

Abstract
The perforant path input (pp) is a major direct source of specific sensory information for the CA1 hippocampal region. The termination area of this pathway, the stratum lacunosum-moleculare, has the highest concentration of dopamine receptors in the hippocampus. We have examined the properties of the pp input and its modulation by dopamine. The input is glutamatergic and has a larger NMDA component than the Schaffer collateral (sc) input. Dopamine strongly inhibits the response to pp stimulation (IC50 ∼3 μm) but not the response to sc stimulation. Dopamine reduces both the NMDA and AMPA components of transmission at the pp and increases paired-pulse facilitation. In the sc, the NMDA component but not the AMPA component is decreased, and paired-pulse facilitation is not affected. The effect of dopamine on the pp does not depend on GABAA inhibition but is reduced by the antagonists of both D1 and D2 families of dopamine receptors. The effect is not completely blocked by the combination of D1 and D2 antagonists, but is completely blocked by the atypical neuroleptic clozapine. Our results provide the first evidence for strong dopaminergic control of transmission in the perforant path. By inhibiting this pathway, dopamine hyperfunction and/or NMDA hypofunction abnormalities implicated in schizophrenia may isolate CA1 from its main source of sensory information.
Keywords: AMPA, CA1, clozapine, dopamine, D1, D2, eticlopride, GABAA, haloperidol, hippocampus, NMDA, perforant path, SCH 23390, Schaffer collaterals, schizophrenia, U-101958
The hippocampus has an important role in memory (Buzsaki, 1989; Jensen et al., 1996; Eichenbaum, 1997), in habituation, in the detection of novelty (Vinogradova, 1984; Levy, 1989), and in the spatial mapping of the environment (Skaggs and McNaughton, 1992; O’Keefe, 1993). The hippocampal CA1 region receives dopaminergic input from midbrain sources and has all five types of dopamine receptors, a dopamine uptake system, DARPP, and other machinery of dopaminergic target cells (for review, see Otmakhova and Lisman, 1996). An increase in the hippocampal dopaminergic function improves learning in animals (Grecksch and Matthies, 1982; Packard and White, 1991; Gasbarri et al., 1996; Bernabeu et al., 1997). There has been substantial recent progress in understanding how dopamine affects hippocampal synaptic plasticity. Studies on the Schaffer collateral input to the CA1 region show that dopamine enhances long-term potentiation (Frey et al., 1993; Otmakhova and Lisman, 1996) and inhibits depotentiation (Otmakhova and Lisman, 1998).
Although dopamine receptors are relatively widespread in the hippocampus, they are most concentrated in the distal dendritic region of the CA1 field, the stratum lacunosum-moleculare (Swanson et al., 1987; Goldsmith and Joyce, 1994). This suggests that it would be of interest to examine the dopaminergic modulation of the synaptic inputs into this stratum. Anatomical and physiological work indicates that the principal input into this stratum is a direct projection from the entorhinal cortex (Lopes da Silva et al., 1990), but there are also inputs from the nucleus reuniens of the thalamus (Dolleman-Van der Weel et al., 1997). After previous convention, we term these inputs the perforant path input (pp). Although the pp has not received as much attention as other pathways, it appears to have an important role.In vivo recordings show that cells in the entorhinal cortex, the source of the perforant path to CA1, generate responses specific to particular stimuli and modalities (Vinogradova, 1984). Such responses are also seen in CA1 but cannot be brought there via the indirect (dentate gyrus, CA3) pathway because sensory specificity is rarely observed in these intermediary structures. Furthermore, destruction of the dentate gyrus actually increases the fraction of CA1 and CA3 neurons with specific sensory responses (Vinogradova, 1984) and does not strongly affect their place fields (McNaughton et al., 1989). The direct pp input therefore appears to be the main source of specific sensory information for CA1 and CA3 fields (Vinogradova, 1984;McNaughton et al., 1989).
Elucidation of the role of dopamine in the hippocampus is relevant to schizophrenia and other dopamine-dependent brain disorders. The hippocampus has been implicated in schizophrenia because the disease is associated with abnormalities in hippocampal structure (Bogerts and Falkai, 1995; Fukuzako et al., 1995) and corticohippocampal interactions (Shenton et al., 1992; Heckers et al., 1998). Dopamine hyperfunction has been previously implicated in schizophrenia (Gray et al., 1995; Joyce and Meador-Woodruff, 1997), but there has been little information about how dopamine affects synaptic transmission in the hippocampus. Previous works indicate that dopamine does not affect the sc synaptic transmission (Gribkoff and Ashe, 1984; Marciani et al., 1984; Pockett, 1985) (but see Hsu, 1996). Here we show that dopamine strongly suppresses the pp input to the CA1 hippocampal region.
MATERIALS AND METHODS
Transverse slices (400-μm-thick) from the dorsal hippocampus of 28-to 45-d-old male Long–Evans rats were used in this study. Part of the dentate gyrus and the CA3 field were cut from the slices as shown on Figure 1A. Slices were superfused with artificial CSF (ACSF) at a flow rate of 1.5–2.5 ml/min. ACSF contained (in mm): NaCl 120, NaHCO4 26, NaH2PO4 1, KCl 2.5, CaCl2 2.5, MgSO4 1.3, and d-glucose 10 and was oxygenated (95% O2 and 5% CO2). Experiments were done at the temperature 29.2–30.2°C. All electrodes (glass pipettes filled with ACSF,r = 0.2–0.3 MΩ) were placed in the CA1 hippocampal region, closer to the subiculum than to CA3 (Fig.1A). Two electrodes were placed in the distal of the stratum radiatum 120–160 μm apart from each other for stimulating and recording from the sc synapses. Another pair of similar electrodes was positioned in the stratum lacunosum-moleculare to stimulate and record from the pp synapses. The thickness of the stratum lacunosum-moleculare is not even over the stretch of dorsal hippocampus. Procedurally, in each slice we subdivided this stratum by eye into two equal parts (sublayers, bands): proximal, close to st. radiatum, and distal, close to fissure, and placed our electrodes on the distal band. According to the literature, the region of CA1 adjoining the subiculum is a site of lateral pp projections (Swanson et al., 1987; Lopes da Silva et al., 1990). The distance between the pp electrodes was ∼100 μm. Data acquisition and initial “on-line” analysis were done using a PC through an LM-900 interface (Dagan Corporation, Minneapolis, MN) using a custom-made AXOBASIC program. We alternated the stimulation/recording between pp and sc inputs; each input was stimulated every 30 sec.
Fig. 1.
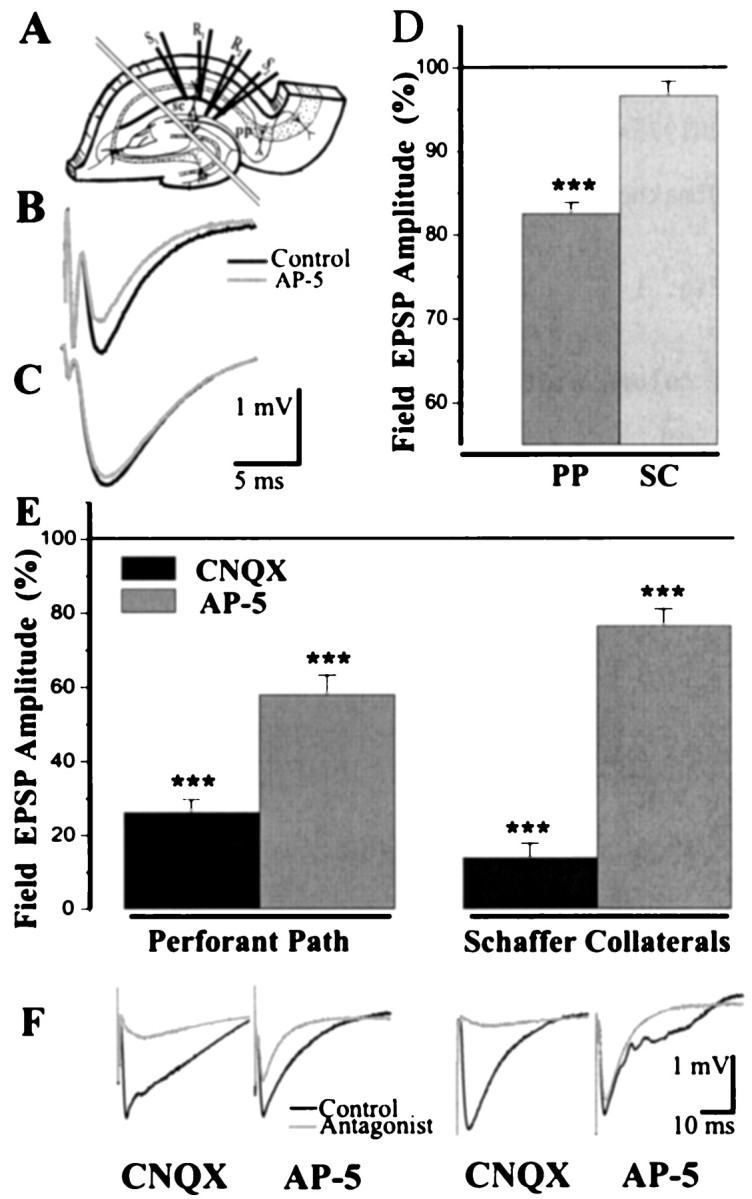
Differences in the pp and sc fEPSP.A, Electrode positions for the simultaneous pp and sc fEPSP recording. Parallel lines signify the cut made to isolate the inputs. B, An example of the pp fEPSP in regular ACSF and the effect of NMDA receptors blockade.C, The sc fEPSP in regular ACSF shows no substantial effect of NMDA antagonist. D, Averaged data on the effect of NMDA antagonist ± AP-5 (100 μm) on the pp and sc fEPSP amplitude in regular ACSF. E, Averaged data on the effect of NMDA and AMPA antagonists on the pp and sc fEPSP amplitude in low Mg2+, picrotoxin, and tetrodotoxin containing ACSF. Horizontal lines (100%) represent the fEPSP amplitude before drug application. Data in columns were taken at 10 min after the start of application. Significance in paired t test: ***p < 0.001. F, Field EPSP traces from individual experiments with NMDA and AMPA antagonist applications in low Mg2+, picrotoxin, and tetrodotoxin containing ACSF. Pathway labels below Ealso refer to F.
All drugs were purchased at Research Biochemicals (Natick, MA). Water soluble drugs were dissolved in the ACSF or water with 0.02% ascorbic acid for stock solutions and then diluted in ACSF and oxygenated before the experiment. None of the drugs was oxidation-protected during the application to avoid the necessity of additional controls for antioxidant action. Water insoluble drugs (neuroleptics and a specific D4 antagonist) were initially dissolved in DMSO for stocks and then sonicated in ACSF immediately before each experiment. The final concentration of DMSO during perfusion did not exceed 0.05–0.1%. Water soluble antagonists were applied starting 10 min before dopamine and throughout dopamine application. Water insoluble antagonists were applied at a higher concentration starting 25 min before dopamine.
For statistical analysis, responses were collected and averaged in 1 or 5 min periods. Maximal initial field EPSP (fEPSP) slope (millivolts per millisecond), amplitude (millivolts), and fiber volley amplitude were calculated. In most cases, data for each experiment were normalized relative to baseline. One minute statistics (Mean ± SEM) were used for the time plots (see Figs. 3-5), whereas 5 min interval data were used for all statistical comparisons. As a standard requirement, an a priori α value of 0.05 was established before all experiments. The effect of drug was estimated in each slice relative to baseline and then analyzed for the whole experimental series using two-tailed pairedt test for means (Microsoft Excel statistical package). For between-slice comparisons, a Student’s two-tailed t test was used. Concentration–response curve fitting (on a logarithmic concentration scale) was done in Microcal Origin statistical package. Dopamine antagonist potency was estimated by comparing the dopamine effect in the presence or absence of the antagonist on the same slices by two factors ANOVA for repeated measurements (Microsoft Excel). Considered factors were drug (presence or absence of the antagonist; df = 1), time (from the start of dopamine application including 5 min of washout; in 5 min bins, df = 3), and drug * time interaction (df = 3). If ANOVA showed a significant drug effect, apost hoc paired t test (two-tailed) was administered to check the significance of the effect in each 5 min interval. Figures show means and SEs.
Fig. 3.
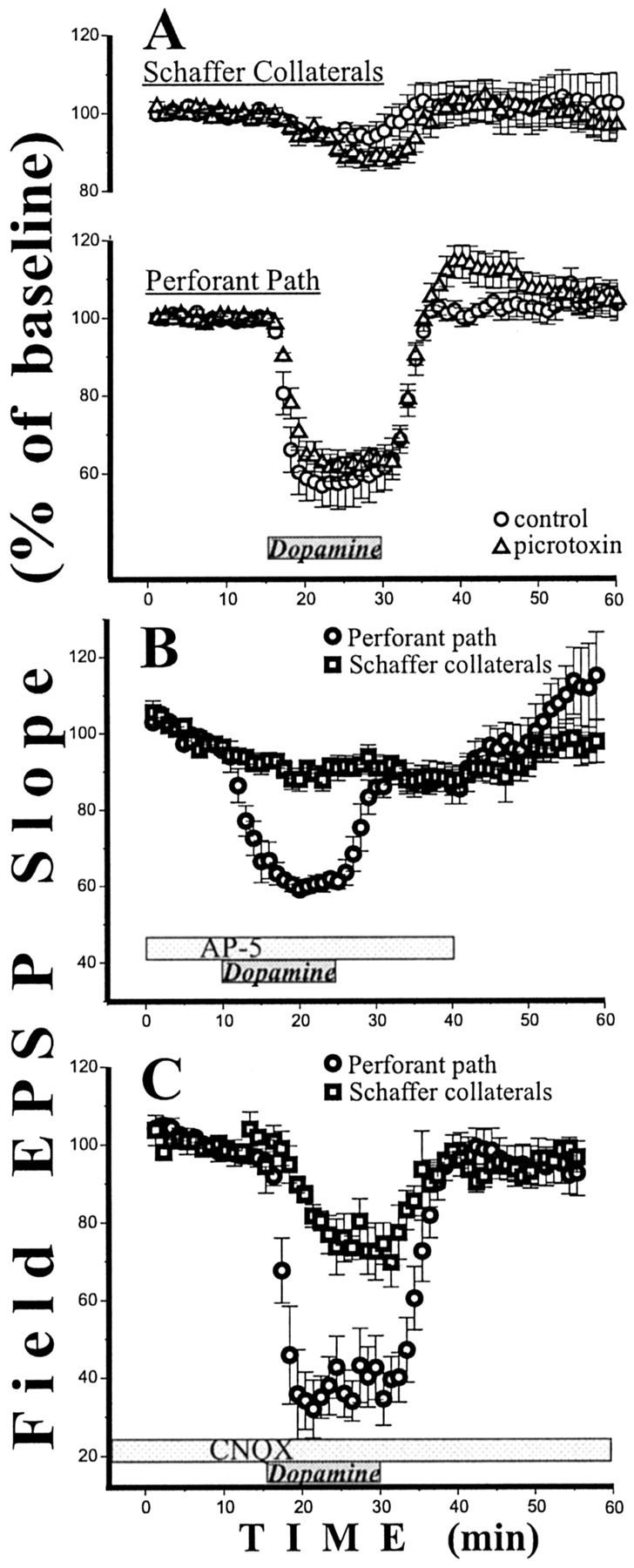
Dopamine-induced suppression of pp fEPSP slope does not depend on GABAA inhibition and involves both NMDA and AMPA components of the response. A, Blockade of GABAA inhibition by picrotoxin (n = 6) did not affect the magnitude of dopamine-induced suppression but increased the pp fEPSP slope during washout. B, Dopamine strongly suppressed the isolated AMPA fEPSP slope in the pp but not in the sc. C, Isolated NMDA component of the fEPSP in both the pp and the sc show strong inhibition by dopamine. Time of drug applications is marked by rectangles. Experiments inB and C were done in 0.1 Mg2+ in the presence of picrotoxin; tetrodotoxin was not used.
Fig. 4.
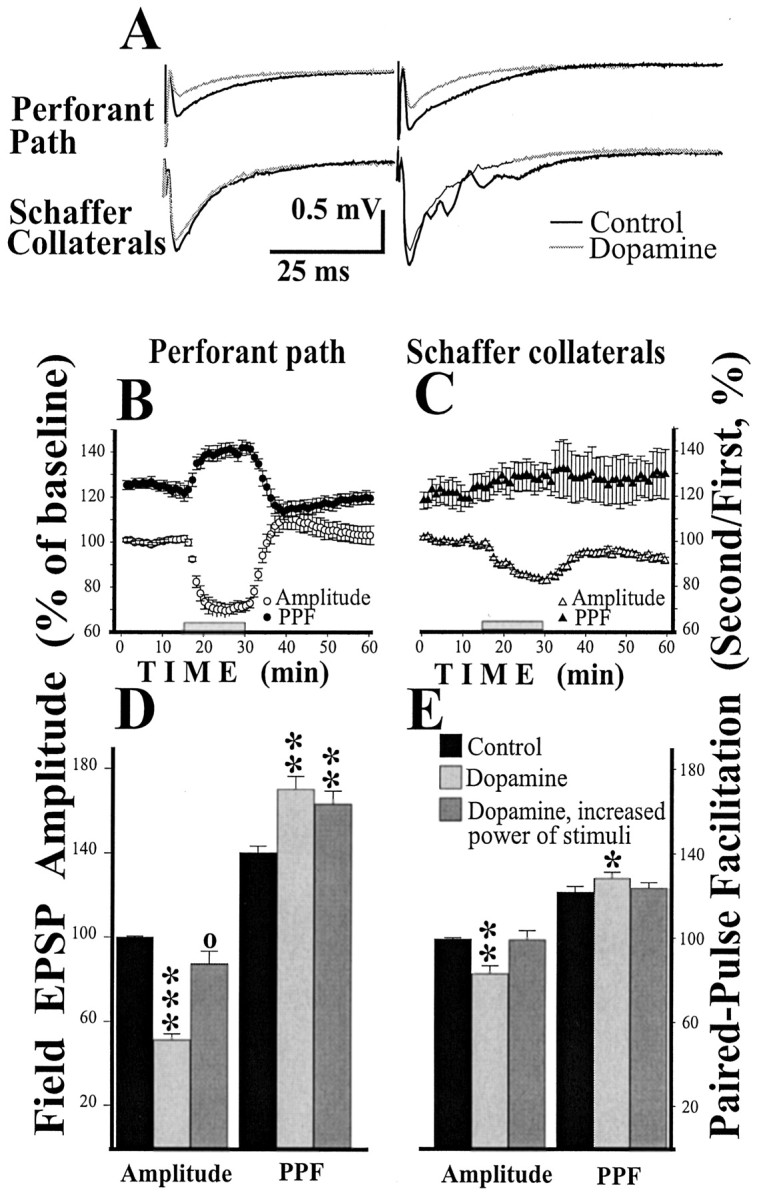
Dopamine significantly increased paired-pulse facilitation in the pp input. A, Individual examples of fEPSP during paired-pulse stimulation. All experiments were done in normal Mg2+ ACSF with 50 μmpicrotoxin added. B, Changes in fEPSP amplitude and PPF of amplitude during dopamine application in the pp. C, Changes in the fEPSP amplitude and PPF in the sc. Apparent failure of reversal in amplitude during the washout of dopamine is probably caused by the small, slow decline of responses (∼10%/hr) generally seen in the sc input during picrotoxin applications. Time of dopamine applications is marked by rectangles. D, Increase of power of stimuli during dopamine application does not affect PPF of the pp fEPSP amplitude. E, Increase of power of stimuli during dopamine application decreases PPF of the sc fEPSP amplitude back to the baseline level. Pathway labels forB and C also refer to Dand E. The dopamine effect (second column) was estimated between 5 and 10 min of dopamine application relative to the baseline before application. Third columns represent measurement between 5 and 10 min after the increase in power of stimuli in the presence of dopamine. Significance relative to baseline in paired ttest: ○p < 0.1; *p < 0.05; **p < 0.01; ***p < 0.001.
Fig. 5.
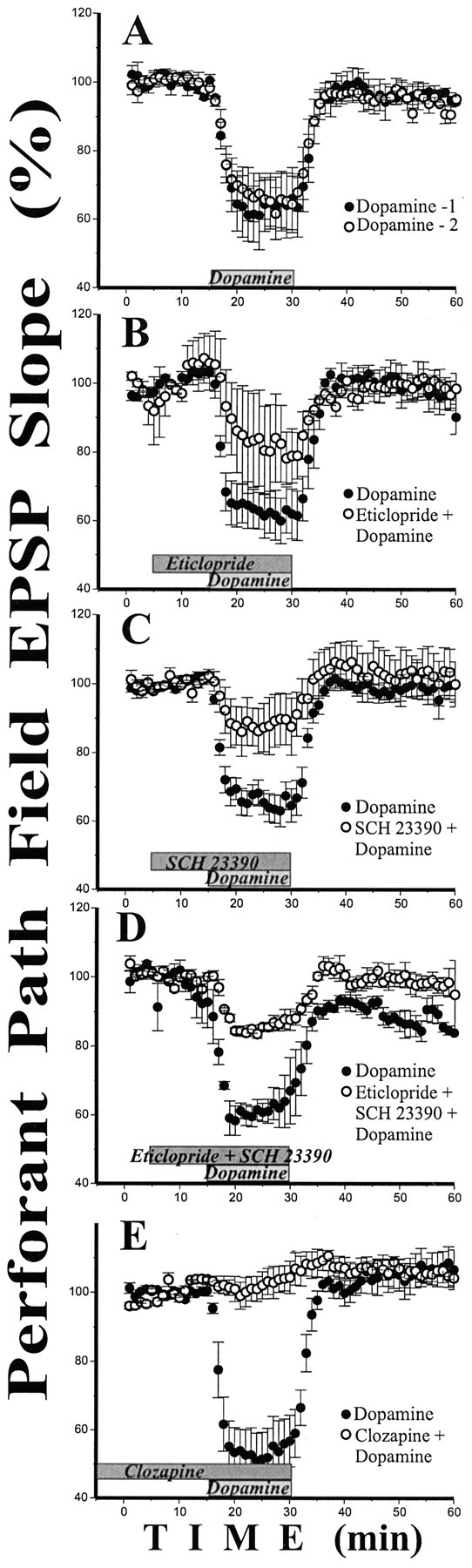
Dopamine antagonists inhibit dopamine-induced suppression of the pp fEPSP. A, Repetitive dopamine applications did not affect dopamine action on the pp fEPSP slope.B, D2 antagonist (−)eticlopride (5 μm) inhibits the effect of dopamine on the pp fEPSP slope.C, D1 antagonist (+)SCH 23390 (5 μm) inhibits the effect of dopamine on the pp fEPSP slope.D, There is no full inhibition of the effect of dopamine on the pp even by a mixture of D1 and D2 antagonists (5 μm each). E, Clozapine (20 μm) completely blocks dopamine-induced suppression of fEPSP slope in the pp. Time of drug applications is marked byrectangles.
RESULTS
Properties of the pp input
To selectively stimulate and record from the perforant path input to CA1, stimulating and recording electrodes were placed in the distal region of the stratum lacunosum-moleculare. Stimulation at this site could indirectly activate CA1 cells through the pp axons that stimulate CA3 or dentate granule cells and subsequent CA3–CA1 transmission (Yeckel and Berger, 1990). Such indirect transmission was eliminated by a cut separating CA1 from CA3 (Fig. 1A). In all our experiments, the pp and the sc inputs were measured simultaneously. The field EPSP evoked by the pp stimulation appeared as a negative deviation of potential at the pp recording electrode and as a positive deviation at the sc electrode. Conversely, stimulation of the sc caused negative deviation of potential at the sc recording electrode and positive deviation at the pp electrode. This indicates that the two electrodes stimulate separate populations of axons that selectively synapse in the two strata.
The general characteristics of the pp and sc responses were quite different. Higher (2–5 times) stimulus strength was required to induce the same fEPSP in pp as in the sc. As stimulus strength increased, the fiber volley and fEPSP amplitude in the sc input increased proportionally, causing the appearance of population spike at a fiber volley amplitude of 0.4–0.5 mV. In the pp, increasing stimulus strength led to an increase in the fiber volley, but the fEPSP easily saturated. At a fiber volley/fEPSP ratio of ∼1, the fEPSP usually could not be further increased. In our standard recordings in the sc, stimulus strength was selected to be 50–60% of the threshold value for evoking a population spike. The pp was stimulated with current sufficient to achieve 50–75% of maximal slope of fEPSP. Under these conditions, the fiber volley amplitude in sc (0.25 ± 0.01 mV) was smaller than in pp (0.78 ± 0.004 mV; p < 0.0001;n = 31; paired t test), whereas the fEPSP amplitude (1.32 ± 0.04 mV) was larger (0.74 ± 0.03 mV;p < 0.0001; n = 31) (Fig.1B,C). Thus, the pp response had much higher fiber volley/fEPSP ratio (1.09 ± 0.07) compared with the sc (0.23 ± 0.04; p > 0.0001;n = 31). This difference is expected from what is known about the connectivity. Schaffer collaterals synapse onto a large fraction of CA1 pyramidal cells. In contrast, perforant path axons make “point to point” topographical connections with CA1 in transverse dimension (Swanson et al., 1987; Lopes da Silva et al., 1990) and may thus pass through a given region without making synaptic contacts. These characteristic features of the pp response are often not present if the stimulating and recording electrodes are placed only slightly closer to the cell body in the proximal region of the stratum-lacunosum moleculare. If these characteristics were not present in a given slice, the electrodes were moved or a new slice was used.
The NMDA component of the fEPSP
The pp input, like the sc, is glutamatergic and has both NMDA and AMPA components (Colbert and Levy, 1992), but it was unclear from previous work whether the relative contribution of the two components was similar in the two pathways. To compare the contribution of the NMDA component in the pp and sc pathways, we first applied the NMDA antagonist, (±)AP-5 (100 μm) in ACSF having a standard Mg2+/Ca2+ ratio. Under these conditions, AP-5 did not affect the sc fEPSP amplitude (p > 0.1; n = 6; Fig.1D) but reduced the pp fEPSP by 18% (p < 0.001; n = 6; Fig.1D).
As a second approach, we measured the effects of NMDA antagonists under conditions that largely removed the Mg2+ block of the NMDA channels (0.1 mm Mg2+ in ACSF). Picrotoxin (50 μm) was added to inhibit the GABAA IPSP. Under these conditions, the excitability of CA1 neurons was increased, causing the appearance of population spikes even with weak stimulation. To minimize the resulting distortion of fEPSP, pyramidal cell excitability was decreased using a low concentration (10 nm) of the Na+ channel blocker tetrodotoxin. As a result, the number and amplitude of spikes in the field potential became much lower, and their latency markedly increased. This allowed more reliable amplitude measurements of the fEPSP (Fig. 1F). The strength of stimuli was adjusted so that fEPSP amplitude was approximately the same in both inputs. Under these conditions, application of 100 μm NMDA antagonist, (±)AP-5 decreased the amplitude of the pp responses by 42% (n = 6; p < 0.001; Fig.1E), whereas the sc response was decreased by only 23% (n = 6; p < 0.001; pairedt test). The difference between inputs was significant (p < 0.05; unpaired t test).
As a third approach, we compared the effects of the AMPA antagonist CNQX on the two pathways in low Mg2+ picrotoxin and tetrodotoxin (Fig. 1E). CNQX (10 μm) left a residual response of only 14% in the sc (p < 0.001; paired t test). The pp response was also strongly decreased by CNQX (p< 0.001; paired t test). However, there was a much larger (24%) residual response in the pp as compared with the sc input (p < 0.05; unpaired t test). All our pharmacological tests are thus in agreement in indicating the NMDA component of transmission is larger in the pp than in the sc pathway.
Selective suppression of the pp fEPSP by dopamine
Application of 1–100 μm dopamine strongly decreased the pp fEPSP (Fig. 2A) measured as a maximal initial slope. The degree of suppression varied with concentration and was maximally 45 ± 2%; IC50 = 3.05 ± 0.5 μm (Fig. 2A). In some experiments the suppression was as large as 75–80% (Fig.2A, inset). There was nearly full recovery within 5–10 min of washout. Dopamine is easily oxidized in solutions, and it was usually not oxidation-protected in our experiments. In control experiments using a very high concentration of antioxidant ascorbic acid (400 μm), normal suppression (∼30%;n = 2) occurred when dopamine (20 μm) was applied. This indicates that the effect is unlikely to be mediated by a breakdown product of dopamine. Because usually dopamine was not oxidation-protected, its actual concentration may have been lower than the nominal value.
Fig. 2.

Concentration dependence of dopamine effect on fEPSP in CA1. A, Concentration–response data (Mean ± SEM) for the dopamine effect on the maximal initial slope of fEPSP in the pp and sigmoidal fit using logistical model.B, Concentration–response data for dopamine effect in the sc input (maximal fEPSP slope). Insets, fEPSP traces from a representative experiment when three concentrations of dopamine (1, 5, and 20 μm) were applied. Experiments were done in regular ACSF. The dopamine effect was estimated between 10 and 15 min of dopamine application relative to the baseline before application. Significance in paired t test: *p < 0.05; ***p < 0.001.
During the same dopamine applications, we also examined the effect of dopamine on the sc transmission. Dopamine produced at most only a minor decrease in the fEPSP slope in the sc input (<10%, Fig.2B). The small dopamine effect on the sc input appeared to have a reversed concentration dependence. The suppression was statistically detectable at 1 μm concentration but was absent at 100 μm. At all tested concentrations (1–100 μm), dopamine did not change the fiber volley in pp and sc inputs, indicating no change in axon excitability. For all further analysis we used a 20 μm concentration of dopamine. We conclude that dopamine powerfully and selectively suppresses the fEPSP of the pp.
Dopaminergic fibers often terminate on GABAergic inhibitory interneurons (Carr and Sesack, 1996; Mrzljak et al., 1996; Delle Donne et al., 1997; Lewis et al., 1998). It was, therefore, possible that the effect of dopamine on the pp was mediated by GABAergic interneurons. In this case, the effect should disappear if GABAergic inhibition was eliminated. Because it has already been shown that GABAB receptors do not affect the pp input (Ault and Nadler, 1982; Colbert and Levy, 1992), we concentrated on GABAA inhibition. In the presence of 50 μmpicrotoxin, a GABAA antagonist, application of 20 μm dopamine caused the same pathway-specific decrease of field EPSP as in control ACSF (p > 0.4; Fig.3A). The only small difference was that during the washout, the pp fEPSP in picrotoxin transiently became higher (p < 0.05) than in the baseline. We conclude that the strong suppression of the pp fEPSP does not depend on GABAergic processes.
Dopamine decreases both NMDA and AMPA components of pp fEPSP, but only the NMDA component in sc
We next checked whether dopamine affects the isolated NMDA and AMPA components of fEPSP. For these experiments we used ACSF with low Mg2+ (0.1 mm) and 50 μmpicrotoxin. First we determined the magnitude of the dopamine (20 μm, 15 min) effect in these new conditions for fEPSP containing both NMDA and AMPA components. The slope of the pp fEPSP was decreased by 47% after dopamine application (n = 5;p < 0.001). In sc there also was a small but significant decrease (10%; n = 5; p < 0.05). To isolate the AMPA component in the same slices, we added a 100 μm concentration of the NMDA receptor antagonist (±)AP-5. Fifteen minutes after the beginning of (±)AP-5 perfusion, dopamine was applied for the second time (Fig. 3B). The AMPA component of the pp fEPSP was decreased by ∼37% (p < 0.001). In sc the isolated AMPA response was not affected by dopamine (n = 5; p> 0.35; Fig. 3B). In a separate series of experiments we isolated the NMDA component using 10 μm CNQX. Dopamine (20 μm, 15 min) decreased the NMDA fEPSP of the pp by 65% (n = 6; p < 0.001; Fig.3C). In sc the NMDA fEPSP was also decreased (25%;n = 6; p < 0.001). These results indicate that dopamine decreases both the AMPA and NMDA components in the pp, but only the NMDA component in the sc pathway.
Dopamine increases paired-pulse facilitation in the pp
It was of interest to study the effect of dopamine on paired-pulse facilitation (PPF). It is generally thought that changes in PPF are caused by a presynaptic action (for example, see Benke et al., 1998). Experiments were performed in the ACSF with regular Mg2+ concentration in the presence of 50 μm picrotoxin to avoid the interference of GABAA inhibition. Stimuli were applied every 30 sec in pairs of pulses with interpulse delay of 50 msec. PPF was calculated for fEPSP using the formula: PPF = second/first * 100%. Dopamine (20 μm, 15 min) decreased the pp responses (Fig.4A,B,D) as described above. Associated with this decrease was an increase in PPF (Fig. 4B,D). Measured after 10 min of dopamine perfusion, PPF in pp was increased by 20% for the amplitude (p < 0.01; paired t test) and by 18% for the slope (p < 0.001). An important control was to determine whether the change in PPF might result secondarily from the reduction in amplitude of the response. To control for this possibility, we conducted six additional experiments (Fig. 4D,E) in which the strength of stimulation was increased to compensate for the reduction in fEPSP produced by dopamine. As before, dopamine increased PPF in the pp (Fig.4D) by 23% for amplitude (p< 0.05) and by 30% for slope (p < 0.01). After the compensatory increase in power of stimulation to return fEPSP to the baseline level, PPF was still significantly higher than in baseline (by 23% for amplitude, p < 0.01, and by 22% for slope, p < 0.01).
In both series of the above experiments, PPF was simultaneously measured in the sc. In the first series, dopamine had no significant effect on PPF in the sc input (p > 0.09 for amplitude and p > 0.1 for slope, paired ttest; n = 6; Fig. 4C). In the second series dopamine slightly (by 5%) increased PPF for amplitude but not for slope (p > 0.1). PPF changes in amplitude returned to a baseline after the compensating increase in the strength of stimuli. The simplest interpretation of these findings is that dopamine acts postsynaptically in the sc and selectively reduces the NMDA component. It therefore has negligible effect on PPF. In contrast, dopamine has a presynaptic effect in the pp and therefore affects PPF.
Dopamine receptor antagonists inhibit dopamine effect in pp
To determine whether D1 or D2 receptors families contribute to the dopamine action on pp we used the D1/D5 antagonist (+)SCH 23390 and several D2 receptor antagonists in regular ACSF. Dopamine was first applied alone to establish the magnitude of dopamine action in each slice. Then after 30 min of washout, dopamine was applied for a second time in the presence of antagonist. Dopamine responses (fEPSP slopes) with and without antagonist were compared in each slice using two-factor ANOVA for repeated measurements followed by post hoc two-tailed paired t test. The time factor was significant in all experiments, but the “time * drug” interaction was not. In all these experiments we did not observe any dopamine effects in the sc input. An important control was to test whether the dopamine response might change as a result of repetitive applications. In four slices we applied dopamine twice (30 min washout interval). No differences appeared between the reactions on the first and the second dopamine application (F = 0.36; p > 0.5; n = 4; Fig.5A).
The D2 receptor family antagonist (−)eticlopride is effective against D2, D3, and D4 dopamine receptors. (−)Eticlopride (5 μm) did not affect baseline synaptic transmission but significantly inhibited the dopamine effect in pp (by 30–35%; F = 5.87; p < 0.03; post hoc pairedt test p < 0.07; n = 4; Fig. 5B). Another D2 family antagonist, the neuroleptic haloperidol, also inhibited dopamine-induced depression (30–35%;F = 7.80; p < 0.01; post hoc p < 0.01; n = 5). Among the D2 receptor family, D4 receptors are most strongly represented in the hippocampus (Mrzljak et al., 1996; Defagot et al., 1997; Joyce and Meador-Woodruff, 1997). The specific D4 antagonist U-101958 (Schlachter et al., 1997) at 20 μm concentration caused a similar inhibition of the dopamine effect (by 25–30%; F = 5.02; p < 0.04; post hoc p< 0.05; n = 4). D1/D5 antagonist (+)SCH 23390 (5 μm) also strongly decreased the effect of dopamine (by 60–65%, F = 29.82; p < 0.0001;post hoc p < 0.05; n = 5; Fig. 5C) without affecting the baseline.
Because neither D1 nor D2 antagonists alone could fully block the dopamine-induced suppression of pp we applied a mixture of the SCH 23390 and eticlopride (5 μm each). The inhibition of dopamine effect by the mixture was very strong (by 70–75%,F = 252.94; p < 0.0001; post hoc p < 0.005; n = 4; Fig.5D) but was still not full. We then tested one of the less specific dopamine antagonists, clozapine, which has affinity to D1, D2, α-adrenergic, and some serotonin receptors (Baldessarini et al., 1992; Bunney, 1992; Coward, 1992; Meltzer, 1994; Jackson and Mohell, 1996; Newman-Tancredi et al., 1997). Clozapine (20 μm) completely blocked the effect of dopamine on the pp (100%,F = 113.12; p < 0.0001;n = 4; Fig. 5E).
DISCUSSION
Dopamine selectively controls inputs to CA1
Our results provide the first evidence that dopamine suppresses the perforant path input to CA1 pyramidal cells. The effect shows all the characteristics of a specific receptor response: saturability, reversibility, and a low half-maximal concentration (Fig. 2). Both AMPA and NMDA components are inhibited. The inhibition of transmission is strong, sometimes up to 80%, and is thus one of the most powerful examples of dopaminergic modulation of transmission so far observed. Dopamine does not affect AMPA-mediated transmission at the nearby Schaffer collateral synapses on the same neurons. The effect is thus pathway-specific. This is the first demonstration of the input-specific dopamine effect in the brain. Pathway-specific effects in CA1 have been also shown for muscarinic (Hasselmo and Schnell, 1994) and GABAB receptors (Ault and Nadler, 1982; Colbert and Levy, 1992; Hasselmo and Schnell, 1994). Both selectively suppress the sc input. In contrast, dopamine selectively suppresses the pp input.
Our findings give some insight to the sites of dopamine action in the stratum lacunosum-moleculare. We found that the dopamine effect in pp was not affected by picrotoxin so does not involve inhibitory interneurons (Fig. 3A). Dopamine suppressed the NMDA and the AMPA components of the pp response (Fig. 3) and increased paired-pulse facilitation (Fig. 4). This would be most consistent with a presynaptic action of dopamine. In the sc, as was shown before, dopamine had no detectable effect on normal AMPA-mediated transmission (Gribkoff and Ashe, 1984; Marciani et al., 1984; Pockett, 1985) but decreased the isolated NMDA response. Furthermore, paired-pulse facilitation was not affected. This pattern is consistent with a postsynaptic site of modulation specific to the NMDA conductance. It should be noted, however, that the NMDA component of the pp response was inhibited significantly stronger than the AMPA. That suggests the possibility that dopamine may also have a postsynaptic action on NMDA receptor channels in the pp, similar to what is observed in the sc input.
We have made some progress in understanding the receptor subtypes involved in the dopamine effect, but the complete picture remains to be worked out. The dopamine effect could be decreased by either D2 or D1 receptor families antagonist (Fig. 5), implicating both D1 and D2 receptors as a major target of dopamine action. This is consistent with the presence of both subtypes of receptors in stratum lacunosum-moleculare (Swanson et al., 1987; Goldsmith and Joyce, 1994). However, even the mixture of both D1 and D2 antagonists did not completely block dopamine-induced suppression of the pp. There is precedent for activation of other monoamine receptors by dopamine (Malenka and Nicoll, 1986; Aguayo and Grossie, 1994). Such action might contribute to the effect of dopamine in the pp input, because adrenergic and serotonergic receptors are also concentrated in the stratum lacunosum-moleculare (Swanson et al., 1987).
D1 and D2 receptors are often coupled to the opposing intercellular processes, but they act similarly in mediating the dopamine effect on the pp. There is precedent for cooperative action of D1 and D2 receptors at both the behavioral and the cellular levels (Bertorello et al., 1990; Piomelli et al., 1991; Calabresi et al., 1992; Momiyama et al., 1993a,b; Surmeier and Kitai, 1993; Keefe and Gerfen, 1995;Wan et al., 1996; Hu and White, 1997; Shi et al., 1997). Our data leaves open the possibility that D1 and D2 receptors could modulate transmission at different sites and perhaps through complex processes. For instance, D2 type receptors might inhibit glutamate release and/or modify postsynaptic targets on pyramidal cell dendrites, whereas D1 type might increase local noradrenaline (Hajos-Korcsok and Sharp, 1996) or acetylcholine (Acquas et al., 1994; Hersi et al., 1995) release. These neuromodulators in turn might affect the pp transmission.
We find that the atypical antipsychotic agent clozapine completely abolishes the effect of dopamine on the pp. Clozapine is one of the most effective drugs in treating schizophrenia. It is known that in addition to blocking a broad range of dopamine receptors, clozapine also acts as an antagonist to α-adrenergic, and some serotonin receptors (Baldessarini et al., 1992; Bunney, 1992; Coward, 1992;Meltzer, 1994; Jackson and Mohell, 1996; Newman-Tancredi et al., 1997). The broad spectrum of clozapine actions is thought to contribute to its antipsychotic function. Because the effect of dopamine is so large at the pp and because the effectiveness of clozapine is so high, the pp may serve as excellent site for investigating the details of the action of clozapine. Moreover, there are reasons for suspecting that this site might be important in schizophrenia (see functional significance).
Differences between the pp and the sc inputs
Several lines of results indicate that NMDA-mediated transmission is more important at the pp than at the sc. In regular ACSF, the fEPSP of Schaffer collateral is hardly affected by the NMDA antagonist. In contrast, the pp response was reduced by ∼20% (Fig. 1). We also observed this difference in low Mg2+ solutions that should greatly reduce the voltage-dependent block of NMDA receptors by Mg2+. Under these conditions, we similarly found that blocking NMDA channels produced a much larger fractional reduction in the amplitude of the fEPSP in the pp than in the sc. Conversely, an AMPA antagonist, CNQX, produced a much larger block of the fEPSP in the sc than in the pp. Our conclusion that the pp has a larger NMDA component of transmission than the sc is consistent with data shown byColbert and Levy (1992). The pp input to the CA3 has also been shown to have a large NMDA component (Berzhanskaya et al., 1998).
Our finding of the differences in neuromodulation and transmission in stratum radiatum and stratum lacunosum-moleculare is generally consistent with available histological data. It was shown that these regions have substantial differences in receptor and channel distribution. D1 and D2 dopamine, 5-HT1 serotonin, α-adrenergic, and nicotinic receptors seem to concentrate in the stratum lacunosum-moleculare (Swanson et al., 1987; Goldsmith and Joyce, 1994). Putatively presynaptic high-conductance Ca2+-dependent K+ channels (Knaus et al., 1996) and metabotropic glutamate receptors mGgluR2 (Neki et al., 1996) also are more concentrated in the area of the pp input. The GluRD subunit of AMPA receptors, and muscarinic receptors, are more concentrated in the area of the sc input (Baude et al., 1995). It is therefore reasonable that dopamine inhibits the pp synaptic transmission stronger than the sc, but the activation of muscarinic receptors has stronger effects in the sc than in the pp (Hasselmo and Schnell, 1994).
We found that for a given fiber volley, the amplitude of the fEPSP in the pp was smaller than in the sc input. This seems to be consistent with available data. Schaffer collaterals give massive divergent (and convergent) input to a large number of pyramidal cells. The pp, on the other hand, mostly consists of highly specific point-to-point (nondivergent) corticohippocampal connections (Swanson et al., 1987;Lopes da Silva et al., 1990). Thus, many of the stimulated axons traveling through the stratum lacunosum-moleculare in the vicinity of the recording electrode may not synapse on the local population of pyramidal cells.
Functional role of dopaminergic modulation of perforant path
The “direct” pp connection from entorhinal cortex to CA1 is the main source of specific sensory information to CA1 (Vinogradova, 1984; McNaughton et al., 1989). Despite its distal location, it can control the CA1 output acting directly and via inhibitory interneurons.In vitro and in vivo studies show that the stimulation of the pp can induce asynchronous spiking in CA1 pyramidal neurons (Spencer and Kandel, 1962; Bragin and Otmakhov, 1979a,b;Doller and Weight, 1982; Vinogradova, 1984; Yeckel and Berger, 1990). Excitation is usually followed by strong inhibition (Bragin and Otmakhov, 1979a; Empson and Heinemann, 1995; Levy et al., 1995) that temporarily blocks the effects of the indirect sc input.
The “indirect” path via the dentate and CA3 does not carry specific sensory information because these regions do not generally respond to specific features of the sensory stimulus (Vinogradova, 1984). Direct evidence shows that the responses of the dentate gyrus or CA3 fields depend strongly on training (Deadwyler and Hampson, 1997). An emerging view of the dentate gyrus/CA3 is that it uses the current sensory information to make predictions based on the contents of long-term memory (Jensen et al., 1996). These predictions would then be sent to CA1 by the sc, where they converge with actual sensory information arriving from the pp. The comparison of the information brought by these two inputs leads to the detection of novelty or mismatch from expectations (Vinogradova, 1984; Levy, 1989; Hasselmo and Schnell, 1994). Our results suggest that dopamine hyperfunction or NMDA hypofunction would isolate CA1 from specific sensory information coming from the entorhinal cortex via the pp and lead to an error in the mismatch computation.
Both dopaminergic hyperfunction (Joyce, 1993; Gray et al., 1995; Joyce and Meador-Woodruff, 1997) and NMDA hypofunction (Carlsson, 1995;Halberstadt, 1995; Olney and Farber, 1995; Javitt, 1996) are thought to underlie schizophrenia. There is converging evidence that schizophrenia involves hippocampal malfunction (Shenton et al., 1992; Bogerts and Falkai, 1995; Fukuzako et al., 1995; Heckers et al., 1998). Specifically, there is a decrease in temporolimbic volume that correlates with the severity of frontal syndromes (Bilder et al., 1995;Turetsky et al., 1995). Decreased hippocampal function is associated with delusional syndrome (Schroder et al., 1995), and the degree of thought disorder correlates with the asymmetry in phosphorous metabolism in temporal cortices, specifically, hyperactivation of the right compared with the left temporal region (Deicken et al., 1995). Conscious recollection deficit in schizophrenic patients is associated with reduced hippocampal activation (Heckers et al., 1998). Appearance of verbal hallucinations in schizophrenia is associated with hippocampal activation (Silbersweig et al., 1995).
The only previous connection between dopamine and hippocampal function were the reports that dopamine could affect synaptic plasticity in the sc, specifically facilitate LTP (Frey et al., 1993; Otmakhova and Lisman, 1996), and inhibit depotentiation (Otmakhova and Lisman, 1998). Through these cellular actions, dopamine hyperfunction might increase random memory associations and disrupt the ability to inhibit incorrect associations, which is known to occur in schizophrenia. Although schizophrenia may involve long-term memory impairments, it clearly involves information-processing aberrations (Schroder et al., 1995;Bazin and Perruchet, 1996; Brebion et al., 1996). It may therefore be important that dopamine and NMDA antagonist can reduce the cortical input to CA1, and thereby affect information processing. This suggests a connection between the three known features of schizophrenia: the disruption of corticohippocampal interactions, dopamine hyperfunction, and NMDA hypofunction.
Footnotes
This work was supported by Grants 2R01 NS27337/09 and 1R01 NS35083/01 from the National Institutes of Health and Grant RG3-96-015 from the Alzheimer Association to J. Lisman, and Grant 1F32 MH11720–01 from the National Institutes of Health, the National Alliance for Research on Schizophrenia and Depression Young Investigator Award, and the Scottish Rite Schizophrenia Research Program, NMJ, to N. Otmakhova. The authors appreciate the support from W. M. Keck Foundation and are very grateful to Dr. Robert Greene for remarks on this manuscript.
Correspondence should be addressed to John E. Lisman, Biology Department, CCS, Brandeis University, 415 South Street, Waltham, MA 02254.
REFERENCES
- 1.Acquas E, Day JC, Fibiger HC. The potent and selective dopamine D1 receptor agonist A-77636 increases cortical and hippocampal acetylcholine release in the rat. Eur J Pharmacol. 1994;260:85–87. doi: 10.1016/0014-2999(94)90013-2. [DOI] [PubMed] [Google Scholar]
- 2.Aguayo LG, Grossie J. Dopamine inhibits a sustained calcium current through activation of alpha adrenergic receptors and a GTP-binding protein in adult rat sympathetic neurons. J Pharmacol Exp Ther. 1994;269:503–508. [PubMed] [Google Scholar]
- 3.Ault B, Nadler JV. Baclofen selectively inhibits transmission at synapses made by axons of CA3 pyramidal cells in the hippocampal slice. J Pharmacol Exp Ther. 1982;223:291–297. [PubMed] [Google Scholar]
- 4.Baldessarini RJ, Huston-Lyons D, Campbell A, Marsh E, Cohen BM. Do central antiadrenergic actions contribute to the atypical properties of clozapine? Br J Psychiatry [Suppl] 1992;17:12–16. [PubMed] [Google Scholar]
- 5.Baude A, Nusser Z, Molnar E, McIlhinney RA, Somogyi P. High-resolution immunogold localization of AMPA type glutamate receptor subunits at synaptic and non-synaptic sites in rat hippocampus. Neuroscience. 1995;69:1031–1055. doi: 10.1016/0306-4522(95)00350-r. [DOI] [PubMed] [Google Scholar]
- 6.Bazin N, Perruchet P. Implicit and explicit associative memory in patients with schizophrenia. Schizophr Res. 1996;22:241–248. doi: 10.1016/s0920-9964(96)00052-7. [DOI] [PubMed] [Google Scholar]
- 7.Benke TA, Luthi A, Isaac JT, Collingridge GL. Modulation of AMPA receptor unitary conductance by synaptic activity. Nature. 1998;393:793–797. doi: 10.1038/31709. [DOI] [PubMed] [Google Scholar]
- 8.Bernabeu R, Bevilaqua L, Ardenghi P, Bromberg E, Schmitz P, Bianchin M, Izquierdo I, Medina JH. Involvement of hippocampal cAMP/cAMP-dependent protein kinase signaling pathways in a late memory consolidation phase of aversively motivated learning in rats. Proc Natl Acad Sci USA. 1997;94:7041–7046. doi: 10.1073/pnas.94.13.7041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bertorello AM, Hopfield JF, Aperia A, Greengard P. Inhibition by dopamine of (Na(+)+K+)ATPase activity in neostriatal neurons through D1 and D2 dopamine receptor synergism. Nature. 1990;347:386–388. doi: 10.1038/347386a0. [DOI] [PubMed] [Google Scholar]
- 10.Berzhanskaya J, Urban NN, Barrionuevo G. Electrophysiological and pharmacological characterization of the direct perforant path input to hippocampal area CA3. J Neurophysiol. 1998;79:2111–2118. doi: 10.1152/jn.1998.79.4.2111. [DOI] [PubMed] [Google Scholar]
- 11.Bilder RM, Bogerts B, Ashtari M, Wu H, Alvir JM, Jody D, Reiter G, Bell L, Lieberman JA. Anterior hippocampal volume reductions predict frontal lobe dysfunction in first episode schizophrenia. Schizophr Res. 1995;17:47–58. doi: 10.1016/0920-9964(95)00028-k. [DOI] [PubMed] [Google Scholar]
- 12.Bogerts B, Falkai P. Postmortem brain abnormalities in schizophrenia. In: Shriqui CL, Nasrallah HA, editors. Contemporary issues in the treatment of schizophrenia. American Psychiatric; Washington, DC: 1995. pp. 43–59. [Google Scholar]
- 13.Bragin AG, Otmakhov NA. Comparison of direct influences of the perforant path on hippocampal CA1 and CA3 neurons in vitro. Neurophysiology. 1979a;11:220–225. [PubMed] [Google Scholar]
- 14.Bragin AG, Otmakhov NA. Investigation of heterosynaptic interactions in hippocampal areas CA1 and CA3 in vitro. Neurophysiology. 1979b;11:389–396. [PubMed] [Google Scholar]
- 15.Brebion G, Smith MJ, Gorman JM, Amador X. Reality monitoring failure in schizophrenia: the role of selective attention. Schizophr Res. 1996;22:173–180. doi: 10.1016/s0920-9964(96)00054-0. [DOI] [PubMed] [Google Scholar]
- 16.Bunney BS. Clozapine: a hypothesised mechanism for its unique clinical profile. Br J Psychiatry [Suppl]: 1992;17:17–21. [PubMed] [Google Scholar]
- 17.Buzsaki G. Two-stage model of memory trace formation: a role for “noisy” brain states. Neuroscience. 1989;31:551–570. doi: 10.1016/0306-4522(89)90423-5. [DOI] [PubMed] [Google Scholar]
- 18.Calabresi P, Maj R, Mercuri NB, Bernardi G. Coactivation of D1 and D2 dopamine receptors is required for long-term synaptic depression in the striatum. Neurosci Lett. 1992;142:95–99. doi: 10.1016/0304-3940(92)90628-k. [DOI] [PubMed] [Google Scholar]
- 19.Carlsson A. Neurocircuitries and neurotransmitter interactions in schizophrenia. Int Clin Psychopharmacol [Suppl] 1995;3:21–28. [PubMed] [Google Scholar]
- 20.Carr DB, Sesack SR. Hippocampal afferents to the rat prefrontal cortex: synaptic targets and relation to dopamine terminals. J Comp Neurol. 1996;369:1–15. doi: 10.1002/(SICI)1096-9861(19960520)369:1<1::AID-CNE1>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 21.Colbert CM, Levy WB. Electrophysiological and pharmacological characterization of perforant path synapses in CA1: mediation by glutamate receptors. J Neurophysiol. 1992;68:1–8. doi: 10.1152/jn.1992.68.1.1. [DOI] [PubMed] [Google Scholar]
- 22.Coward DM. General pharmacology of clozapine. Br J Psychiatry [Suppl]: 1992;17:5–11. [PubMed] [Google Scholar]
- 23.Deadwyler SA, Hampson RE. The significance of neural ensemble codes during behavior and cognition. Annu Rev Neurosci. 1997;20:217–244. doi: 10.1146/annurev.neuro.20.1.217. [DOI] [PubMed] [Google Scholar]
- 24.Defagot MC, Malchiodi EL, Villar MJ, Antonelli MC. Distribution of D4 dopamine receptor in rat brain with sequence-specific antibodies. Brain Res Mol Brain Res. 1997;45:1–12. doi: 10.1016/s0169-328x(96)00235-5. [DOI] [PubMed] [Google Scholar]
- 25.Deicken RF, Calabrese G, Merrin EL, Vinogradov S, Fein G, Weiner MW. Asymmetry of temporal lobe phosphorous metabolism in schizophrenia: a 31 phosphorous magnetic resonance spectroscopic imaging study. Biol Psychiatry. 1995;38:279–286. doi: 10.1016/0006-3223(94)00372-A. [DOI] [PubMed] [Google Scholar]
- 26.Delle Donne KT, Sesack SR, Pickel VM. Ultrastructural immunocytochemical localization of the dopamine D2 receptor within GABAergic neurons of the rat striatum. Brain Res. 1997;746:239–255. doi: 10.1016/s0006-8993(96)01226-7. [DOI] [PubMed] [Google Scholar]
- 27.Dolleman-Van der Weel MJ, Lopes da Silva FH, Witter MP. Nucleus reuniens thalami modulates activity in hippocampal field CA1 through excitatory and inhibitory mechanisms. J Neurosci. 1997;17:5640–5650. doi: 10.1523/JNEUROSCI.17-14-05640.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Doller HJ, Weight FF. Perforant pathway activation of hippocampal CA1 stratum pyramidale neurons: electrophysiological evidence for a direct pathway. Brain Res. 1982;237:1–13. doi: 10.1016/0006-8993(82)90553-4. [DOI] [PubMed] [Google Scholar]
- 29.Eichenbaum H. Declarative memory: insights from cognitive neurobiology. Annu Rev Psychol. 1997;48:547–572. doi: 10.1146/annurev.psych.48.1.547. [DOI] [PubMed] [Google Scholar]
- 30.Empson RM, Heinemann U. Perforant path connections to area CA1 are predominantly inhibitory in the rat hippocampal-entorhinal cortex combined slice preparation. Hippocampus. 1995;5:104–107. doi: 10.1002/hipo.450050203. [DOI] [PubMed] [Google Scholar]
- 31.Frey U, Huang Y-Y, Kandel E. Effects of cAMP stimulate a late stage of LTP in hippocampal CA1 neurons. Science. 1993;260:1661–1664. doi: 10.1126/science.8389057. [DOI] [PubMed] [Google Scholar]
- 32.Fukuzako H, Kodama S, Fukuzako T, Yamada K, Hokazono Y, Ueyama K, Hashiguchi T, Takenouchi K, Takigawa M, Takeuchi K, Manchandza S. Shortening of the hippocampal formation in first-episode schizophrenic patients. Psychiatry Clin Neurosci. 1995;49:157–161. doi: 10.1111/j.1440-1819.1995.tb02221.x. [DOI] [PubMed] [Google Scholar]
- 33.Gasbarri A, Sulli A, Innocenzi R, Pacitti C, Brioni JD. Spatial memory impairment induced by lesion of the mesohippocampal dopaminergic system in the rat. Neuroscience. 1996;74:1037–1044. doi: 10.1016/0306-4522(96)00202-3. [DOI] [PubMed] [Google Scholar]
- 34.Goldsmith S, Joyce J. Dopamine D2 receptor expression in hippocampus and parahippocampal cortex of rat, cat, and human in relation to tyrosine hydroxylase-immunoreactive fibers. Hippocampus. 1994;4:354–373. doi: 10.1002/hipo.450040318. [DOI] [PubMed] [Google Scholar]
- 35.Gray NS, Pilowsky LS, Gray JA, Kerwin RW. Latent inhibition in drug naive schizophrenics: relationship to duration of illness and dopamine D2 binding using SPET. Schizophr Res. 1995;17:95–107. doi: 10.1016/0920-9964(95)00034-j. [DOI] [PubMed] [Google Scholar]
- 36.Grecksch G, Matthies H. Involvement of hippocampal dopaminergic receptors in memory consolidation in rats. In: Marsan C, Matthies H, editors. Neuronal plasticity and memory formation. Raven; New York: 1982. pp. 353–357. [Google Scholar]
- 37.Gribkoff VK, Ashe JH. Modulation by dopamine of population responses and cell membrane properties of hippocampal CA1 neurons in vitro. Brain Res. 1984;292:327–338. doi: 10.1016/0006-8993(84)90768-6. [DOI] [PubMed] [Google Scholar]
- 38.Hajos-Korcsok E, Sharp T. 8-OH-DPAT-induced release of hippocampal noradrenaline in vivo: evidence for a role of both 5-HT1A and dopamine D1 receptors. Eur J Pharmacol. 1996;314:285–291. doi: 10.1016/s0014-2999(96)00560-2. [DOI] [PubMed] [Google Scholar]
- 39.Halberstadt AL. The phencyclidine-glutamate model of schizophrenia. Clin Neuropharmacol. 1995;18:237–249. doi: 10.1097/00002826-199506000-00004. [DOI] [PubMed] [Google Scholar]
- 40.Hasselmo ME, Schnell E. Laminar selectivity of the cholinergic suppression of synaptic transmission in rat hippocampal region CA1: computational modeling and brain slice physiology. J Neurosci. 1994;14:3898–3914. doi: 10.1523/JNEUROSCI.14-06-03898.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Heckers S, Rauch S, Golf D, Savage G, Schaster D, Fischman A, Alpert N. Impaired recruitment of the hippocampus during conscious recollection in schizophrenia. Nat Neurosci. 1998;1:318–323. doi: 10.1038/1137. [DOI] [PubMed] [Google Scholar]
- 42.Hersi AI, Richard JW, Gaudreau P, Quirion R. Local modulation of hippocampal acetylcholine release by dopamine D1 receptors: a combined receptor autoradiography and in vivo dialysis study. J Neurosci. 1995;15:7150–7157. doi: 10.1523/JNEUROSCI.15-11-07150.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hsu KS. Characterization of dopamine receptors mediating inhibition of excitatory synaptic transmission in the rat hippocampal slice. J Neurophysiol. 1996;76:1887–1895. doi: 10.1152/jn.1996.76.3.1887. [DOI] [PubMed] [Google Scholar]
- 44.Hu XT, White FJ. Dopamine enhances glutamate-induced excitation of rat striatal neurons by cooperative activation of D1 and D2 class receptors. Neurosci Lett. 1997;224:61–65. doi: 10.1016/s0304-3940(97)13443-7. [DOI] [PubMed] [Google Scholar]
- 45.Jackson D, Mohell N. A review of the pharmacology of new antipsychotic drugs. In: Stone T, editor. CNS neurotransmitters and neuromodulators: dopamine. CRS; Boca Raton: 1996. pp. 185–200. [Google Scholar]
- 46.Javitt DC. Glutamate receptors and schizophrenia: opportunities and caveats [news]. Mol Psychiatry. 1996;1:16–17. [PubMed] [Google Scholar]
- 47.Jensen O, Idiart M, Lisman JE. Physiologically realistic formation of autoassociative memory in networks with theta/gamma oscillations: role of fast NMDA channels. Learning and Memory. 1996;3:243–256. doi: 10.1101/lm.3.2-3.243. [DOI] [PubMed] [Google Scholar]
- 48.Joyce JN. The dopamine hypothesis of schizophrenia: limbic interactions with serotonin and norepinephrine. Psychopharmacology (Berl) 1993;112:S16–S34. doi: 10.1007/BF02245004. [DOI] [PubMed] [Google Scholar]
- 49.Joyce JN, Meador-Woodruff JH. Linking the family of D2 receptors to neuronal circuits in human brain: insights into schizophrenia. Neuropsychopharmacology. 1997;16:375–384. doi: 10.1016/S0893-133X(96)00276-X. [DOI] [PubMed] [Google Scholar]
- 50.Keefe KA, Gerfen CR. D1–D2 dopamine receptor synergy in striatum: effects of intrastriatal infusions of dopamine agonists and antagonists on immediate early gene expression. Neuroscience. 1995;66:903–913. doi: 10.1016/0306-4522(95)00024-d. [DOI] [PubMed] [Google Scholar]
- 51.Knaus HG, Schwarzer C, Koch RO, Eberhart A, Kaczorowski GJ, Glossmann H, Wunder F, Pongs O, Garcia ML, Sperk G. Distribution of high-conductance Ca(2+)-activated K+ channels in rat brain: targeting to axons and nerve terminals. J Neurosci. 1996;16:955–963. doi: 10.1523/JNEUROSCI.16-03-00955.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Levy WB. A computational approach to hippocampal function. In: Hawkins RD, Bower GH, editors. Computational models of learning in simple neural systems. Academic; New York: 1989. pp. 243–305. [Google Scholar]
- 53.Levy WB, Colbert CM, Desmond NL. Another network model bites the dust: entorhinal inputs are no more than weakly excitatory in the hippocampal CA1 region. Hippocampus. 1995;5:137–140. doi: 10.1002/hipo.450050209. [DOI] [PubMed] [Google Scholar]
- 54.Lewis DA, Sesack SR, Levey AI, Rosenberg DR. Dopamine axons in primate prefrontal cortex: specificity of distribution, synaptic targets, and development. Adv Pharmacol. 1998;42:703–706. doi: 10.1016/s1054-3589(08)60845-5. [DOI] [PubMed] [Google Scholar]
- 55.Lopes da Silva FH, Witter MP, Boeijinga PH, Lohman AH. Anatomic organization and physiology of the limbic cortex. Physiol Rev. 1990;70:453–511. doi: 10.1152/physrev.1990.70.2.453. [DOI] [PubMed] [Google Scholar]
- 56.Malenka RC, Nicoll RA. Dopamine decreases the calcium-activated afterhyperpolarization in hippocampal CA1 pyramidal cells. Brain Res. 1986;379:210–215. doi: 10.1016/0006-8993(86)90773-0. [DOI] [PubMed] [Google Scholar]
- 57.Marciani MG, Calabresi P, Stanzione P, Bernardi G. Dopaminergic and noradrenergic responses in the hippocampal slice preparation. Evidence for different receptors. Neuropharmacology. 1984;23:303–307. doi: 10.1016/0028-3908(84)90191-6. [DOI] [PubMed] [Google Scholar]
- 58.McNaughton BL, Barnes CA, Meltzer J, Sutherland RJ. Hippocampal granule cells are necessary for normal spatial learning but not for spatially-selective pyramidal cell discharge. Exp Brain Res. 1989;76:485–496. doi: 10.1007/BF00248904. [DOI] [PubMed] [Google Scholar]
- 59.Meltzer HY. An overview of the mechanism of action of clozapine. J Clin Psychiatry [Suppl] 1994;55:47–52. [PubMed] [Google Scholar]
- 60.Momiyama T, Sasa M, Takaori S. Enhancement of D2 receptor agonist-induced inhibition by D1 receptor agonist in the ventral tegmental area. Br J Pharmacol. 1993a;110:713–718. doi: 10.1111/j.1476-5381.1993.tb13870.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Momiyama T, Todo N, Sasa M. A mechanism underlying dopamine D1 and D2 receptor-mediated inhibition of dopaminergic neurones in the ventral tegmental area in vitro. Br J Pharmacol. 1993b;109:933–940. doi: 10.1111/j.1476-5381.1993.tb13710.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mrzljak L, Bergson C, Pappy M, Huff R, Levenson R, Goldman-Rakic PS. Localization of dopamine D4 receptors in GABAergic neurons of the primate brain. Nature. 1996;381:245–248. doi: 10.1038/381245a0. [DOI] [PubMed] [Google Scholar]
- 63.Neki A, Ohishi H, Kaneko T, Shigemoto R, Nakanishi S, Mizuno N. Pre- and postsynaptic localization of a metabotropic glutamate receptor, mGluR2, in the rat brain: an immunohistochemical study with a monoclonal antibody. Neurosci Lett. 1996;202:197–200. doi: 10.1016/0304-3940(95)12248-6. [DOI] [PubMed] [Google Scholar]
- 64.Newman-Tancredi A, Audinot-Bouchez V, Gobert A, Millan MJ. Noradrenaline and adrenaline are high affinity agonists at dopamine D4 receptors. Eur J Pharmacol. 1997;319:379–83. doi: 10.1016/s0014-2999(96)00985-5. [DOI] [PubMed] [Google Scholar]
- 65.O’Keefe J. Hippocampus, theta, and spatial memory. Curr Opin Neurobiol. 1993;3:917–924. doi: 10.1016/0959-4388(93)90163-s. [DOI] [PubMed] [Google Scholar]
- 66.Olney JW, Farber NB. Glutamate receptor dysfunction and schizophrenia. Arch Gen Psychiatry. 1995;52:998–1007. doi: 10.1001/archpsyc.1995.03950240016004. [DOI] [PubMed] [Google Scholar]
- 67.Otmakhova NA, Lisman JE. D1/D5 dopamine receptor activation increases the magnitude of early long-term potentiation at CA1 hippocampal synapses. J Neurosci. 1996;16:7478–7486. doi: 10.1523/JNEUROSCI.16-23-07478.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Otmakhova NA, Lisman JE. D1/D5 dopamine receptors inhibit depotentiation at CA1 synapses via cAMP-dependent mechanism. J Neurosci. 1998;18:1270–1279. doi: 10.1523/JNEUROSCI.18-04-01270.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Packard MG, White NM. Dissociation of hippocampus and caudate nucleus memory systems by posttraining intracerebral injection of dopamine agonists. Behav Neurosci. 1991;105:295–306. doi: 10.1037//0735-7044.105.2.295. [DOI] [PubMed] [Google Scholar]
- 70.Piomelli D, Pilon C, Giros B, Sokoloff P, Martres MP, Schwartz JC. Dopamine activation of the arachidonic acid cascade as a basis for D1/D2 receptor synergism. Nature. 1991;353:164–167. doi: 10.1038/353164a0. [DOI] [PubMed] [Google Scholar]
- 71.Pockett S. Dopamine changes the shape of action potentials in hippocampal pyramidal cells. Brain Res. 1985;342:386–390. doi: 10.1016/0006-8993(85)91143-6. [DOI] [PubMed] [Google Scholar]
- 72.Schlachter SK, Poel TJ, Lawson CF, Dinh DM, Lajiness ME, Romero AG, Rees SA, Duncan JN, Smith MW. Substituted 4-aminopiperidines having high in vitro affinity and selectivity for the cloned human dopamine D4 receptor. Eur J Pharmacol. 1997;322:283–286. doi: 10.1016/s0014-2999(97)00013-7. [DOI] [PubMed] [Google Scholar]
- 73.Schroder J, Buchsbaum MS, Siegel BV, Geider FJ, Niethammer R. Structural and functional correlates of subsyndromes in chronic schizophrenia. Psychopathology. 1995;28:38–45. doi: 10.1159/000284898. [DOI] [PubMed] [Google Scholar]
- 74.Shenton ME, Kikinis R, Jolesz FA, Pollak SD, LeMay M, Wible CG, Hokama H, Martin J, Metcalf D, Coleman M, McCarley RW. Abnormalities of the left temporal lobe and thought disorder in schizophrenia. A quantitative magnetic resonance imaging study. N Engl J Med. 1992;327:604–612. doi: 10.1056/NEJM199208273270905. [DOI] [PubMed] [Google Scholar]
- 75.Shi WX, Smith PL, Pun CL, Millet B, Bunney BS. D1–D2 interaction in feedback control of midbrain dopamine neurons. J Neurosci. 1997;17:7988–7994. doi: 10.1523/JNEUROSCI.17-20-07988.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Silbersweig DA, Stern E, Frith C, Cahill C, Holmes A, Grootoonk S, Seaward J, McKenna P, Chua SE, Schnorr L, Jones T, Frackowiak RSJ. A functional neuroanatomy of hallucinations in schizophrenia. Nature. 1995;378:176–179. doi: 10.1038/378176a0. [DOI] [PubMed] [Google Scholar]
- 77.Skaggs WE, McNaughton BL. Computational approaches to hippocampal function. Curr Opin Neurobiol. 1992;2:209–211. doi: 10.1016/0959-4388(92)90014-c. [DOI] [PubMed] [Google Scholar]
- 78.Spencer W, Kandel E. Hippocampal neuron responses in relation to normal and abnormal function. In: Passouant P, editor. Physiologiede l’hippocampe. Centre National de la Recherche Scientifique; Paris: 1962. pp. 71–103. [Google Scholar]
- 79.Surmeier DJ, Kitai ST. D1 and D2 dopamine receptor modulation of sodium and potassium currents in rat neostriatal neurons. Prog Brain Res. 1993;99:309–324. doi: 10.1016/s0079-6123(08)61354-0. [DOI] [PubMed] [Google Scholar]
- 80.Swanson L, Kohler C, Bjorklund A. The limbic region. I: The septohippocampal system. In: Bjorklund A, Hokfelt T, Swanson L, editors. Integrated systems of the CNS. Elsevier Science; New York: 1987. pp. 125–269. [Google Scholar]
- 81.Turetsky B, Cowell PE, Gur RC, Grossman RI, Shtasel DL, Gur RE. Frontal and temporal lobe brain volumes in schizophrenia. Relationship to symptoms and clinical subtype. Arch Gen Psychiatry. 1995;52:1061–1070. doi: 10.1001/archpsyc.1995.03950240079013. [DOI] [PubMed] [Google Scholar]
- 82.Vinogradova O. Functional organization of the limbic system in the process of registration of information: facts and hypotheses. In: Isaacson RL, Pribram KH, editors. The hippocampus. Plenum; New York: 1984. pp. 1–69. [Google Scholar]
- 83.Wan FJ, Taaid N, Swerdlow NR. Do D1/D2 interactions regulate prepulse inhibition in rats? Neuropsychopharmacology. 1996;14:265–274. doi: 10.1016/0893-133X(95)00133-X. [DOI] [PubMed] [Google Scholar]
- 84.Yeckel MF, Berger TW. Feedforward excitation of the hippocampus by afferents from the entorhinal cortex: redefinition of the role of the trisynaptic pathway. Proc Natl Acad Sci USA. 1990;87:5832–5836. doi: 10.1073/pnas.87.15.5832. [DOI] [PMC free article] [PubMed] [Google Scholar]