

Abstract
Betel quid is one of the most widely used psychoactive substances, and is consumed by approximately 10% of the world’s population. In addition to its carcinogenicity, betel quid has also been reported to affect many organs, including the brain, heart, lungs, gastrointestinal tract, and reproductive organs. As betel quid contains several neurotoxic ingredients, we hypothesize that it also possesses ototoxicity and may lead to sensorineural hearing impairment (SNHI). In this study, we investigated the contribution of betel quid consumption to SNHI in a large clinical cohort, and validated the pathogenetic mechanisms in ex vivo tissue explants. We enrolled a total of 2364 volunteers, and determined their audiologic results based on Z-scores converted from their original frequency-specific hearing thresholds. Using generalized linear regression, we identified a positive correlation between betel quid consumption and the Z-scores across different frequencies. Subsequently, we explored the toxicity of arecoline, the main neuroactive component of betel quid, on tissue explants from murine cochleae. Arecoline reduced cell activity in the explant cultures and induced apoptosis in the hair cells, probably through the effects of oxidative stress. These findings have expanded the potential hazards of betel quid to common neurological disorders, and provide insights into preventive strategies against SNHI caused by neurotoxic substances.
Subject terms: Peripheral neuropathies, Experimental models of disease, Risk factors
Introduction
Sensorineural hearing impairment (SNHI) is the most common sensory disorder in humans. According to the report of the World Health Organization, it is estimated that approximately 466 million people worldwide have disabling hearing loss greater than 40 dB, and 34 millions of these are children (http://www.who.int/en/news-room/fact-sheets/detail/deafness-and-hearing-loss). SNHI in children is caused predominantly by genetic mutations1. By contrast, SNHI in adults, similar to other neurodegenerative disorders, involves an interplay between genetic and environmental factors2–4. In addition to noise exposure5, a number of toxic substances have been documented to contribute to SNHI, including ototoxic antibiotics, diuretics, chemotherapeutic agents, and tobacco3,6. In mammals, including humans, the damages result in permanent hearing loss due to the quiescence nature of the sensory epithelium of the cochlea7,8.
Betel quid or areca nut (the primary ingredient in betel quid) is among the most widely used psychoactive substances worldwide, along with tobacco, alcohol, and caffeine. Globally, more than 600 million people are estimated to consume betel quid or areca nut in some form9–12. Consumption is especially prevalent in the Asia-pacific region, including South and Southeast Asia, tropical Pacific, east Africa, as well as in emigrant communities arising therefrom. Both betel quid and areca nut are classified as carcinogenic to human beings by the International Agency for Cancer Research, and have been confirmed as important risk factors for oral, pharyngeal, and esophageal cancers13. In addition, the consumption of betel quid has also been related to various disorders, including myocardial infarction14, cardiac arrhythmia15, hepatotoxicity16, asthma17, diabetes18, metabolic syndrome19, premature delivery20, and infertility21. There is growing evidence that the predominant psychoactive agent in betel quid, arecoline, plays a critical role in the carcinogenesis process22–24.
Interestingly, experiments in cell lines revealed that arecoline of the betel quid causes neurotoxicity by enhancing oxidative stress and suppressing the antioxidant protective systems25. Despite these findings in cell lines, the real contribution of betel quid consumption to neurological disorders (such as SNHI) in humans remained unexplored. Accordingly, we investigated the effects of betel quid consumption on SNHI in a large clinical cohort, and then validated the pathogenetic mechanisms in ex vivo tissue explants.
Materials and Methods
Subject recruitment
From 2007 to 2017, Han Chinese volunteers aged between 40–80 years were prospectively recruited during routine health examination from the Health Management Center of National Taiwan University Hospital to establish a cohort for epidemiological and pathogenetic research on SNHI26–28. The subjects were interviewed by trained assistants with the use of a structured questionnaire detailing demographic data and family history of hearing loss. The questionnaire also served to obtain information on medical history including betel quid consumption, smoking, alcohol consumption, and systemic diseases, such as coronary artery disease, hypertension, diabetes, hyperlipidemia, and chronic hepatitis. Height and body weight were measured after an overnight fast to determine the body mass index (BMI).
All subjects and/or their parents provided informed consent before testing, and all procedures were approved by the Research Ethics Committees of National Taiwan University Hospital. All methods were performed in accordance with the relevant guidelines and regulations.
Audiologic evaluations
All subjects underwent an otoscopic examination by an otologist to inspect ear pathology, and the audiologic results were assessed in terms of air- and bone-conduction thresholds of pure-tones using an audiometer (Grason-Stadler Inc., GSI 10). Air-conduction thresholds were measured at 0.25, 0.5, 1, 2, 4, and 8 kHz; and bone-conduction thresholds were measured at 0.5, 1, 2, and 4 kHz. Subjects with asymmetric sensorineural hearing loss, conductive hearing loss, and a 4-kHz dip on an audiogram were excluded since these audiologic features indicate the presence of other pathologies. Subjects with a history of any of the following were also excluded: external- or middle-ear diseases, hearing loss before the age of 30 years, high environmental noise exposure, exposure to ototoxic medication, brain tumor, vertigo, chronic renal failure under dialysis, head and neck radiation exposure, heavy smoking, alcoholism, or substance abuse.
Frequency-specific thresholds were converted to sex- and age-independent Z-scores, which represent the number of standard deviations by which the hearing threshold differs from the median at a given frequency29. Z-scores for each frequency were denoted as Z250, Z500, Z1000, Z2000, Z4000, and Z8000. Corresponding to the hearing levels at the low-tone, mid-tone, and high-tone frequencies, Z250 and Z500 scores were averaged as Zlow, Z1000 and Z2000 scores as Zmid, and Z4000 and Z8000 as Zhigh26. Only Z-scores of the better hearing ear were used for subsequent analyses.
Generalized linear regression
To investigate the associations between risk factors and Z-scores, we used the generalized linear regression model (GLM). All of the models were adjusted by age and gender. Coefficients (β) of all the risk factors and the 95% confidence intervals (CI) were estimated using the SAS Software version 9.1 (SAS Institute). Statistical significance was set at p < 0.05 based on two-sided estimation.
Preparation of cochlear explants
Ex vivo experiments were carried out by culturing cochlear explants containing the organ of Corti and spiral ganglions harvested from 3–5-day-old C57BL/6 mice (Fig. 1A)26. The explants dissected from the otic capsule were oriented with the hair cell-side up, attached to the Matrigel (Corning) coated on cell culture dishes (Greiner Labortechnik) in the Dulbecco’s Modified Eagle Medium (DMEM) (GIBCO) with 7% fetal bovine serum, and then kept at 37 °C in a humidified incubator under 5% CO2. To explore the toxic effects of arecoline on the cochlear explants, we used gradient concentrations, 200 μM, 800 μM, 2 mM, and 10 mM of arecoline, and examined the tissue morphology, hair cell arrangement, cell activity, and cytotoxicity. Rhodamine-phalloidin (Invitrogen) was used to label the f-actin to examine the cell morphology and the hair bundles of the hair cells. The cells were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). All animal experiments were carried out in accordance with animal welfare guidelines and approved by the Institutional Animal Care and Use Committee (IACUC) of National Taiwan University College of Medicine.
Figure 1.

The effects of arecoline on cochlear explants. (A) The sketch of the cross section of the cochlear duct and the organ of Corti within it. Cochlear explants used in this study contained the region between the dashed lines with the removal of the tectorial membrane (TM). (B) The morphology of cochlear explants under stereo microscopy after 3 days of culture without and with 10 mM arecoline. HB, hair bundles; IHC, inner hair cells; OHC, outer hair cells; TM, tectorial membrane. Bar = 1 mm.
Cell activity and cytotoxicity
The cell activity of cochlear explants was investigated using the alamarBlue assay, which measures the reducing power of living cells quantitatively30. The alamarBlue reagent (10% v/v) was added into the culture medium and incubated for 24 h, and the culture medium was then transferred into 96-well plates. Subsequently, the fluorescent signals were examined by an ELISA reader using a fluorescence excitation wavelength of 570 nm and measuring emission at 585 nm.
The lactate dehydrogenase (LDH) Cytotoxicity Assay Kit (Pierce) was used to determine the amount of LDH released from the damaged cells31. The culture medium of the cochlear explants incubated for 24 h was collected at the indicated time points, and mixed with the LDH reaction mixtures at room temperature for 30 min. The reaction was terminated by adding the stop solution, and subsequently the absorbance was read by an ELISA reader at 490 nm and 690 nm. The absorbance values were processed according to the manufacturer’s instructions to calculate the LDH values.
TUNEL assay
The terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was performed by using the APO-BrdU TUNEL Assay Kit (Invitrogen). It is based on the binding of 5-bromo-2′-deoxyuridine 5′-triphosphate (BrdUTP) by the terminal deoxynucleotidyl transferase (TdT) at the 3′ hydroxyl termini of fragmented DNA32. This was followed by detecting BrdU with an anti-BrdU antibody using standard immunofluorescent techniques. After rinsing with ice cold phosphate-buffered saline (PBS) twice, the cochlear explants were permeabilized with 0.1% Triton X-100 in PBS. After washing, the explants were incubated in the TUNEL reaction mix for 1 h at 37 °C in the dark, and then labeled using the Alexa Fluor 488 dye-conjugated anti-BrdU antibody. Hair cells were stained by the phalloidin–rhodamine probe (Invitrogen) and then counterstained with DAPI. The cochlear explants were mounted and photographed using a confocal microscope (Zeiss LSM 780). The apical, middle, and basal parts of each explant were scanned for comparison.
Cellular oxidative stress
Reactive oxygen species (ROS) within the cochlear explants were examined using the CellROX Green fluorogenic probe (Invitrogen)33. At 30 min before the indicated time points, CellROX Green Reagent was added to the cochlear explants at a final concentration of 5 μM and incubated at 37 °C in a 5% CO2 humidified incubator for 30 min. Then the medium was removed, and the cochlear explants were washed 3 times with ice cold PBS, fixed with 4% paraformaldehyde in PBS, and permeabilized with 0.5% Triton X-100. The hair bundles were labeled with phalloidin and the cells were counterstained with DAPI. The stained cochlear explants were then examined by Zeiss LSM 780 laser scanning confocal microscopy immediately. The apical, middle, and basal parts of each cochlear explant were scanned for comparison. The images were processed by ImageJ (NIH, Bethesda, MD, USA).
Results
Betel quid consumption contributed to SNHI in humans
In total, 2364 subjects, including 1306 men and 1058 women, with a mean age of 54.0 ± 7.7 years, were included in the analyses. The basic characteristics of these subjects are shown in Table 1. Fifty-five (2.3%) of the 2364 subjects consumed betel quid.
Table 1.
Basic characteristics of the 2364 subjects.
| Variables | Value* |
|---|---|
| Age (years) | 54.0 ± 7.7 |
| Male sex | 1306 (55.1) |
| Coronary heart disease | 66 (2.8) |
| Hypertension | 398 (16.8) |
| Diabetes | 136 (5.8) |
| Hyperlipidemia | 123 (5.2) |
| Chronic hepatitis | 129 (5.5) |
| BMI | 24.0 ± 3.2 |
| Smoking | |
| No | 2064 (87.3) |
| Yes | 300 (12.7) |
| Alcohol consumption | |
| No | 1670 (70.6) |
| Yes | 694 (29.4) |
| Betel quid consumption | |
| No | 2309 (97.7) |
| Yes | 55 (2.3) |
*mean ± S.D. or n (%).
Under the generalized linear regression model, correlations with statistical significance were identified between a number of risk factors and the Z scores (Table 2), including the positive correlation between smoking and the Zmid (95% CI 0.01~0.15, p = 0.03) and Zhigh (95% CI 0.10~0.36, p = 0.0004) scores, as well as the positive correlation between the alcohol consumption and the Zhigh score (95% CI 0.01~0.20, p = 0.02). The consumption of betel quid correlated positively with the Zlow (95% CI 0.08~0.34, p = 0.002), Zmid (95% CI 0.16~0.47, p < 0.0001), and Zhigh (95% CI 0.40~0.94, p < 0.0001) scores, indicating that betel quid consumption contributed to elevated hearing thresholds across different frequencies in humans.
Table 2.
Generalized linear regression analyses of all risk factors affecting Z scores.
| Zlow | Zmid | Zhigh | |||||||
|---|---|---|---|---|---|---|---|---|---|
| β | 95% CI | p value | β | 95% CI | p value | β | 95% CI | p value | |
| Coronary heart disease | 0.02 | (−0.10, 0.14) | 0.73 | 0.04 | (−0.10, 0.18) | 0.54 | 0.01 | (−0.24, 0.26148) | 0.93 |
| Hypertension | −0.05 | (−0.11, 0.003) | 0.07 | −0.05 | (−0.11, 0.01) | 0.10 | −0.14 | (−0.25, −0.03) | 0.01 |
| Diabetes | 0.10 | (0.01, 0.19) | 0.02 | 0.15 | (0.05, 0.25) | 0.003 | 0.25 | (0.07, 0.43) | 0.01 |
| Hyperlipidemia | −0.01 | (−0.10, 0.08) | 0.75 | −0.02 | (−0.12, 0.08) | 0.68 | 0.01 | (−0.17, 0.20) | 0.90 |
| Chronic hepatitis | −0.03 | (−0.11, 0.06) | 0.57 | 0.002 | (−0.10, 0.10) | 0.95 | 0.11 | (−0.07, 0.29) | 0.21 |
| BMI | 0.01 | (−0.001, 0.01) | 0.10 | 0.01 | (0.01, 0.02) | 0.0002 | 0.02 | (0.004, 0.03) | 0.01 |
| Smoking | 0.05 | (−0.01, 0.11) | 0.14 | 0.08 | (0.01, 0.15) | 0.03 | 0.23 | (0.10, 0.36) | 0.0004 |
| Alcohol consumption | −0.01 | (−0.06, 0.03) | 0.62 | −0.01 | (−0.06, 0.05) | 0.78 | 0.11 | (0.01, 0.21) | 0.02 |
| Betel quid consumption | 0.21 | (0.08, 0.34) | 0.002 | 0.31 | (0.16, 0.47) | <0.0001 | 0.67 | (0.40, 0.94) | <0.0001 |
Consistent with our previous studies that demonstrated a significant association between obesity and SNHI26,27, BMI correlated positively with the Zmid (95% CI 0.01~0.02, p = 0.0002) and Zhigh (95% CI 0.004~0.03, p = 0.01) scores. Of note, diabetes and betel quid consumption were the two factors that were significantly associated with all the Zlow, Zmid, and Zhigh scores.
Arecoline caused derangement of hair cells in the organ of Corti
In this study we focused on the organ of Corti region to evaluate the effects of arecoline. Figure 1A shows the cross-section of the cochlear duct and the organ of Corti, which is composed of many types of cells, and the most important are the hair cells with hair bundles (HB) on the apical membrane. In contrast to untreated controls which showed a striped pattern with translucent bands at the organ of Corti (Fig. 1B, left panel), cochlear explants treated with arecoline revealed an opaque appearance (Fig. 1B, right panel). In the untreated cochlear explants, three rows of outer hair cells and one row of inner hair cells retained the original regular arrangement and the v-shaped hair bundles (Fig. 2A,B). On the contrary, the cochlear explants with the short-term and high-dose treatment of 4 mM or 10 mM arecoline showed disrupted hair cell arrangement with the disappearance of some hair bundles, resulting in gaps in the hair cell regions, which phenomena is not caused by the long-period culture (Fig. 2C,D, arrowheads).
Figure 2.

The effects of arecoline on cochlear hair cells. (A) A schematic diagram showing the orientation of both outer hair cells (OHCs) and inner hair cells (IHCs). The mechanosensitive hair bundles (HB) of hair cells are clusters of stereocilia on top of each hair cell and can be stained with phalloidin. (B–D) The hair bundles labeled by phalloidin after 30 minutes of arecoline treatment. In the control group (B), the three rows of outer hair cells and one row of inner hair cells retained the original regular arrangement and v-shaped hair bundles. By contrast, explants treated with 4 mM (C) or 10 mM (D) arecoline showed disrupted hair cell arrangement with disappearance of some hair bundles, resulting in vacancies in the hair cell regions (arrowheads). HB, hair bundles; IHC, inner hair cells; OHC, outer hair cells. Bar = 10 μm (B–D).
Arecoline decreased cell activity and exerted cytotoxicity in cochlear explants
After treatment with 10 mM arecoline, the cell activity of cochlear explants decreased dramatically on day 3 and stayed at a low level until day 7, indicating that cells were seriously damaged within 3 days of treatment (Fig. 3A). A rather moderate decline in the cell activity was observed after treatment with 2 mM arecoline, which showed similar cell activity level on day 3 as the cochlear explants treated with 800 μM or 200 μM arecoline, but started to decline on day 5, and finally diminished to the same level as the 10 mM arecoline-treated cochlear explants on day 7. This finding suggested that 2 mM of arecoline is sufficient to cause toxic effects in cochlear explants.
Figure 3.
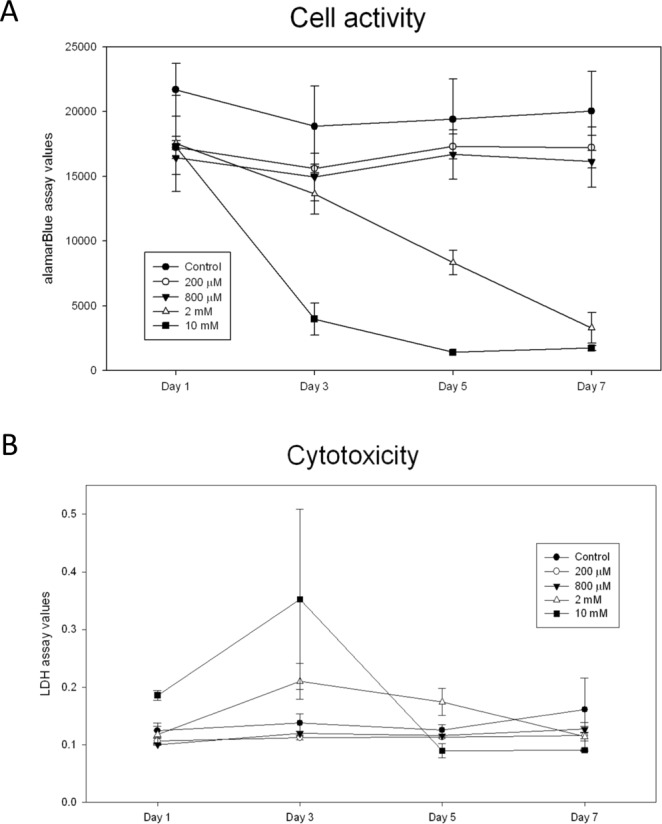
The cell activity and cytotoxicity assays of the cochlear explants. (A) The cell activity determined by the alamarBlue® assay. Experiments were performed under 5 different conditions: control, 200 μM, 800 μM, 2 mM, and 10 mM arecoline (n = 3). The data were recorded on day 1, 3, 5, and 7. (B) The cytotoxicity determined by LDH values. Experiments were also performed under 5 different conditions: control, 200 μM, 800 μM, 2 mM, and 10 mM arecoline. The data were recorded on day 1, 3, 5, and 7 (n = 3). Data are expressed as mean ± SEM.
Figure 3B illustrates the cytotoxicity in terms of LDH levels after treatment with various concentrations of arecoline. Parallel to the results in the cell activity, treatment with 200 μM or 800 μM arecoline did not cause obvious cell death in the cochlear explants, whereas treatment with 2 mM or 10 mM arecoline led to a surge in LDH on day 3, concurrent with the decrease in cell activity shown in Fig. 3A. The LDH levels then decreased on day 5 commensurate to that of the control group, indicating that the cell death caused by arecoline mainly occurred within the first 3 days.
Arecoline induced cell death in cochlea explants
To specify the cell types where death events occurred after arecoline treatment, we performed the TUNEL assay to label the DNA breaks of the apoptotic cells. On day 3, compared to the relatively normal appearance observed in the controls and cochlear explants treated with 800 μM arecoline, the cochlear explants treated with 2 mM arecoline showed positive nuclear BrdU stains at the organ of Corti (Fig. 4A, lower panel, arrowheads).
Figure 4.

The cell death detected by the TUNEL assay. The BrdU labeled the nuclei of dead cells (green) on day 3 (A) and day 7 (B). Red: phalloidin-labeled f-actin and hair bundles; blue: DAPI-labeled nuclei. On day 3 after treatment with 2 mM arecoline, positive green TUNEL signals appeared mainly within the hair cells (arrowheads), which then progressed to a severely damaged pattern of a wide-spread fragmented nuclei on day 7. After treatment with 800 μM arecoline, there were no positive green TUNEL signals on day 3, but some apoptotic cells (arrows) within the hair cell region could be observed on day 7. (C) The numbers of hair cells with distinguishable hair bundles were counted per 100 μm section of the middle region of cochlear duct. At day 3, the 2 mM arecoline-treated explants retained significantly fewer hair cells with distinguishable hair bundles than the untreated and 800 μM arecoline-treated explants, which further decreased to 0 cells at day 7. At day 7, the 800 μM arecoline-treated explants also revealed significantly fewer hair cells with distinguishable hair bundles than the untreated explants. Error bars represent SEM. HC, hair cells; IHC, inner hair cells; OHC: outer hair cells. Bar = 10 μm (A,B). *p < 0.01.
On day 7, cochlear explants treated with 800 μM arecoline revealed some degeneration of the hair bundles accompanied with some positive BrdU stains at the organ of Corti region (Fig. 4B, middle panel, arrows), whereas the cochlear explants treated with 2 mM arecoline exhibited a severely injured pattern, where phalloidin stains showed no discernible hair bundles and demonstrated an apoptotic pattern, accompanied by scattered BrdU stains (Fig. 4B, lower panel). These findings indicated that apoptosis after arecoline treatment developed predominantly in the hair cells. The numbers of hair cells with distinguishable hair bundles were counted per 100 μm section of cochlear duct (Fig. 4C). At day 3, the 2 mM arecoline-treated cochlear explants retained significantly fewer hair cells with distinguishable hair bundles (n = 14 ± 6) than the untreated (n = 62 ± 7) and 800 μM arecoline-treated cochlear explants (n = 66 ± 1) (both p < 0.01), which further decreased to 0 cells at day 7. At day 7, the 800 μM arecoline-treated cochlear explants also revealed significantly fewer hair cells with distinguishable hair bundles (n = 9 ± 4) than the untreated cochlear explants (n = 52 ± 3).
Arecoline induced ROS generation in the cochlea
On day 2, there were detectable ROS signals (green) in the untreated cochlear explants (Fig. 5A), while fuzzy fluorescent signals for ROS were detected around the hair cell band region in the 2 mM arecoline-treated cochlear explants (Fig. 5B). With the arecoline concentration being increased to 10 mM, ROS signals could be detected as early as 3 h after treatment, especially at the nuclei of the hair cells (Fig. 5C, arrows). It is noteworthy that there were some weak signals could be detected at the hair bundles, but we only focused on the nucleus-shaping signal in this study. Meanwhile, derangement and disappearance of hair bundles were also observed.
Figure 5.

ROS generated in the cochlear explant. Cellular oxidative stress within the cochlea explant was examined using a fluorogenic probe which could be oxidized by reactive oxygen species (ROS). On day 2, there were no detectable ROS signals (green) in the untreated explants (A), while fuzzy fluorescent signals were detected around the hair cell band region (marked as HC region) in the 2 mM arecoline-treated explants (B). (C) The ROS signals could be detected within 3 h when the arecoline concentration was increased to 10 mM, especially in the hair cells (arrows). Disruption and disappearance of hair bundles within the hair cell region could also be observed. The z-projection column demonstrates the summed-up signals of all the optical sections, which represents the total ROS signal in the explant. The z-section column is specifically for the section at the nuclei level. The images of 10 mM group showed that many ROS signals were detected at the nuclei, indicating the oxidative stress generated (C, lower panel). HC, hair cells; IHC, inner hair cells; OHC: outer hair cells. Bar = 80 μm (A,B) and 30 μm (C).
Discussion
In this study, we observed that the consumption of betel quid constituted a significant risk factor for SNHI in a large clinical cohort. To elucidate the underlying mechanisms, we then investigated the toxicity of arecoline, the main neuroactive component of the betel quid, on tissue explants harvested from murine cochleae. Our findings revealed that arecoline could reduce the cell activity and induce apoptosis in the cochlear explants, and confirmed oxidative stress as one of the possible mechanisms.
The cochlea is a highly differentiated and structured organ containing multiple types of cells, including inner hair cells, outer hair cells, supporting cells, spiral ganglion neuronal cells, as well as other specialized epithelial cells. The auditory function and physiological homeostasis are maintained by intricate interactions between these specialized cells. As such, even though several cell lines are available for hearing research34, organotypic cultures remained an invaluable experimental model. In this study, we dissected and cultured the whole cochlear explants from mice to simulate the effects of arecoline in the in vivo condition. Our findings revealed that arecoline started to exhibit cytotoxic effects on the cochlea explants at a concentration between 800 μM and 2 mM (Fig. 3A,B). Notably, this cytotoxic concentration is higher than that of 50–200 μM in the neural cell lines25, indicating that cochlear explants are more resistant to arecoline toxicity than neural cell lines. It has been reported that the supporting cells in the cochlea could protect hair cells by secreting Heat shock protein 70 (HSP70)35. Therefore, it is conceivable that the intact organotypic structure of the cochlear explant might provide the hair cells with additional protective effects against toxic agents like arecoline.
Next, we conducted multi-parameter cell viability and cytotoxicity assessments to delineate the profile of tissue adaptability in response to arecoline treatment. The alamarBlue assay measures the metabolic activity of the mitochondrial enzymes of living cells (Fig. 3A), whereas the LDH assay detects the lactate dehydrogenase released from the cytosol of lysed cells36 (Fig. 3B). As the surge of LDH on day 3 after arecoline treatment represented the destruction of the integrity of the cytoplasmic membrane, it could be inferred that arecoline caused irreversible changes in the cells of the cochlear explants, resulting in decreased cellular metabolic activity, and eventually cell death.
To specify the types of cells damaged by the arecoline treatment, we further performed TUNEL assays to investigate the apoptotic process. Our findings showed that, corresponding to the morphological changes such as hair bundle degeneration, apoptosis mainly occurred in the hair cell region of the cochlear explants (Fig. 4). Different ototoxic substances could cause specific damage to different sub-populations of cells in the cochlea. For instance, ouabain leads to degeneration of neuronal cells in the spiral ganglion37, erythromycin causes dysfunction in the epithelial cells of the stria vascularis38, aminoglycosides induce death of the outer and inner hair cells39,40, whereas platinum-containing chemotherapeutic agents result in a wider range of damage involving hair cells, neuronal cells, and epithelial cells of the stria vascularis41. As our findings revealed that the hair cells were the predominant cell type damaged by arecoline, it could be assumed that the ototoxic mechanisms of arecoline might resemble those of the aminoglycosides and platinum-containing chemotherapeutic agents. Previous studies demonstrated that overproduction of ROS might play a pivotal role in the ototoxic mechanisms of aminoglycosides39 and platinum-containing chemotherapeutic agents41; and oxidative stress has also been implicated in the carcinogenesis related to betel quid chewing42.
The postulation regarding the ototoxic mechanisms of arecoline was confirmed by our observation that strong ROS signals were detected in the hair cell regions of the cochlear explants after arecoline treatment (Fig. 5B,C). This finding of increased ROS levels in the cochlear explants is in agreement with previous studies in neural cell lines which reported that arecoline might induce neurotoxicity through increased oxidative stress25,43. Elevated ROS generation and subsequent apoptosis have been implicated in various types of hearing loss, including noise-induced hearing loss, ototoxicity-induced hearing loss, and age-related hearing impairment4,44,45. In a healthy cell, a delicate balance of pro- and anti-apoptotic factors exists, allowing it to live and proliferate. In stress situations like noise exposure, exposure to ototoxic drugs, and aging, this balance might be disturbed, and specific cells in the cochlea may enter the apoptotic death program. The knowledge concerning the pathogenetic roles of ROS and apoptosis have advanced the preventive strategies against hearing loss. For example, the application of certain antioxidant molecules, such as Q-ter46, N-acetylcysteine47,48, D-methionine49–51, and glutathione52, has been demonstrated to be effective in preventing the development of noise-induced and ototoxicity-induced hearing loss.
In the hair cell region, the outer hair cells appeared to be more vulnerable to the arecoline treatment than the inner hair cells. Previous studies revealed that outer hair cells were susceptible to various apoptosis-inducing stimuli, and significant apoptosis in outer hair cells was observed in noise-induced hearing loss53, as well as in hearing loss caused by aminoglycosides40,54 and cisplatin55. Different subpopulations of cells in the cochlea might exhibit different expression patterns of ROS-related proteins. For instance, the superoxide-producing NADPH oxidase 3 (NOX3) is highly expressed in the hair cells of the organ of Corti56, contributing to differences in the ROS concentration among different cochlear cells. In addition, the differences in the expression of certain apoptosis-regulating molecules (e.g. Bcl2 and Bcl6) between inner and outer hair cells, as shown in recent transcriptomic studies57,58, might also be a possible explanation for the susceptibility of outer hair cells to early cell death.
To our knowledge, this study is amongst the first to document the relevance of betel quid consumption to common neurological diseases such as SNHI. However, some limitations of this study deserve discussion. First, the cytotoxic concentrations of arecoline determined in the cochlear explants were not validated in vivo, because it was impossible to obtain cochlear tissue or fluid from the human subjects. However, as our findings have confirmed that arecoline resulted in increased ROS in the hair cells, it would not be surprising that long-term accumulation of oxidative stress might cause damage to the inner ear in humans and contribute to SNHI in individuals who consume betel quid. Second, although our results showed that increased ROS could be the crucial factor which induced hair cell apoptosis after arecoline treatment, it must be emphasized that there might be other apoptosis-inducing stimuli which do not depend on ROS production that could also contribute to the apoptotic process. It has been reported that increased Ca2+ in the outer hair cells59 or the activation of calcineurin60 may also trigger apoptotic pathways without ROS production. Further studies are warranted to elucidate the comprehensive mechanisms of arecoline in the pathogenesis of SNHI.
In conclusion, we identified that betel quid consumption might contribute to SNHI in humans, and confirmed that arecoline of the betel quid could induce apoptosis in hair cells through oxidative stress in ex vivo experiments. These findings have important public health implications by expanding the potential hazards of betel quid to common neurological disorders, and provide insights into the preventive strategies against SNHI or other neurological diseases caused by environmental neuro-toxic substances.
Acknowledgements
The authors are grateful to all the participants for their cooperation in the present study. This work was supported by research grants from the Ministry of Science and Technology of the Republic of China (MOST-106-2314-B-303-010-MY3) and the National Taiwan University Hospital (201607021RINB). We thank the staff of the imaging core at the First Core Lab, National Taiwan University College of Medicine, for their technical assistance.
Author Contributions
C.J. Hsu, and C.C. Wu conceived and designed the study., C.K. Liao, H.J. Lin, C.H. Tsai, and Y.L.L. Lee collected the clinical data and performed clinical data statistical analyses. Y.H. Chan, Y.C. Lu, and C.J. Hu performed the cochlear explants experiments analyzed the data. Y.H. Chan, Y.F. Cheng, and CCW drafted manuscript. C.J. Hsu and T.C. Liu supervised the whole study and provided critical revision. All authors reviewed the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Morton CC, Nance WE. Newborn hearing screening–a silent revolution. N Engl J Med. 2006;354:2151–2164. doi: 10.1056/NEJMra050700. [DOI] [PubMed] [Google Scholar]
- 2.Seidman, M. D., Ahmad, N. & Bai, U. Molecular mechanisms of age-related hearing loss. Ageing Res Rev1, 331–343, S1568163702000041 (2002). [DOI] [PubMed]
- 3.Van Eyken, E., Van Camp, G. & Van Laer, L. The complexity of age-related hearing impairment: contributing environmental and genetic factors. Audiol Neurootol12, 345–358, 000106478 (2007). [DOI] [PubMed]
- 4.Op de Beeck K, Schacht J, Van Camp G. Apoptosis in acquired and genetic hearing impairment: the programmed death of the hair cell. Hear Res. 2011;281:18–27. doi: 10.1016/j.heares.2011.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wilson D. H., Walsh P. G., Sanchez L., Davis A. C., Taylor A. W., Tucker G., Meagher I. The epidemiology of hearing impairment in an Australian adult population. International Journal of Epidemiology. 1999;28(2):247–252. doi: 10.1093/ije/28.2.247. [DOI] [PubMed] [Google Scholar]
- 6.Fetoni AR, Picciotti PM, Paludetti G, Troiani D. Pathogenesis of presbycusis in animal models: a review. Experimental gerontology. 2011;46:413–425. doi: 10.1016/j.exger.2010.12.003. [DOI] [PubMed] [Google Scholar]
- 7.Batts SA, Shoemaker CR, Raphael Y. Notch signaling and Hes labeling in the normal and drug-damaged organ of Corti. Hear Res. 2009;249:15–22. doi: 10.1016/j.heares.2008.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kim YH, Raphael Y. Cell division and maintenance of epithelial integrity in the deafened auditory epithelium. Cell Cycle. 2007;6:612–619. doi: 10.4161/cc.6.5.3929. [DOI] [PubMed] [Google Scholar]
- 9.Gupta, P. C. & Ray, C. S. Epidemiology of betel quid usage. Ann Acad Med Singapore33, 31–36, https://www.ncbi.nlm.nih.gov/pubmed/15389304 (2004). [PubMed]
- 10.Gupta PC, Warnakulasuriya S. Global epidemiology of areca nut usage. Addict Biol. 2002;7:77–83. doi: 10.1080/13556210020091437. [DOI] [PubMed] [Google Scholar]
- 11.Nair, U., Bartsch, H. & Nair, J. Alert for an epidemic of oral cancer due to use of the betel quid substitutes gutkha and pan masala: a review of agents and causative mechanisms. Mutagenesis19, 251–262, https://www.ncbi.nlm.nih.gov/pubmed/15215323 (2004). [DOI] [PubMed]
- 12.Garg A, Chaturvedi P, Gupta PC. A review of the systemic adverse effects of areca nut or betel nut. Indian J Med Paediatr Oncol. 2014;35:3–9. doi: 10.4103/0971-5851.133702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mehrtash H, et al. Defining a global research and policy agenda for betel quid and areca nut. Lancet Oncol. 2017;18:e767–e775. doi: 10.1016/S1470-2045(17)30460-6. [DOI] [PubMed] [Google Scholar]
- 14.Chen, Y. C. et al. Areca Nut Chewing Complicated with Non-Obstructive and Obstructive ST Elevation Myocardial Infarction. Acta Cardiol Sin32, 103–107, https://www.ncbi.nlm.nih.gov/pubmed/27122938 (2016). [DOI] [PMC free article] [PubMed]
- 15.Chiang, W. T., Yang, C. C., Deng, J. F. & Bullard, M. Cardiac arrhythmia and betel nut chewing–is there a causal effect? Vet Hum Toxicol40, 287–289, https://www.ncbi.nlm.nih.gov/pubmed/9778766 (1998). [PubMed]
- 16.Zhou J, Sun Q, Yang Z, Zhang J. The hepatotoxicity and testicular toxicity induced by arecoline in mice and protective effects of vitamins C and e. Korean J Physiol Pharmacol. 2014;18:143–148. doi: 10.4196/kjpp.2014.18.2.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Taylor, R. F., al-Jarad, N., John, L. M., Conroy, D. M. & Barnes, N. C. Betel-nut chewing and asthma. Lancet339, 1134-1136, https://www.ncbi.nlm.nih.gov/pubmed/1349367 (1992). [DOI] [PubMed]
- 18.Boucher BJ, Mannan N. Metabolic effects of the consumption of Areca catechu. Addict Biol. 2002;7:103–110. doi: 10.1080/13556210120091464. [DOI] [PubMed] [Google Scholar]
- 19.Hsu HF, et al. Effects of arecoline on adipogenesis, lipolysis, and glucose uptake of adipocytes-A possible role of betel-quid chewing in metabolic syndrome. Toxicol Appl Pharmacol. 2010;245:370–377. doi: 10.1016/j.taap.2010.04.008. [DOI] [PubMed] [Google Scholar]
- 20.Chang YF, Liu TY, Liu ST. Arecoline inhibits and destabilizes agrin-induced acetylcholine receptor cluster formation in C2C12 myotubes. Food Chem Toxicol. 2013;60:391–396. doi: 10.1016/j.fct.2013.07.079. [DOI] [PubMed] [Google Scholar]
- 21.Wu PF, et al. A characterization of the antioxidant enzyme activity and reproductive toxicity in male rats following sub-chronic exposure to areca nut extracts. J Hazard Mater. 2010;178:541–546. doi: 10.1016/j.jhazmat.2010.01.118. [DOI] [PubMed] [Google Scholar]
- 22.Hu CW, Chao MR. Direct-acting DNA alkylating agents present in aqueous extracts of areca nut and its products. Chem Res Toxicol. 2012;25:2386–2392. doi: 10.1021/tx300252r. [DOI] [PubMed] [Google Scholar]
- 23.Tsai YS, et al. Arecoline, a major alkaloid of areca nut, inhibits p53, represses DNA repair, and triggers DNA damage response in human epithelial cells. Toxicology. 2008;249:230–237. doi: 10.1016/j.tox.2008.05.007. [DOI] [PubMed] [Google Scholar]
- 24.Lin Ming-Hsun, Chou Fen-Pi, Huang Hui-Pei, Hsu Jeng-Dong, Chou Ming-Yung, Wang Chau-Jong. The tumor promoting effect of lime-piper betel quid in JB6 cells. Food and Chemical Toxicology. 2003;41(11):1463–1471. doi: 10.1016/S0278-6915(03)00124-8. [DOI] [PubMed] [Google Scholar]
- 25.Shih YT, et al. Arecoline, a major alkaloid of the areca nut, causes neurotoxicity through enhancement of oxidative stress and suppression of the antioxidant protective system. Free Radic Biol Med. 2010;49:1471–1479. doi: 10.1016/j.freeradbiomed.2010.07.017. [DOI] [PubMed] [Google Scholar]
- 26.Wu CC, et al. Contribution of adiponectin and its type 1 receptor to age-related hearing impairment. Neurobiology of aging. 2015;36:2085–2093. doi: 10.1016/j.neurobiolaging.2015.02.030. [DOI] [PubMed] [Google Scholar]
- 27.Hwang JH, Wu CC, Hsu CJ, Liu TC, Yang WS. Association of central obesity with the severity and audiometric configurations of age-related hearing impairment. Obesity. 2009;17:1796–1801. doi: 10.1038/oby.2009.66. [DOI] [PubMed] [Google Scholar]
- 28.Lin YH, Wu CC, Hsu CJ, Hwang JH, Liu TC. The Grainyhead-Like 2 Gene (GRHL2) Single Nucleotide Polymorphism Is Not Associated with Age-Related Hearing Impairment in Han Chinese. Laryngoscope. 2011;121:1303–1307. doi: 10.1002/lary.21771. [DOI] [PubMed] [Google Scholar]
- 29.Fransen Erik, Van Laer Lut, Lemkens Nele, Caethoven Goele, Flothmann Kris, Govaerts Paul, Van de Heyning Paul, Van Camp Guy. A Novel Z-Score–Based Method to Analyze Candidate Genes for Age-Related Hearing Impairment. Ear and Hearing. 2004;25(2):133–141. doi: 10.1097/01.AUD.0000120362.69077.0B. [DOI] [PubMed] [Google Scholar]
- 30.Larson, E. M., Doughman, D. J., Gregerson, D. S. & Obritsch, W. F. A new, simple, nonradioactive, nontoxic in vitro assay to monitor corneal endothelial cell viability. Invest Ophthalmol Vis Sci38, 1929–1933, https://www.ncbi.nlm.nih.gov/pubmed/9331256 (1997). [PubMed]
- 31.Decker, T. & Lohmann-Matthes, M. L. A quick and simple method for the quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis factor (TNF) activity. J Immunol Methods115, 61–69, https://www.ncbi.nlm.nih.gov/pubmed/3192948 (1988). [DOI] [PubMed]
- 32.Bortner, C. D., Oldenburg, N. B. & Cidlowski, J. A. The role of DNA fragmentation in apoptosis. Trends Cell Biol5, 21–26, https://www.ncbi.nlm.nih.gov/pubmed/14731429 (1995). [DOI] [PubMed]
- 33.Soares J, et al. A tryptophanol-derived oxazolopiperidone lactam is cytotoxic against tumors via inhibition of p53 interaction with murine double minute proteins. Pharmacol Res. 2015;95-96:42–52. doi: 10.1016/j.phrs.2015.03.006. [DOI] [PubMed] [Google Scholar]
- 34.Rivolta MN, Holley MC. Cell lines in inner ear research. J Neurobiol. 2002;53:306–318. doi: 10.1002/neu.10111. [DOI] [PubMed] [Google Scholar]
- 35.May LA, et al. Inner ear supporting cells protect hair cells by secreting HSP70. J Clin Invest. 2013;123:3577–3587. doi: 10.1172/JCI68480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bopp SK, Lettieri T. Comparison of four different colorimetric and fluorometric cytotoxicity assays in a zebrafish liver cell line. BMC Pharmacol. 2008;8:8. doi: 10.1186/1471-2210-8-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yuan Y, et al. Ouabain-induced cochlear nerve degeneration: synaptic loss and plasticity in a mouse model of auditory neuropathy. J Assoc Res Otolaryngol. 2014;15:31–43. doi: 10.1007/s10162-013-0419-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Liu, J., Marcus, D. C. & Kobayashi, T. Inhibitory effect of erythromycin on ion transport by stria vascularis and vestibular dark cells. Acta Otolaryngol116, 572–575, https://www.ncbi.nlm.nih.gov/pubmed/8831844 (1996). [DOI] [PubMed]
- 39.Wu WJ, Sha SH, Schacht J. Recent advances in understanding aminoglycoside ototoxicity and its prevention. Audiol Neurootol. 2002;7:171–174. doi: 10.1159/000058305. [DOI] [PubMed] [Google Scholar]
- 40.Hirose K, Li SZ, Ohlemiller KK, Ransohoff RM. Systemic lipopolysaccharide induces cochlear inflammation and exacerbates the synergistic ototoxicity of kanamycin and furosemide. J Assoc Res Otolaryngol. 2014;15:555–570. doi: 10.1007/s10162-014-0458-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rybak LP, Whitworth CA, Mukherjea D, Ramkumar V. Mechanisms of cisplatin-induced ototoxicity and prevention. Hear Res. 2007;226:157–167. doi: 10.1016/j.heares.2006.09.015. [DOI] [PubMed] [Google Scholar]
- 42.Wong RH, et al. Sulfotransferase 1A1 and glutathione S-transferase P1 genetic polymorphisms modulate the levels of urinary 8-hydroxy-2′-deoxyguanosine in betel quid chewers. Arch Toxicol. 2008;82:313–321. doi: 10.1007/s00204-007-0248-8. [DOI] [PubMed] [Google Scholar]
- 43.Jiang JM, et al. Arecoline Induces Neurotoxicity to PC12 Cells: Involvement in ER Stress and Disturbance of Endogenous H2S Generation. Neurochem Res. 2016;41:2140–2148. doi: 10.1007/s11064-016-1929-6. [DOI] [PubMed] [Google Scholar]
- 44.Fujimoto C, Yamasoba T. Oxidative stresses and mitochondrial dysfunction in age-related hearing loss. Oxid Med Cell Longev. 2014;2014:582849. doi: 10.1155/2014/582849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kamogashira T, Fujimoto C, Yamasoba T. Reactive oxygen species, apoptosis, and mitochondrial dysfunction in hearing loss. Biomed Res Int. 2015;2015:617207. doi: 10.1155/2015/617207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fetoni AR, Piacentini R, Fiorita A, Paludetti G, Troiani D. Water-soluble Coenzyme Q10 formulation (Q-ter) promotes outer hair cell survival in a guinea pig model of noise induced hearing loss (NIHL) Brain Res. 2009;1257:108–116. doi: 10.1016/j.brainres.2008.12.027. [DOI] [PubMed] [Google Scholar]
- 47.Ohinata Y, Miller JM, Schacht J. Protection from noise-induced lipid peroxidation and hair cell loss in the cochlea. Brain Research. 2003;966:265–273. doi: 10.1016/S0006-8993(02)04205-1. [DOI] [PubMed] [Google Scholar]
- 48.Wu YJ, Muldoon LL, Neuwelt EA. The chemoprotective agent N-acetylcysteine blocks cisplatin-induced apoptosis through caspase signaling pathway. J Pharmacol Exp Ther. 2005;312:424–431. doi: 10.1124/jpet.104.075119. [DOI] [PubMed] [Google Scholar]
- 49.Campbell KCM, et al. Prevention of noise- and drug-induced hearing loss with D-methionine. Hearing Res. 2007;226:92–103. doi: 10.1016/j.heares.2006.11.012. [DOI] [PubMed] [Google Scholar]
- 50.Sha, S. H. & Schacht, J. Antioxidants attenuate gentamicin-induced free radical formation in vitro and ototoxicity in vivo: D-methionine is a potential protectant. Hear Res142, 34–40, https://www.ncbi.nlm.nih.gov/pubmed/10748326 (2000). [DOI] [PubMed]
- 51.Campbell KCM, Rybak LP, Meech RP, Hughes L. D-Methionine provides excellent protection from cisplatin ototoxicity in the rat. Hearing Res. 1996;102:90–98. doi: 10.1016/S0378-5955(96)00152-9. [DOI] [PubMed] [Google Scholar]
- 52.Hight NG, McFadden SL, Henderson D, Burkard RF, Nicotera T. Noise-induced hearing loss in chinchillas pre-treated with glutathione monoethylester and R-PIA. Hearing Res. 2003;179:21–32. doi: 10.1016/S0378-5955(03)00067-4. [DOI] [PubMed] [Google Scholar]
- 53.Hu, B. H., Henderson, D. & Nicotera, T. M. Involvement of apoptosis in progression of cochlear lesion following exposure to intense noise. Hear Res166, 62–71, https://www.ncbi.nlm.nih.gov/pubmed/12062759 (2002). [DOI] [PubMed]
- 54.Suzuki M, Ushio M, Yamasoba T. Time course of apoptotic cell death in guinea pig cochlea following intratympanic gentamicin application. Acta Otolaryngol. 2008;128:724–731. doi: 10.1080/00016480701714244. [DOI] [PubMed] [Google Scholar]
- 55.Alam Shaheen Ara, Ikeda Katsuhisa, Oshima Takeshi, Suzuki Masaaki, Kawase Tetsuaki, Kikuchi Toshihiko, Takasaka Tomonori. Cisplatin-induced apoptotic cell death in Mongolian gerbil cochlea. Hearing Research. 2000;141(1-2):28–38. doi: 10.1016/S0378-5955(99)00211-7. [DOI] [PubMed] [Google Scholar]
- 56.Poirrier, A. L., Pincemail, J., Van Den Ackerveken, P., Lefebvre, P. P. & Malgrange, B. Oxidative Stress in the Cochlea: An Update. Curr Med Chem17, 3591–3604, https://doi.org/<Go to ISI>://WOS:000284654300011 (2010). [DOI] [PubMed]
- 57.Li Y, et al. Transcription Factors Expressed in Mouse Cochlear Inner and Outer Hair Cells. PLoS One. 2016;11:e0151291. doi: 10.1371/journal.pone.0151291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Liu H, et al. Characterization of transcriptomes of cochlear inner and outer hair cells. J Neurosci. 2014;34:11085–11095. doi: 10.1523/JNEUROSCI.1690-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Fridberger, A., Flock, A., Ulfendahl, M. & Flock, B. Acoustic overstimulation increases outer hair cell Ca2+ concentrations and causes dynamic contractions of the hearing organ. Proc Natl Acad Sci USA95, 7127–7132, https://www.ncbi.nlm.nih.gov/pubmed/9618550 (1998). [DOI] [PMC free article] [PubMed]
- 60.Minami SB, Yamashita D, Schacht J, Miller JM. Calcineurin activation contributes to noise-induced hearing loss. J Neurosci Res. 2004;78:383–392. doi: 10.1002/jnr.20267. [DOI] [PubMed] [Google Scholar]