

Abstract
Human milk has antimicrobial compounds and immunomodulatory activities. We investigated glycerol monolaurate (GML) in human milk versus bovine milk and infant formula for antimicrobial and anti-inflammatory activities. Human milk contained approximately 3000 µg/ml of GML, compared to 150 μg/ml in bovine milk and none in infant formula. For bacteria tested (Staphylococcus aureus, Bacillus subtilis, Clostridium perfringens, Escherichia coli), except Enterococcus faecalis, human milk was more antimicrobial than bovine milk and formula. The Enterococcus faecalis strain, which was not inhibited, produced reutericyclin, which is an analogue of GML and functions as a growth stimulant in bacteria that produce it. Removal of GML and other lipophilic molecules from human milk by ethanol extraction resulted in a loss of antibacterial activity, which was restored by re-addition of GML. GML addition caused bovine milk to become antimicrobial. Human milk but not bovine milk or formula inhibited superantigen and bacterial-induced IL-8 production by model human epithelial cells. GML may contribute beneficially to human milk compared to bovine milk or infant formula.
Subject terms: Paediatric research, Infection, Acute inflammation
Introduction
Human milk contains many known antimicrobial and immunomodulatory molecules, including immunoglobulins, antimicrobial peptides, and fatty acids1–5. The cumulative effect of the properties of human milk, including ability of bacterial species to utilize human milk oligosaccharides, is selection for bacteria such as bifidobacteria, lactobacilli, and enterococci, genera of bacteria that have acids and hydrogen peroxide as major end products of metabolism6–10. The reduced pH caused by these bacteria, and additionally, by known and unknown components of human breast milk make it more difficult for potentially pathogenic organisms to colonize6,7; for example potentially pathogenic Escherichia coli are inhibited6,7. Human milk, including human milk oligosaccharides11,12, also may reduce the occurrence of allergic disorders, including asthma, food allergy, and atopic dermatitis, the latter two of which have been linked to Staphylococcus aureus colonization in young children13–15.
We and others have characterized glycerol monolaurate (GML) as a fatty acid monoester with broad antimicrobial and anti-inflammatory properties16–21. Lauric acid, one of the esterase cleavage products of GML, also has potent antimicrobial and anti-inflammatory properties, but these properties require nearly 400-fold more lauric acid than GML21. Other fatty acid monoesters have significantly reduced antimicrobial activity compared to GML21. The antimicrobial properties of GML and free fatty acids extend to nearly all Gram-positive bacterial species except certain lactobacilli, bifidobacteria, and enterococci21,22. These resistant bacteria appear to be positively selected for due to GML mimicking a quorum-sensing growth stimulant known as reutericyclin or related tetramic acids21,23. The ultimate bacterial killing effect by GML appears to be due to interference with plasma membrane functions, resulting in loss of potential difference across the membrane23. Lactobacilli that produce reutericyclin have been incorporated into probiotic capsules in attempt to increase lactobacilli in the gastrointestinal tract24.
GML also prevents harmful pro-inflammatory processes in vivo at mucosal surfaces16–20,25, although in vitro studies with purified GML show toxicity to tissue culture cells in vitro at concentrations ≥100 μg/ml26–28. The differences between in vivo versus in vitro activities has not been investigated, but may in part be related to the modulation of GML toxicity by human serum albumin29. Our studies have shown, for example, that inflammation at the human and non-human primate vaginal mucosa facilitates simian immunodeficiency virus infection in rhesus macaque monkeys and production of menstrual toxic shock syndrome16,18–20,30. The production of local inflammation initially depends on microbial stimulation of epithelial cells, among other cells, to produce pro-inflammatory chemokines, such as IL-8 and MIP-3α, which attract innate and adaptive immune cells into the submucosal areas and facilitating barrier disruption19,20,30. Our studies also show that vaginal pathogens, but not lactobacilli and latex beads, activate epithelial cells to produce many expected pro-inflammatory chemokines16,19,22,30. GML interferes with normal signal transduction in epithelial cells and locally-recruited immune cells through membrane effects, but does not kill the host cells in vivo26–28. It is noteworthy that a small percentage of women (1%) have reutericyclin-producing lactobacilli and enterococci vaginally, and in such cases, there are minimal to no other vaginal microbes present; one of these enterococci is used in our current studies21. This raises the possibility that reutericyclin and its possible analogue GML have similar effects, both in killing other bacteria and preventing harmful inflammation, in the gastrointestinal tract21. Reutericyclin is expensive to purchase and evaluate, whereas GML is inexpensive and readily available for use in tests.
The goals of this study were to determine: (1) the amount of GML present in whole human versus whole bovine milk and commercial formula, and (2) the possible contribution of GML to the antimicrobial and anti-inflammatory properties of human milk compared to bovine milk and commercial formula. Our hypothesis was that GML may be present in human milk samples but not bovine milk and commercial formula. Furthermore, GML presence, along with other factors, may inhibit the growth of pathogens and at the same time exhibit anti-inflammatory activity. Overall, these studies provide important data on the advantages of use of human milk for infants.
Results
GML quantified in human and bovine milk samples and commercial formula
Four human milk, 6 bovine whole milk from cows, and 2 store-purchased whole bovine milk samples were quantified for GML (Fig. 1). All milk samples had been pasteurized. Whether compared to whole bovine milk obtained from individual cows or to whole bovine milk purchased from a local grocery store, the four tested human milk samples had approximately 20 times more GML than the 8 bovine samples. There was minimal variation in GML concentrations in the human milk samples. Commercial infant formula (Similac® Advance) contained no detectable GML.
Figure 1.
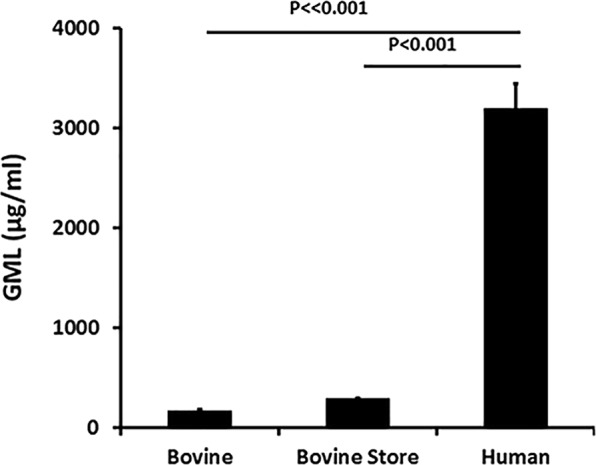
Glycerol monolaurate (GML) concentrations in 6 individual pasteurized whole bovine, 2 whole bovine pasteurized samples purchased from a local grocery store, and 4 pasteurized whole human milk samples. Data are recorded as mean ± SD. Student’s t test was used to assess differences in means. P ≪ 0.001 indicates mean differences much greater than p < 0.001.
Effect of whole human versus bovine milk and formula on growth of bacteria
We tested the human milk, bovine milk samples, and the commercial infant formula sample for ability to inhibit the growth of selected Gram-positive aerobes S. aureus (a potential gastrointestinal pathogen known for its roles in food poisoning31 and enterocolitis32,33; Fig. 2A) and B. subtilis (highly susceptible to killing by GML34 and essentially a positive control; Fig. 2B); we tested human and bovine milk for effect on the Gram-positive anaerobe C. perfringens (anaerobe; potential gastrointestinal pathogen and microflora; Fig. 2C); and we tested human and bovine milk and commercial infant formula for effect on the Gram-negative Escherichia coli (potential gastrointestinal and urinary tract pathogen, microflora, representative of Enterobacteriaceae; Fig. 2D). For all three Gram-positive bacteria, the 4 human milk samples comparably inhibited the growth of the strains compared to the bovine milk samples and commercial formula. B. subtilis was more susceptible to killing by the human milk samples than either of the other two organisms; we previously observed the same differential susceptibility when we studied the effects of purified GML on Gram-positive bacteria21,34. Both aerobes and obligate anaerobes were significantly inhibited by human milk samples. There were only minimal differences in inhibitory activity among the human milk samples. The Gram-negative organism, Escherichia coli, which is clearly both a commensal and one of the leading causes of gastrointestinal and urinary tract infections worldwide, was likewise inhibited by human milk but not whole bovine milk or infant formula. It is likely that the effect on E. coli, and likely on other organisms depended on synergy among GML and other antimicrobial compounds in human milk; E. coli is typically not susceptible to GML alone, even at 3000 µg/ml35.
Figure 2.

Effect of whole pasteurized human milk and bovine milk samples, and commercial infant formula on growth of (A) S. aureus, (B) B. subtilis, (C) C. perfringens, and (D) E. coli. Log colony-forming units (CFUs)/ml are shown for each treatment. Starting inoculum was 1–2 × 105/ml. Samples were cultured to stationary phase for 24 hours with aeration (200 RPM; A,B,D) and stationary in an anaerobic chamber (C). Data represent means ± SD. Student’s t test was used to determine significant differences in means. P ≪ 0.001 indicates mean differences much greater than p < 0.001.
The stationary phases of all tested organisms, for comparison to milk and formula, in excellent growth medium (Todd Hewitt broth, Difco Laboratories, Detroit, MI) were: S. aureus MN8 7 × 109/ml; B. subtilis 1 × 109/ml; C. perfringens 5 × 108/ml; and E. coli 2 × 109/ml. Thus, the stationary phases of these organisms were generally 0.5–1 log higher in Todd Hewitt medium than whole bovine milk and infant formula.
We next tested the effect of whole human and bovine milk and infant formula on the growth of an Enterococcus faecalis strain, essentially a negative control for inhibitory activity, (Fig. 3) obtained from a vaginal swab from a woman with pure culture22 of this organism. The strain produces reutericyclin, which may be an analogue of GML22. The growth of this organism was not inhibited by human or bovine milk, or infant formula. The stationary phase of growth for this organism in excellent growth medium (Todd Hewitt) was 5.0 × 108/ml. Based on the data obtained, the failure to inhibit the growth of Enterococcus faecalis by the milk sample provides evidence that GML in human milk is contributing to the antimicrobial activity against some but not all bacteria, and additionally shows that human milk can support the growth of some bacteria. This demonstrates that human milk is not nutritionally-deficient, but instead is actively antimicrobial.
Figure 3.

Effect of whole human and bovine milk and commercial infant formula on the growth of a reutericyclin-producing Enterococcus faecalis. Log colony-forming units (CFU)/ml are shown for each treatment. Starting inoculum was 105/ml. Samples were cultured with shaking at 200 RPM for 24 hours. Data represent mean ± SD. Student’s t test was used to determine significant differences in means; NS = Not Significant.
Effect of removal of GML and other lipids by ethanol extraction
We treated two human milk samples with 80% ethanol to precipitate molecules >10,000 molecular weight and leave GML and other soluble lipophilic molecules in the ethanol fraction. The precipitates were resolubilized with distilled water. We then tested the GML-free milk samples, containing 0.03 M glucose, an amount below the standard amount present in human milk in the form of lactose36. When GML and other lipophilic molecules were removed from the whole human milk samples, the antimicrobial activity against S. aureus was also removed (Fig. 4). We then added back 500 or 3000 µg/ml, amounts of highly-pure, food-grade GML that has been used by us in multiple studies on humans17,25; the 3000 µg/ml amount was designed in this experiment to overcome GML reduction in activity by human albumin29. Antimicrobial activity was restored by addition of 3000 µg/ml (Fig. 4) but not by addition of 500 µg/ml (data not shown). The fact that 3000 µg/ml GML add-back was more strongly bactericidal after ethanol extraction of human milk, suggests that additional nutrients besides glucose or antimicrobial factors were removed by the extraction process.
Figure 4.
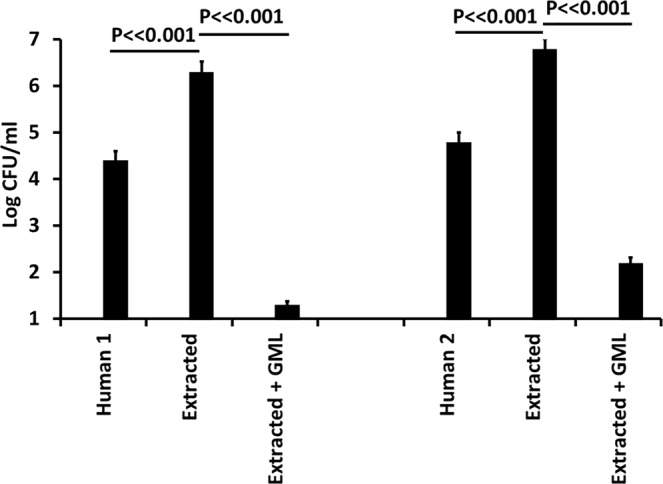
Effect of GML removal from two whole human milk samples (human 1, human 2) on growth of Staphylococcus aureus MN8, and effect of re-addition of 3000 µg/ml of GML. Extracted + GML refers to the resolubilized precipitates +3000 µg/ml GML. Log colony-forming units (CFUs)/ml are shown for each treatment. Starting bacterial inoculum was 4.9 × 104/ml. Data represent means ± SD. Student’s t test was used to determine significant differences in means. P ≪ 0.001 indicates mean differences much greater than p < 0.001.
S. aureus inhibitory activity of bovine milk after addition of GML
In Fig. 5, we examined the antimicrobial activity of bovine milk after addition of 0–5000 µg/ml of highly-pure GML. Whole bovine milk without GML addition was not antimicrobial. However, successive increases in GML amounts added resulted in antimicrobial activity, with milk containing 3000 and 5000 µg/ml having potent bactericidal activity.
Figure 5.
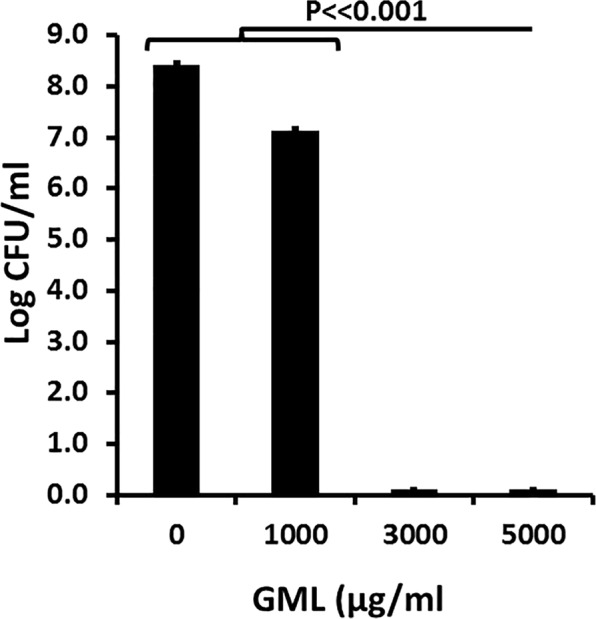
Dose response of GML (0–5000 µg/ml) added to bovine milk samples. Log colony-forming units/ml are shown for all samples. Starting inoculum was approximately 105/ml of bacteria. Samples incubated with shaking (200 RPM) for 24 hour at 37 °C. Data represent means ± SD. Student’s t test was used to determine significant differences in means. P ≪ 0.001 indicates mean differences much greater than p < 0.001.
Anti-inflammatory properties of human milk but not bovine milk
In addition to its antimicrobial activity, GML prevents harmful inflammation initiated by pathogens on mucosal surfaces16,18–21,25. Also, human milk has been used topically to help in the treatment of atopic dermatitis in children37. We thus hypothesized that whole human milk samples, but not whole bovine milk samples, have anti-inflammatory activities against epithelial cells. This was tested on confluent layers of human squamous epithelial cells (HSECs) used only as the model cell system (Fig. 6). In this model test system, as expected the superantigen toxic shock syndrome toxin-1 (TSST-1) caused significant production (in 6 hours) of the chemokine IL-8, as only one of TSST-1 induced cytokines from HSECs38. The two representative human milk samples alone added to the HSECs did not induce production of IL-8, but the bovine samples alone caused low-level IL-8 production, but not to the extent of TSST-1. When the two separate human milk samples were added to HSECs in the presence of TSST-1, there was minimal IL-8 produced, indicating both samples were anti-inflammatory. The HSECs alone were not killed by the human or bovine milk samples. Neither bovine sample reduced IL-8 production caused by TSST-1. This experiment was repeated one additional time with similar results. The ethanol-extracted milk samples shown in Fig. 4 lost ability to reduce IL-8 production by TSST-1 (data not shown). We did not fractionate the ethanol-extracted fraction to determine which component(s) were removed that resulted in loss of activity. However, it is likely that GML and other ethanol-soluble molecules contributed to anti-inflammatory activity of human milk samples.
Figure 6.
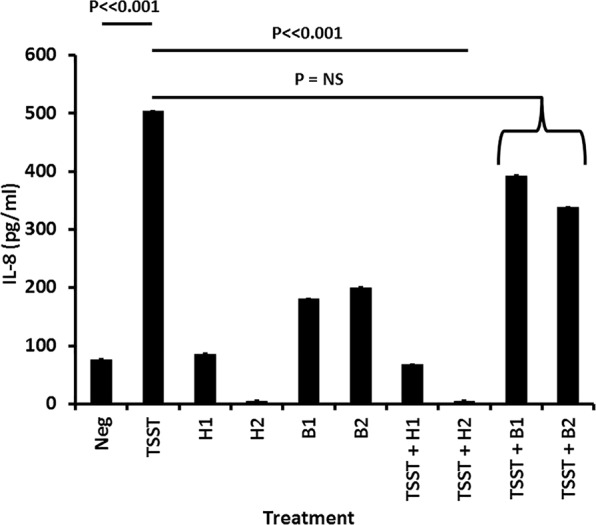
Anti-inflammatory activity of two whole human milk samples versus two whole bovine milk samples during stimulation of human squamous epithelial cells with the superantigen toxic shock syndrome toxin-1 (TSST). Data represent means ± SD of secreted chemokine IL-8, as the representative cytokine, after 6 hours incubation. Assays performed in triplicate in 96 well microtiter plates with confluent human epithelial cells + 200 µl keratinocyte serum-free medium with or without 20 µl milk samples. Student’s t test was used to determine significant differences in means. P ≪ 0.001 indicates mean differences much greater than p < 0.001; NS = Not Significant.
Human milk samples were also tested for ability to reduce IL-8 production from HSECs by the studied bacteria. These bacteria had previously been shown in our laboratory, some published and some unpublished, to stimulate IL-8 production from the HSECs over the 6 hour time period16,22,30. For all microbes, human milk reduced the IL-8 production to or near to background.
Collectively, our data suggest that human milk is both inhibitory to pathogen growth and exhibits anti-inflammatory activity with both activities in part dependent on GML. In contrast, bovine milk and commercial formula lacked or had greatly reduced GML and exhibited neither antimicrobial nor anti-inflammatory properties.
Discussion
It has long been known that breast feeding is highly beneficial to babies through antimicrobial and anti-inflammatory activities. Human milk fed babies select for bifidobacteria, lactobacilli, and enterococci, with limited microbial diversity6,7. These organisms are fermenters, producing primarily lactic acid, to some extent acetic acid, and hydrogen peroxide. These organisms plus many components of human milk help keep out potentially pathogenic bacteria. Bovine and commercial formula-fed infants in general have more microbial diversity such that bifidobacteria, lactobacilli, and enterococci numbers are reduced relatively39,40.
It is our experience that approximately 1 in 100 women have pure cultures of lactobacilli vaginally (in one case Enterococcus faecalis), with no other flora present22. These women stably maintain this mono-flora colonization. The lactobacilli and Enterococcus faecalis produce antimicrobial compounds, including a broadly antimicrobial and anti-inflammatory compound, reutericyclin, which is a quorum-sensing growth stimulant for these reutericyclin-producing bacteria23,41. At the same time, the bacteria have an immunity gene that protects them from reutericyclin41.
These observations led us to hypothesize that human breast milk may have an analogue of reutericyclin to cause the predominance of bifidobacteria, lactobacilli, and enterotococci in the gastrointestinal tract. In multiple prior studies we have shown that GML acts as an analogue of reutericyclin, with the same broad spectrum of antimicrobial activity, functioning as a quorum-sensing growth stimulant for lactobacilli, and having anti-inflammatory activity16–21,25. The mechanism of bacterial killing by GML, like reutericyclin, appears to be dissipation of the potential difference across bacteria plasma membranes21.
In the current study, we first observed that whole pasteurized human milk contains large amounts of GML, compared to 20-fold lesser amounts in whole pasteurized bovine milk. Milk in general contains albumin that potentially interferes with GML activity29. However, the amount of GML in human milk samples exceeds the interference by albumin, and additionally human milk contains many other antimicrobial compounds that may synergize with GML. This must be the case since E. coli, for example, is not susceptible to GML alone35. Thus, we suggest that a significant part of the antimicrobial activity we saw in whole human milk against tested bacteria results from GML alone and in concert with other antimicrobials. Enhancement of GML activity by other antimicrobials has been observed previously42,43. Other investigators have also shown that free fatty acids have antimicrobial activity44–46. For example, Kabara and colleagues showed that among saturated free fatty acids, lauric acid, the fatty acid contained in GML, has the greatest antimicrobial activity, with the same bacterial inhibitory spectrum as GML45. We have recently duplicated these findings, but our studies showed that GML is nearly 400-fold more active against S. aureus than lauric acid21. Indeed, Kabara and colleagues, who did much of the pioneering related to GML, later focused many of their studies on GML because of its increased activity46. Despite these findings, it is important to consider that human milk contains multiple other antimicrobial compounds that may synergize or act additively with GML.
Although we do not know why human milk contains so much GML, compared to bovine milk, this observation is supported by based differences in gastrointestinal tracts of humans and cows. Cows have four stomachs that must adapt to processing grasses and related plants through cellulases. This means cows must also have a quite different gut microbiome than humans, one that may depend on not having high concentrations of GML.
Most microbial pathogens stimulate harmful inflammation at mucosal surfaces17,19–21,25. We have previously provided strong evidence that the superantigen, TSST-1, as studied in the current manuscript, and simian immunodeficiency virus (SIV) cause harmful inflammation at such mucosal surfaces17,19–21,25. The net effect is to open the mucosal barrier to TSST-1 to result in menstrual TSS, and SIV to result in infection, and to attract innate and adaptive immune cells that ultimately are the direct targets for disease causation. In this manuscript, we show that whole human milk samples are anti-inflammatory as tested with IL-8 production, whereas ethanol-extracted human milk and whole bovine milk samples are not. While the significance of this is incompletely known, the data are consistent with slowing but not stopping development of harmful pathogen responses so long as babies are breast-fed39,40. It is recognized that intestinal microbiome organisms are essential for immune system development. However, over-reactivity may lead to harmful effects, such as asthma and atopic dermatitis.
Collectively, our study suggests that there are great benefits to human milk compared to bovine milk and formula. Positive effects of human milk appear to be due in part to the presence of GML combined with other known and unknown factors39,40. Future studies are needed to determine whether or not supplementation of bovine milk and commercial infant formula with GML will be beneficial.
Methods
All experiments with use of whole human milk and use of human epithelial cells were performed in accordance with accepted regulatory requirements. The human epithelial cells have been used by our laboratory in multiple publications since 200538. University of Iowa Institutional Review Board approval and consent (approval number 199910006) were obtained prior to obtaining and using these cells. The human milk samples were purchased from Mother’s Milk Bank of Iowa, Coralville, IA.; IRB approval was not required for use of these samples. The milk samples were not identified as to donor.
Bacteria and TSST-1
Staphylococcus aureus MN8 is a commonly used strain from a patient with menstrual TSS, but this clonal group of S. aureus is also associated with enterocolitis32,33,38. B. subtilis34, C. perfringens34, and E. coli21 are Schlievert laboratory strains of stock cultures. Enterococcus faecalis was obtained from a vaginal culture from a woman with pure culture of this organism; the strain produces reutericyclin22. These organisms are all maintained as −80 °C frozen cultures. We chose these strains for study to include bacteria that are considered typically highly susceptible to GML (Gram-positives) and bacteria that are typically resistant to GML (Gram-negative E. coli). However, we have shown that the reutericyclin-producing Enterococcus faecalis strain (Gram-positive) is resistant to GML22. Prior to addition of microbes to milk samples, the organisms were cultured overnight in Todd Hewitt (Difco, Detroit, MI) broths. The aerobic organisms were cultured at 37 °C with aeration (shaking at 200 RPM); C. perfringens was cultured stationary in an anaerobic chamber. Subsequently, each organism was diluted to the desired starting inoculum for addition to milk samples.
The superantigen TSST-1 was purified after culture of S. aureus RN6390 containing the tstH gene on plasmid pCE10747. TSST-1 was purified by ethanol precipitation from cultures followed by isoelectric focusing48,49. Toxin thus purified is homogeneous when tested by SDS-PAGE48,49.
Whole human and bovine milk and commercial infant formula samples
In all studies, for consistency, milk sample were pasteurized either by us or prior to their purchase from the commercial providers. Non-purchased samples were pasteurized by incubation at 65 °C for 30 minutes. Whole human milk samples were purchased from the Mother’s Milk Bank of Iowa, Coralville, IA. Whole bovine milk samples were obtained from cows at the dairy center at Mississippi State University by Dr. Keun Seok Seo. Commercially-available, previously pasteurized, bovine milk was purchased from HyVee Grocery Store, Coralville, IA. Infant formula was Similac Advance, also purchased from HyVee; this infant formula is a product of Abbott Laboratories, Columbus, OH. The formula was diluted two-fold with sterile distilled water as directed by the manufacturer. Milk fat was not removed from any samples, but instead was thoroughly mixed in the milk samples.
Glycerol monolaurate (GML)
Highly-pure, human, food grade GML was purchased from Colonial, South Pittsburgh, TN. The pellets were solubilized in absolute ethanol as a stock culture at 100 mg/ml for addition to milk samples. GML in milk samples was measured by mass spectrometry on a fee-for-service basis at the University of Iowa Metabolomics Center, compared to known standards. GML concentrations in milk samples were considered validated after addition of 3000 μg/ml of purified GML to bovine milk demonstrated of antimicrobial activity against S. aureus, and thus overcoming albumin’s ability to interfere with GML activity29. GML and other lipophilic compounds were removed from human milk samples by treatment with ethanol (80% final concentration) to precipitate milk molecules >10,000 molecular weight. The precipitates were dried and then resolubilized in distilled water to the original volume.
Human squamous epithelial cells
We used human squamous epithelial cells (HSECs) as a model system38 to test if human and bovine milk samples exhibit anti-inflammatory activity. These gently immortalized vaginal epithelial cells were cultured in triplicate to confluence in 96 well, flat-bottom microtiter plates in keratinocyte serum-free medium. Once confluent, the cells were treated for 6 hours in the absence and presence of 100 µg of the superantigen toxic shock syndrome toxin-1 (TSST-1) and with and without 20 µl of milk samples. The supernates were then tested for the pro-inflammatory chemokine IL-8 by Quantikine kit (R&D systems Minneapolis, MN). The experiment was repeated one additional time. Previously, we have shown that TSST-1 stimulates production of many pro-inflammatory cytokines by HSECs and keratinocytes, including MIP-3α and IL-8, whose genes may be up-regulated 500 and 80-fold, respectively38. Our studies were not designed to assess the effects of TSST-1 with and without milk samples on other innate and adaptive immune cells. However, our prior studies showed that a major effect of TSST-1 and SIV was on epithelial cells to produce harmful inflammation, these cells being present in many times the numbers of other immune cells19,20,22. Human milk samples alone were tested for resistance to killing by human milk through performing the 6 hour incubations with milk alone and then performing trypan blue dye exclusion. Finally, we tested all stationary phase (grown in Todd Hewitt) bacteria for stimulation of the epithelial cells and inhibition of IL-8 production by human milk in the same types of studies with use of 20 μl of washed bacteria (4000 × g, 10 minutes; resuspended in PBS)16,22.
Statistics
Means, standard deviations, and Student’s t test analyses were performed with Microsoft Excel 2010.
Acknowledgements
This research was supported by HHS National Institutes of Health research grants U19 AI117673 and NIAMS AR41256.
Author Contributions
P.M.S. conceived the studies, performed experimentation, and wrote and edited the manuscript. S.H.K. and K.S.S. performed experimentation and edited the manuscript. D.Y.M.L. conceived the studies and edited the manuscript.
Competing Interests
This research has been supported by grants from the National Institutes of Health awarded to the University of Iowa and National Jewish Health. The funders had no input in conceptualization, design, data collection, analysis, decision to publish, or preparation of the manuscript.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Slade HB, Schwartz SA. Mucosal immunity: the immunology of breast milk. J Allergy Clin Immunol. 1987;80:348–358. doi: 10.1016/0091-6749(87)90041-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lonnerdal B. Nutritional and physiologic significance of human milk proteins. Am J Clin Nutr. 2003;77:1537S–1543S. doi: 10.1093/ajcn/77.6.1537S. [DOI] [PubMed] [Google Scholar]
- 3.Murakami M, Dorschner RA, Stern LJ, Lin KH, Gallo RL. Expression and secretion of cathelicidin antimicrobial peptides in murine mammary glands and human milk. Pediatr Res. 2005;57:10–15. doi: 10.1203/01.PDR.0000148068.32201.50. [DOI] [PubMed] [Google Scholar]
- 4.Zhang F, et al. Antimicrobial activity and mechanism of the human milk-sourced peptide Casein201. Biochem Biophys Res Commun. 2017;485:698–704. doi: 10.1016/j.bbrc.2017.02.108. [DOI] [PubMed] [Google Scholar]
- 5.Desbois AP, Smith VJ. Antibacterial free fatty acids: activities, mechanisms of action and biotechnological potential. Applied microbiology and biotechnology. 2010;85:1629–1642. doi: 10.1007/s00253-009-2355-3. [DOI] [PubMed] [Google Scholar]
- 6.Martin R, et al. Human milk is a source of lactic acid bacteria for the infant gut. J Pediatr. 2003;143:754–758. doi: 10.1016/j.jpeds.2003.09.028. [DOI] [PubMed] [Google Scholar]
- 7.Fernandez L, et al. The human milk microbiota: origin and potential roles in health and disease. Pharmacol Res. 2013;69:1–10. doi: 10.1016/j.phrs.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 8.Ramani S, et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nature communications. 2018;9:5010. doi: 10.1038/s41467-018-07476-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stewart CJ, et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. 2018;562:583–588. doi: 10.1038/s41586-018-0617-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yu ZT, Chen C, Newburg DS. Utilization of major fucosylated and sialylated human milk oligosaccharides by isolated human gut microbes. Glycobiology. 2013;23:1281–1292. doi: 10.1093/glycob/cwt065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yu ZT, Nanthakumar NN, Newburg DS. The human milk oligosaccharide 2′-fucosyllactose quenches Campylobacter jejuni-induced inflammation in human epithelial cells HEp-2 and HT-29 and in mouse intestinal mucosa. J Nutr. 2016;146:1980–1990. doi: 10.3945/jn.116.230706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Newburg DS, Ko JS, Leone S, Nanthakumar NN. Human Milk Oligosaccharides and synthetic galactosyloligosaccharides contain 3’-, 4-, and 6’-galactosyllactose and attenuate inflammation in human T84, NCM-460, and H4 cells and intestinal tissue ex vivo. J Nutr. 2016;146:358–367. doi: 10.3945/jn.115.220749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rajani PS, Seppo AE, Jarvinen KM. Immunologically active components in human milk and development of atopic disease, with emphasis on food allergy, in the pediatric population. Front Pediatr. 2018;6:218. doi: 10.3389/fped.2018.00218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jones AL, Curran-Everett D, Leung DYM. Food allergy is associated with Staphylococcus aureus colonization in children with atopic dermatitis. J Allergy Clin Immunol. 2016;137:1247–1248 e1243. doi: 10.1016/j.jaci.2016.01.010. [DOI] [PubMed] [Google Scholar]
- 15.Simpson EL, et al. Patients with atopic dermatitis colonized with Staphylococcus aureus have a distinct phenotype and endotype. J Invest Dermatol. 2018;138:2224–2233. doi: 10.1016/j.jid.2018.03.1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schlievert PM, et al. Glycerol monolaurate does not alter rhesus macaque (Macaca mulatta) vaginal lactobacilli and is safe for chronic use. Antimicrob Agents Chemother. 2008;52:4448–4454. doi: 10.1128/AAC.00989-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Strandberg KL, et al. Glycerol monolaurate inhibits Candida and Gardnerella vaginalis in vitro and in vivo but not Lactobacillus. Antimicrob Agents Chemother. 2010;54:597–601. doi: 10.1128/AAC.01151-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Peterson ML, Schlievert PM. Glycerol monolaurate inhibits the effects of Gram-positive select agents on eukaryotic cells. Biochemistry. 2006;45:2387–2397. doi: 10.1021/bi051992u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li Q, et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature. 2009;458:1034–1038. doi: 10.1038/nature07831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Haase AT, et al. Glycerol monolaurate microbicide protection against repeat high-dose SIV vaginal challenge. PLoS One. 2015;10:e0129465. doi: 10.1371/journal.pone.0129465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schlievert PM, Peterson ML. Glycerol monolaurate antibacterial activity in broth and biofilm cultures. PLoS One. 2012;7:e40350. doi: 10.1371/journal.pone.0040350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brosnahan AJ, Merriman JA, Salgado-Pabón W, Ford B, Schlievert PM. Enterococcus faecalis inhibits superantigen toxic shock syndrome toxin-1-induced interleukin-8 from human vaginal epithelial cells through tetramic acids. PLoS One. 2013;8:e61255. doi: 10.1371/journal.pone.0061255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kaufmann GF, et al. Revisiting quorum sensing: Discovery of additional chemical and biological functions for 3-oxo-N-acylhomoserine lactones. Proc Natl Acad Sci USA. 2005;102:309–314. doi: 10.1073/pnas.0408639102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mu Q, Tavella VJ, Luo XM. Role of Lactobacillus reuteri in human health and diseases. Frontiers in microbiology. 2018;9:757. doi: 10.3389/fmicb.2018.00757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Strandberg KL, et al. Reduction in Staphylococcus aureus growth and exotoxin production and in vaginal interleukin 8 levels due to glycerol monolaurate in tampons. Clin Infect Dis. 2009;49:1711–1717. doi: 10.1086/644614. [DOI] [PubMed] [Google Scholar]
- 26.Witcher KJ, Novick RP, Schlievert PM. Modulation of immune cell proliferation by glycerol monolaurate. Clin Diagn Lab Immunol. 1996;3:10–13. doi: 10.1128/cdli.3.1.10-13.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang MS, Sandouk A, Houtman JC. Glycerol Monolaurate (GML) inhibits human T cell signaling and function by disrupting lipid dynamics. Scientific reports. 2016;6:30225. doi: 10.1038/srep30225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang Michael S., Tran Phuong M., Wolff Alexander J., Tremblay Mikaela M., Fosdick Micaela G., Houtman Jon C. D. Glycerol monolaurate induces filopodia formation by disrupting the association between LAT and SLP-76 microclusters. Science Signaling. 2018;11(528):eaam9095. doi: 10.1126/scisignal.aam9095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang MS, Houtman JC. Human serum albumin (HSA) suppresses the effects of glycerol monolaurate (GML) on human T cell activation and function. PLoS One. 2016;11:e0165083. doi: 10.1371/journal.pone.0165083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Brosnahan AJ, Schlievert PM. Gram-positive bacterial superantigen outside-in signaling causes toxic shock syndrome. Febs J. 2011;278:4649–4667. doi: 10.1111/j.1742-4658.2011.08151.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kokan NP, Bergdoll MS. Detection of low-enterotoxin-producing Staphylococcus aureus strains. Appl Environ Microbiol. 1987;53:2675–2676. doi: 10.1128/aem.53.11.2675-2676.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kotler DP, Sandkovsky U, Schlievert PM, Sordillo EM. Toxic shock-like syndrome associated with staphylococcal enterocolitis in an HIV-infected man. Clin Infect Dis. 2007;44:e121–123. doi: 10.1086/518286. [DOI] [PubMed] [Google Scholar]
- 33.Lin Z, Kotler DP, Schlievert PM, Sordillo EM. Staphylococcal enterocolitis: forgotten but not gone? Dig Dis Sci. 2010;55:1200–1207. doi: 10.1007/s10620-009-0886-1. [DOI] [PubMed] [Google Scholar]
- 34.Schlievert, P. M., Kilgore, S. H., Kaus, G. M., Ho, T. D. & Ellermeier, C. D. Glycerol monolaurate (GML) and a nonaqueous five-percent GML gel Kill Bacillus and Clostridium spores. mSphere3, 10.1128/mSphereDirect.00597-18 (2018). [DOI] [PMC free article] [PubMed]
- 35.Schlievert PM, Deringer JR, Kim MH, Projan SJ, Novick RP. Effect of glycerol monolaurate on bacterial growth and toxin production. Antimicrob Agents Chemother. 1992;36:626–631. doi: 10.1128/AAC.36.3.626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jenness R. The composition of human milk. Semin Perinatol. 1979;3:225–239. [PubMed] [Google Scholar]
- 37.Kasrae H, Amiri Farahani L, Yousefi P. Efficacy of topical application of human breast milk on atopic eczema healing among infants: a randomized clinical trial. Int J Dermatol. 2015;54:966–971. doi: 10.1111/ijd.12764. [DOI] [PubMed] [Google Scholar]
- 38.Peterson ML, et al. The innate immune system is activated by stimulation of vaginal epithelial cells with Staphylococcus aureus and toxic shock syndrome toxin 1. Infect Immun. 2005;73:2164–2174. doi: 10.1128/IAI.73.4.2164-2174.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Thursby E, Juge N. Introduction to the human gut microbiota. Biochem J. 2017;474:1823–1836. doi: 10.1042/BCJ20160510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rodriguez JM, et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb Ecol Health Dis. 2015;26:26050. doi: 10.3402/mehd.v26.26050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lin XB, et al. Genetic determinants of reutericyclin biosynthesis in Lactobacillus reuteri. Appl Environ Microbiol. 2015;81:2032–2041. doi: 10.1128/AEM.03691-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hess DJ, Henry-Stanley MJ, Wells CL. The natural surfactant glycerol monolaurate significantly reduces development of Staphylococcus aureus and Enterococcus faecalis iofilms. Surg Infect (Larchmt) 2015;16:538–542. doi: 10.1089/sur.2014.162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hess DJ, Henry-Stanley MJ, Wells CL. Antibacterial synergy of glycerol monolaurate and aminoglycosides in Staphylococcus aureus biofilms. Antimicrob Agents Chemother. 2014;58:6970–6973. doi: 10.1128/AAC.03672-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Desbois AP, Lang S, Gemmell CG, Coote PJ. Surface disinfection properties of the combination of an antimicrobial peptide, ranalexin, with an endopeptidase, lysostaphin, against methicillin-resistant Staphylococcus aureus (MRSA) J Appl Microbiol. 2010;108:723–730. doi: 10.1111/j.1365-2672.2009.04472.x. [DOI] [PubMed] [Google Scholar]
- 45.Kabara JJ, Swieczkowski DM, Conley AJ, Truant JP. Fatty acids and derivatives as antimicrobial agents. Antimicrob Agents Chemother. 1972;2:23–28. doi: 10.1128/AAC.2.1.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Carpo BG, Verallo-Rowell VM, Kabara J. Novel antibacterial activity of monolaurin compared with conventional antibiotics against organisms from skin infections: an in vitro study. Journal of drugs in dermatology: JDD. 2007;6:991–998. [PubMed] [Google Scholar]
- 47.Murray DL, et al. Localization of biologically important regions on toxic shock syndrome toxin 1. Infect Immun. 1996;64:371–374. doi: 10.1128/iai.64.1.371-374.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Blomster-Hautamaa DA, Kreiswirth BN, Novick RP, Schlievert PM. Resolution of highly purified toxic-shock syndrome toxin 1 into two distinct proteins by isoelectric focusing. Biochemistry. 1986;25:54–59. doi: 10.1021/bi00349a009. [DOI] [PubMed] [Google Scholar]
- 49.Blomster-Hautamaa DA, Schlievert PM. Preparation of toxic shock syndrome toxin-1. Methods Enzymol. 1988;165:37–43. doi: 10.1016/S0076-6879(88)65009-9. [DOI] [PubMed] [Google Scholar]