

Abstract
In vitro and in vivo experiments were conducted to study the effects of synbiotic supplementation on Salmonella enterica ser. Enteritidis (SE) proliferation, cecal content load, and broiler carcass contamination. Lactobacillus reuteri, Enterococcus faecium, Bifidobacterium animalis, and Pediococcus acidilactici culture supernatants decreased (P < 0.05) the in vitro proliferation of SE at 1:1 supernatant: pathogen dilution. A total of 240 Cobb-500 broiler chicks were randomly allotted to three treatment groups (8 replicates/group with 10 birds/replicate): control (basal diet), antibiotic (Virginiamycin at 20 mg/kg feed), synbiotic (PoultryStar® ME at 0.5 g/kg feed containing L. reuteri, E. faecium, B. animalis, P. acidilactici and a Fructooligosaccharide) from day of hatch. At 21 d of age, all birds in experimental groups were orally inoculated with 250 μl of 1 X 109 CFU SE. Antibiotic supplementation increased (P < 0.05) body weight and feed consumption, compared to the control group. Birds in the synbiotic supplementation had intermediate body weight and feed consumption that were not significantly different from both the control and antibiotic group at 42 d of age in SE infected birds. No significant effects were observed in feed efficiency at 42 d of age among the groups. Antibiotic and synbiotic supplementation decreased (P < 0.05) SE load in cecal contents by 0.90 and 0.85 log units/ g and carcass SE load by 1.4 and 1.5 log units/mL of rinsate compared to the control group at 42 d of age (21 dpi). The relative abundance of IL-10, IL-1, TLR-4, and IFNγ mRNA was decreased (P < 0.05) in the antibiotic and synbiotic supplementation groups compared to the control birds at 42 d of age (21 dpi). It can be concluded that synbiotic supplementation decreased SE proliferation in vitro and decreased SE load in the cecal contents and broiler carcass.
Introduction
Salmonellosis is a foodborne illness, caused by the gram-negative enteric bacterium Salmonella and is of major public health importance in developing countries. The primary sources of human Salmonella infections are consumption of contaminated meat or eggs of Salmonella-positive chickens [1] and up to 9% of samples from poultry production can be positive for Salmonella [2]. Chicks acquire Salmonella via vertical transmission from parents and horizontal transfer from environmental facilities [3]. Most of the initial infection takes place early during post-hatch, although, Salmonella infection can occur during any part of the production cycle [4]. Salmonella control in poultry flocks is difficult since cleaning and disinfection fail to eliminate Salmonella in poultry [5]. Though HACCP (Hazard Analysis Critical Control Program) has reduced Salmonella contamination of chicken carcasses [6], recent multistate outbreak of multidrug-resistant- S. Heidelberg highlights the need to develop effective control measures to reduce Salmonella in the poultry industry [7].
In healthy humans, the Salmonella infectious dose is 106 to 108 [8], while chickens infected with Salmonella are persistent carriers [9]. Salmonella survives in the chicken intestine by inducing T regulatory cells (Tregs) [9]. Induced Tregs secrete Interleukin-10 (IL-10) and suppress the host immune responses, which could help Salmonella to escape host immune responses [9]. Virginiamycin is a commonly used antibiotic in poultry production and has been shown to affect Salmonella prevalence and abundance in poultry intestine [10].
S. Enteritidis is the predominant Salmonella serovar in human cases related to poultry contamination in US [11]. Numerous on-farm control strategies have been evaluated for control Salmonella shedding in poultry, including vaccination [12]. However, these control strategies have limited success in controlling Salmonella contamination in chicken [13], and hence, it is necessary to identify alternative on-farm strategies to control Salmonella infection in broilers.
Currently, the poultry industry applies probiotics and prebiotics to control issues associated with gut health. Probiotics are live fed microbial supplements and can maintain the microbial balance between beneficial and pathogenic bacteria in the gut [14] by producing antibacterial substances or through competitive exclusion by competing for attachment sites in the gut [15]. Prebiotics are non-digestible carbohydrates that act as a substrate for Bifidobacteria and lactic acid bacteria (LAB) in the colon [16]. Fructo-oligosaccharides, galacto-oligosaccharides, and mannan-oligoasacchardes are commonly applied as prebiotics in poultry production [17]. Prebiotics protect against Salmonella colonization by competing for the binding sites [18] and increasing the short-chain fatty acids concentrations in the intestine [19].
Intestinal colonization load of Salmonella play a role in carcasses contamination at slaughter, hence, reducing Salmonella colonization in chickens may potentially reduce salmonellosis incidence in humans [20]. Though extensive studies have been conducted to evaluate the effects of several newly developed and commercially available probiotics on intestinal colonization of Salmonella in birds, very little research has been undertaken to identify if the reduced intestinal Salmonella colonization translates into decreased carcass contamination. The objective of this study is to identify the effects of four probiotic strains of commercially available synbiotic compound (PoultryStar® ME at 0.5 g/kg feed containing L. reuteri, E. faecium, B. animalis, P. acidilactici and a prebiotic Fructooligosaccharide) on Salmonella proliferation and to identify whether in vivo synbiotic supplementation can decrease the Salmonella load in the chicken intestine and bacterial load on carcass.
Materials and methods
All animal protocols were approved by the Institutional Animal Care and Use Committee at the University of Georgia.
in vitro study
Cell-free probiotic supernatants on S. Enteritidis in vitro proliferation
Single isolated colonies of Lactobacillus reuteri, Enterococcus faecium, Bifidobacterium animalis, and Pediococcus acidilactici probiotic strains were inoculated into 50 mL of MRS (DeMan-Rogosa-Sharpe; Sigma Aldrich, St Louis, MO, USA) broth and incubated overnight at 37° C. Once the overnight probiotic cultures reached an optical density between 0.9–1.2 at 600 nm wavelength (O.D 600), cultures were centrifuged at 4,500 X g for 10 min and the supernatant was collected. The supernatant was filter-sterilized using 0.22μm filter (EMD Millipore, MA, USA) to collect cell-free supernatant. A primary isolate of S. Enteritidis [9] was inoculated into 15 mL of Tryptic Soy broth and incubated at 37° C for 12 h.
A volume of 10 μl of S. Enteritidis overnight culture (O.D 600 = 0.1) was incubated with 0:1, 1:1, 5:1, or 10:1 supernatant: pathogen dilutions in triplicates (n = 3) in 96-well flat-bottom plate. The total incubated volume was adjusted to 110 μl using MRS broth. The 96-well plates were incubated at 37°C for 24 h. After incubation, the absorbance was measured at 600nm and the effect of probiotic culture supernatant inhibition on Salmonella proliferation was reported as optical density (OD) values. This assay was conducted in triplicates in three independent experiments (n = 3).
in vivo study
Birds and S. Enteritidis infection
A total of 240 Cobb-500 broiler chicks were randomly allotted to one of three treatment groups, control (basal diet; corn-soybean meal diet), antibiotic (Virginiamycin at 20 mg/Kg feed; Stafac®20, Phibro Animal Health, Teaneck, NJ), and synbiotic (Poultrystar® MEUS at 0.5g/Kg feed; Biomin America Inc., Overland Park, KS) from day of hatch. Experimental basal feed was a corn-soybean meal diet (Table 1). The synbiotic (PoultryStar® ME, BIOMIN America, Inc.) contained four live strains isolated from adult chickens (L. reuteri, E. faecium, B. animalis, and P. acidilactici) with the prebiotic, Fructooligosaccharide. Each treatment was replicated in eight floor pens with 10 chicks per pen (n = 8). Chickens had ad libitum access to water and feed during the entire experimental period. Bodyweight and feed consumption were measured at weekly intervals, and body weight gain and feed conversion ratio (FCR) were calculated. At 21 d of age, all birds in experimental groups were inoculated orally with 250 μl of 1 X 109 colony forming units (CFU) of nalidixic acid-resistant S. Enteritidis, the same strain used for in vitro study. The nalidixic acid-resistant variants were used to assess the recovery of Salmonella from carcass rinses.
Table 1. Primer sequences and PCR conditions for housekeeping genes under study1.
| Gene name | Primer sequence1 | Annealing temperature |
|---|---|---|
| IL-10 |
F: 5’- CAGACCAGCACCAGTCATCA-3’ R: 5’- CGAACGTCTCCTTGATCTGC-3’ |
57.5ºC |
| IL-1β |
F: 5’- CTACACCCGCTCACAGTCCT-3’ R: 5’- TCACTTTCTGGCTGGAGGAG-3’ |
57.5ºC |
| TLR-4 |
F: 5’- ACCTACCCATCGGACACTTG-3’ R: 5’- TGCCTGAGAGGTCAGGTT-3’ |
60.0ºC |
| IFNγ |
F: 5’- CTGATGGCGTGAAGAAGGTG -3’ R: 5’- CTCCTCTGAGACTGGCTCCTTT -3’ |
57.4ºC |
| β-actin |
F: 5’- ACCGGACTGTTACCAACACC-3’ R: 5’-GACTGCTGCTGACACCTTCA-3’ |
57.0ºC |
1Primer sets F, forward; R, reverse.
Effect of synbiotic supplementation on cecal S. Enteritidis load post-Salmonella infection in broiler birds
On 3, 7, 14, and 21 d post-infection, cecal contents were collected from one bird per pen (eight birds per treatment) and analyzed for S. Enteritidis load by real-time PCR. Bacterial genomic DNA was isolated as described earlier by [21] with some modifications. Cecal contents (0.2g) were washed two times with 1X PBS. The cell pellet was resuspended in EDTA and treated with 20 mg/ml lysozyme for 30 min at 37°C, followed by treatment with lysis buffer containing 20% SDS and 0.1 mg/ml proteinase K (Sigma Aldrich, St Louis, MO) for 5 min at 80°C. The samples were incubated with 5μL of RNase at 37°C for 30 min. The cell lysate was incubated with 6M sodium chloride on ice for 10 min. The supernatant was collected after centrifugation at 400 X g for 10 min. The DNA in the supernatant was precipitated with isopropanol and washed once in ice-cold ethanol. The DNA pellet was resuspended in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0) and stored at -20°C until further use.
The DNA extracted from all the treatment groups was analyzed for S. Enteritidis load by real-time PCR using S. Enteritidis specific primers F-GCAGCGGTTACTATTGCAGC and R-CTGTGACAGGGACATTTAGCG [22]. The threshold cycle (Cq) values were determined by CFX software (Bio-Rad, Hercules, CA) when the fluorescence rises exponentially 2-fold above background. The copy numbers of S. Enteritidis specific was expressed in log units as described previously [23].
Effect of synbiotic supplementation on S. Enteritidis carcass rinsate load post-Salmonella infection in broiler birds
At 21 d post-infection, chickens were removed from feed, but not water, for 8 h prior to slaughter, and one chicken from each pen from each of the eight replications (n = 8) was randomly selected for slaughtering. The chickens were processed at the USDA-ARS, Athens processing facility following standard processing protocols for stunning and bleeding. Three counter-current scald tanks were used for scalding. The scalding tanks temperature was 128°, 130°, and 130° Fahrenheit. Chickens were immersed in each scalding tank for 90 seconds, followed by defeathering for 30 seconds. Chickens from antibiotic treatment group were processed first followed by that in the synbiotic treatment group and control treatment group. Scald tank water was changed, all equipment was cleaned and sanitized before, chickens from each treatment group were processed. After removal of the feathers, head, and hocks, the excess fluid was drained from the carcass, which was then transferred to a sterile bag (Cryovac, Charlotte, NC). A 400 milliliter (ml) volume of sterile buffered peptone water (BPW) (Difco Laboratories) was added into each bag. The sterile carcass bags were shaken in a rocking motion for two minutes. One mL of chicken carcass rinsate from each sample was transferred to a polypropylene culture tube containing 9 mL BPW for Salmonella enumeration. A 3-tube most probable number (MPN) technique was used to enumerate the Salmonella load in the carcass as described earlier [24]. Briefly, one mL of the chicken carcass rinsate was mixed with 9 mL BPW and incubated at 37°C for 24 h. At 24 h incubation, 0.1 mL of pre-enriched sample was inoculated into 9.9 mL of Rappaport-Vassiliadis broth (Sigma Aldrich, St Louis, MO) for selective enrichment of Salmonella and incubated at 42°C for 24 h. 10 μL of the enrichment culture was streaked on Xylose- Lysine- Tergitol 4 (XLT4) agar selective media (Hardy Diagnostics) and incubated at 37 °C for 24 h. The number of CFU Salmonella, recovered from each rinse sample was determined by manual counting of colonies. Salmonella suspected positive black colonies were resuspended in the PBS for confirmation through real-time PCR using S. Enteritidis specific primers. Rinse fluid CFU from each rinse sample was converted to Log10 CFU/mL of recovered rinse fluid.
Synbiotic supplementation on IL-10, IL-1, TLR-4, and IFNγ mRNA amounts in the cecal tonsils
At 3, 7, 14, and 21 d post-infection, cecal tonsils were collected and analyzed for IL-10 IL-1, Toll-like receptor (TLR-4), and Interferon-γ (IFNγ) mRNA content by real-time PCR. On 3, 7, 14, and 21 d post-infection, one bird per pen from each of the eight replications were randomly chosen for sample collection (n = 8). Total RNA was collected from cecal tonsils and reverse transcribed into cDNA [25]. mRNA content for IL-10, IL-1β, TLR4, and IFNγ were analyzed by real-time PCR (CFX96 Touch Real Time System, BioRad) using SyBr green after normalizing for β-actin mRNA [26]. Primer sequences are provided in Table 1. Fold change from the reference was calculated [27] as ES (Ct Sample)/ER (Ct Reference), where ES and ER are the sample and reference PCR amplification efficiencies as determined by LinRegPCR program [28], and Ct is the threshold cycle. Ct was determined by the CFX software (Biorad, Hercules, CA) when the fluorescence rises exponentially two-fold times above background. The reference group was the control diet group.
Statistical analysis
A one-way ANOVA was used to determine the effect probiotic culture supernatant on Salmonella growth and the effects of antibiotic and synbiotic supplementation on dependent variables (JMP, SAS Institute Inc., Cary, NC). The averages of plate counts were converted to log CFU/ml and were analyzed using one- way ANOVA. When the main effects were significant (P < 0.05), differences between means were analyzed by Tukey's least-square means comparison.
Results
In vitro experiment
Effect of cell-free probiotic supernatants on S. Enteritidis in vitro proliferation
All the four probiotics culture supernatants inhibit the growth of S. Enteritidis at 1:1 supernatant: pathogen dilution compared to the 0:1 dilution group. The proliferation of S. Enteritidis at 1:1 dilution decreased by 99.5, 95.8, 93.1, and 95.7% for L. reuteri, E. faecium, B. animalis, and P. acidilactici, respectively, when compared to the 0:1 dilution group. The inhibition at 5:1 dilution was 96.8, 87.4, 87.4, and 92.9% for L. reuteri, E. faecium, B. animalis, and P. acidilactici, respectively, when compared to the 0:1 dilution group. While the inhibition at 10:1 dilution was 96.3, 93.0, 88.8, and 97.1% for L. reuteri, E. faecium, B. animalis, and P. acidilactici, respectively, when compared to the 0:1 dilution group (Fig 1).
Fig 1. Effect of cell- free probiotic supernatants on S. Enteritidis in vitro proliferation.
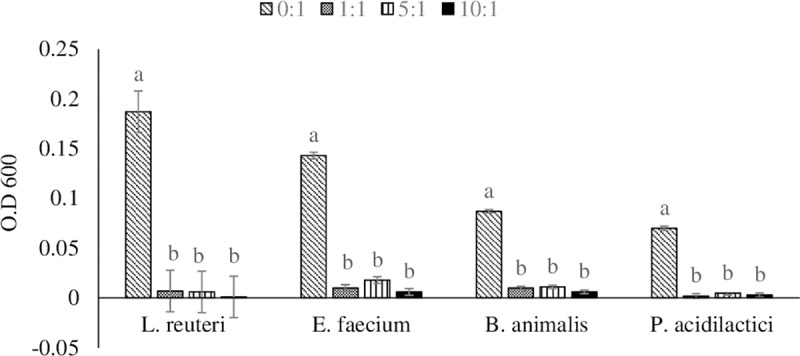
Overnight culture of single isolated colonies of Lactobacillus reuteri, Enterococcus faecium, Bifidobacterium animalis, and Pediococcus acidilactici probiotic strains were centrifuged at 4,500 X g for 10 min to collect supernatants. The supernatant was filtered using a 0.22μm filter to collect cell-free supernatant. 10 μl of S. Enteritidis overnight culture was incubated with 0:1, 10:1, 5:1, or 1:1 cell-free supernatant: pathogen dilutions. The absorbance was measured at 600nm at 24 hours. n = 3. Values ± SEM of pooled samples of 3 wells/treatment. Bars (± SEM) with no common superscript (a, b) differ significantly (P < 0.05).
In vivo experiment
Effect of synbiotic supplementation on production parameters in post-Salmonella challenge
Synbiotic supplementation had no significant effects on body weight and feed consumption at 21 d and 42 d of age (P > 0.05) compared to control groups (Table 2). At 21 d and 42 d of age antibiotic supplementation had significantly increased BW and feed consumption (P < 0.01) compared to the control groups. Antibiotic and synbiotic supplementation increased BW by 230 and 170 g and feed consumption by 270 and 200 g compared to the control S. Enteritidis challenge group at 42 d of age, respectively. Synbiotic supplementation had no significant effects on feed conversion ratio at 21 (P = 0.12) and 42 d of age (P = 0.16) compared to control groups in response to S. Enteritidis challenge.
Table 2. Effect of synbiotic supplementation on production parameters post-Salmonella infection in broiler birds.
| 0-21d | 0-42d | |
|---|---|---|
| Body weight (kg) | ||
| Control | 0.79b | 2.65b |
| Virginiamycin | 0.89a | 2.89a |
| Synbiotic | 0.82b | 2.82ab |
| SEM | 0.01 | 0.05 |
| P value | P < 0.01 | P = 0.01 |
| Feed consumption (kg) | ||
| Control | 1.08b | 4.47b |
| Virginiamycin | 1.17a | 4.74a |
| Synbiotic | 1.10b | 4.67ab |
| SEM | 0.02 | 0.06 |
| P value | 0.01 | 0.01 |
| Feed conversion ratio | ||
| Control | 1.37 | 1.69 |
| Virginiamycin | 1.32 | 1.64 |
| Synbiotic | 1.33 | 1.66 |
| SEM | 0.02 | 0.02 |
| P value | 0.12 | 0.16 |
Birds were fed either basal diet (Control) or supplemented with 20 mg/Kg feed Virginiamycin (antibiotic) or 0.05% synbiotic product (Poultrystar® ME; Biomin America Inc) day-of-hatch through 42d of age. At 21 d of age, birds were challenged with 1 X 109 CFU of Salmonella enterica ser. Enteritidis. Means with no common superscript (a, b, ab) within a column differ significantly (P < 0.05). n = 8.
Effect of synbiotic supplementation on S. Enteritidis load in the cecal content post-Salmonella challenge
Antibiotic and synbiotic supplementation had significant effects on S. Enteritidis load in the cecal content at 3 (P < 0.01), 7 (P < 0.01), 14 (P < 0.01), and 21 (P = 0.01) d post-infection (Fig 2). Antibiotic and synbiotic supplementation decreased S. Enteritidis load in the cecal content at by 0.90 and 0.85 log units, respectively compared to the control group at 21 d post-Salmonella infection.
Fig 2. Effect of synbiotic supplementation on S. Enteritidis load in the cecal content post-Salmonella infection in broilers.

Birds were fed either basal diet (Control) or supplemented with 20 mg/Kg feed Virginiamycin (antibiotic) or 0.05% synbiotic product from day-of-hatch through 42d of age. At 21 d of age, birds were challenged with 1 X 109 CFU of S. Enteritidis. At 3, 7, 14, and 21 d post-infection, cecal content were analyzed for S. Enteritidis load by real-time PCR collected and expressed as log values. Bars (± SEM) with no common superscript (a, b, c) differ significantly (P < 0.05). n = 8.
Effect of synbiotic supplementation on carcass S. Enteritidis load post- Salmonella challenge
Antibiotic and synbiotic supplementation had significant effects on chilled carcass S. Enteritidis load at 21 d (P = 0.02) post-infection (Fig 3). Antibiotic and synbiotic supplementation decreased carcass S. Enteritidis rinsate load at by 1.4 and 1.5 log units, respectively compared to the control group at 21 d post-Salmonella infection.
Fig 3. Effect of synbiotic supplementation on chilled carcass S. Enteritidis rinsate load post-Salmonella infection in broiler birds.

Birds were fed either basal diet (Control) or supplemented with 20 mg/Kg feed Virginiamycin (antibiotic) or 0.05% synbiotic product from day-of-hatch through 42d of age. At 21 d of age, birds were challenged with 1 X 109 CFU of S. Enteritidis. At 21 d post-infection, the carcass S. Enteritidis most probable number (MPN) was analyzed by plating on XLT agar and expressed as log MPN/ml. Bars (± SEM) with no common superscript (a, b) differ significantly (P < 0.05). n = 8.
Effect of synbiotic supplementation on cecal tonsil gene expression post-Salmonella challenge
Antibiotic and synbiotic supplementation reduced cecal tonsil IL-10 mRNA content at 21 (P < 0.01) d post-infection (Fig 4A).
Fig 4. Effect of synbiotic supplementation on cecal tonsil mRNA content post-Salmonella infection in broilers.

Birds were fed either basal diet (Control) or supplemented with 20 mg/Kg feed Virginiamycin (antibiotic) or 0.05% synbiotic product from day-of-hatch through 42d of age. At 21 d of age, birds were challenged with 1 X 109 CFU of S. Enteritidis. At 3, 7, 14 and 21 d post-infection (p.i.), relative IL-10 (Fig 4A), IL-1 (Fig 4B), TLR-4 (Fig 4C), and IFN γ (Fig 4D) mRNA content in cecal tonsils were analyzed from 1 bird/pen; 8 pens/diet after correcting for β-actin mRNA and normalizing to the mRNA content of the control group. Bars (± SEM) with no common superscript (a, b) differ significantly. n = 8.
Antibiotic and synbiotic supplementation had significant effects on cecal tonsil IL-1 mRNA content at 3 (P = 0.01), 7 (P = 0.01), and 21 (P < 0.01) d post-infection (Fig 4B). At 21 d post-infection, birds in the antibiotic and synbiotic supplementation group had 60, and 50% decreased IL-1 mRNA compared to the control group, respectively.
Antibiotic and synbiotic supplementation had significant effects on cecal tonsil TLR-4 mRNA content at 3 (P < 0.01), 7 (P < 0.01), 14 (P <0.01) and 21 (P < 0.01) d post-infection (Fig 4C). At 21 d post-infection, birds in the antibiotic and synbiotic supplementation group had 70, and 56% decreased TLR-4 mRNA compared to the control group, respectively.
Antibiotic and synbiotic supplementation had significant effects on cecal tonsil IFNγ mRNA content at 3 (P < 0.01), 7 (P < 0.01), 14 (P < 0.01) and 21 (P < 0.01) d post-infection (Fig 4D). At 21 d post-infection, birds in the antibiotic and synbiotic supplementation group had by 0.61, and 0.65% decreased IFNγ mRNA compared to the control group, respectively.
Discussion
This study identified that all four probiotics strains supernatants had decreased in vitro proliferation of S. Enteritidis separately, and synbiotic supplementation from the day of hatch decreased the Salmonella load in the chicken cecal contents and decreased carcass contamination in broiler birds.
Supernatants from probiotic strains L. reuteri, P. acidilactici, B. animalis and E.faecium decreased the proliferation of S. Enteritidis in vitro. Our results are consistent with previous studies conducted with Lactic acid bacteria (LAB) and Enterococcus bacteria. LAB produce antimicrobial substances such as organic acids, bacteriocins [29], and peptidoglycan hydrolases [30] which can be expected to decrease Salmonella proliferation. In addition, LAB have been shown to have a competitive advantage over pathogenic microorganism in the gut because LAB can tolerate low intestinal pH and bile [31]. L. reuteri exhibits inhibitory effects against both S. Enteritidis and S. Typhimurium [32]. Similarly, E. faecium and P. acidilactici produce enterocins and pediocins respectively, which have been shown to inhibit the growth of gram-positive and gram-negative pathogenic bacteria [33]. B. animalis produces lactic acid and other bactericidal substances to inhibit the growth of Salmonella [34]. Supernatants from all four probiotic strains efficiently inhibited the proliferation of S. Enteritidis, suggesting that in vivo supplementation of these probiotic strains to the chickens might be beneficial during a Salmonella infection.
In this study, we demonstrated that in vivo synbiotic supplementation from the day of hatch decreased the S. Enteritidis load in the cecal tonsils and decreased carcass contamination in broiler birds. Our laboratory has previously shown that L. reuteri, P. acidilactici, B. animalis, and E.faecium can successfully colonize the chicken intestine [23]. The consistent effect of the probiotics in decreasing the proliferation of S. Enteritidis both in vitro and in vivo suggest that probiotics will be a major tool in combating Salmonella load in birds.
In this study, chickens fed synbiotics and antibiotics had decreased Salmonella load in both cecal content and carcass rinsate. In pigs, synbiotics decrease Salmonella loads in the intestine and decrease Salmonella contamination of carcasses which suggest that decreasing intestinal Salmonella load in the intestine would be the ideal approach to decrease carcass load [35]. In chickens, Salmonella can colonize the gut efficiently and thereby can be shed in the feces for an extended period without showing symptoms. Salmonella-contaminated feces play a major role in carcass contamination and horizontal transmission in chicken [36].; Probiotics which act against gut microbes through competitive exclusion treatments has been shown to reduce Salmonella flock prevalence by up to 70–85% [37]. In our study, synbiotic supplementation decreased the Salmonella load in both cecal content and carcass rinsate suggesting that synbiotics not only efficiently colonized the intestine but also secreted antibacterial substances in the gut lumen to decrease the S. Enteritidis load in the carcass.
TLR-4 is a pattern recognition receptor and recognizes the lipopolysaccharide of gram-negative bacteria and is an indicator of immune stimulation following Salmonella infection [38]. Chickens in the antibiotic and synbiotic supplemented groups had decreased cecal tonsil TLR-4 mRNA content compared to the control groups post-Salmonella infection throughout the study.
Synbiotics may protect against Salmonella infection through different mechanisms, including modulation of cytokine responses. Probiotic bacteria such as E. faecium and L reuteri exert immunomodulatory activities by altering the host cytokine expression profiles [39, 40]. Salmonella can also stimulate the host immune cells to modify host cytokines and chemokines [40]. IFNγ is an inflammatory cytokine that acts to improve the host defense against intracellular pathogen like Salmonella. Chickens in the control group infected with Salmonella had higher IFNγ compared to the Salmonella-infected birds fed antibiotic and synbiotic all-time points of this study. Considering that supplementation of synbiotic and antibiotic in birds challenged with Salmonella decreased Salmonella load in the cecal content, the decreased Salmonella load likely contributed to the decreased IFNγ mRNA in the cecal tonsil. Salmonella infection increase IL-10 mRNA content in the cecal tonsils [41]. IL-10 is a regulatory cytokine produced by T regulatory cells, and upregulation of IL-10 by Salmonella is a pathogen defense mechanism to create a persistent infection of chickens [9]. Synbiotic and antibiotic supplementation reduced Salmonella load in the cecal content of Salmonella infected birds and the reduced Salmonella load likely contributed to the decreased IL-10 mRNA in the cecal tonsil.
In conclusion, our study confirmed that the in vivo synbiotic supplementation improves the body weight and feed intake and reduce the colonization of Salmonella in the cecal content of broiler chickens. Therefore, administration of synbiotics can reduce or replace the use of antibiotics in poultry production and reduce the incidence of Salmonella load in the carcass.
Acknowledgments
The authors acknowledge Gabriel Oluwamuyiwa Akerele, Jarred Oxford and Keila Yanis Acevedo Villanueva for their help in animal husbandry and sample collection.
Data Availability
All relevant data are within the manuscript.
Funding Statement
This work was funded partially supported by BIOMIN fund awarded to RS and Hatch grant and USDA-NIFA grant 2017-05035 awarded to RKS. The funders had no role in study design, data role of authors in this manuscript is mentioned in authors contribution section collection and analysis, decision to publish, or preparation of the manuscript. The specific contribution of authors in this manuscript is mentioned in authors contribution section.
References
- 1.Kao JY, Zhang M, Miller MJ, Mills JC, Wang B, Liu M, et al. Helicobacter pylori immune escape is mediated by dendritic cell-induced Treg skewing and Th17 suppression in mice. Gastroenterology. 2010;138(3):1046–54. Epub 2009/11/26. S0016-5085(09)02060-5 [pii] 10.1053/j.gastro.2009.11.043 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bailey JS, Stern NJ, Fedorka-Cray P, Craven SE, Cox NA, Cosby DE, et al. Sources and movement of Salmonella through integrated poultry operations: a multistate epidemiological investigation. J Food Prot. 2001;64(11):1690–7. Epub 2001/12/01. 10.4315/0362-028x-64.11.1690 . [DOI] [PubMed] [Google Scholar]
- 3.Rothrock MJ Jr., Ingram KD, Gamble J, Guard J, Cicconi-Hogan KM, Hinton A Jr., et al. The characterization of Salmonella enterica serotypes isolated from the scalder tank water of a commercial poultry processing plant: Recovery of a multidrug-resistant Heidelberg strain. Poult Sci. 2015;94(3):467–72. 10.3382/ps/peu060 . [DOI] [PubMed] [Google Scholar]
- 4.Byrd JA, DeLoach JR, Corrier DE, Nisbet DJ, Stanker LH. Evaluation of salmonella serotype distributions from commercial broiler hatcheries and grower houses. Avian Dis. 1999;43(1):39–47. 10.2307/1592760 WOS:000081987800006. [DOI] [PubMed] [Google Scholar]
- 5.Davies RH, Wray C. Studies of contamination of three broiler breeder houses with Salmonella enteritidis before and after cleansing and disinfection. Avian Dis. 1996;40(3):626–33. Epub 1996/07/01. . [PubMed] [Google Scholar]
- 6.White PL, Naugle AL, Jackson CR, Fedorka-Cray PJ, Rose BE, Pritchard KM, et al. Salmonella Enteritidis in meat, poultry, and pasteurized egg products regulated by the US food safety and inspection service, 1998 through 2003. Journal of Food Protection. 2007;70(3):582–91. 10.4315/0362-028x-70.3.582 WOS:000244736600007. [DOI] [PubMed] [Google Scholar]
- 7.Vugia DJ, Samuel M, Farley MM, Marcus R, Shiferaw B, Shallow S, et al. Invasive Salmonella infections in the United States, FoodNet, 1996–1999: incidence, serotype distribution, and outcome. Clin Infect Dis. 2004;38 Suppl 3:S149–56. 10.1086/381581 . [DOI] [PubMed] [Google Scholar]
- 8.TODD ECD, GREIG JD, BARTLESON CA, MICHAELS BS. Outbreaks Where Food Workers Have Been Implicated in the Spread of Foodborne Disease. Part 4. Infective Doses and Pathogen Carriage. Journal of Food Protection. 2008;71(11):2339–73. 10.4315/0362-028x-71.11.2339 [DOI] [PubMed] [Google Scholar]
- 9.Shanmugasundaram R, Kogut MH, Arsenault RJ, Swaggerty CL, Cole K, Reddish JM, et al. Effect of Salmonella infection on cecal tonsil regulatory T cell properties in chickens. Poult Sci. 2015;94(8):1828–35. 10.3382/ps/pev161 . [DOI] [PubMed] [Google Scholar]
- 10.Lu J, Hofacre C, Smith F, Lee M. Effects of feed additives on the development on the ileal bacterial community of the broiler chicken. Animal. 2008;2(5):669–76. 10.1017/S1751731108001894 [DOI] [PubMed] [Google Scholar]
- 11.Morningstar-Shaw B, Mackie T, Barker D, Palmer E. Salmonella Serotypes Isolated from Animals and Related Sources. Ames: United States Department of Agriculture (USDA), National Veterinary Services Laboratories; 2016. [Google Scholar]
- 12.Varmuzova K, Faldynova M, Elsheimer-Matulova M, Sebkova A, Polansky O, Havlickova H, et al. Immune protection of chickens conferred by a vaccine consisting of attenuated strains of Salmonella Enteritidis, Typhimurium and Infantis. Veterinary research. 2016;47(1):94 10.1186/s13567-016-0371-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Koutsoumanis K, Allende A, Alvarez-Ordonez A, Bolton D, Bover-Cid S, Chemaly M, et al. Salmonella control in poultry flocks and its public health impact. Efsa J. 2019;17(2). ARTN 5596 10.2903/j.efsa.2019.5596 WOS:000459860800006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fuller. Probiotics in man and animals. Journal of Applied Microbiology. 1989;66(5):365–78. [PubMed] [Google Scholar]
- 15.Tuomola EM, Ouwehand AC, Salminen SJ. The effect of probiotic bacteria on the adhesion of pathogens to human intestinal mucus. FEMS Immunol Med Microbiol. 1999;26(2):137–42. Epub 1999/10/28. 10.1111/j.1574-695X.1999.tb01381.x . [DOI] [PubMed] [Google Scholar]
- 16.Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. 1995;125(6):1401–12. 10.1093/jn/125.6.1401 . [DOI] [PubMed] [Google Scholar]
- 17.Ricke SC. Potential of fructooligosaccharide prebiotics in alternative and nonconventional poultry production systems. Poult Sci. 2015;94(6):1411–8. Epub 2015/02/27. 10.3382/ps/pev049 . [DOI] [PubMed] [Google Scholar]
- 18.Durant JA, Corrier DE, Ricke SC. Short-chain volatile fatty acids modulate the expression of the hilA and invF genes of Salmonella Typhimurium. J Food Protect. 2000;63(5):573–8. 10.4315/0362-028x-63.5.573 WOS:000086925700003. [DOI] [PubMed] [Google Scholar]
- 19.Ricke SC. Potential of fructooligosaccharide prebiotics in alternative and nonconventional poultry production systems. Poultry Science. 2015;94(6):1411–8. 10.3382/ps/pev049 WOS:000355189700035. [DOI] [PubMed] [Google Scholar]
- 20.Khaksefidi A, Rahimi S. Effect of Probiotic inclusion in the diet of broiler chickens on performance, feed efficiency and carcass quality. Asian Austral J Anim. 2005;18(8):1153–6. 10.5713/ajas.2005.1153 WOS:000229447500016. [DOI] [Google Scholar]
- 21.Zhu XY, Zhong T, Pandya Y, Joerger RD. 16S rRNA-based analysis of microbiota from the cecum of broiler chickens. Appl Environ Microbiol. 2002;68(1):124–37. Epub 2002/01/05. 10.1128/AEM.68.1.124-137.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.De Medici D, Croci L, Delibato E, Di Pasquale S, Filetici E, Toti L. Evaluation of DNA Extraction Methods for Use in Combination with SYBR Green I Real-Time PCR To Detect Salmonella enterica Serotype Enteritidis in Poultry. Appl Environ Microb. 2003;69(6):3456–61. 10.1128/aem.69.6.3456-3461.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shanmugasundaram R, Mortada MH, Murugesan RG, Selvaraj RK. In-vitro Characterization and Analysis of Probiotic Species in the Chicken Intestine by Real-time Polymerase Chain Reaction. Poult Sci. 2019:In Press. [DOI] [PubMed] [Google Scholar]
- 24.Santos FBO, Li X, Payne JB, Sheldon BW. Estimation of Most Probable Number Salmonella Populations on Commercial North Carolina Turkey Farms. The Journal of Applied Poultry Research. 2005;14(4):700–8. 10.1093/japr/14.4.700 [DOI] [Google Scholar]
- 25.Selvaraj RK, Klasing KC. Lutein and Eicosapentaenoic Acid Interact to Modify iNOS mRNA Levels through the PPAR{gamma}/RXR Pathway in Chickens and HD11 Cell Lines. J Nutr. 2006;136(6):1610–6. 10.1093/jn/136.6.1610 [DOI] [PubMed] [Google Scholar]
- 26.Shanmugasundaram R, Selvaraj RK. In vitro human TGF-beta treatment converts CD4(+)CD25(-) T cells into induced T regulatory like cells. Vet Immunol Immunopathol. 2010;137(1–2):161–5. Epub 2010/08/06. 10.1016/j.vetimm.2010.04.017 . [DOI] [PubMed] [Google Scholar]
- 27.Selvaraj RK, Shanmugasundaram R, Klasing KC. Effects of dietary lutein and PUFA on PPAR and RXR isomer expression in chickens during an inflammatory response. Comp Biochem Physiol a: Mol Integ Physiol. 2010;157(3):198–203. [DOI] [PubMed] [Google Scholar]
- 28.Ramakers C, Ruijter JM, Deprez RH, Moorman AF. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett. 2003;339(1):62–6. Epub 2003/03/06. S0304394002014234 [pii]. 10.1016/s0304-3940(02)01423-4 . [DOI] [PubMed] [Google Scholar]
- 29.Corcionivoschi N, Drinceanu D, Stef L, Luca I, Julean C, Mingyart O. Probiotics—identification and ways of action. Innovative Romanian Food Biotechnology. 2010;6:1–11. [Google Scholar]
- 30.Turner MS, Hafner LM, Walsh T, Giffard PM. Identification, characterisation and specificity of a cell wall lytic enzyme from Lactobacillus fermentum BR11. FEMS Microbiol Lett. 2004;238(1):9–15. Epub 2004/09/01. 10.1016/j.femsle.2004.07.008 . [DOI] [PubMed] [Google Scholar]
- 31.Ghadban GS. Probiotics in broiler production—a review. Arch Geflugelkd. 2002;66(2):49–58. WOS:000175786700001. [Google Scholar]
- 32.Brisbin JT, Parvizi P, Sharif S. Differential cytokine expression in T-cell subsets of chicken caecal tonsils co-cultured with three species of Lactobacillus. Benef Microbes. 2012;3(3):205–10. 10.3920/BM2012.0014 . [DOI] [PubMed] [Google Scholar]
- 33.Jamuna M, Jeevaratnam K. Isolation and partial characterization of bacteriocins from Pediococcus species. Appl Microbiol Biotechnol. 2004;65(4):433–9. 10.1007/s00253-004-1576-8 . [DOI] [PubMed] [Google Scholar]
- 34.Bielecka M, Biedrzycka E, Biedrzycka E, Smoragiewicz W, Smieszek M. Interaction of Bifidobacterium and Salmonella during associated growth. Int J Food Microbiol. 1998;45(2):151–5. Epub 1999/01/30. 10.1016/s0168-1605(98)00150-0 . [DOI] [PubMed] [Google Scholar]
- 35.Pesciaroli M, Cucco L, De Luca S, Massacci FR, Maresca C, Medici L, et al. Association between pigs with high caecal Salmonella loads and carcass contamination. International Journal of Food Microbiology. 2017;242:82–6. 10.1016/j.ijfoodmicro.2016.11.021 WOS:000392045200011. [DOI] [PubMed] [Google Scholar]
- 36.Rose N, Beaudeau F, Drouin P, Toux JY, Rose V, Colin P. Risk factors for Salmonella enterica subsp. enterica contamination in French broiler-chicken flocks at the end of the rearing period. Prev Vet Med. 1999;39(4):265–77. Epub 1999/05/18. . [DOI] [PubMed] [Google Scholar]
- 37.FAO/WHO. Salmonella and Campylobacter in chicken meat. 2009 Microbiological Risk Assessment Series No. 19. Rome 56 pp.
- 38.Kogut MH, He HQ, Kaiser P. Lipopolysaccharide binding protein/CD14/TLR4-dependent recognition of Salmonella LPS induces the functional activation of chicken heterophils and up-regulation of pro-inflammatory cytokine and chemokine gene expression in these cells. Animal Biotechnology. 2005;16(2):165–81. 10.1080/10495390500264896 WOS:000233299700007. [DOI] [PubMed] [Google Scholar]
- 39.Kim YG, Ohta T, Takahashi T, Kushiro A, Nomoto K, Yokokura T, et al. Probiotic Lactobacillus casei activates innate immunity via NF-kappa B and p38 MAP kinase signaling pathways. Microbes Infect. 2006;8(4):994–1005. 10.1016/j.micinf.2005.10.019 WOS:000237754100005. [DOI] [PubMed] [Google Scholar]
- 40.Haghighi HR, Abdul-Careem MF, Dara RA, Chambers JR, Sharif S. Cytokine gene expression in chicken cecal tonsils following treatment with probiotics and Salmonella infection. Vet Microbiol. 2008;126(1–3):225–33. 10.1016/j.vetmic.2007.06.026 WOS:000251924700024. [DOI] [PubMed] [Google Scholar]
- 41.Arai T, Hiromatsu K, Nishimura H, Kimura Y, Kobayashi N, Ishida H, et al. Effects of in vivo administration of anti-IL-10 monoclonal antibody on the host defence mechanism against murine Salmonella infection. Immunology. 1995;85(3):381–8. Epub 1995/07/01. [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the manuscript.