

Abstract
Allothetic and idiothetic navigation strategies use very different cue constellations and computational processes. Allothetic navigation requires the use of the relationships between relatively stable external (visual, olfactory, auditory) cues, whereas idiothetic navigation requires the integration of cues generated by self-movement and/or efferent copy of movement commands. The flexibility with which animals can switch between these strategies and the neural structures that support these strategies are not well understood. By capitalizing on the proclivity of foraging rats to carry large food pellets back to a refuge for eating, the present study examined the contribution of the hippocampus to the use of allothetic versus idiothetic navigation strategies. Control rats and fimbria–fornix-ablated rats were trained to follow linear, polygonal, and octagonal scent trails that led to a piece of food. The ability of the rats to return to the refuge with the food via the shortest route using allothetic cues (visual cues and/or the odor trail available) or using ideothetic cues (the odor trail removed and the rats blindfolded or tested in infrared light) was examined. Control rats “closed the polygon” by returning directly home in all cue conditions. Fimbria–fornix rats successfully used allothetic cues (closed the polygon using visual cues or tracked back on the string) but were insensitive to the direction and distance of the refuge and were lost when restricted to idiothetic cues. The results support the hypothesis that the hippocampal formation is necessary for navigation requiring the integration of idiothetic cues.
Keywords: allothetic cue, dead reckoning, idiothetic cue, fimbria–fornix, hippocampus, hippocampal lesions, odor tracking, path integration, piloting, spatial learning, spatial navigation
Allothetic and idiothetic navigation strategies use very different cue constellations and computational processes (Darwin, 1873; Barlow, 1964; O’Keefe and Nadel, 1978;Mittelstaedt and Mittelstaedt, 1980; Landeau et al., 1984; Etienne, 1987; Gallistel, 1990; Maurer and Séguinot, 1995). Allothetic navigation requires the use of the relationships between relatively stable external (visual, olfactory, auditory) cues, whereas idiothetic navigation requires the integration of cues generated by self-movement. An animal can obtain self-movement information from vestibular receptors, muscle and joint receptors, and efference copy of commands that generate movement. An animal may also use flows in visual, auditory, and olfactory stimuli caused by its movements. Using an allothetic strategy an animal can use geometrical calculations to determine directions and distances to places in its environment, whereas using an idiothetic strategy it can integrate and double-integrate cues generated by its movements to make the same determinations. The ability to estimate direction and distance are colloquially called “sense of direction” and “sense of distance.”
Although there is considerable evidence that the hippocampus is involved in allothetic navigation (O’Keefe and Nadel, 1978; O’Keefe and Speakman, 1987), there is also evidence from behavioral (Whishaw et al., 1997; Whishaw and Maaswinkel, 1998; Maaswinkel and Whishaw, 1999), modeling (Samsonovich and McNaughton, 1997), and electrophysiological (O’Mare et al., 1994;; Sharp et al., 1995;Taube and Burton, 1995; Blair and Sharp, 1996; McNaughton et al., 1996; Wiener, 1996; Golob and Taube, 1997) studies that the hippocampal formation is involved in idiothetic navigation. The relative contribution of the hippocampus to the two forms of navigation is still uncertain, however. Ordinarily, it is difficult to be certain that an animal is using an allothetic versus an idiothetic strategy because animals are very flexible in their use of strategies and cues (Etienne et al., 1996; Dudchenko et al., 1997; Martin et al., 1997;Maaswinkel and Whishaw, 1999). The objective of the present study was to solve the problem of cue specification in order to examine the relative contribution of the hippocampus in the use of these strategies. The rats were trained in a new task in which they followed linear or polygon scented trails to obtain a large food pellet hidden on an open field. Because rats have a proclivity to carry the food back to the refuge, accuracy and the cues used to return to the home base were dependent variables (Whishaw and Tomie, 1997). To force an animal to use an idiothetic strategy to reach its refuge with the food, the rats were tested when blindfolded or under infrared light, a spectral wavelength in which they cannot see, and in some experiments the scent trail was additionally removed once an animal reached the food. To examine the relative contribution of the hippocampus, fimbria–fornix (FF) lesions, which disrupt information flow in the hippocampal formation (Bland, 1986), impair memory (Gaffan and Gaffan, 1991), and produce spatial deficits (Whishaw and Jarrard, 1995), were used.
MATERIALS AND METHODS
Animals
Twelve adult female Long–Evans rats (University of Lethbridge vivarium), weighing 250–300 gm, were housed in groups in wire mesh cages in a laboratory with room temperature maintained at 20–21°C and lighted on a 12 hr light/dark cycle (8 A.M. to 8 P.M.). Six rats received sham operations and six received fimbria–fornix lesions before testing.
Surgery. For surgery, the rats were anesthetized with sodium pentobarbital (40 mg/kg, i.p.) and atropine methyl nitrate (5 mg/kg, i.p.). To make fimbria–fornix lesions, 1.5 mA cathodal current was passed for 40 sec through 00 stainless steel insect pins, insulated with epoxylite except at the surface of their tips. Lesions were made at two sites in each hemisphere using coordinates in reference to bregma and the surface of the dura: 1.3 mm posterior, 1.5 mm lateral, and 3.6 mm ventral, and 1.5 mm posterior, 0.5 mm lateral, and 3.3 mm ventral (Whishaw and Jarrard, 1996). The control rats received anesthesia only.
Feeding
Feeding was restricted to maintain the rats at 90% of their expected body weights. Large (750 mg) rodent pellets (Bio-Serv, Frenchtown, NJ) were used for reward during behavioral testing. Rats reliably carry these pellets to a refuge for eating (Whishaw et al., 1995a,b). After testing each day, the rats were supplementally fed with LabDiet Laboratory Rodent Pellets in their home cage.
Apparatus
The open field consisted of a 204-cm-diameter circular wooden table, similar to a Barnes’s spatial testing apparatus (Barnes, 1979), that was painted white and elevated 64 cm above the floor (Whishaw and Maaswinkel, 1998). Eight 11.5-cm-diameter holes were cut in the table, spaced equidistant around its perimeter and centered 13.5 cm from the table’s edge (Fig.1A). A cage, similar to a rat’s home cage, could be inserted beneath a hole to serve as a refuge. The apparatus was located in a test room in which many cues, including windows covered by blinds, counters, a refrigerator, cupboards, a desk with computers, etc., were present. A camera was located above the center of the table so that the behavior of the animals could be videorecorded (Whishaw and Tomie, 1997).
Fig. 1.

A, The foraging table showing the location of the refuge hole (●) and a string in one of its configurations with a food pellet located at its end. B, An example of the string that the rats tracked and food pellets that they carried back to the refuge. C, An example of a rat tracking along the string to the food pellet.
Strings and scent
The rats were trained to follow a scented string to the food pellet. The string was ∼2 mm diameter, but pieces were of variable length and were placed on the table to form varying patterns on the table (Fig. 1A,B). The strings were scented with almond extract. The odor remained on the string for ∼20 min after it was wiped onto the string with a soaked piece of cotton gauze. Preliminary experiments indicated that rats trained to follow the scented string (Fig. 1C) did not follow an odor tract left by a piece of string that had just been removed.
Masks and blindfolds
Masks and blindfolds, used to control the rats use of visual cues, were constructed of felt and attached by a Velcro collar fixed around a rat’s neck [see Whishaw and Maaswinkel (1998), their Fig. 2]. They were fastened across a rat’s face by an elastic chin strap that was attached to the neck collar. The elastic strap was flexible so that a rat could grasp food pellets with its mouth and chew and swallow them. A mask allowed the rats to see, whereas a blindfold occluded vision. The effectiveness of the blindfolds was tested on rats trained to swim to a visible platform located in a swimming pool. Well trained rats wearing a mask swam directly to the platform from any starting point on the periphery of the pool, whereas rats wearing the blindfold swam around the edge of the pool or swam in a haphazard manner. The rats were adapted to masks and blindfolds by having them wear the apparel for at least 30 min/d for 5 d before testing. Before the formal tests, a mask or blindfold was placed on the animals for 30 min before the test.
Infrared testing
In addition to testing the rats while their vision was occluded by blindfolds, some of the experiments were performed under infrared light. The test room was light proof, and the room lights were turned off. Under infrared light, the animals’ movements were recorded with a Sony infrared camera. The experimenter used an infrared spotter to orient in the test room.
Training
All of the training was conducted with straight scented trails. A piece of string of variable length, scented with almond extract, was stretched from the edge of the refuge hole and led to the food pellet (Fig. 1). Initially, the string was quite short (<10 cm) and led in different directions, but as the animals became proficient in following the string, its length was increased to up to >150 cm. Training lasted ∼14 d, by which time the rats were proficient in following the string to retrieve food under both mask and blindfolded conditions.
Analysis
The behavior of the rats was filmed on all of the tests. From the ongoing trials and the video recordings the following behavioral measures were made.
Correct trial. A correct trial was a trial during which a rat found a food pellet and returned directly to the starting hole without stopping at any other potential exit hole.
Retrieval. A retrieval was defined as an exit from the home cage and a return with a food pellet.
Error. An incorrect trial was one during which a rat found a food pellet but stopped at one of the other potential exits before returning to the exit from which its excursion began. A rat was deemed to have stopped at an exit if its snout was brought to within ∼2 cm of a hole (errors were usually unambiguous because the rats stopped and inserted their heads into the holes). In some experiments, if the rat followed the string home, rather than taking a more direct route, that was classified as a error.
Travel distance. The outward and homeward distances were analyzed with a movement-tracking device to compute distance traveled.
Response times. Using a stop watch, an observer recorded separately the time taken to find a food pellet and the time taken to return to the home cage with the food.
Heading angles. Measures of heading angles were made after a rat retrieved a food pellet and began its homeward trip. Once the rat had moved one body length with the food, the angle that it was pointing, measured through the long axis of the animal’s body relative to the most direct line to the refuge, was measured.
Statistical analysis
Group comparisons were made using ANOVA’s and ttests (Winer, 1962).
Histology
At the completion of the experiments, the rats were deeply anesthetized and perfused with saline and saline–formalin, and the brains were removed and stored in a 30% sucrose–formalin solution. The brains were cut in 40 μm sections on a cryostat, and alternate sections were stained with cresyl violet and for acetylcholinesterase.
Procedure
Testing began once the rats were reliably following the string to retrieve the food. The following four tests were administered.
Return accuracy on a linear scent track. A scented string 125 cm long and an unscented string of the same length were both stretched in a straight line on the table. The location of the strings varied from trial to trial, and trials were given in a pseudo-random order, with one trial given each day. Each rat received three trials while wearing a mask and three trials while wearing a blindfold.
Return accuracy on a linear track with scent removed. The rats were tested with strings of length 20, 30, 50, 100, and 150 cm. Strings were placed on the table so that one end was at the target distance and the other led into the rats’ refuge and then to the experimenter. Once a rat reached the food pellet, the string was gently pulled from the table. All of the rats received three trials wearing masks and three trials wearing blindfolds at each of the distances. Order of distance presentation varied from trial to trial as did string direction.
Return accuracy from a polygon scent track. The rats received four tests with the scented track positioned on the table in a polygon pattern (curved or angular). The length, direction, and pattern of the string arrangement were different for each test, but in all cases the distance home from the food location was much shorter than the string distance. For three tests, the rats performed once with masks and once with blindfolds. For the fourth test, the rats wore no head gear, and one trial was given in normal light and one trial was given in infrared light.
Travel distance on an octagonal scent track. The string led from the refuge cage and then formed an octagon (75 cm diameter) in the center of the table. No food pellet was present. Each rat received one trial under room lights on 1 d and another trial under infrared light on a second day. Once a rat reached the octagon, the string connecting the refuge with the octagon was removed. If a rat left the octagon and entered the refuge the trial was complete. If a rat completed four complete rotations around the octagon, the trial was ended by removing the rat.
Test from a novel location, with and without vision. Each rat completed two trials following the scented string from a novel location. On one trial the rats wore a mask, and on the second trial they wore blindfolds.
RESULTS
Histological results
The dorsal fornix and the fimbria were completely sectioned in all of the rats that were given lesions (Fig.2, top left vsright). The lesions were selective and did not damage the septum, the septal portions of the hippocampus, or the hippocampal commissure. The tract made by the electrodes and the lesion did a small amount of damage to the supracollasal septohippocampal pathways just above the lesion and along the path made by the penetration. Previous work has shown that supracallosal damage does not produce additional impairments on spatial tasks (Sutherland and Rodriguez, 1989; Jeltsch et al., 1994). Stains for acetylcholinesterase (AChE) revealed extensive depletion of AChE throughout the hippocampus (Fig. 2,bottom left vs right), a marker that serves to confirm the completeness of the lesion. From previous work it is known that the lesion used in the present experiment reduces cholinergic markers by ∼70% in the dorsal hippocampus (Cassel et al., 1991;Jeltsch et al., 1994), in addition to damaging many other afferent and efferent pathways.
Fig. 2.
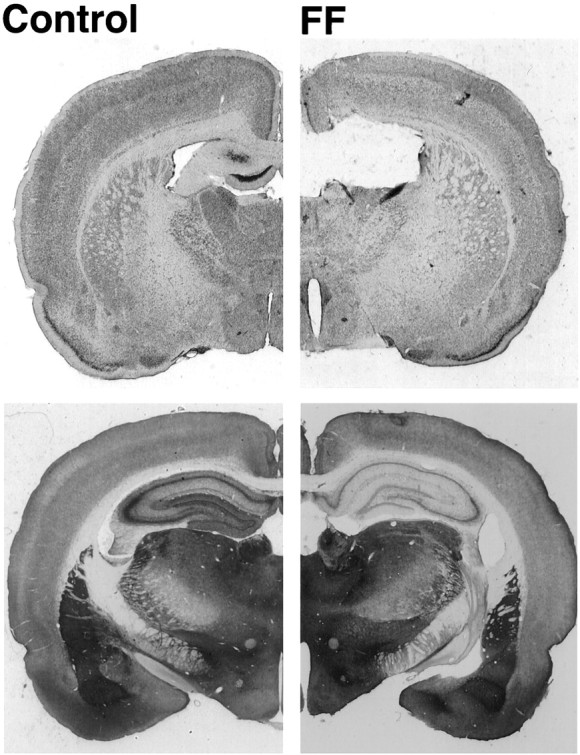
Photomicrographs of a control and lesion brain.Top, Cresyl violet sections shows the intact fimbria–fornix (FF) at the septal tip of the hippocampus (left) and an illustration of the absent fimbria–fornix (right). Bottom, Acetylcholinesterase sections through the hippocampus illustrating dense acetylcholinesterase staining in CA1 and dentate gyrus (left) and absence of staining in the FF lesion section (right). The slight cut in the left topof the cresyl violet section is an electrode tract.
General behavioral observations
After a rat in a refuge cage was placed beneath a hole, the rat typically poked its head out of the hole a number of times before it exited. It exited by pulling itself up with its forepaws and pushing with its hind paws. Once on the table, it paused and scanned the table in search of the scented string. On its outward journeys it walked along the string, which it sniffed with lateral scanning motions of the head. If a rat deviated away from the string, it quickly circled to relocate the string. Once a rat found a piece of food, it grasped it in its mouth and set off for the refuge. Once it arrived at the refuge, it inserted its head into the hole and adjusted the position of its feet so that it could drop down into the cage beneath the hole. In general, travel speed home was faster than travel speed out. If the rat was wearing a mask its travel speed was slightly slower than when it was sighted, both when tracking and returning to the refuge. Travel speed under infrared light was typically faster than travel speed under masked conditions. In all of the experiments, the major group differences were caused by the very poor performance of the fimbria–fornix rats when they could not use vision or olfaction. Although the fimbria–fornix rats were able to return home quickly in the light and to follow the string to the refuge in the absence of vision, they became lost and frequently ate their food on the table in the absence of these cues.
Return accuracy on a linear scent track
To evaluate tracking accuracy, a straight scented string and a straight nonscented string were placed on the table. On all trials, when masked and blindfolded, both the control and the fimbria–fornix rats followed the scented string to the food and ignored the nonscented string. They also returned directly back to the refuge, along the path of the scented string. Figure 3illustrates the return paths of the control and fimbria–fornix rats on one tracking problem. Although there were no group differences, there were significant differences in response times. The rats were faster in the mask than in the blindfolded condition (F(1,10) = 10.4; p < 0.01), and they were faster in returning to the home cage with the food than they were in traveling out to the food (F(1,10) = 2.7; p < 0.05).
Fig. 3.
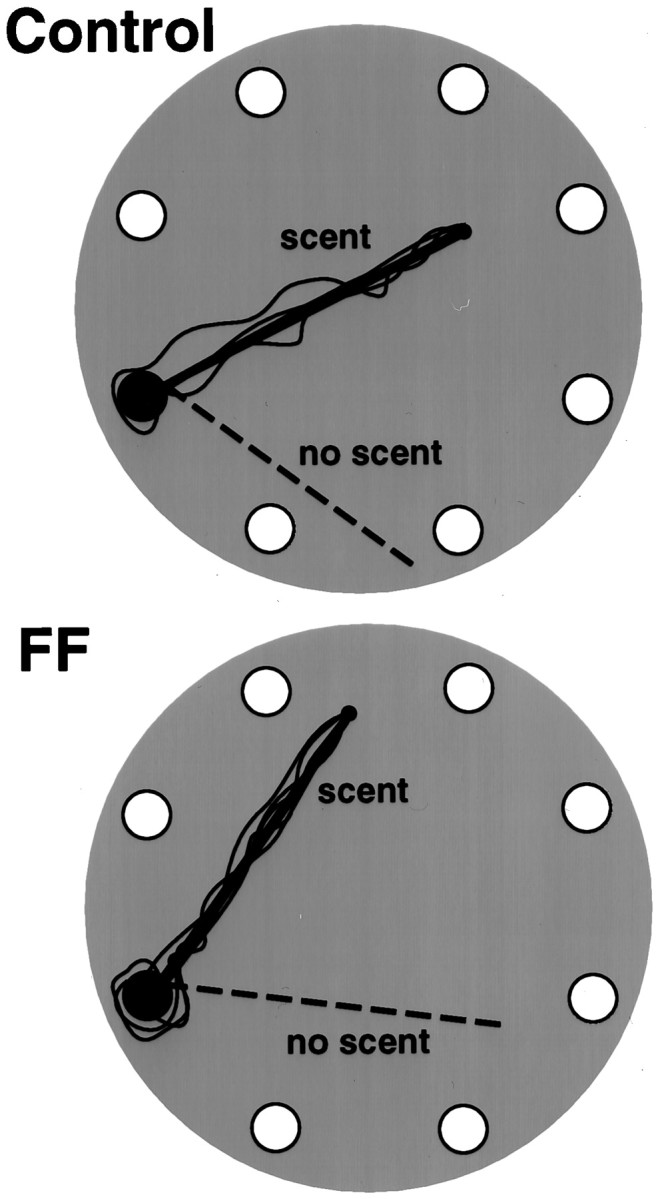
Illustrations of the homeward trajectories (solid lines) of the control (top) and fimbria–fornix (FF) rats along a scented string in the blindfold condition. The dotted line is a nonscented string.
Return accuracy on a linear track with scent removed
The rats were presented with strings of different length (Fig.4A), and the strings were removed just as the rats grasped the food, thus requiring that the rats return home without the scented track, either with vision or when blindfolded. The main findings were that the control rats were equally accurate in sighted and blindfolded conditions. When sighted, the fimbria–fornix rats performed almost as well as the control rats; when blindfolded, they made more errors, had longer return times, and had more deviant heading directions than they did when sighted and than did the control rats.
Fig. 4.
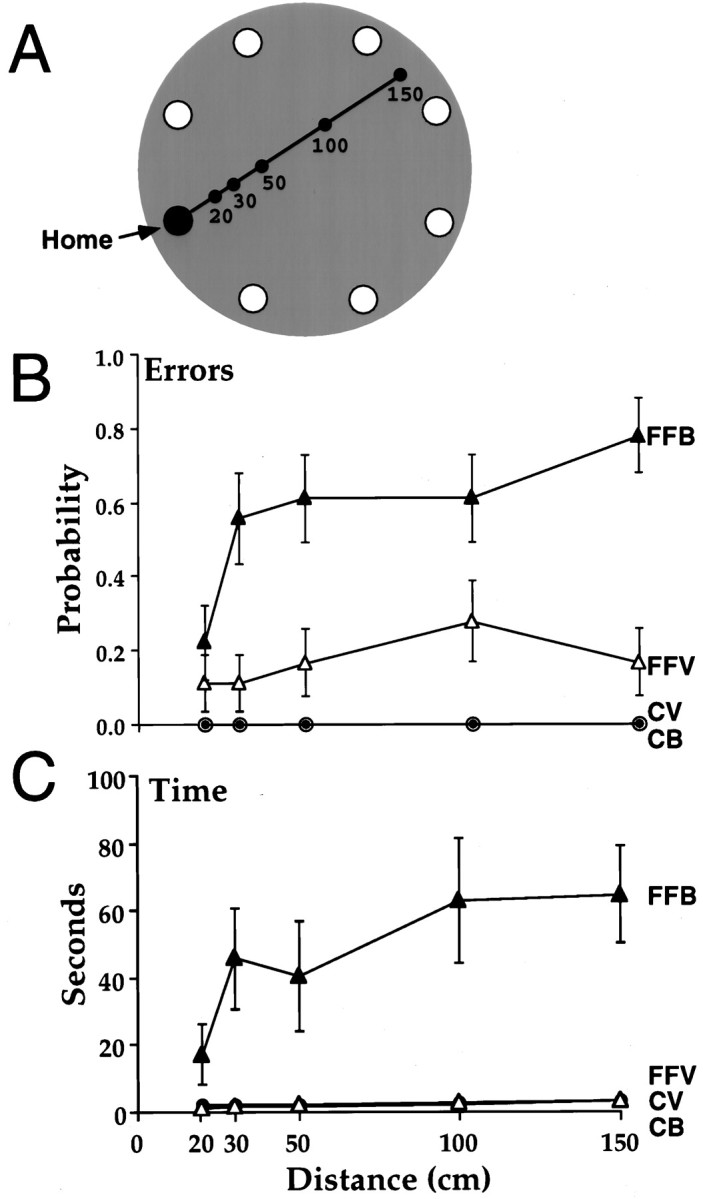
A, The rats traveled one of five linear distances to find a food pellet. The scent string was removed when they reached the food pellet. B, Errors as a function of distance on the return trip (mean and SE) consisted of a visit to an incorrect hole. C, Latencies to return home as a function of distance (mean and SE). FFB, Fimbria–fornix blindfold; FFV, fimbria–fornix vision;CV, control vision; CB, control blindfold.
Errors
A measure of errors, visits to incorrect holes on the return trip, gave significant effects of group (F(1,10) = 26.1; p = 0.005), visual condition (F(1,10) = 23.8; p = 0.006), and visual condition by group (F(1,10) = 23.8;p = 0.006). All of these effects were attributable to errors made by the fimbria–fornix rats (Fig. 4B). There were also significant effects of distance traveled out versus back (F(4,40) = 3.81; p = 0.01) and distance by group (F(4,40) = 3.8;p = 0.01). The interaction effect was attributable mainly to the much poorer performance of the fimbria–fornix group in the blindfolded condition than in the sighted condition. The fimbria–fornix rats did make errors in the light, but these errors occurred mainly when the strings were adjacent to other holes, rather than when they occupied the center of the board. Sighted fimbria–fornix rats had a strong tendency to stop at incorrect holes if they were adjacent to their route home.
Time
A measure of time taken to return to the refuge with the food gave a significant effect of group (F(1,10) = 15.171;p = 0.003), visual condition (F(1,10) = 15.5; p = 0.028), and visual condition by group (F(1,10) = 15.1;p = 0.003). All of the significant effects of time were attributable to the slow returns of the fimbria–fornix rats when they were blindfolded (Fig. 4C). On many trials they lost their way, ate their food, and only reached the refuge after haphazard walks around the foraging table. Although the fimbria–fornix rats returned more quickly at the shortest distance, the effect of distance did not quite reach significance (F(4,40) = 2.49; p = 0.058).
Heading angle
Heading angles, consisting of the direction the rat pointed after traveling one body length with the food relative to the straight line direction to the food, are summarized by group, condition, and distance in Figure 5. There were significant effects of group (F(1,10) = 14.9;p = 0.003), visual condition (F(1,10) = 13.8; p = 0.004), and visual condition by group (F(1,10) = 13.1; p = 0.05). These effects were caused by the more deviant angles of the rats with FF lesions. There were also significant effects of distance traveled to reach the food (F(4,40) = 4.44; p = 0.04) and distance traveled by group (F(4,40) = 2.94;p = 0.03). These effects were attributable to the more deviant heading angles of the blindfolded fimbria–fornix rats at longer distances.
Fig. 5.

Turning angles of rats after food retrieval as a function of distance traveled to reach the food. Note that the fimbria–fornix rats were inaccurate at all distances when blindfolded.
Behavioral analysis
When the way in which the rats retrieved the food and turned toward home was examined, differences were observed between the turning movements made by control and fimbria–fornix rats (Fig.6). The control rats typically stretched forward to retrieve the food and then recoiled along the main axis of their body so that they faced in the direction from which they had come. Forward stretching was much less obvious in the fimbria–fornix rats, which did not recoil and turn but simply pivoted as they walked. In blindfolded conditions, the first response of the fimbria–fornix rats with the food was often to begin to eat before attempting to return home. Counts on whether the rats recoiled along their outward path or moved in some other direction gave a significant group effect (F(1,10) = 39.2; p< 0.001).
Fig. 6.

Turning strategies of control and fimbria–fornix rats on retrieving a food pellet when the scent string was removed. The control rat stretches forward to retrieve the food and then recoils backward and pivots to face the refuge. The fimbria–fornix rat walks over to the food and fails to pivot back to the return path.
Return accuracy from polygon scent tracks
The rats were tested on three polygons under masked and blindfolded conditions (Fig.7A). Response times and distances out and back were averaged for the three problems. The main findings were that the control rats took the most direct route home under both masked and blindfolded conditions, whereas the fimbria–fornix rats took the direct route home only while they were sighted. An examination of the behavior of the rats showed that both control and fimbria–fornix rats tracked the string on the outward trip in both mask and blindfolded conditions. The control rats always returned directly to the starting hole via the shortest route under both visual and blindfolded conditions. The fimbria–fornix group returned directly home when sighted, but when blindfolded they followed the scent track home. Accordingly, the outward distance for both groups of rats in both conditions was very similar to the outward string length. The homeward distance for the control and sighted fimbria–fornix rats was very close to the shortest distance back to the starting hole. The travel distance of the blindfolded fimbria–fornix rats was much longer than the string distance back. This was because many of the rats walked off course and had to find their way back to the string, or else they circled, as if searching for the refuge, before they reached it.
Fig. 7.

A, Three polygon string tracks that the rats were required to follow. B, Time (mean and SEs on the three problems) taken to travel to the food (Out) and return (Back) in control and fimbria–fornix (FF) rats in mask and blindfold conditions. C, Distance traveled to reach the food by control and fimbria–fornix rats in masked and blindfolded conditions. The top portion of the gray bars(Out) represents the distance to the food along the string. The bottom portion of the gray bars (Back) represents the shortest distance from the food to the home refuge. Note that the performance of the groups is quite similar and approximates the string out and shortest back distance, with the exceptions of the return back performance of the fimbria–fornix rats when blindfolded.
Masks or blindfolds
Travel times
Although there was no group effect for time (F(1,20) = 3.39; p = 0.09, the effects of visual condition (F(1,10) = 40;p = 0.001) and direction traveled (F(1,20) = 5.0; p = 0.49) were significant. The fimbria–fornix rats were faster on their outward trips than were the control rats, especially in the mask condition, and there were no effects of blindfolding within the groups. What is perhaps more important, the interactions between group and visual condition (F(1,10) = 20.5; p = 0.001) and group by direction traveled (F(1,10)= 10.8; p = 0.008) were significant. The interactions were attributable to slower returns in both control and fimbria–fornix rats in the blindfold condition and the especially slow return times for the fimbria–fornix rats in the blindfolded condition (Fig.7B).
Travel distance
Analysis of the travel distance gave significant effects for all of the main effects, groups (F(1,20) = 13.8; p = 0.004), visual condition (F(1,10) = 24; p = 0.001), and direction (F(1,20) = 16.3; p < 0.002). Outward travel distances in both groups in both visual conditions were similar and approximated the string distance, but there were group differences in homeward distance, as indicated by the significant interactions between group by visual condition (F(1,10) = 14.92; p = 0.003), group by direction (F(1,10) = 7.52;p = 0.001), and vision by direction (F(1,10) = 5.9; p = 0.03). There was no effect of blindfolding on the homeward trips of the control group, whose return trip approximated the distance of the shortest route home, but the fimbria–fornix group took long routes when blindfolded (Fig. 7C).
Room or infrared light
The rats were given one polygon problem under room light and another under infrared light, and their return paths are illustrated in Figure 8. All six control rats returned directly to the home under light and dark conditions, whereas five of the fimbria–fornix rats returned directly home under light conditions. All of the fimbria–fornix rats returned home along the track in the dark (Fig. 8A). As occurred in the blindfold conditions, return latencies for the fimbria–fornix rats in the dark were significantly higher than latencies for the fimbria–fornix rats in the light or for the control group in either the light or dark (p < 0.01) (Fig. 8B).
Fig. 8.

A, Homeward trajectories for control and fimbria–fornix (FF) rats under room light and infrared light on one polygon string problem. Returns of fimbria–fornix rats were inaccurate in the dark, because they were largely following the scent string back to the refuge. Return latencies were mean and SEs. Note the significantly longer latencies of the fimbria–fornix rats under infrared light.
Travel distance on an octagonal track
The rats were allowed access to an octagonal tract via a string bridge. Once they were on the octagonal track, the connecting bridge to their home was removed (Fig.9A). The control rats each made a single trip around the track, and when they arrived back at the starting location without finding food, they went to the refuge hole and entered the refuge cage. The fimbria–fornix rats made many trips around the octagon (Fig. 9B), and most were removed at the end of their fourth circuitry. Thus, the control rats made fewer circles around the octagon (control mean = 1 vs FF mean = 3.83) and spent much less time tracking the string (F(1,10) = 15.3; p = 0.003) than did the fimbria–fornix rats (Fig. 9C).
Fig. 9.

A, A string led from the refuge hole to an octagonal circuit on which there was no food. Once the rat was on the octagon, the string connecting it to the home cage was removed. B, Travel paths made by control and fimbria–fornix rats. Note that the control rats averaged one circle on the octagon before returning to the home cage, whereas the fimbria–fornix rats made many turns around the octagon.C, Time (mean and SE) spent traveling on the octagon for control and fimbria–fornix rats.
Test from a novel location, with and without vision
The return routes and choices made by control and fimbria–fornix rats in tests in which their refuge cage was moved to a new location are shown in Figure 10. When masked, all of the rats, with the exception of two control rats, returned to the familiar home location (Fig. 10A). The two control rats returned directly to the new home location. When blindfolded, all of the control rats returned by fairly direct paths to the new location. The fimbria–fornix rats were inaccurate and encountered incorrect locations or returned to the new location by following the string (Fig. 10B).
Fig. 10.

Paths and destinations of control and fimbria–fornix rats when the home cage was moved to a new location (shaded refuge). Note that in the mask condition in which the rats could see, most control rats and all fimbria–fornix rats carried the food to the old home location. In the blindfold condition in which the rats could not see, all control rats carried food to the new home location via a direct route, whereas fimbria–fornix rats reached other nearby holes or followed the string back to the new home location.
DISCUSSION
The experiments examined the contribution of the hippocampus to allothetic versus idiothetic navigation by exploiting a novel task in which rats attempted to return home after following a scented string from their refuge to a food pellet located at the end of the string. Control rats navigated efficiently using both allothetic and idiothetic cues. The rats with fimbria–fornix lesions successfully navigated using allothetic cues but were impaired when forced to use idiothetic cues. These results are consistent with the hypothesis that the integrity of the hippocampal formation is essential for idiothetic navigation.This may also be the first demonstration in tracking in an animal other than the dog.
Task demands
The novel feature of this study was the use of a scented string that allowed the subjects to track an odor trail to a food pellet. The use of tracking facilitates an analysis of the rats’ homing behavior in a number of ways. First, the rats’ outward path and the cue are specified. Rats are very capable of using surface olfactory cues (Maaswinkel and Whishaw, 1999), and so the use of a scented string forces them to focus on a particular cue. Second, all of the rats could be presented with exactly the same outward journey. This removes the variability that occurs when they take haphazard and individual outward routes in search of randomly located food. The third, and perhaps the most important, feature of the procedure is that it was possible to determine when the rats switched cues and navigation strategies between the outward and homeward trip.
In the course of the study, the rats received a large number of tracking problems, which included linear tracks, polygon tracks, and an octagonal track. What was similar in all of the problems, however, was the comparison that was made between the rats’ ability to navigate home when they could use allothetic cues (vision or olfaction) and their ability to return home using idiothetic cues (when blindfolded or tested in infrared light).
Cue use
When they were sighted, both the control and the fimbria–fornix rats traveled relatively directly home after a circuitous outward trip. In doing so, they likely used stable visual room cues to return to their home base. When blindfolded and presented with a polygon track, the control rats still took a relatively direct route home, ignoring the scent track and thus “closing the polygon.” In doing so they were likely integrating the self-movement cues generated on the outward trip.
In contrast to the ability of the control rats to switch from an allothetic to an idiothetic navigation strategy, the fimbria–fornix rats’ impairment in idiothetic navigation was starkly revealed when they wore blindfolds or were tested in infrared light. Rather than taking a direct route home after following a polygon track, they relocated the scent track and retraced its route back to the starting location. Consequently, their homebound route was as circuitous as their outbound route. This was not simply a matter of conveniently “taking an easy route home.” When the scent track was removed, the fimbria–fornix rats became disoriented, ate their food on the table, and then walked around almost haphazardly until they regained the refuge. Thus, in a situation in which they were forced to use idiothetic strategy, they were unable to do so. This result fails to confirm the suggestion that hippocampal rats preferentially use idiothetic navigation strategies (Pearce et al., 1998).
The comparative behavior of the control and fimbria–fornix rats suggested that control rats may be prepared, and the fimbria–fornix rats may be unprepared, for making a direct homeward trip. The fimbria–fornix rats’ inability to reach home via a direct route was apparent at all but the shortest outward trips. Nevertheless, after a trip of 20 cm (one body length), the fimbria–fornix rats circled after finding the food pellet, so it was not possible to demonstrate that they used an idiothetic strategy after even the shortest trip. The control rats also typically stretched forward as they tracked and then recoiled after grasping the food pellet and so turned back along the axis of their body, thus reversing their trajectory. Their behavior seemed to suggest that they were always anchored to the home base (Dudchenko et al., 1997; Martin et al., 1997). The fimbria–fornix rats walked over the food “as if they were going to pick it up and eat it on the spot” and then turned in an almost haphazard direction once they grasped it, suggesting that they did not have a similar “home anchor.”
That both control and fimbria–fornix rats used visual cues was confirmed by a probe test in which the animals were started from a novel location. Most of the control rats and all of the fimbria–fornix rats returned to the familiar location, thus signifying that they had previously learned that location in relation to visual room cues. That the control rats could switch to an idiothetic strategy was confirmed in a second probe trial in which the rats started from a novel location when blindfolded. All of the control rats returned to the novel location, whereas the fimbria–fornix rats tracked back along the string. The only strategy that would have allowed the control rats to take a direct route home to a novel location is idiothetic.
Distance
The experiments also suggested that the fimbria–fornix rats did not know the homeward distance, confirming a result of a previous study that described impaired homing on a linear track of variable distance (Whishaw, 1993). When blindfolded, the fimbria–fornix rats frequently picked up the food and circled as if looking for the home base at that location. If they tracked home, they frequently stopped and circled on the way, as if again looking for the home. To confirm they they displayed an impairment in distance estimates, the rats were given an octagonal track to follow on which there was no food. It was hypothesized that once the rats had completed a tour around the octagon, they would realize that they had returned to the proximity of the refuge and so would leave the track to return to the refuge. Although the control rats did return to the refuge after a single turn on the track, the fimbria–fornix rats remained on the octagonal track and continued to circle until they were removed.
Allothetic versus idiothetic strategies
In a number of previous studies it has been demonstrated that rats with hippocampal damage can form place responses using visual cues (Whishaw and Jarrard, 1995, 1996; Whishaw et al., 1995b; Whishaw and Maaswinkel, 1998; L. D. Day, M. Weisend, R. J. Sutherland, T. Schallert, unpublished observation). This was confirmed in the present study. In fact, the ability of the fimbria–fornix rats to return to the home base using visual cues was essential to the demonstration that they are impaired in idiothetic navigation. Nevertheless, it was interesting that even when they were sighted, the return routes of the fimbria–fornix rats were recognizably less direct than those of the control rats. Possibly, the control rats use allothetic and idiothetic strategies concurrently (Whishaw and Mittleman, 1986; Dudchenko et al., 1997; Martin et al., 1997), whereas the fimbria–fornix rats are able to use only allothetic cues. The fimbria–fornix rats’ dependence on allothetic cues was manifest in another way. They had a strong tendency to stop at incorrect refuge holes when the holes were adjacent to a homeward route. It is well known that hippocampal rats are more strongly responsive to local cues than are control rats (O’Keefe and Nadel, 1978), so that is likely that in learning to enter their home refuge they developed strong tendencies to approach similar stimuli encountered elsewhere.
Anatomical localization
The hippocampal formation is a complex of nuclei and pathways (Amaral and Witter, 1995), and there is behavioral evidence for heterogeneity in its function (Grey and McNaughton, 1983;Jarrard, 1993; Sharp et al., 1995; Whishaw and Jarrard, 1995). Because fimbria–fornix lesions disconnect a number of hippocampal structures, it is not possible to specify the structures that are involved in idiothetic navigation. Cells that code head direction are found in subicular complex (Taube, 1990, 1995; Sharp, 1997), parietal and posterior cingulate cortex (Chen et al., 1994), thalamus (Mizumori and Williams, 1993; Taube, 1995), and striatum (Wiener, 1996), suggesting a role for these structures in idiothetic navigation. On logical grounds, others have concluded that the behavior of place cells in the hippocampus proper suggests that they may also play a role in idiothetic navigation (McNaughton et al., 1996). Future studies will have to separately assess the roles of different anatomical structures on dissociative tasks such as the one used in the present study.
Conclusion
Contemporary research suggests that the hippocampus plays some central role in spatial navigation [but see Squire (1992)], but there are divergent views concerning this role (O’Keefe and Nadel, 1978;Worden, 1992; Muller et al., 1996; Whishaw and Maaswinkel, 1998). The present finding supports the idea that the hippocampus plays a role in idiothetic navigation. This conclusion is consistent with previous suggestions that the hippocampus contains an innate spatial framework within which it can generate vectors between points (Whishaw et al., 1995a,b, 1997; McNaughton et al., 1996; Samsonovich and McNaughton, 1997). Although the present results support a role for the hippocampus in idiothetic navigation, they are not definitive in assigning this as an exclusive function. Hippocampal animals are also impaired in learning new allothetic spatial problems in which the demands of working memory are high (Shapiro and O’Connor, 1992; Angeli et al., 1993; Whishaw and Jarrard, 1995, 1996). This may mean that idiothetic information aids in the transition between one problem and the next (Whishaw et al., 1997), or it may indicated that the impairment in idiothetic navigation is but one aspect of a general hippocampal function in spatial behavior and memory (O’Keefe and Nadel, 1978). The resolution of this question must await more definitive studies.
Footnotes
This work was supported by the Medical Research Council of Canada.
Correspondence should be addressed to Ian Q. Whishaw, Department of Psychology, University of Lethbridge, Lethbridge, Alberta, T1K 3M4, Canada.
REFERENCES
- 1.Amaral DG, Witter MP. Hippocampal formation. In: Paxinos G, editor. The rat nervous system. Academic; San Diego: 1995. pp. 443–493. [Google Scholar]
- 2.Angeli SJ, Murray EA, Mishkin M. Hippocampectomized monkeys can remember one place but not two. Neuropsychologia. 1993;31:1021–1030. doi: 10.1016/0028-3932(93)90030-4. [DOI] [PubMed] [Google Scholar]
- 3.Barlow JS. Inertial navigation as a basis for animal navigation. J Theor Biol. 1964;6:76–117. doi: 10.1016/0022-5193(64)90067-0. [DOI] [PubMed] [Google Scholar]
- 4.Barnes CA. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 1979;93:74–104. doi: 10.1037/h0077579. [DOI] [PubMed] [Google Scholar]
- 5.Blair HT, Sharp PE. Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat. Behav Neurosci. 1996;110:643–660. doi: 10.1037//0735-7044.110.4.643. [DOI] [PubMed] [Google Scholar]
- 6.Bland BH. The physiology and pharmacology of hippocampal formation theta rhythms. Prog Neurobiol. 1986;26:1–54. doi: 10.1016/0301-0082(86)90019-5. [DOI] [PubMed] [Google Scholar]
- 7.Cassel JC, Kelche C, Peterson GM, Ballough GP, Goepp I, Will B. Graft induced behavioral recovery from subcallosal septo-hippocampal damage in rats depends on maturity stage of donor tissue. Neuroscience. 1991;45:571–586. doi: 10.1016/0306-4522(91)90272-p. [DOI] [PubMed] [Google Scholar]
- 8.Chen LL, Lin LH, Green EJ, Barnes CA, McNaughton BL. Head direction cells in the rat posterior cortex. I. Anatomical distribution and behavioral modulation. Exp Brain Res. 1994;101:8–23. doi: 10.1007/BF00243212. [DOI] [PubMed] [Google Scholar]
- 9.Darwin C. On the origin of certain instincts. Nature. 1873;7:417–418. [Google Scholar]
- 10.Dudchenko PA, Goodridge JP, Seiterle DA, Taube JS. Effects of repeated disorientation on the acquisition of spatial tasks in rats: dissociation between the appetitive radial arm maze and aversive water maze. J Exp Psychol. 1997;23:194–210. doi: 10.1037//0097-7403.23.2.194. [DOI] [PubMed] [Google Scholar]
- 11.Etienne AS. The control of short-distance homing in the golden hamster. In: Ellen P, Thinus-Blanc C, editors. Cognitive processes in spatial orientation in animal and man. Martinus Nijhoff; Dordrecht, The Netherlands: 1987. pp. 223–251. [Google Scholar]
- 12.Etienne A, Maurer R, Seguinot V. Path integration in mammals and its interaction with visual landmarks. J Exp Biol. 1996;199:201–209. doi: 10.1242/jeb.199.1.201. [DOI] [PubMed] [Google Scholar]
- 13. Gaffan D, Gaffan EA. 1991. Amnesia in man following transection of the fornix. 114:2611–2618 [DOI] [PubMed]
- 14.Gallistel CR. The organization of learning. MIT; Cambridge, MA: 1990. [Google Scholar]
- 15.Golob EJ, Taube JS. Head direction cells and episodic spatial information in rats without a hippocampus. Proc Natl Acad Sci USA. 1997;94:7645–7650. doi: 10.1073/pnas.94.14.7645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Grey JA, McNaughton N. Comparison between the behavioural effects of septal and hippocampal lesions: a review. Neurosci Biobehav Rev. 1983;7:119–188. doi: 10.1016/0149-7634(83)90014-3. [DOI] [PubMed] [Google Scholar]
- 17.Jarrard LE. On the role of the hippocampus in learning and memory in the rat. Behav Neural Biol. 1993;60:9–26. doi: 10.1016/0163-1047(93)90664-4. [DOI] [PubMed] [Google Scholar]
- 18.Jeltsch H, Cassel JC, Jackisch R, Neufang B, Green PL, Kelche C, Hertting G, Will B. Lesions of supracallosal or infracallosal hippocampal pathways in the rat: behavioural, neurochemical, and histochemical effects. Behav Neural Biol. 1994;62:121–133. doi: 10.1016/s0163-1047(05)80033-0. [DOI] [PubMed] [Google Scholar]
- 19.Landeau B, Spelke E, Gleitman H. Spatial knowledge in a young blind child. Cognition. 1984;16:225–260. doi: 10.1016/0010-0277(84)90029-5. [DOI] [PubMed] [Google Scholar]
- 20.Maaswinkel H, Whishaw IQ. Homing with locale, taxon, and dead reckoning strategies by foraging rats: sensory hierarchy in spatial navigation. Behav Brain Res. 1999;99:143–152. doi: 10.1016/s0166-4328(98)00100-4. [DOI] [PubMed] [Google Scholar]
- 21.Martin GM, Harley CW, Smith AR, Hoyles ES, Hynes CA. Opaque transportation with rotation blocks reliable goal location on a plus maze but does not prevent goal location in the Morris maze. J Exp Psychol. 1997;23:183–193. doi: 10.1037//0097-7403.23.2.183. [DOI] [PubMed] [Google Scholar]
- 22.Maurer R, Séguinot V. What is modeling for? A critical review of the models of path integration. J Theor Biol. 1995;175:457–475. [Google Scholar]
- 23.McNaughton BL, Barnes CA, Gerrard JL, Gothard K, Jung JJ, Knierim JJ, Kudrimoti H, Quin Y, Skaggs WE, Suster M, Weaver KL. Deciphering the hippocampal polyglot: the hippocampus as a path integration system. J Exp Biol. 1996;199:173–185. doi: 10.1242/jeb.199.1.173. [DOI] [PubMed] [Google Scholar]
- 24.Mittelstaedt M-L, Mittelstaedt M. Homing by path integration in a mammal. Naturwissenschafen. 1980;67:566–567. [Google Scholar]
- 25.Mizumori SJY, Williams JD. Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats. J Neurosci. 1993;13:4015–4028. doi: 10.1523/JNEUROSCI.13-09-04015.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Muller RU, Stead M, Pach J. The hippocampus as a cognitive graph. J Gen Physiol. 1996;107:663–694. doi: 10.1085/jgp.107.6.663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O’Keefe J, Nadel L. The hippocampus as a cognitive map. Clarendon; UK: 1978. [Google Scholar]
- 28.O’Keefe J, Speakman A. single unit activity in the rat hippocampus during a spatial memory task. Exp Brain Res. 1987;68:1–27. doi: 10.1007/BF00255230. [DOI] [PubMed] [Google Scholar]
- 29.O’Mare S, Rolls ET, Berthoz A, Desner RP. Neurons responding to whole-body motion in the primate hippocampus. J Neurosci. 1994;14:6511–6523. doi: 10.1523/JNEUROSCI.14-11-06511.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pearce JM, Roberts ADL, Good M. Hippocampal lesions disrupt navigation based on cognitive maps but not heading vectors. Nature. 1998;369:75–77. doi: 10.1038/23941. [DOI] [PubMed] [Google Scholar]
- 31.Samsonovich A, McNaughton BL. Path integration and cognitive mapping in a continuous attractor neural network model. J Neurosci. 1997;17:5900–5920. doi: 10.1523/JNEUROSCI.17-15-05900.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sharp PE. Subicular cells generate similar spatial firing patterns in two geometrically and visually distinctive environments: comparison with hippocampal place cells. Behav Brain Res. 1997;85:71–92. doi: 10.1016/s0166-4328(96)00165-9. [DOI] [PubMed] [Google Scholar]
- 33.Sharp PE, Blair HT, Etkin D, Tzanetos DBJ. Influences of vestibular and visual motion information on the spatial firing patterns of hippocampal place cells. Neuroscience. 1995;15:173–189. doi: 10.1523/JNEUROSCI.15-01-00173.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shapiro ML, O’Connor C. N-methyl-d-aspartate receptor antagonist MK-801 and spatial memory representation: working memory is impaired in an unfamiliar environment but not in a familiar environment. Behav Neurosci. 1992;106:604–612. doi: 10.1037//0735-7044.106.4.604. [DOI] [PubMed] [Google Scholar]
- 35.Squire L. Memory and the hippocampus: a synthesis from findings with rats, monkey, and humans. Psychol Rev. 1992;99:195–231. doi: 10.1037/0033-295x.99.2.195. [DOI] [PubMed] [Google Scholar]
- 36.Sutherland RJ, Rodriguez AJ. The role of the fornix/fimbria and some related subcortical structures in place learning and memory. Behav Brain Res. 1989;32:129–144. doi: 10.1016/s0166-4328(89)80059-2. [DOI] [PubMed] [Google Scholar]
- 37.Taube JS. Head direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. J Neurosci. 1990;172:49–84. doi: 10.1523/JNEUROSCI.10-02-00420.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Taube JS. Head direction cells recorded in the anterior thalamic nuclei of freely moving rats. J Neurosci. 1995;15:70–85. doi: 10.1523/JNEUROSCI.15-01-00070.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Taube JS, Burton HL. Head direction cell activity monitored in a novel environment and during a cue conflict situation. J Neurosci. 1995;15:1953–1971. doi: 10.1152/jn.1995.74.5.1953. [DOI] [PubMed] [Google Scholar]
- 40.Whishaw IQ. Activation, travel distance, and environmental change influence food carrying in rats with hippocampal, medial thalamic and septal lesions: implications for studies on hoarding and theories of hippocampal function. Hippocampus. 1993;3:373–385. doi: 10.1002/hipo.450030311. [DOI] [PubMed] [Google Scholar]
- 41.Whishaw IQ, Jarrard L. Similarities vs. differences in place learning and circadian activity in rats after fimbria-fornix section or ibotenate removal of hippocampal cells. Hippocampus. 1995;5:595–604. doi: 10.1002/hipo.450050610. [DOI] [PubMed] [Google Scholar]
- 42.Whishaw IQ, Jarrard LE. Evidence for extrahippocampal involvement in place learning and hippocampal involvement in path integration. Hippocampus. 1996;6:513–524. doi: 10.1002/(SICI)1098-1063(1996)6:5<513::AID-HIPO4>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 43.Whishaw IQ, Maaswinkel H. Rats with fimbria–fornix lesions are impaired in path integration: a role for the hippocampus in “sense of direction.”. J Neurosci. 1998;18:3050–3058. doi: 10.1523/JNEUROSCI.18-08-03050.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Whishaw IQ, Mittleman G. Visits to starts, routes, places by rats (Rattus norvegicus) in swimming pool navigation tasks. J Comp Psychol. 1986;100:422–431. [PubMed] [Google Scholar]
- 45.Whishaw IQ, Tomie J. Piloting and dead reckoning dissociated by fimbria-fornix lesions in a rat food carrying task. Behav Brain Res. 1997;89:87–97. doi: 10.1016/s0166-4328(97)00068-5. [DOI] [PubMed] [Google Scholar]
- 46.Whishaw IQ, Coles BKL, Bellerive CHM. Food carrying: a new method for naturalistic studies of spontaneous and forced alternation. J Neurosci Methods. 1995a;61:139–143. doi: 10.1016/0165-0270(95)00035-s. [DOI] [PubMed] [Google Scholar]
- 47.Whishaw IQ, Cassel J-C, Jarrard LE. Rats with fimbria–fornix lesions display a place response in a swimming pool: a dissociation between getting there and knowing where. J Neurosci. 1995b;15:5779–5788. doi: 10.1523/JNEUROSCI.15-08-05779.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Whishaw IQ, McKenna J, Maaswinkel H. Hippocampal lesions and path integration. Curr Opin Neurobiol. 1997;7:228–234. doi: 10.1016/s0959-4388(97)80011-6. [DOI] [PubMed] [Google Scholar]
- 49.Wiener SI. Spatial, behavioral and sensory correlates of hippocampal CA1 complex spike cell activity: implications for information processing functions. Prog Neurobiol. 1996;49:335–361. doi: 10.1016/0301-0082(96)00019-6. [DOI] [PubMed] [Google Scholar]
- 50.Winer BJ. Statistical principles in experimental design. McGraw-Hill; London: 1962. [Google Scholar]
- 51.Worden R. Navigation by fragment fitting: a theory of hippocampal function. Hippocampus. 1992;2:165–188. doi: 10.1002/hipo.450020208. [DOI] [PubMed] [Google Scholar]