internal solution A, holding potential −20 mV). Left panel, Mean values of the percentage of inhibition of the peak glycine responses by strychnine (50 or 500 nm applied with preincubation) in cultures of purified motoneurons (white bars) and in cultures of all spinal neurons (hatched bars). The glycine concentration was either 40 μm (close to the EC50) or 100 μm. Middle panel, Effect of strychnine applied with preincubation first at 50 nm, then at 500 nm, on the response to glycine (40 μm) of a purified motoneuron (MN). Right panel, Effect of 500 nm strychnine, applied first without preincubation, then with preincubation, on the response to glycine (100 μm) in a culture of all spinal neurons (SN).
The question remains of the association of large gephyrin clusters postsynaptic to cholinergic synapses with GlyRα/β-IR. This was approached by sequential detection of gephyrin (Fig.2A1) and GlyRα/β (Fig. 2A2) on motoneurons cultured alone for 7 DIV. We found that large gephyrin-IR clusters were not associated with GlyRα/β-IR, suggesting that gephyrin microdomains postsynaptic to cholinergic afferences do not recruit GlyRα/β at this site. In contrast, many small, round-shaped clusters of gephyrin found at nonsynaptic loci colocalized with GlyRα/β-IR clusters of comparable size and shape.
Fig. 2.

Simultaneous detection of gephyrin-IR (A1, A3, A5) and GlyRα/β-IR (A2, A4, A6) on motoneurons cultured alone for 7 DIV. Motoneurons display large and small, round-shaped Geph-IR clusters. Large Geph-IR clusters do not colocalize with GlyRα/β-IR (crossed arrows). Most (arrowheads) but not all (arrows) small Geph-IR clusters colocalize with GlyRα/β-IR clusters.Geph, Gephyrin-IR; GlyRα/β, GlyRα/β subunits-IR. A1–2, Pairs of digitized images acquired with CCD camera (A1, FITC channel;A2, TRITC channel). A3–4, A5–6, Higher magnification of the plain and dotted outlined regions in A1–2, respectively. Scale bar:A1–2, 10 μm; A3–4,A5–6, 1.2 μm.
Our data indicate that in cultured motoneurons that do not receive glycinergic and GABAergic functional innervation, one can observe clusters of gephyrin, GlyRα/β, and GABAARβ2/3. Gephyrin accumulates in front of synaptic cholinergic boutons, and these postsynaptic gephyrin clusters hardly recruit GlyRα/β and GABAARβ2/3 (for quantification, see Fig. 8).
Fig. 8.

Quantifications of the synaptic localization of gephyrin, GlyR, and GABAAR on motoneurons cultured alone (A1–3) or with spinal interneurons (B1–3). In each case, the open bars give the proportion of synapses with the indicated postsynaptic IR (Gephyrin, GlyRα/β, orGABAARβ2/3), and the filled bars give the proportion of nonsynaptic clusters per cell. Gephyrin, GlyRα/β, and GABAARβ2/3 clusters were classified as synaptic when adjacent to synapsin-IR. Results are means (± SEM) from 10–18 cells. The levels of significance (ANOVA, Scheffé F test) are indicated by one (p < 0.05), two (p < 0.01), or three (p < 0.001) symbols. ∉, Significance between 3 and 7 DIV; ¥, significance between 7 and 11 DIV; $, significance between 3 and 11 DIV.
Functional and pharmacological properties of GlyR and GABAAR in motoneurons cultured alone
In an attempt to understand the different localizations of GlyRs and gephyrin in motoneurons cultured alone (in particular the absence of GlyRs at gephyrin-rich synaptic loci), we investigated the functional and pharmacological properties of the receptors of these cells. Glycine (applied at 10–320 μm by a fast perfusion system) evoked responses in all the motoneurons recorded in the whole-cell configuration of the patch-clamp technique (n > 50). A concentration–response curve obtained from a single motoneuron is illustrated in Figure 3A. The response (recorded here for an inward driving force of 20 mV only) was already detectable at 10 μm glycine. Raising the concentration increased the peak response and revealed some desensitization. For each cell in which a complete curve was obtained, the concentration dependence of the peak response was fitted by a Hill equation (Fig. 3A, right graph and legend). The EC50, Hill coefficient (nH), and maximum response (Imax) values thus derived were then averaged for two series of motoneurons (Fig. 3C, bars labeled MN1 and MN2; see legend); EC50 and nH were always close to 40 μm and 2, respectively. For comparison, identical experiments (Fig. 3B) were performed on cultured neurons from the whole spinal cord (spinal neuron). The physiological properties of neurons in both types of cultures were almost identical (Fig. 3C). For example, the mean EC50 and nH values found using internal solution A were 37.1 ± 7.4 μm (4) and 1.92 ± 0.19 (4) for purified motoneurons (MN2), and 38.3 ± 8.5 μm (6) and 2.25 ± 0.20 (6) for cultures of all spinal neurons (SN), respectively. The Imax values were more variable and usually slightly larger for purified motoneurons, which can be explained by their greater surface area.
The voltage-sensitivity of glycine responses of motoneurons cultured alone was investigated. The I–V curves of the responses to 10 μm glycine obtained from three motoneurons with symmetrical chloride concentrations are illustrated in Figure3D. As shown by the plot of the normalized responses of these cells and by the records obtained in one of them for two symmetrical driving forces, the voltage dependence was clearly nonlinear, favoring outward responses. The reversal potential was close to ECl (here 0 mV); this was confirmed in an experiment using asymmetrical chloride concentrations (internal solution B), bringing the reversal potential to −50 mV (data not shown).
The blocking effect of the classical GlyR antagonist strychnine was investigated in the two types of cultures. In motoneurons cultured alone, when strychnine was applied with preincubation (both between and during glycine applications) at either 50 or 500 nm, the peak responses to 40 μm glycine were reduced by 52.8 ± 5.3% (5) or 96.2 ± 1.1% (5), respectively (Fig.3E, white bars and records labeledMN). The responses recorded in cultures of all spinal neurons showed a similar strong sensitivity to strychnine (Fig.3E, hatched bars). For all neurons studied, when strychnine was applied without preincubation (i.e., only with glycine), its blocking effect was apparently much lower. This is illustrated for the effect of 500 nm strychnine on the response to 100 μm glycine in a culture of all spinal neurons (Fig. 3E, traces labeled SN), and the results were confirmed in four similar experiments (relatively small blockade between 21 and 44%). The requirement for preincubation with strychnine was confirmed in motoneurons cultured alone (the peak response to 40 μm glycine, which could be completely blocked by 500 nm strychnine with preincubation, was only reduced by 60 ± 11% (3) without preincubation).
Low sensitivity of glycine responses to picrotoxinin (PTX) is usually considered to be indicative of the presence of β subunits in functional heteromeric receptors (Pribilla et al., 1992; Pistis et al., 1997). Therefore, the blocking effect of a high concentration of PTX (100 μm, applied with preincubation) on the responses to glycine of motoneurons cultured alone was investigated. The responses to 40 μm glycine, recorded at −20 mV in four experiments with internal solution A, were reduced by only 44 ± 10%. In addition, the responses to 100 μm glycine, recorded between −30 and −90 mV in three other experiments with internal solution B, were reduced by only 26–28%, confirming the low sensitivity to PTX of GlyR in motoneurons.
In conclusion, the nonsynaptic GlyRs of motoneurons cultured alone retain the main pharmacological properties of the receptors found in cultures of all spinal neurons.
Cultured neurons from embryonic spinal cord (Ransom et al., 1977), as well as motoneurons from slices of embryonic or neonatal rat spinal cord (Gao and Ziskind-Conhaim, 1995), display chloride responses not only to glycine but also to GABA. The responses to 5 or 10 μm GABA were tested in motoneurons cultured alone (data not shown). Large responses of at least 0.8 nA were recorded for a driving force of only 20 mV (four cells). These responses were little affected (reduced by only 9.5 ± 7.6% in three experiments using 5 μm GABA) by a concentration of strychnine (500 nm applied with preincubation) that almost completely blocked glycine responses. In contrast, these GABA responses were almost completely blocked (result qualitatively confirmed in three cells) by a low concentration of bicuculline (10 μm) known to be ineffective on glycine responses (Lewis and Faber, 1993). Thus motoneurons cultured alone also display functional GABAA receptors.
Cellular distribution of gephyrin, GlyR, and GABAAR in motoneurons cocultured with dorsal root ganglia
Initially, embryonic DRG explants were added to purified motoneurons to provide a glutamatergic presynaptic innervation to motoneurons. To our surprise, we found that a large proportion of DRG neurons were GAD67-positive, indicating the presence of GABAergic neurons in the culture. These results were consistent with previous reports showing that embryonic (Roy et al., 1991; Chauvet et al., 1995) and adult (Schoenen et al., 1989; Roy et al., 1991; Chauvet et al., 1995) DRG neurons express a GABAergic phenotype. In our cultures, the colocalization of GAD67-IR puncta with synaptophysin-IR over Islet1-positive neurons indicate that DRG differentiate GABAergic synapses impinging on motoneurons. Gephyrin formed clusters on somata and neurites of motoneurons that accumulated in front of synapsin-IR profiles (Fig. 4A1–2). Most clusters of gephyrin were adjacent to GAD-positive boutons (Fig.4B1–2), which were also synaptophysin positive (data not shown). Occasionally, gephyrin-IR clusters were detected in front of synaptophysin-positive but GAD-negative terminals (data not shown). On Islet1-positive motoneurons, GABAARβ2/3 were present in front of synapsin-positive boutons (Fig. 4C1–2); some clusters were also detected at nonsynaptic loci as describedin vivo (Todd et al., 1996). Gephyrin was also detected at the level of GABAARβ3 clusters (Fig.4D1–2). The GlyRα/β staining pattern was identical to that observed on motoneurons cultured alone. GlyRα/β-IR clusters were present on the soma and along the length of the dendrites of motoneurons (Fig. 4E1). These GlyRα/β clusters did not form under synapsin-positive varicosities (Fig. 4E1–2) or under GAD-IR blobs (Fig.4F1–2). In some instances, a diffuse GlyRα/β-IR was observed over the soma and dendrites (Fig.4F1).
Fig. 4.
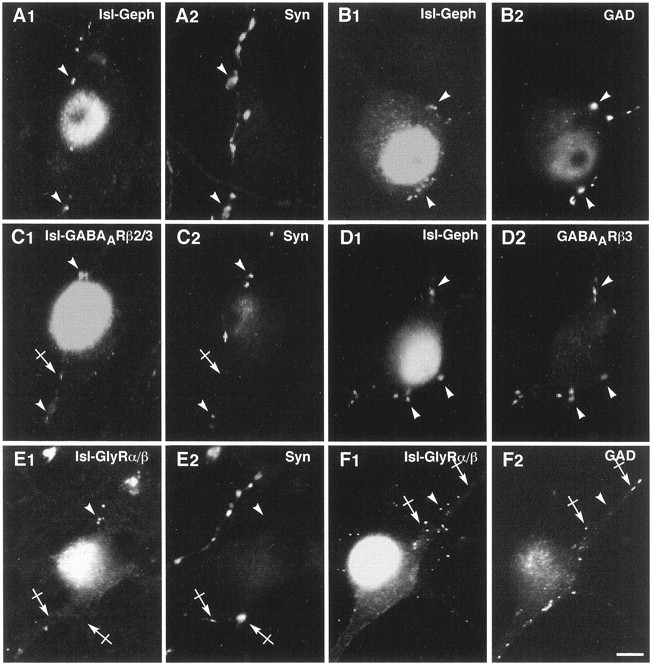
Gephyrin and GABAAR but not GlyR form postsynaptic clusters on motoneurons cocultured with DRG neurons. In all pairs of images, motoneurons are identified by the nuclear Islet1 staining. A, B, Geph-IR is concentrated in front of synapsin-IR boutons (arrowheads in A1–2) displaying GAD-IR (arrowheads in B1–2).C, D, GABAARβ2/3 accumulate at synaptic (arrowheads in C1–2) and nonsynaptic loci (crossed arrow in C1–2). Geph-IR clusters colocalize with GABAARβ3 clusters (arrowheads in D1–2). E, F, GlyRα/β (E1, F1) form clusters (arrowheads) that are not adjacent to synapsin-IR (E1–2, crossed arrows) or GAD-IR (F1–2, crossed arrows) boutons.GABAARβ2/3, GABAARβ2/3 subunits-IR;GABAARβ3, GABAARβ3 subunits-IR; GAD, GAD67-IR;Geph, gephyrin-IR, GlyRα/β, GlyRα/β subunits-IR; Isl, Islet1-IR;Syn, synapsin-IR. A1–A2, B1–B2, C1–C2, D1–D2, E1–E2, F1–F2, Pairs of digitized images acquired with CCD camera (A1, B1, C1, D1, E1, F1, TRITC channel;A2, B2, C2, D2, E2, F2, FITC channel). Scale bar, 10 μm.
These results again indicate that gephyrin accumulates under synaptic boutons, but when these boutons are GABAergic there is also a postsynaptic accumulation of GABAARβ2/3 but not of GlyRα/β.
Cellular distribution of gephyrin, GlyR, and GABAAR in motoneurons cocultured with spinal interneurons
Dissociated spinal interneurons were cocultured with motoneurons to supply glycinergic and GABAergic innervation to motoneurons. In these cultures, motoneurons were identified by the Islet1 nuclear staining. At the somatic and dendritic surface, gephyrin, GlyRα/β, and GABAARβ2/3 formed numerous patches in front of synapsin-IR boutons (Fig.5A1–2, B1–2, C1–2). A few synapsin-stained endings (Fig.5A1–2, B1–2, C1–2, crossed arrows) did not colocalize with these receptor components; they may correspond to excitatory synapses (O’Brien et al., 1997). These data indicate that gephyrin, GlyRα/β, and GABAARβ2/3 clusters present at the periphery of motoneurons innervated by spinal interneurons face synaptic boutons.
Fig. 5.

Gephyrin, GlyR, and GABAAR distribution on motoneurons cocultured with spinal neurons. In all pairs of images, motoneurons are identified by the nuclear Islet1 staining. A–C, Accumulation of Geph (A2), GlyRα/β (B2), and GABAARβ2/3 (C2) in front of most (arrowheads) but not all (crossed arrows) synapsin-IR boutons (A1, B1, C1). D–F, Double-staining of GAD (D1, E1, F1) and Geph (D2); GlyRα/β (E2) or GABAARβ2/3 (F2) show closely apposed signals (arrowheads). Some Geph and GlyRα/β spots are not adjacent to GAD-IR boutons (crossed arrows inD1–2, E1–2, respectively). G, H, Double-immunofluorescence showing that most GABAARβ3-IR spots (arrowheads in G1, H1) are associated with Geph-IR (G2) and GlyRα/β-IR (H2) clusters. Few Geph-IR clusters do not colocalize with GABAARβ3-IR (crossed arrows inG1–2).GABAARβ2/3, GABAARβ2/3 subunits-IR;GABAARβ3, GABAARβ3 subunits-IR; GAD, GAD67-IR;Geph, gephyrin-IR; GlyRα/β, GlyRα/β subunits-IR; Isl, Islet1-IR;Syn, synapsin-IR. A1–A2, B1–B2, C1–C2, D1–D2, E1–E2, F1–F2, G1–G2, H1–H2, Pairs of digitized images acquired with CCD camera (A1, B1, C1, D1, E1, F1, G1, H1, FITC channel; A2, B2, C2, D2, E2, F2, G2, H2, TRITC channel). Scale bar, 10 μm.
Previous reports have shown that in the ventral horn of the spinal cordin vivo, boutons containing both glycine and GABA are presynaptic to gephyrin-IR synapses (Triller et al., 1987; Todd et al., 1996; Colin et al., 1998) and that corelease of these two neurotransmitters occurs (Jonas et al., 1998). Furthermore, some GABAAR and GlyR clusters are colocalized in front of GABAergic terminals (Bohlhalter et al., 1994; Todd et al., 1996). The relationship of gephyrin, GlyRα/β, and GABAARβ2/3 to GABAergic axons was therefore analyzed. Gephyrin (Fig. 5D2) and GlyRα/β (Fig.5E2) clusters were often apposed to GAD-positive terminals (Fig. 5D1, E1). However, gephyrin and GlyRα/β clusters were not always adjacent to GAD-IR profiles, suggesting that some of them are apposed to boutons enriched in glycine only. In contrast, all somato-dendritic GABAARβ2/3-IR clusters detected on motoneurons were apposed to GAD-IR boutons (Fig.5F1–2). Simultaneous experiments in which GABAARβ3 was detected with gephyrin or with GlyRα/β indicated that most but not all GABAARβ3-IR clusters were also immunoreactive for gephyrin (Fig. 5G1–2) or GlyRα/β (Fig.5H1–2). This raised the question regarding whether gephyrin is able to accumulate receptors other than GlyR and GABAAR. This was determined by examining the cellular distribution of the GluR1 subunit of AMPA receptors on neurons cultured for 11 DIV (Fig. 6). The large majority of gephyrin clusters did not colocalize with GluR1 clusters (Fig. 6A1–2). However, we found that 9.8 ± 2.2% (10 cells for a total of 422 gephyrin clusters) of gephyrin clusters were associated with GluR1-IR. Similarly, most GlyRα/β clusters were not stained for GluR1 clusters (Fig.6B1–2). However, 8.0 ± 1.5% (10 cells for a total of 713 GlyRα/β clusters) of GlyRα/β clusters colocalized with GluR1-IR. For this quantification, we selected neurons with large numbers of GluR1 clusters. Interestingly, these neurons had a lower count of gephyrin and GlyR clusters.
Fig. 6.

Comparison of GluR1-IR with gephyrin- or GlyRα/β-IR clusters on spinal interneurons. Spinal neurons immunolabeled at 11 DIV for GluR1 (A1, B1) and Geph (A2) or GlyRα/β (B2). Most Geph- or GlyRα/β-IR (arrowheads) and GluR1-IR (crossed arrows) do not colocalize. Few Geph- and GlyRα/β-IR clusters colocalized with GluR1-IR (arrows).Geph, gephyrin-IR; GluR1, glutamate receptor subunit GluR1; GlyRα/β, GlyRα/β subunits-IR. A1–A2, B1–B2, Pairs of digitized images acquired with CCD camera (A1, B1, FITC channel;A2, B2, TRITC channel). A3–4, B3–4,Higher magnification of a region outlined in A1–2, B1–2, respectively. Scale bar, A1–2, B1–2, 10 μm; A3–4, B3–4, 2.5 μm.
These data indicate that gephyrin, GABAARβ2/3, and GlyRα/β can be detected at the same synapse on motoneurons apposed to GABAergic terminals. Some synapses had only one of these two receptors. Few gephyrin-IR synapses were associated with a noninhibitory receptor subunit, i.e., GluR1.
Quantification of gephyrin, GlyR, and GABAAR-IR
The proportion of cells displaying immunoreactivity for gephyrin, GlyRα/β, and GABAARβ3 was determined at 7 DIV on motoneurons cultured alone or with spinal interneurons (Table1). The immunoreactivity corresponding to GlyRα/β and GABAARβ3 was present at the neuronal surface and had a clustered or diffuse distribution. The latter could only be observed when motoneurons were cultured alone. Diffuse staining of gephyrin was never observed on motoneurons cultured alone, therefore indicating that diffuse GlyRα/β and GABAARβ3 are not associated with gephyrin. We found that the percentage of motoneurons displaying immunoreactivity for either gephyrin or GABAARβ3 increased when interneurons were added to the culture. In contrast, the presence of interneurons in the culture decreased the number of motoneurons with GlyRα/β-IR.
Table 1.
Percentage of motoneurons with gephyrin, GlyRα/β, and GABAARβ3 immunoreactivity
| % of cells with | Motoneurons | Motoneurons + interneurons | ||||
|---|---|---|---|---|---|---|
| GlyRα/β | GABAARβ3 | Gephyrin | GlyRα/β | GABAARβ3 | Gephyrin | |
| Clusters | 71.0 ± 4.6 | 56.2 ± 13.1 | 68.2 ± 3.7 | 63.6 ± 1.9 | 90.8 ± 3.7 | 93.1 ± 1.2 |
| Diffuse | 22.0 ± 3.4 | 26.3 ± 0.3 | n.d. | n.d. | n.d. | n.d. |
| Total | 90.0 ± 3.0 | 76.2 ± 2.5 | 68.2 ± 3.7 | 63.6 ± 1.9 | 90.8 ± 3.7 | 93.1 ± 1.2 |
The number of motoneurons displaying gephyrin-, GlyRα/β-, and GABAARβ3-IR was computed on motoneurons cultured for 7 DIV alone or in the presence of interneurons. Results expressed as percentages (mean ± SEM) were obtained from three independent experiments. The IR could be either diffused or clustered. Totals indicate the percentage of cells with IR independent from its pattern; n.d., IR not detected. Statistical analysis (ANOVA, SchefféF-test) indicates that the presence of interneurons in the culture decreases the number of motoneurons displaying GlyRα/β-IR (p < 0.01), whereas it increases the number of motoneurons with gephyrin-IR (p < 0.001) and GABAARβ3-IR (p < 0.05).
The mean surface areas of gephyrin, GlyRα/β, and GABAARβ3 clusters were determined at 7 DIV on maximum intensity projections of a series of confocal sections spanning the whole thickness of the neuronal somata. The quantifications were performed on motoneurons cultured alone or with interneurons and on interneurons of the latter coculture (Table2). We found that the presence of interneurons increased significantly the surface area of GlyRα/β, GABAARβ3, and gephyrin clusters on motoneurons. On motoneurons cultured alone for 7 DIV, most GlyRα/β and GABAARβ2/3 clusters were nonsynaptic, but when cultured with interneurons they were detected in front of synaptic contacts (see Fig. 8). The GlyRα/β, GABAARβ3, and gephyrin clusters were slightly but significantly smaller on motoneurons cultured with interneurons than on interneurons themselves.
Table 2.
Quantification of GlyRα/β, GABAARβ3, and gephyrin clusters surface areas (μm2)
| Motoneurons | Motoneurons + interneurons | Interneurons | |
|---|---|---|---|
| GlyRα/β | 0.251 ± 0.004∉∉∉ | 0.360 ± 0.009¥ | 0.388 ± 0.007$$$ |
| GABAARβ3 | 0.337 ± 0.010∉∉∉ | 0.502 ± 0.015¥¥¥ | 0.659 ± 0.020$$$ |
| Gephyrin | 0.311 ± 0.008∉∉∉ | 0.423 ± 0.009¥¥¥ | 0.479 ± 0.012$$$ |
Neurons were stained at 7 DIV for gephyrin, GlyRα/β, or GABAARβ3 subunit. For each type of culture and antibody, quantifications were performed on maximum-intensity confocal projections as described in Materials and Methods. Thirty cells were obtained from three independent experiments that were shown to be statistically equivalent (ANOVA, Scheffé F-test). Values are averages ± SEM. The levels of significance (ANOVA, Scheffé F-test) are indicated by one (p < 0.05) or three (p< 0.001) symbols. ∉, Significance between motoneurons cultured alone and motoneurons cultured with interneurons; ¥, significance between interneurons and motoneurons cultured with interneurons; $, significance between interneurons and motoneurons alone.
The proportion of synapses apposed to gephyrin, GlyRα/β, or GABAARβ2/3 clusters and the proportion of nonsynaptic clusters were quantified on images collected with a CCD camera and are shown for motoneurons cultured alone for 3, 7, and 11 DIV (Fig. 7). Whether or not the gephyrin-, GlyRα/β-, and GABAARβ2/3-IR were adjacent to synapsin-IR, the proportion of apposition varied depending on the nature of the postsynaptic marker and stage of maturation (Fig.8). The same analysis was performed on motoneurons cultured with interneurons (data not shown). The mean number of synaptic boutons increased significantly at the surface of motoneurons cultured with or without interneurons during maturation, indicating a progressive establishment of synaptic connections (Table3). The mean numbers of gephyrin, GlyRα/β, or GABAARβ2/3 clusters per cell were not significantly different on motoneurons cultured alone, regardless of the stage in culture. In contrast, when motoneurons were cocultured with interneurons, the mean numbers of gephyrin, GlyRα/β, or GABAARβ2/3 clusters per cell increased significantly during the formation of synaptic contacts (between 3 and 7 DIV). On motoneurons cultured with interneurons, the number of gephyrin clusters was much smaller than the sum of GlyRα/β and GABAARβ2/3 clusters. This indicates that a large proportion of postsynaptic aggregates were composed of mosaics of receptors, as already seen by double-labeling experiments (Fig. 5G1–2, H1–2). The level of synaptic localization of the postsynaptic markers was further quantified on motoneurons cultured alone (Fig. 8A1–3) or in the presence of interneurons (Fig. 8B1–3). When motoneurons were cultured alone, the proportion of synapses showing gephyrin clusters was already high (77.7 ± 6.5%) at 3 DIV and varied little until 11 DIV (90.1 ± 3.1%), whereas the proportion of nonsynaptic gephyrin clusters per cell decreased progressively during maturation (Fig. 8A1). In contrast, only 9–14% of synapses formed between motoneurons were apposed to GlyRα/β clusters at any of the stages analyzed, and the proportion of nonsynaptic clusters decreased slightly between 7 and 11 DIV (Fig.8A2). The percentage of synapses with GABAARβ2/3 clusters increased slightly during maturation to 36.1 ± 3.9% at 11 DIV, whereas that of nonsynaptic GABAARβ2/3 clusters decreased in parallel (Fig.8A3). The scheme was different when motoneurons were cocultured with interneurons. The proportion of synapses showing gephyrin clusters increased significantly between 3 and 7 DIV and then remained stable until the end of the experiment, whereas the proportion of nonsynaptic gephyrin clusters decreased dramatically between 3 and 7 DIV (Fig. 8B1). The evolution of the level of synaptically and nonsynaptically localized GlyRα/β (Fig.8B2) and GABAARβ2/3 (Fig.8B3) clusters was a mirror image of what was found on motoneurons cultured alone.
Fig. 7.

Relationships during in vitromaturation of gephyrin, GlyR, and GABAAR with synaptic terminals on motoneurons cultured alone. Motoneurons double-stained for synapsin and gephyrin (A1–2, B1–2, C1–2), synapsin and GlyRα/β (D1–2, E1–2, F1–2), and synapsin and GABAARβ2/3 (G1–2, H1–2, I1–2) at 3, 7, and 11 DIV (left, middle, andright columns, respectively). A–C, At 3 DIV, Geph-IR clusters accumulate in front of most but not all synaptic boutons. Numerous Geph-IR clusters are also detected at nonsynaptic loci. At 7 and 11 DIV, the number of postsynaptic Geph-IR clusters increased, and the number of nonsynaptic Geph-IR clusters decreased.D–F, At all stages, most GlyRα/β clusters are detected at nonsynaptic sites. G–I, At 3 and 7 DIV, few GABAARβ2/3 clusters are in front of synapsin-IR terminals. Their number increases at 11 DIV. Arrows, Apposed synapsin and postsynaptic markers (Geph, GlyRα/β, or GABAARβ2/3); crossed arrows, presence of synapsin-IR but not of the above-mentioned postsynaptic markers;arrowheads, presence of Geph-, GlyRα/β-, or GABAARβ2/3-IR clusters without adjacent synapsin-IR.GABAARβ2/3, GABAARβ2/3 subunits-IR; Geph, gephyrin-IR;GlyRα/β, GlyRα/β subunits-IR;Syn, synapsin-IR. A1–A2, B1–B2, C1–C2, D1–D2, E1–E2, F1–F2, G1–G2, H1–H2, I1–I, Pairs of digitized images acquired with CCD camera (A1, B1, C1, D1, E1, F1, G1, H1, I1, FITC channel; A2, B2, C2, D2, E2, F2, G2, H2, I2, TRITC channel). Scale bar, 10 μm.
Table 3.
Quantifications of the mean numbers of synapses and of gephyrin, GlyR, and GABAAR clusters per cell duringin vitro maturation
| Number per cell | Motoneurons | Motoneurons + interneurons | ||||
|---|---|---|---|---|---|---|
| 3 DIV | 7 DIV | 11 DIV | 3 DIV | 7 DIV | 11 DIV | |
| Synapses | 5.3 ± 0.8 | 16.1 ± 2.0∉∉∉ | 24.9 ± 2.7¥ | 9.6 ± 0.9 | 50.5 ± 2.8∉∉∉ | 63.7 ± 2.4¥¥¥ |
| Gephyrin | 24.4 ± 3.8 | 28.9 ± 4.7 | 37.7 ± 4.6 | 10.8 ± 1.7 | 40.5 ± 3.1∉∉∉ | 67.4 ± 3.7¥¥¥ |
| GlyRα/β | 22.1 ± 3.1 | 24.7 ± 3.2 | 23.8 ± 3.3 | 4.2 ± 1.1 | 49.1 ± 4.0∉∉∉ | 54.4 ± 3.9 |
| GABAARβ2/3 | 14.5 ± 2.8 | 22.7 ± 3.3 | 16.7 ± 2.0 | 2.6 ± 0.8 | 45.2 ± 5.9∉∉∉ | 53.3 ± 4.3 |
Neurons were stained at indicated stages for synapsin, gephyrin, GlyRα/β, or GABAARβ2/3 subunits. Values are averages ± SEM of 10–18 cells. The levels of significance (ANOVA, Scheffé F test) are indicated by one (p < 0.05), two (p < 0.01), or three (p < 0.001) symbols. ∉, Significance of differences between 3 and 7 DIV; ¥, significance between 7 and 11 DIV.
These data suggest that the presynaptic glycinergic and/or GABAergic innervation supplied by interneurons (1) increases the number, size, and synaptic localization of GlyRα/β, GABAARβ2/3, and gephyrin clusters and (2) controls negatively the number of nonsynaptic clusters of receptors.
DISCUSSION
Role of presynaptic innervation in the formation of GlyR and GABAAR microdomains
The role of presynaptic innervation in the formation of postsynaptic clusters was investigated on motoneurons cultured alone, with DRG, or with spinal interneurons. On motoneurons cultured alone, GlyRα/β and GABAARβ2/3 formed nonsynaptic clusters. Therefore, clustering is independent of the corresponding presynaptic innervation, a situation comparable to that found in muscle (Kuromi and Kidokoro, 1984). Some motoneurons also expressed large patches of diffuse GlyRα/β and GABAARβ3. GlyR and GABAAR-IR were present on the somato-dendritic compartment, suggesting that targeting of receptors does not depend on innervation.
When motoneurons were contacted by GABAergic boutons, GABAARβ2/3 accumulated in front of these boutons, and the nonsynaptic GABAARβ3 disappeared. Nonsynaptic GlyRα/β could still be detected. In the presence of glycinergic and GABAergic innervations, both GlyRα/β and GABAARβ3 were detected in front of synaptic boutons where they could form mosaics, as in vivo at synapses containing (Triller et al., 1987; Todd et al., 1996; Colin et al., 1998) and releasing (Jonas et al., 1998) glycine and GABA. The amount of nonsynaptic GlyR decreases dramatically. Thus, the presence of GlyR and GABAAR as well as the disappearance of nonsynaptic receptor clusters are directly related to the presence of the corresponding presynaptic innervation. These results are consistent with what has been observed at the neuromuscular junction, where, after contact by the motor nerve, nAChR clusters become highly concentrated postsynaptically and disappear from nonsynaptic sites [see references in Hall and Sanes (1993)]. The influence of the afferent innervation has been investigated previously for central synapses. In micro-island hippocampal cultures, isolated GABAergic neurons do not form AMPA-type glutamate receptor clusters (Rao et al., 1997), and in isolated spinal cord neurons, the GluR1 subunit of AMPA receptors remains diffuse unless the cells receive glutamatergic innervation (O’Brien et al., 1997). Our data add further evidence indicating that presynaptic innervation is implicated in the diminution of nonsynaptic receptors.
In muscle, the size of nAChR clusters is regulated by the presynaptic nerve (Anderson and Cohen, 1977; Kuromi and Kidokoro, 1984). We show that a similar regulation also occurs at central synapses because the presence of presynaptic innervation increases the size and number of gephyrin, GlyRα/β, and GABAARβ3 clusters. This notion is also supported by a set of data obtained in vivo. The mean diameter and surface area of GlyR and gephyrin clusters increased along a somato-dendritic gradient on the goldfish Mauthner cell (Triller et al., 1990) and on cat spinal Ia interneurons and α- or γ-motoneurons (Alvarez et al., 1997). This was correlated with an increase in the size of the corresponding presynaptic releasing sites (Pierce and Mendell, 1993; Sur et al., 1995b). These observations and our present data are in favor of control by presynaptic innervation of the shape of inhibitory amino acid receptor clusters. This control could involve interactions of the receptors with the cytoskeleton (Kirsch and Betz, 1995).
Functional and pharmacological properties of the nonsynaptic GlyR of motoneurons cultured alone
We showed that the nonsynaptic GlyR of motoneurons cultured alone retain most functional and pharmacological properties of the postsynaptic receptors usually studied in cultures of all spinal neurons (Béchade et al., 1996). Their I–V curves are nonlinear, as described previously for spinal neurons (Bormann et al., 1987). Surprisingly, whereas nonlinearity has already been observed for various GlyR expressed in Xenopus oocytes (with asymmetrical Cl− concentrations) (Schmieden et al., 1989;Morales et al., 1994), linear I–V curves have been reported in symmetrical Cl− in transfected mammalian cells (Sontheimer et al., 1989; Bormann et al., 1993; Rundström et al., 1994; Lynch et al., 1995). The nonlinearity observed here in motoneurons favors inhibitory responses (Faber and Korn, 1987). We found similar EC50 for glycine (close to 40 μm) in motoneurons cultured alone and in spinal neurons. This result does not support the notion that nonsynaptic and synaptic GlyRs have different affinities for glycine related to their respective gephyrin-regulated density (Kuhse et al., 1995).
The strychnine sensitivity observed here both in motoneurons cultured alone and in spinal neurons (IC50 close to 50 nm) was higher than expected from the sensitivity reported for embryonic GlyRα2 coexpressed with gephyrin in human embryonic kidney 293 cells (IC50 of 1800 nm) (Takagi et al., 1992). This discrepancy could be explained by differences in experimental protocols, i.e., preincubation with strychnine (Boehm et al., 1997) and speed of the perfusion (slow applications of glycine leading to desensitization and underestimation of control responses). Embryonic spinal cord cultures were reported to express GlyR with low strychnine affinity (Becker et al., 1988; Hoch et al., 1989), and the α2* receptors were proposed to account for these results (Kuhse et al., 1990, 1991). Although α2* receptors might be present in our cultures, their low affinity for glycine (EC50 12 mm) excludes their contribution to the responses recorded here. α2 and/or α1 subunits could account for our results because they display a high affinity for glycine (Rundström et al., 1994) and strychnine (Grenningloh et al., 1990). The low sensitivity to picrotoxinin of the responses recorded in motoneurons cultured alone demonstrates that these cells express functional heteromeric receptors containing β subunits (known to lower the picrotoxinin sensitivity of GlyR) (Pribilla et al., 1992; Pistis et al., 1997). The presence of the latter subunit in the hetero-oligomer would allow GlyR to interact with gephyrin (Meyer et al., 1995) when they colocalize. Thus, the difference between the localizations of these two proteins in motoneurons cultured alone do not result from the absence of the β subunit. The nonsynaptic GlyR clusters detected on motoneurons cultured alone might result from an accumulation mediated by gephyrin, also detected at nonsynaptic loci (Fig. 8A1).
Gephyrin and postsynaptic accumulation of inhibitory amino acid receptors
Small clusters of gephyrin were detected in axons of motoneurons cultured alone. This axonal gephyrin, also observed in hippocampal cultured neurons (Craig et al., 1996), could anchor proteins other than GlyR and GABAAR. In spinal neurons, GlyRs are associated with the subsynaptic cytoskeleton via gephyrin (Kirsch et al., 1993b; Kirsch and Betz, 1995). It has recently been demonstrated that gephyrin is involved in the clustering of GABAAR (Essrich et al., 1998). Furthermore, it has been hypothesized that the interaction between gephyrin and GABAAR is most likely indirect (Essrich et al., 1998) because the GABAAR is stably attached to the subsynaptic microtubules by the GABAAR-associated protein (Wang et al., 1999). When motoneurons were cultured alone, gephyrin was detected at nonsynaptic sites but also at motoneuron-to-motoneuron postsynaptic sites, whereas GlyRα/β and GABAARβ2/3 subunits were mostly nonsynaptic. Therefore, on motoneurons cultured alone, postsynaptic gephyrin clusters likely to be associated with the neuronal nAChR are unable to accumulate GlyR and to a lesser extent GABAAR. In cocultures with DRG, postsynaptic gephyrin was associated with GABAAR facing GAD-IR boutons but not with GlyR. In cocultures with interneurons, postsynaptic gephyrin was found together with GlyRα/β and GABAARβ3 mosaics in front of glycine- and GABA-containing boutons. Strychnine treatment of spinal cultured neurons has shown that postsynaptic GlyR cluster formation depends on functional GlyR activity (Kirsch and Betz, 1998; Lévi et al., 1998). It has been suggested that during development GlyR could mediate a Ca2+ influx initiating gephyrin postsynaptic accumulation and trapping of randomly inserted membrane GlyR (Kirsch and Betz, 1998). A similar role of Ca2+ could theoretically account for the accumulation of gephyrin at GABAergic (Reichling et al., 1994) or cholinergic (McGehee and Role, 1995) synapses in motoneurons cultured alone. The Ca2+ hypothesis (Betz, 1998;Kirsch and Betz, 1998) alone is therefore not likely to account for the specific accumulation of given types of receptors at different synapses showing gephyrin clusters.
At least five variants of gephyrin are differentially expressed in the CNS (Prior et al., 1992; Kawasaki et al., 1997). Therefore, the IR detected on motoneurons could correspond to distinct gephyrin isoforms with specific interacting and/or aggregating properties for nAChR, GlyR, or GABAAR. The control of the expression and localization of gephyrin variants by the innervation may ensure the specificity of receptor accumulation (Betz, 1998). Variability of gephyrin also results from post-translational modifications such as phosphorylation (Prior et al., 1992). Independently of the postulated association of given forms of gephyrin with given receptors, the postsynaptic membrane has to be informed of the secretory phenotype of the presynaptic element to ensure the complementarity of the connected membranes. This could be achieved by interactions of gephyrin with transmembrane molecules other than neurotransmitter receptors. The scheme would then be comparable to that proposed for the recruitment of transmitter receptors or channels at β-neurexin–neuroligin junctions via interactions with PSD95–PDZ domains (Irie et al., 1997; Missler and Südhof, 1998).
Footnotes
This work was supported by grants from the Institut de la Recherche sur la Moelle Epinière, the Association Française contre les Myopathies, and European Community (Biomed II, CT 9 72 37 4). S.L. is a recipient of fellowships from the Institut Lilly and the Fondation pour la Recherche Médicale. We thank Drs. C. Henderson and V. Arce for help with the motoneuron purification and culture procedures and Professor Ascher for critical reading of this manuscript.
Correspondence should be addressed to Antoine Triller, Laboratoire de Biologie Cellulaire de la Synapse Normale et Pathologique, Institut National de la Santé et de la Recherche Médicale U-497, Ecole Normale Supérieure, 46 Rue d’Ulm, F-75005 Paris, France.
REFERENCES
- 1.Alvarez FJ, Dewey DE, Harrington DA, Fyffe REW. Cell-type specific organization of glycine receptor clusters in the mammalian spinal cord. J Comp Neurol. 1997;379:150–170. [PubMed] [Google Scholar]
- 2.Anderson MJ, Cohen MW. Nerve-induced and spontaneous redistribution of acetylcholine receptors on cultured muscle cells. J Physiol (Lond) 1977;268:757–773. doi: 10.1113/jphysiol.1977.sp011880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aoki C, Venkatesan C, Go CG, Mong JA, Dawson TM. Cellular and subcellular localization of NMDA-R1 subunit immunoreactivity in the visual cortex of adult and neonatal rats. J Neurosci. 1994;14:5202–5222. doi: 10.1523/JNEUROSCI.14-09-05202.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Arakawa Y, Sendtner M, Thoenen H. Survival effect of ciliary neurotrophic factor (CNTF) on chick embryonic motoneurons in culture: comparison with other neurotrophic factors and cytokines. J Neurosci. 1990;10:3507–3515. doi: 10.1523/JNEUROSCI.10-11-03507.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Baude A, Sequier JM, McKernan RM, Olivier KR, Somogyi P. Differential subcellular distribution of the α6 subunit versus the α1 and β2/3 subunits of the GABAA benzodiazepine receptor complex in granule cells of the cerebellar cortex. Neuroscience. 1992;51:739–748. doi: 10.1016/0306-4522(92)90513-2. [DOI] [PubMed] [Google Scholar]
- 6.Baude A, Molnar E, Latawiec D, McIlhinney RAJ, Somogyi P. Synaptic and nonsynaptic localization of the GluR1 subunit of the AMPA-type excitatory amino acid receptor in the rat cerebellum. J Neurosci. 1994;14:2830–2843. doi: 10.1523/JNEUROSCI.14-05-02830.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Baude A, Nusser Z, Molnar E, McIlhinney RA, Somogyi P. High-resolution immunogold localization of AMPA type glutamate receptor subunits at synaptic and nonsynaptic sites in rat hippocampus. Neuroscience. 1995;69:1031–1055. doi: 10.1016/0306-4522(95)00350-r. [DOI] [PubMed] [Google Scholar]
- 8.Béchade C, Colin I, Kirsch J, Betz H, Triller A. Expression of glycine receptor α subunits and gephyrin in cultured spinal neurons. Eur J Neurosci. 1996;8:429–435. doi: 10.1111/j.1460-9568.1996.tb01226.x. [DOI] [PubMed] [Google Scholar]
- 9.Becker C-M, Hoch W, Betz H. Glycine receptor heterogeneity in rat spinal cord during postnatal development. EMBO J. 1988;7:3717–3726. doi: 10.1002/j.1460-2075.1988.tb03255.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Betz H. Ligand-gated ion channels in the brain: the amino acid receptor superfamily. Neuron. 1990;5:383–392. doi: 10.1016/0896-6273(90)90077-s. [DOI] [PubMed] [Google Scholar]
- 11.Betz H. Gephyrin, a major player in GABAergic postsynaptic membrane assembly. Nat Neurosci. 1998;1:541–543. doi: 10.1038/2777. [DOI] [PubMed] [Google Scholar]
- 12.Binder LI, Frankfurter A, Kim H, Caceres A, Payne MR, Rebhun LI. Heterogeneity of microtubule-associated protein 2 during rat brain development. Proc Natl Acad Sci USA. 1984;81:5613–5617. doi: 10.1073/pnas.81.17.5613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Boehm S, Harvey RJ, von Holst A, Rohrer H, Betz H. Glycine receptors in cultured chick sympathetic neurons are excitatory and trigger neurotransmitter release. J Physiol (Lond) 1997;504.3:683–694. doi: 10.1111/j.1469-7793.1997.683bd.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bohlhalter S, Mohler H, Fritschy JM. Inhibitory neurotransmission in rat spinal cord: co-localization of glycine- and GABAA-receptors at GABAergic synaptic contacts demonstrated by triple immunofluorescence staining. Brain Res. 1994;642:59–69. doi: 10.1016/0006-8993(94)90905-9. [DOI] [PubMed] [Google Scholar]
- 15.Bormann J, Hamill OP, Sakmann B. Mechanism of anion permeation through channels gated by glycine and γ-aminobutyric acid in mouse cultured spinal neurones. J Physiol (Lond) 1987;385:243–286. doi: 10.1113/jphysiol.1987.sp016493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bormann J, Rundström N, Betz H, Langosch D. Residues within transmembrane segment M2 determine chloride conductance of glycine receptor homo- and hetero-oligomers. EMBO J. 1993;12:3729–3737. doi: 10.1002/j.1460-2075.1993.tb06050.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chandler CE, Parsons LM, Hosang M, Shooter EM. A monoclonal antibody modulates the interaction of nerve growth factor with PC12 cells. J Biol Chem. 1984;259:6882–6889. [PubMed] [Google Scholar]
- 18.Chauvet N, Drian MJ, Privat A. Immunocytochemical study of phenotypic plasticity of cultured dorsal root ganglion neurons during development. Int J Dev Neurosci. 1995;13:673–683. doi: 10.1016/0736-5748(95)00066-6. [DOI] [PubMed] [Google Scholar]
- 19.Chesnoy-Marchais D. Potentiation of chloride responses to glycine by three 5-HT3 antagonists in rat spinal neurones. Br J Pharmacol. 1996;118:2115–2125. doi: 10.1111/j.1476-5381.1996.tb15651.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Colin I, Augustin A, Rostaing P, Triller A. Localization of components of glycinergic synapses during rat spinal cord development. J Comp Neurol. 1998;398:359–372. [PubMed] [Google Scholar]
- 21.Craig AM, Blackstone CD, Huganir RL, Banker G. The distribution of glutamate receptors in cultured rat hippocampal neurons: postsynaptic clustering of AMPA-selective subunits. Neuron. 1993;10:1055–1068. doi: 10.1016/0896-6273(93)90054-u. [DOI] [PubMed] [Google Scholar]
- 22.Craig AM, Banker G, Chang W, McGrath ME, Serpinskaya AS. Clustering of gephyrin at GABAergic but not glutamatergic synapses in cultured rat hippocampal neurons. J Neurosci. 1996;16:3166–3177. doi: 10.1523/JNEUROSCI.16-10-03166.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.De Camilli P, Cameron R, Greengard P. Synapsin I (Protein I), a nerve terminal-specific phosphoprotein. I. Its general distribution in synapses of the central and peripheral nervous system demonstrated by immunofluorescence in frozen and plastic sections. J Cell Biol. 1983;96:1337–1354. doi: 10.1083/jcb.96.5.1337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dodd J, Morton SB, Karagogeos D, Yamamoto M, Jessell TM. Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron. 1988;1:105–116. doi: 10.1016/0896-6273(88)90194-8. [DOI] [PubMed] [Google Scholar]
- 25.Ericson J, Thor S, Edlund T, Jessell TM, Yamada T. Early stages of motor neuron differentiation revealed by expression of homeobox gene Islet-1. Science. 1992;256:1555–1560. doi: 10.1126/science.1350865. [DOI] [PubMed] [Google Scholar]
- 26.Essrich C, Lorez M, Benson JA, Fritschy JM, Lüscher B. Postsynaptic clustering of major GABAAR subtypes requires the γ2 subunit and gephyrin. Nat Neurosci. 1998;1:563–571. doi: 10.1038/2798. [DOI] [PubMed] [Google Scholar]
- 27.Faber DS, Korn H. Voltage-dependence of glycine-activated Cl− channels: a potentiometer for inhibition? J Neurosci. 1987;7:807–811. doi: 10.1523/JNEUROSCI.07-03-00807.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gao BX, Ziskind-Conhaim L. Development of glycine and GABA-gated currents in rat spinal motoneurons. J Neurophysiol. 1995;74:113–121. doi: 10.1152/jn.1995.74.1.113. [DOI] [PubMed] [Google Scholar]
- 29.Grenningloh G, Schmieden V, Schofield PR, Seeburg PH, Siddique T, Mohandas TK, Becker C-M, Betz H. Alpha subunit variants of the human glycine receptor: primary structure, functional expression and chromosomal localization of the corresponding genes. EMBO J. 1990;9:771–776. doi: 10.1002/j.1460-2075.1990.tb08172.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hall ZW, Sanes JR. Synaptic structure and development: the neuromuscular junction. Cell [Suppl] 1993;72:99–121. doi: 10.1016/s0092-8674(05)80031-5. [DOI] [PubMed] [Google Scholar]
- 31.Henderson CE, Camu W, Mettling C, Gouing A, Poulsen K, Karihaloo M, Rullamas J, Evans T, McMahon SB, Armanini MP, Berkemeier L, Philips HS, Rosenthal A. Neurotrophins promote motor neuron survival and are present in embryonic limb bud. Nature. 1993;363:266–270. doi: 10.1038/363266a0. [DOI] [PubMed] [Google Scholar]
- 32.Henderson CE, Phillips HS, Pollock RA, Davies AM, Lemeulle C, Armanini M, Simpson LC, Moffet B, Vandlen RA, Koliatsos VE, Rosenthal A. GDNF: a potent survival factor for motoneurons present in peripheral nerve and muscle. Science. 1994;266:1062–1064. doi: 10.1126/science.7973664. [DOI] [PubMed] [Google Scholar]
- 33.Henderson CE, Bloch-Gallego E, Camu W. Purified embryonic motoneurons. In: Cohen J, Wilkin G, editors. Nerve cell culture: a practical approach. Oxford UP; London: 1995. pp. 69–81. [Google Scholar]
- 34.Hoch W, Betz H, Becker C-M. Primary cultures of mouse spinal cord express the neonatal isoform of the inhibitory glycine receptor. Neuron. 1989;3:339–348. doi: 10.1016/0896-6273(89)90258-4. [DOI] [PubMed] [Google Scholar]
- 35.Irie M, Hata Y, Takeuchi M, Ichtchenko K, Toyoda A, Hirao K, Takai Y, Rosahl TW, Südhof TC. Binding of neuroligins to PSD-95. Science. 1997;277:1511–1515. doi: 10.1126/science.277.5331.1511. [DOI] [PubMed] [Google Scholar]
- 36.Jonas P, Bischofberger J, Sandkühler J. Corelease of two fast neurotransmitters at a central synapse. Science. 1998;281:419–424. doi: 10.1126/science.281.5375.419. [DOI] [PubMed] [Google Scholar]
- 37.Kawasaki BT, Hoffman KB, Yamamoto RS, Bahr BA. Variants of the receptor/channel clustering molecule gephyrin in brain: distinct distribution patterns, developmental profiles, and proteolytic cleavages by calpain. J Neurosci Res. 1997;49:381–388. doi: 10.1002/(sici)1097-4547(19970801)49:3<381::aid-jnr13>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 38.Kirsch J, Betz H. Widespread expression of gephyrin, a putative glycine receptor-tubulin linker protein, in rat brain. Brain Res. 1993;621:301–310. doi: 10.1016/0006-8993(93)90120-c. [DOI] [PubMed] [Google Scholar]
- 39.Kirsch J, Betz H. The postsynaptic localization of the glycine receptor-associated protein gephyrin is regulated by the cytoskeleton. J Neurosci. 1995;15:4148–4156. doi: 10.1523/JNEUROSCI.15-06-04148.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kirsch J, Betz H. Glycine-receptor activation is required for receptor clustering in spinal neurons. Nature. 1998;392:717–720. doi: 10.1038/33694. [DOI] [PubMed] [Google Scholar]
- 41.Kirsch J, Malosio ML, Wolters I, Betz H. Distribution of gephyrin transcripts in the adult and developing rat brain. Eur J Neurosci. 1993a;5:1109–1117. doi: 10.1111/j.1460-9568.1993.tb00965.x. [DOI] [PubMed] [Google Scholar]
- 42.Kirsch J, Wolters I, Triller A, Betz H. Gephyrin antisense oligonucleotides prevent glycine receptor clustering in spinal neurons. Nature. 1993b;366:745–748. doi: 10.1038/366745a0. [DOI] [PubMed] [Google Scholar]
- 43.Kuhse J, Schmieden V, Betz H. A single amino acid exchange alters the pharmacology of neonatal rat glycine receptor subunit. Neuron. 1990;5:867–873. doi: 10.1016/0896-6273(90)90346-h. [DOI] [PubMed] [Google Scholar]
- 44.Kuhse J, Kuryatov A, Maulet Y, Malosio ML, Schmieden V, Betz H. Alternative splicing generates two isoforms of the α2 subunit of the inhibitory glycine receptor. FEBS Lett. 1991;283:73–77. doi: 10.1016/0014-5793(91)80557-j. [DOI] [PubMed] [Google Scholar]
- 45.Kuhse J, Betz H, Kirsch J. The inhibitory glycine receptor: architecture, synaptic localization and molecular pathology of a postsynaptic ion-channel complex. Curr Opin Neurobiol. 1995;5:318–323. doi: 10.1016/0959-4388(95)80044-1. [DOI] [PubMed] [Google Scholar]
- 46.Kuromi H, Kidokoro Y. Nerve disperses preexisting acetylcholine receptor clusters prior to induction of receptor accumulation in Xenopus muscle cultures. Dev Biol. 1984;103:53–61. doi: 10.1016/0012-1606(84)90006-x. [DOI] [PubMed] [Google Scholar]
- 47.Lévi S, Vannier C, Triller A. Strychnine-sensitive stabilization of postsynaptic glycine receptor clusters. J Cell Sci. 1998;111:335–345. doi: 10.1242/jcs.111.3.335. [DOI] [PubMed] [Google Scholar]
- 48.Lewis CA, Faber DS. GABA responses and their partial occlusion by glycine in cultured rat medullary neurons. Neuroscience. 1993;52:83–96. doi: 10.1016/0306-4522(93)90184-h. [DOI] [PubMed] [Google Scholar]
- 49.Lynch JW, Rajendra S, Barry PH, Schofield PR. Mutations affecting the glycine receptor agonist transduction mechanism convert the competitive antagonist, picrotoxin, into an allosteric potentiator. J Biol Chem. 1995;270:13799–13806. doi: 10.1074/jbc.270.23.13799. [DOI] [PubMed] [Google Scholar]
- 50.McGehee DS, Role L. Physiological diversity of nicotinic acetylcholine receptors expressed by vertebrate neurons. Annu Rev Physiol. 1995;57:521–546. doi: 10.1146/annurev.ph.57.030195.002513. [DOI] [PubMed] [Google Scholar]
- 51.Meyer G, Kirsch J, Betz H, Langosh D. Identification of a gephyrin binding motif on the glycine receptor β subunit. Neuron. 1995;15:563–572. doi: 10.1016/0896-6273(95)90145-0. [DOI] [PubMed] [Google Scholar]
- 52.Missler M, Sudhof TC. Neurexins: three genes and 1001 products. Trends Genet. 1998;14:20–26. doi: 10.1016/S0168-9525(97)01324-3. [DOI] [PubMed] [Google Scholar]
- 53.Morales A, Nguyen QT, Miledi R. Electrophysiological properties of newborn and adult rat spinal cord glycine receptors expressed in Xenopus oocytes. Proc Natl Acad Sci USA. 1994;91:3097–3101. doi: 10.1073/pnas.91.8.3097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.O’Brien RJ, Mammen AL, Blackshaw S, Ehlers MD, Rothstein JD, Huganir RL. The development of excitatory synapses in cultured spinal neurons. J Neurosci. 1997;17:7339–7350. doi: 10.1523/JNEUROSCI.17-19-07339.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Persohn E, Malherbe P, Richards JG. In situ hybridization histochemistry reveals a diversity of GABAA receptor subunit mRNAs in neurons of the rat spinal cord and dorsal root ganglia. Neuroscience. 1991;42:497–507. doi: 10.1016/0306-4522(91)90392-2. [DOI] [PubMed] [Google Scholar]
- 56.Petralia RS, Wenthold RJ. Light and electron immunocytochemical localization of AMPA-selective glutamate receptors in the rat brain. J Comp Neurol. 1992;318:329–354. doi: 10.1002/cne.903180309. [DOI] [PubMed] [Google Scholar]
- 57.Petralia RS, Yokotani N, Wenthold RJ. Light and electron microscope distribution of the NMDA receptor subunit NMDAR1 in the rat nervous system using a selective anti-peptide antibody. J Neurosci. 1994;14:667–696. doi: 10.1523/JNEUROSCI.14-02-00667.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Pfeiffer F, Simler R, Grenningloh G, Betz H. Monoclonal antibodies and peptide mapping reveal structural similarities between the subunits of the glycine receptor of rat spinal cord. Proc Natl Acad Sci USA. 1984;81:7224–7227. doi: 10.1073/pnas.81.22.7224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pierce JP, Mendell LM. Quantitative ultrastructure of Ia boutons in the ventral horn: scaling and positional relationships. J Neurosci. 1993;13:4748–4763. doi: 10.1523/JNEUROSCI.13-11-04748.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pistis M, Belelli D, Peters JA, Lambert JJ. The interaction of general anaesthetics with recombinant GABAA and glycine receptors expressed in Xenopus laevis oocytes: a comparative study. Br J Pharmacol. 1997;122:1707–1719. doi: 10.1038/sj.bjp.0701563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pribilla I, Takagi T, Langosch D, Bormann J, Betz H. The atypical M2 segment of the β subunit confers picrotoxinin resistance to inhibitory glycine channels. EMBO J. 1992;11:4305–4311. doi: 10.1002/j.1460-2075.1992.tb05529.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Prior P, Schmitt B, Grenningloh G, Pribilla I, Multaup G, Beyreuther K, Maulet Y, Werner P, Langosch D, Kirsch J, Betz H. Primary structure and alternative splice variants of gephyrin, a putative glycine receptor-tubulin linker protein. Neuron. 1992;8:1161–1170. doi: 10.1016/0896-6273(92)90136-2. [DOI] [PubMed] [Google Scholar]
- 63.Ransom BR, Bullock PN, Nelson PG. Mouse spinal cord in cell culture. III. Neuronal chemosensitivity and its relationship to synaptic activity. J Neurophysiol. 1977;40:1163–1177. doi: 10.1152/jn.1977.40.5.1163. [DOI] [PubMed] [Google Scholar]
- 64.Rao A, Cha EM, Craig AM. Is appropriate presynaptic input necessary for the formation of postsynaptic receptor clusters? Soc Neurosci Abstr. 1997;23:656.8. [Google Scholar]
- 65.Reichling DB, Kyrozis A, Wang J, MacDermott AB. Mechanisms of GABA and glycine depolarization-induced calcium transients in rat dorsal horn neurons. J Physiol (Lond) 1994;476:411–421. doi: 10.1113/jphysiol.1994.sp020142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Richards JG, Schoch P, Häring P, Takacs B, Möhler H. Resolving GABAA/benzodiazepine receptors: cellular and subcellular localization in the CNS with monoclonal antibodies. J Neurosci. 1987;7:1866–1886. doi: 10.1523/JNEUROSCI.07-06-01866.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Roy G, Philippe E, Gaulin F, Guay G. Peripheral projections of the chick primary sensory neurons expressing γ-aminobutyric acid immunoreactivity. Neuroscience. 1991;45:177–183. doi: 10.1016/0306-4522(91)90113-3. [DOI] [PubMed] [Google Scholar]
- 68.Rundström N, Schmieden V, Betz H, Bormann J, Langosch D. Cyanotriphenylborate: subunit-specific blocker of glycine receptor chloride channels. Proc Natl Acad Sci USA. 1994;91:8950–8954. doi: 10.1073/pnas.91.19.8950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sanes JR. Genetic analysis of postsynaptic differentiation at the vertebrate neuromuscular junction. Curr Opin Neurobiol. 1997;7:93–100. doi: 10.1016/s0959-4388(97)80126-2. [DOI] [PubMed] [Google Scholar]
- 70.Schmieden V, Grenningloh G, Schofield PR, Betz H. Functional expression in Xenopus oocytes of the strychnine binding 48 kd subunit of the glycine receptor. EMBO J. 1989;8:695–690. doi: 10.1002/j.1460-2075.1989.tb03428.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Schoenen J, Delree P, Leprince P, Moonen G. Neurotransmitter phenotype plasticity in cultured dissociated adult rat dorsal root ganglia: an immunocytochemical study. J Neurosci Res. 1989;22:473–487. doi: 10.1002/jnr.490220414. [DOI] [PubMed] [Google Scholar]
- 72.Schröder S, Hoch W, Becker CM, Grenningloh G, Betz H. Mapping of antigenic epitopes on the α1 subunit of the inhibitory glycine receptor. Biochemistry. 1991;30:42–47. doi: 10.1021/bi00215a007. [DOI] [PubMed] [Google Scholar]
- 73.Sheng M, Wyszynski M. Ion channel targeting in neurons. BioEssays. 1997;19:847–853. doi: 10.1002/bies.950191004. [DOI] [PubMed] [Google Scholar]
- 74.Sontheimer H, Becker C-M, Pritchett DB, Schofield PR, Grenningloh G, Kettenmann H, Betz H, Seeburg PH. Functional chloride channels by mammalian cell expression of rat glycine receptor subunit. Neuron. 1989;2:1491–1497. doi: 10.1016/0896-6273(89)90195-5. [DOI] [PubMed] [Google Scholar]
- 75.Sur C, McKernan R, Triller A. Subcellular localization of the GABAA receptor γ2 subunit in the rat spinal cord. Eur J Neurosci. 1995a;7:1323–1332. doi: 10.1111/j.1460-9568.1995.tb01123.x. [DOI] [PubMed] [Google Scholar]
- 76.Sur C, Triller A, Korn H. Morphology of the release site of inhibitory synapses on the soma and dendrites of an identified neuron. J Comp Neurol. 1995b;351:247–260. doi: 10.1002/cne.903510205. [DOI] [PubMed] [Google Scholar]
- 77.Takagi T, Pribilla I, Kirsch J, Betz H. Coexpression of the receptor-associated protein gephyrin changes the ligand binding affinities of the α2 glycine receptors. FEBS Lett. 1992;303:178–180. doi: 10.1016/0014-5793(92)80513-g. [DOI] [PubMed] [Google Scholar]
- 78.Todd AJ, Watt C, Spike RC, Sieghart W. Colocalization of GABA, glycine, and their receptors at synapses in the rat spinal cord. J Neurosci. 1996;16:974–982. doi: 10.1523/JNEUROSCI.16-03-00974.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Triller A, Cluzeaud F, Pfeiffer F, Betz H, Korn H. Distribution of glycine receptors at central synapses: an immunoelectron microscopy study. J Cell Biol. 1985;101:683–688. doi: 10.1083/jcb.101.2.683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Triller A, Cluzeaud F, Korn H. Gamma-aminobutyric acid-containing terminals can be apposed to glycine receptors at central synapses. J Cell Biol. 1987;104:947–956. doi: 10.1083/jcb.104.4.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Triller A, Seitanidou T, Franksson O, Korn H. Size and shape of glycine receptor clusters in a central neuron exhibit a somato-dendritic gradient. New Biol. 1990;2:637–641. [PubMed] [Google Scholar]
- 82.Tsuchida T, Ensini M, Morton SB, Baldassare M, Edlund T, Jessell TM, Pfaff SL. Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes. Cell. 1994;79:957–970. doi: 10.1016/0092-8674(94)90027-2. [DOI] [PubMed] [Google Scholar]
- 83.Van den Pol AN, Gorcs T. Glycine and glycine receptor immunoreactivity in brain and spinal cord. J Neurosci. 1988;8:472–492. doi: 10.1523/JNEUROSCI.08-02-00472.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Vannier C, Triller A. Biology of the postsynaptic glycine receptor. Int Rev Cytol. 1997;176:201–244. doi: 10.1016/s0074-7696(08)61611-3. [DOI] [PubMed] [Google Scholar]
- 85.Wang H, Bedford FK, Brandon NJ, Moss SJ, Olsen RW. GABA(A)-receptor-associated protein links GABA(A) receptors and the cytoskeleton. Nature. 1999;397:69–72. doi: 10.1038/16264. [DOI] [PubMed] [Google Scholar]
- 86.Wenthold RJ, Yokotani N, Doi K, Wada K. Immunochemical characterization of the non-NMDA glutamate receptor using subunit-specific antibodies. Evidence for a hetero-oligomeric structure in rat brain. J Biol Chem. 1992;267:501–507. [PubMed] [Google Scholar]
- 87.Wisden W, Gundlach AL, Barnard EA, Seeburg PH, Hunt SP. Distribution of GABAA receptor subunit mRNAs in rat lumbar spinal cord. Mol Brain Res. 1991;10:179–183. doi: 10.1016/0169-328x(91)90109-b. [DOI] [PubMed] [Google Scholar]
- 88.Wong E, Schousboe A, Saito K, Wu JY, Roberts E. Immunochemical studies of brain glutamate decarboxylase and GABA-transaminase of six inbred strains of mice. Brain Res. 1974;68:133–142. doi: 10.1016/0006-8993(74)90538-1. [DOI] [PubMed] [Google Scholar]
- 89.Yan Q, Johnson EJ. An immunohistochemical study of the nerve growth factor receptor in developing rats. J Neurosci. 1988;8:3481–3498. doi: 10.1523/JNEUROSCI.08-09-03481.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]