

Abstract
Segmentation is fundamental to the arthropod body plan. Understanding the evolutionary steps by which arthropods became segmented is being transformed by the integration of data from evolutionary developmental biology (evo-devo), Cambrian fossils that allow the stepwise acquisition of segmental characters to be traced in the arthropod stem-group, and the incorporation of fossils into an increasingly well-supported phylogenetic framework for extant arthropods based on genomic-scale datasets. Both evo-devo and palaeontology make novel predictions about the evolution of segmentation that serve as testable hypotheses for the other, complementary data source. Fossils underpin such hypotheses as arthropodization originating in a frontal appendage and then being co-opted into other segments, and segmentation of the endodermal midgut in the arthropod stem-group. Insights from development, such as tagmatization being associated with different modes of segment generation in different body regions, and a distinct patterning of the anterior head segments, are complemented by palaeontological evidence for the pattern of tagmatization during ontogeny of exceptionally preserved fossils. Fossil and developmental data together provide evidence for a short head in stem-group arthropods and the mechanism of its formation and retention. Future breakthroughs are expected from identification of molecular signatures of developmental innovations within a phylogenetic framework, and from a focus on later developmental stages to identify the differentiation of repeated units of different systems within segmental precursors.
Keywords: evo-devo, Arthropoda, palaeontology
1. Introduction
Questions about deep evolutionary history, such as the origin and evolution of animal body plans, demand a combined approach that takes into account multiple sources of evidence. These historical questions are often extremely complex, and are shrouded in the mists of deep time, so that no single approach provides enough information to fully reconstruct the underlying evolutionary events. At the very least, understanding deep evolution requires a sound phylogenetic framework, an adequate fossil record, and a mechanistic/developmental view of morphology [1]. Phylogenetics provides the framework within which we can infer the history of specific characters—how many times a feature evolved and on which branches of the tree of life. Palaeontology calibrates an evolutionary timescale, a putative sequence of events, and direct evidence of extinct morphologies and unique combinations of characters. Development provides insights into the processes by which form is generated, and about the possible transformation series between morphologies. How segmented body plans originated in arthropods exemplifies a question about deep evolutionary history, one for which progress has been informed by these diverse perspectives.
Arthropods are by far the most successful animal phylum and their success is largely an outcome of their segmented body. A segmentally arranged body allows for different measures of autonomy of individual segments or batches of segments, which is manifest in the varied tagmatization of the arthropod body and the ‘Swiss Army knife’-like specialization of appendages to perform diverse functions. This modularity in specialization has allowed arthropods to evolve and adapt to numerous environments and to specialize to myriad locomotory and feeding strategies. The success of arthropods is not confined to the present. Indeed, arthropods have probably been the most common and diverse animals throughout their evolutionary history [2], and fossils show that the segmented body plan evolved very early in arthropod evolution.
A segmented body is found in three phyla: Arthropoda, Chordata and Annelida, and evolved convergently in the three cases [3]. The segmental organization of the body in all three cases probably evolved in a stepwise fashion from an unsegmented ancestor, via recruitment of different metameric organ systems into synchronized repeated body units and the co-option of the development of these organ systems into unified embryonic segments [4]. Beyond this hypothetical scenario (figure 1), we have only patchy data about the evolutionary processes that led to the appearance of the segmented body in arthropods, or about the intermediate stages in this presumed stepwise process. Nonetheless, we can learn about the evolution of the segmented body by understanding the evolution of the developmental process that underlies it—the segmentation process.
Figure 1.

A scenario for the origin and evolution of arthropod segmentation (based on [4]). (a) A simple bilaterian with a short body; (b) extension of the anterior–posterior axis (A-P); (c) several organ systems become independently metameric and distributed along the A-P axis; (d) metamerism of the different systems becomes synchronized; (e) elements of all metameric systems are generated together from an undifferentiated segmental precursor. (Online version in colour.)
2. Evo-devo and the fossil record
Evolutionary developmental biology (evo-devo) is the discipline that focuses on tracing changes in developmental processes over time, through a tree-based comparison of development in different organisms. This approach allows us to pinpoint where in phylogeny specific developmental characters first evolved or where they were modified. This provides insight into the evolution of morphological characters that are under selection, and about the mechanisms behind the generation of the characters and how they can be modified through changes in the developmental process. By contrast, the fossil record provides information about the temporal context of morphological changes; minimum dates for when specific morphologies appeared and in what sequence; while also providing examples of specific character combinations and intermediate character states that are not found in extant species. There is a complementarity between one approach that looks at a single moment in time (the present), but allows mechanistic analyses of developmental processes, and an approach that gives a much deeper temporal perspective but is limited by the unpredictable preservation of a sample of body plans, and of course does not allow experimentation.
Integration of evo-devo approaches and fossil data has been a productive approach in the vertebrate world for years [5–7]. Many of the fundamentals of evo-devo have, however, been established using arthropods [8–14], and this early work was instrumental in consolidating the evo-devo approach as an independent evolutionary discipline. There has been some effort to integrate evo-devo and palaeontological perspectives on the origin of various arthropod features (e.g. insect wings [15,16], head segmentation [17], appendages [18] and trunk segmentation [19]), but this approach remains under-used in the world of arthropod evolutionary biology. With respect to the specific question we are addressing here—the origin of segmental features—palaeontological effort is concentred in the Cambrian, that being the time period when arthropods first appear in the fossil record and when stem-group arthropods are best represented.
The quality of early arthropod fossils and their interpretation have steadily improved via the documentation of Cambrian fossils from sites of exceptional preservation [2], which continue to be discovered [20]. These fossils underpin current hypotheses about how a grade of taxa that constitutes the arthropod stem-group reveals the stepwise acquisition of characters, many related to segmentation. Burgess Shale-type fossils—two-dimensional carbonaceous compressions of the cuticle coupled with mineralization of labile tissue such as the gut and muscle—have been the focus of much research, but have been supplemented by finds from other styles of fossilization (e.g. small carbonaceous fossils [21] and secondarily phosphatized ‘Orsten’ fossils, including larvae [22]). The latter are especially important for preserving series of developmental stages that bridge gaps between embryos (which underpin much evo-devo research) and adult stages, which predominate among Burgess Shale-type fossils [23,24].
In parallel, the representation of species and taxa in the experimental world has increased substantially. This broader selection of species covers a much wider and more representative sample of extant morphological diversity within arthropods than that represented by the very small sample of model species available until only 15 years ago (e.g. hemimetabolous insects [25,26]; diverse arachnids [27–30] and dipterans [31–34]; non-hexapod pancrustaceans: [35–37]; myriapods [38,39]; and many others). This broader range of model species has allowed the addressing of specific evolutionary questions, such as the origin of different respiratory organs [30], the origin of wings [40,41] and the diversity of limbs [18]. Beyond the increase in experimentally tractable model organisms, the explosion of available genomes [42] makes it possible to use bioinformatic approaches to look for genetic and regulatory novelties, and makes it easier to plan and develop functional work on an even larger sample of arthropod diversity. With regards to segmentation in particular, emerging model arthropods have shed light on the diversity of segment generation modes and the evolution of the segmentation process in insects and other arthropods [43–49].
The phylogenetic relationships of extant arthropods used as models in evo-devo are resolved with considerable confidence based on genome scale data (figure 2 top, reviewed by Giribet & Edgecombe [50]). High-level insect phylogeny has a well-supported, morphologically coherent framework based on transcriptomic analyses [51], the pattern of ‘crustacean’ paraphyly with respect to Hexapoda has numerous nodes that are repeatedly recovered with strong support in phylogenomic studies [52,53], and myriapod phylogeny has settled on higher-level clades that reflect groups long recognized based on morphology and development [54,55]. Chelicerate phylogeny remains more ambiguous, but some key nodes bracketing evo-devo models (such as spiders and scorpions being more closely related to each other than either is to mites, i.e. the Arachnopulmonata hypothesis) are robustly supported [56,57].
Figure 2.

Phylogenetic relationships among key experimental model species of Panarthropoda and representative fossil taxa. Names of terminals at the top of the tree are extant genera of experimentally tractable model species. Names of terminals deep within the tree are fossil forms representing stem-groups of various extant taxa. Node names refer to crown-groups. Placement of some of the fossil taxa is still contentious (see text for examples).
The phylogenetic position of fossils on the other hand is often contentious. Over a decade or so, a picture had emerged of the branching sequence in the arthropod stem-group (figure 2; reviewed by Daley et al. [58]), a grade that encompasses a transition from vermiform, lobopod-bearing forms in the deepest nodes of the arthropod stem-group to fully arthrodized and arthropodized forms more proximal to the crown-group. Because it samples morphologies between the most recent common ancestor of Onychophora (which lack segmental sclerites) and extant diversity of Arthropoda, this grade is of particular relevance to understanding the evolution of arthropod segmentation. Some taxa are consistently recovered in the arthropod stem-group in all recent phylogenetic analyses, such as Isoxys and Radiodonta (the clade including Anomalocaris and its relatives), but a few groups that had been assigned to the arthropod stem-group have been reinterpreted as within the arthropod crown-group. This affects some fossil clades that had played a significant role in resolving character acquisition in the arthropod stem-group, such as fuxianhuiids and bivalved hymenocarine arthropods like Canadaspis and Branchiocaris [59,60]. Different interpretations of morphology, especially with regards to mouthpart differentiation, together with alternative character sets and tree reconstruction methods, have prompted a shift of fuxianhuiids and Hymenocarina into the arthropod crown-group, either within or allied to Mandibulata [61,62].
A combined approach, including both the fossil record and experimental data on extant animals, within a unified phylogenetic framework (figure 2) provides the most complete possible picture of the evolution of the arthropod segmented body. The fossil record contributes data about the sequence of character acquisition, most informatively when characters that are clustered at a single node in extant taxa are scattered across multiple nodes in a stem-group. Furthermore, because fossils preserve unique combinations of characters, some of which may be lost in extant taxa, they can serve as the basis for novel hypotheses on the stepwise assembly of the developmental programme that generates segments in extant taxa. Conversely, development provides mechanistic examples of how characters are transformed, which allow linking character states in fossil taxa and suggest putative homologies that are not obvious from the fossils alone. We provide examples of these reciprocities and synergies in the following.
3. Synergistic hypotheses
(a). Arthropodization evolved in the frontal appendage and was co-opted to trunk appendages
The deepest branches of the arthropod stem-group are represented by large-bodied Cambrian lobopodians such as Jianshanopodia [63], Megadictyon [64] and Siberion [65]. These have annulated trunks with a homonomous series of annulated lobopods. The anteriormost appendage pair is specialized as an enlarged raptorial pair of limbs bearing strong, elongate spines along their inner margins, but like the trunk appendages they too are annulated. An annulated, spinose frontal appendage is shared by the so-called gilled lobopodians—exemplified by the early Cambrian Kerygmachela (figure 3a) and Pambdelurion—that are broadly agreed to branch more crownward of the giant lobopodians in the arthropod stem-group. Traces of neural tissue innervating the frontal appendage of Kerygmachela have been interpreted as nerves derived from the first brain ganglion—the protocerebrum [67].
Figure 3.

Annulated (a) and arthropodized (b–e) frontal appendages (fa). (a) The ‘gilled lobopodian’ Kerygmachela kierkegaardi; (b–e) disarticulated Anomalocaris canadensis frontal appendages. Scales: (a–c) 10 mm, (d–e) 5 mm. (a) courtesy of J. Vinther; (b–e) courtesy of A. Daley (from [66]). (Online version in colour.)
The origin of fully arthropodized appendages in the arthropod stem-group can be linked to the group known as Radiodonta, exemplified by the famous Anomalocaris. Radiodonts have an arthropodized frontal appendage, in most members of the group this being the only conspicuously arthropodized appendage in the body. Various theories interpret other segmental body parts, notably a fringe of dorsal setal blades and series of paired ventral flaps, as appendicular derivatives, homologues of the rami of biramous appendages in arthropods [68,69], but the frontal appendage most clearly depicts apomorphies of arthropodization. The radiodont frontal appendage has discrete sclerotized articles/podomeres that are articulated to each other on their dorsal side at hinges and are separated from each other on their ventral sides by desclerotized triangular, telescoping fields of arthrodial membrane [66] (figure 3b–e). Most podomeres bear ventral spines, corresponding to endites in many crown- (and stem-) group arthropods, and a few distal podomeres bear dorsal spines as well.
Extant panarthropods present two character states, each corresponding to one of the two states observed in frontal appendages in the arthropod stem-group: fully arthropodized limbs with a more or less conserved array of elements, and the annulated, lobopodial limbs of onychophorans and tardigrades. The network patterning the arthropodized limb is conserved to a very high degree among all arthropods [18,70]. The gap genes that pattern the proximo-distal axis of arthropod limbs are expressed in the same register in onychophorans [71,72], demonstrating that these genes have a conserved role in limb development in panarthropods that predates arthropodization (figure 4). Thus, it is possible to reconstruct the gene regulatory networks (GRNs) that were instrumental in the evolution of panarthropod limbs and in their subsequent arthropodization.
Figure 4.

Schematic of gap gene expression domains in limbs of onychophorans and in the different arthropod classes. The proximal side of the limb is to the left. Lighter colours represent lower expression levels or downregulation during development. Redrawn with a revised phylogeny from [70]. See source for references. (Online version in colour.)
Radiodonts and more crownward taxa in the arthropod stem-group demonstrate that arthropodization did not happen in a piecemeal manner (e.g. one segment at a time posteriorly from an origin in the protocerebral segment). Rather, it was co-opted by the rest of the head and trunk segments simultaneously [18] and, notably, was abandoned in the protocerebral segment (acknowledging a prolonged debate about whether the labrum is or is not appendicular and specifically a transformed protocerebral appendage; reviewed by Ortega-Hernández et al. [17] and Jockusch [18]). Arthropods immediately crownward of radiodonts in the arthropod stem-group, such as isoxyids, display arthropodization of all head and trunk limbs, presumed to be a shared derived character retained by crown-group arthropods. This is the condition in Deuteropoda (sensu [73]), a monophyletic group united by a deutocerebral first appendage (antenna or chelicera), encompassing all crown-group arthropods and a set of stem-group taxa.
The detailed similarity in the structure of the arthropodized frontal appendage in radiodonts to the arthropodized trunk limbs in more crownward groups suggests that these are serially homologous structures. Homology implies that they are underlain by the same GRN or character identity network (CHiN, [74]), even though they are found on positionally non-equivalent segments. If this is indeed the case, then the modified limb patterning GRN (which allows arthropodization of the limb) was co-opted from the frontal appendage to other appendages. The concept of co-option is a central tenet in the theory of novelty within the evo-devo conceptual framework [75–78]. According to the accepted paradigm, novel structures can arise through the recruitment of existing GRNs to novel locations. For this to happen, all that is required is for a key gene, high in the GRN's hierarchy, to be activated ectopically, and this will suffice for the activation of the entire network and the generation of a complete structure in a new location, in essence creating an evolutionary novelty.
(b). The evolution of tagmatization may be linked to changes in the developmental programme of segment generation
Arthropod tagmata are fundamentally defined by differentiation of segmental structures, such as differential presence or absence of appendages, or differences in the structure of batches of appendages. Evo-devo research has now elucidated many of the mechanisms responsible for the presence or the absence of appendages on specific segments—characteristic for different tagmata. Dll is generally considered to be the main signal for the initiation of the limb development programme [5,79]. The identity of the segments and the identity of the appendages they bear (if any) are mostly under the control of Hox genes, often directly regulating Dll [80], although it is clear that this is only a partial description of the tagma differentiation process. In most cases, Hox gene expression defines broad domains along the anterior–posterior (A-P) axis, but does not define the borders between tagmata [81].
The fossil record predicts that the plesiomorphic state (post-recruitment of arthropodized limbs to the trunk) is a series of more or less homonomous appendages on all post-cephalic segments. However, most arthropod lineages show examples of segment specialization and loss of appendages in some segments. The best-known example is the differentiation of the extant insect body into head, thorax and abdomen, but tagmatization is also well known from extinct groups. Fuxianhuiids, for example, have appendages on the anterior part of the trunk but have a limbless batch of segments in the posterior part of the trunk [82], analogous to the abdomen of extant arthropods (e.g. hexapods). Complex patterns of limb loss and regional differentiation are known in stem- and crown-group chelicerates [83].
Some arthropods display different ontogenetic modes of segment generation in different body regions. In some cases, these differences correlate fully with different tagmata. This is best studied in insects, specifically in the milkweed bug Oncopeltus fasciatus, in which thoracic segments are generated simultaneously, whereas abdominal segments are generated sequentially [48]. While this question has not been addressed specifically in other insects, expression data from the cricket Gryllus bimaculatus [84] suggest this may be a common pattern, at least in hemimetabolous insects. This pattern is not seen in Holometabola, which could indicate a secondary loss of region-specific segmentation modes. In Myriapoda, the head segments are formed separately and not in temporal sequence along the A-P axis, in contrast with the trunk segments, which form in strict A-P sequence [46]. Spiders have a different prosomal and opisthosomal segmentation process [85,86] and in the mite Tetranychus the pair-rule orthologue Pax-3/7 is expressed only in the prosoma [87]. Ontogenetic data for fossil taxa are rare, but there are at least some cases that conform to this pattern. The ontogeny of Fuxianhuia protensa shows that tergites form at a terminal growth zone, and moults involving tergite addition alternate with ones in which a limbless abdominal segment is shed into the appendage-bearing thorax [82]. If fuxianhuiids are stem-group arthropods, as is widely accepted, an association between tagmosis (including presence versus absence of appendages on tagmata) and different modes of segment generation predates the origin of the arthropod crown-group. Thus, we would predict that different modes of segment generation within a single species existed very early in arthropod evolution.
(c). The ancestral crown-group arthropod head was composed of few segments
The fossil record shows that the ancestral arthropod head was composed of fewer segments than that of extant arthropods. Deeply diverging stem-group euarthropods (lobopodians such as Megadictyon and Jianshanopodia and the gilled lobopodians Kerygmachela and Pambdelurion) had a single-segment head, the protocerebral segment bearing the sole appendage pair, a frontal appendage [17,67] (figure 3a). This situation of a head composed of a single protocerebral segment is still found in tardigrades [88,89]. More crownward of these, Radiodonta for the most part correspond to this level of organization, but the number of head segments in this clade is ambiguous because of whether or not three segments associated with small flaps and in some cases paired gnathobase-like structures [90] are a part of the head or the trunk. Taxa closer to the euarthropod crown-group, such as Fuxianhuia [91], have three head segments, in this case being ocular, an antennule and a so-called ‘SPA’—a specialized post-antennal appendage. On this basis, it has been proposed that a three-segmented head in Fuxianhuia corresponds to a three-segmented brain as an ancestral state for Deuteropoda [17], composed of proto-, deuto- and tritocerebral segments. We caution, however, that the head segmentation of Fuxianhuia is under dispute, with evidence that taxa once viewed as having a comparable three-segmented head, such as Branchiocaris [92], later being shown to have mandibles and post-mandibular mouthparts [61].
Evo-devo work shows that there is a distinct difference between the way in which the anterior three segments (the pre-gnathal segments) are patterned in all extant arthropods relative to more posterior segments (figure 5). This is true in insects [48,95,96] and myriapods [93], which have a six-segmented head, and in chelicerates [94], which have a complex cephalothorax (prosoma) rather than a distinct head. Differences include a lack of involvement of pair-rule genes and a different regulation of segment polarity genes in the anterior segments (known from Drosophila [97], reviewed in [96]), lack of Hox gene expression in the anterior two segments [80], and an unusual mode of segment generation, which involves segmental genes being expressed in single stripes or patches in the pre-gnathal domain and then splitting to give two or three stripes that correspond to individual segments [48,93,94]. There is also some evidence for differences in the expression of segment polarity genes in these anterior segments in several species, but this has not been analysed in sufficient detail.
Figure 5.
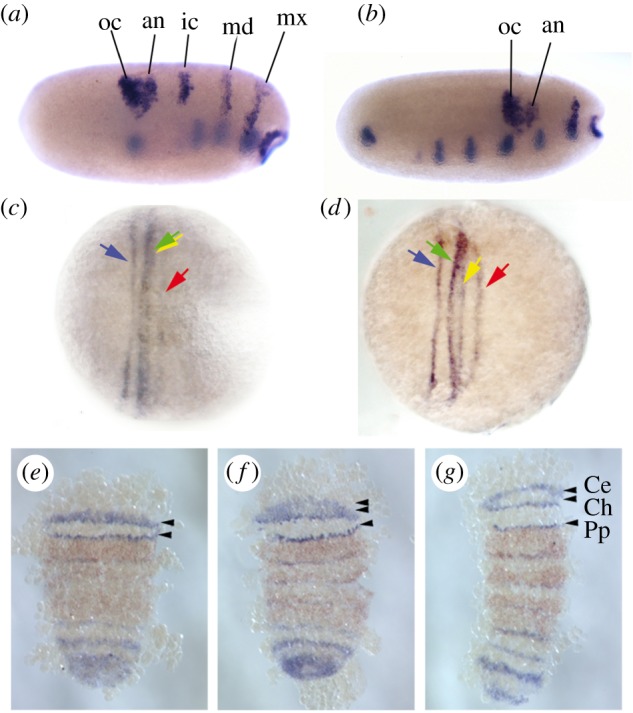
Examples of anterior segmentation through ‘stripe-splitting’ in different arthropod taxa. (a,b) Blastoderm stages of the milkweed bug Oncopeltus fasciatus (Insecta). An anterior patch of wingless expression representing both the ocular (oc) and antennal (an) segment splits to give rise to two distinct expression domains. (c,d) Early development of the centipede Strigamia maritima (Myriapoda), showing the expression of hedgehog. The anterior expression stripe representing the ocular segment (blue arrow) splits first from a preliminary single stripe. This is followed by the separation of the antennal (green arrow) and intercalary (yellow arrow) segmental stripes. The expression stripe representing the mandibular segment (red arrow) appears independently. (e–g) Early germband embryo of the spider Parasteatoda tepidariorum (Chelicerata), showing expression of hedgehog (blue stain) and deformed (orange stain). The anterior expression of hedgehog starts as a single stripe, which splits once to give a stripe representing the pedipalpal segment (Pp) and a second stripe that soon splits again to give the stripes representing the ocular segment (Ce, cephalic lobe) and the cheliceral segment (Ch). Adapted (a,b) from [48], (c,d) from [93] and (e–g) from [94]. (Online version in colour.)
We suggest that the unusual three anterior segments in extant arthropods represent the ancestral three-segment head found in Cambrian taxa that are near or within the arthropod crown-group. Under this hypothesis, the ancestral head segments were already developmentally distinct from the trunk segments at the time of their first appearance, and this difference has been maintained throughout their evolutionary history. It is still seen in all extant arthropods, as well as in Cambrian larvae that provide evidence for simultaneously developing head segments (a so-called ‘head larva’) being an ancestral character of crown-group arthropods [23]. This interpretation, which relies on combined data from fossils and embryos, provides a developmental-mechanistic scenario for the evolution of the three-segmented head from an ancestral single-segment head.
(d). The segmental body of stem-group arthropods included endodermal segmentation
Metameric gut diverticula are a common character in many stem-group arthropods. Based on the typical restriction of these diverticula to the posterior part of the head and the anterior part of the trunk (or much of the length of the trunk) versus a tube-shaped gut more anteriorly and posteriorly, the diverticula provide a basis for distinguishing the midgut from the foregut and hindgut in fossils. The midgut was strongly segmented in the arthropod stem-group (figure 6 [66,98]), including in the ‘gilled lobopodians’ (figure 6a,b) and isoxyids (figure 6c), as well as in some groups that have been interpreted as either stem-group or crown-group arthropods, such as megacheiran ‘great appendage arthropods' (figure 6d) and trilobitomorphs (figure 6e,f). Throughout this evolutionary grade, the morphology of the midgut diverticula is conserved, being relatively large, reniform organs with a distinctive radiating canal system, underpinning their homology. This hypothesis that the gut was segmented at the origin of arthropods (the diverticula being present in giant lobopodians such as Jianshanopodia [98]) could only be formulated based on information from fossils, because gut segmentation is almost non-existent in extant arthropods. An exception to this are remipedes, which are unique in having paired segmental midgut diverticula, up to 32 pairs along the length of the midgut [98]. However, the deeply nested systematic position of remipedes within Pancrustacea (as sister group of hexapods) negates the possibility that their serially repeated midgut diverticula are a plesiomorphy retained from the arthropod stem-group.
Figure 6.

Segmental midgut diverticula (mgd) in Cambrian total-group Arthropoda. (a,b) The ‘gilled lobopodian’ Pambdelurion whittingtoni, scales 20 mm; (c) the isoxyid Isoxys acutangulus, scale 5 mm; (d) the megacheiran Leanchoilia superlata, scale 5 mm; (e,f) the trilobitomorph Kuamaia lata: left side of thorax, showing exopod flap (ef,) lamellar setae (ls) and diverticula on three segments, scales: (e) 2 mm and (f), 5 mm. (Online version in colour.)
We cannot directly study the developmental origin of midgut segmentation, as we have no extant examples that are experimentally tractable. However, we can gain insights from what we know about germ layers in segmentation of arthropod laboratory models. All other metameric organ systems originate from undifferentiated mesoderm and ectoderm segmental precursors, which do not normally contribute to endodermal structures at all [99,100]. It is possible that the endodermal structures are ‘entrained’ to ectodermal or mesodermal structures, and thus their segmentation is secondary. This is in agreement with the model presented above for the evolution of segmental integration [4], which suggests that metamerism in different structures may have evolved independently and was secondarily integrated. Midgut diverticula could be an example of an individually metameric organ system that was not recruited into the main segmentation process and was thus more easily lost.
(e). Early segmental characters can be mismatched along the body axis
Palaeontology provides novel evidence that in some fossil taxa dorsal ectodermal and endodermal structures (tergites and midgut diverticula, respectively) are matched segmentally, but they are out of synchronization with the ganglia of the ventral nerve cord and limbs (which are in turn in segmentally matched to each other). This is evidenced by the distribution of midgut diverticula in the fuxianhiiid Fuxianhuia protensa, which has one pair of diverticula matched to each of its anterior, appendage-bearing trunk segments [101]. It has long been known that fuxianhuiids have many more trunk appendages than tergites [91], and the discovery of the ventral nerve cord in the trunk of the fuxianhuiid Chengjiangocaris kunmingensis showed that ganglia of the nerve cord correspond to the distribution of appendages [102]. The regular, alternating pattern of terminal tergite addition and shedding of abdominal tergites into the thorax in Fuxianhuia, described above, has been interpreted as tergal formation corresponding to a general/primitive arthropod pattern versus a decoupled and phylogenetically derived mode of appendage development [82]. Fuxianhuiids are thus potentially relevant to discussion about dorsoventral decoupling in arthropods more generally [101], or lack of integration of different organ systems into one segmentation system. We must clarify that regardless of fuxianhuiids' exact position (in the arthropod crown-group or in the stem), they probably lost integration rather than displaying an ancestral unintegrated state. This type of mismatch has taken place several times throughout arthropod evolution, and is found both in fossil and extant taxa [103].
These observations about character covariation in fossils could inspire experiments that look at genetic linkages between segmentation mechanisms in the tergites and gut systems and at the possible role of re-segmentation in defining the position of different structures in (extant) model systems [104]. From the perspective of evo-devo, aspects of dorsoventral decoupling have been explored in some detail in millipedes, particularly using Glomeris marginata as a model for gene expression [39,105]. At the level of segment polarity gene expression, the ventral side of the embryo (e.g. sternites and legs) in G. marginata corresponds to patterns seen in other arthropods, whereas dorsal segmentation deviates in many respects. Dorsoventral developmental mismatch is also known from the branchiopod Triops [106] but in much less detail. Dorsal and ventral decoupling of segmentation is manifest even at the level of delimiting the posterior boundary of the head, which conspicuously differs in various groups (reviewed in [107]). The adult dorsoventral axis is represented in the germ band as a medio-lateral axis [108], and the mismatch can easily take place through changes in patterning during the germband stage. A comparative developmental approach to patterning along the dorsoventral axis of the germ band, with the level of segmental mismatch known from the fossil record as a reference point, should be highly informative for understanding the conservation or lability of developmental integration among the units that make up a segment.
4. Discussion
There are groups for which evo-devo gives us an understanding of segmental mechanisms (such as Hexapoda—informed by such models as Drosophila melanogaster and Tribolium castaneum) but palaeontology provides limited insights because key regions of tree space (such as the hexapod stem-group) are essentially unsampled. Conversely, there are other groups for which palaeontology gives us insights into the acquisition of segmental features, such as the arthropod stem-group (examples above) and the chelicerate stem-group, but evo-devo is challenged because extant taxa with features of interest are separated from their closest living relative by long branches, i.e. they have been subjected to so many evolutionary steps that understanding character evolution is difficult. Fossil chelicerates, for example, provide evidence of unique patterns of tagmosis that are not retained by extant forms, yet are critical to inferring ancestral states. Thus, stem-group ‘synziphosurine’ euchelicerates depict a combination of a shield-like prosoma, an articulated set of opisthosomal tergites and a tail spine [83].
As outlined in the examples above, the fossil record gives us a ‘search image’ for features to look for in comparative embryology, e.g. endodermal segmental characters force a rethink of how segments could have been generated ancestrally. It provides a possible explanation for patterns seen in development, such as pre-gnathal segments. Fossils also supply direct ontogenetic data for parts of tree space that are not represented by living taxa. Wholly extinct groups like trilobites, for example, allow for quantitative tree-based analysis of modes of trunk segmentation [109]. Lastly, palaeontology informs on rates of evolution of features of interest from an evo-devo perspective. This is most powerfully understood in the context of molecular time-trees calibrated by fossils, which constrain the origin of many segmental traits in arthropods to a window of time spanning the terminal Ediacaran and the Terreneuvian Series of the Cambrian [51,58,110–112]. Conversely, evo-devo provides mechanistic explanations for phenomena we observe in the fossils. It also raises hypotheses (e.g. germband reconstructions [108]) about the possible sequence of evolutionary events, which can potentially be corroborated by the fossil record, e.g. gradual integration of metameric systems into coherent segments. Evo-devo also provides linkages between different characters through identification of gene pleiotropies and developmental integration—neither of which are obvious from fossils or from general morphology. The two disciplines operate under different conceptual frameworks. These are partly overlapping (notably, both are fundamentally rooted in tree thinking), but each has much to learn from the other.
Looking forward, many of the conflicts in recent phylogenetic trees aiming to place early derived fossil arthropods stem from differing interpretations of fossilized morphology. Burgess Shale-type fossils, as exquisite as they can be, are afflicted by taphonomy (decay, compaction, fragmentation, patchy mineralization, etc.), and teasing apart original anatomy and taphonomic artefacts is often complicated. In spite of this, improved imaging tools are allowing palaeontologists to extract more information from fossils. Arthropod compression fossils have for some years been productively investigated using polarized light and/or immersion in liquid [113], but have more recently provided additional data using such approaches as fluorescence microscopy [114], laminography [115] and computed micro-tomography [116]. These techniques may resolve some of the current controversies about homologies in fossil taxa that complicate character coding.
The future also holds the prospect of genomic sequences as an additional source of information for understanding the evolution of morphology. A continued improvement in taxon sampling and an increased quality of genome sequencing and genome annotation will allow us to look for molecular signatures of developmental innovations within a phylogenetic framework. This will make it easier to generalize from the small sample of experimentally tractable species to species in under-sampled regions of the tree, and to possibly extrapolate to extinct taxa using our understanding of development from extant diversity.
Finally, a glaring gap in current work on the evo-devo of segmentation can be potentially closed by bringing in insights from the fossil record. Most work on the development of segments focuses on the molecular processes leading to the undifferentiated segmental precursors. In terms of the segmental organization of the body, it is hardly an exaggeration to say that everything important has already happened by the germband stage and the expression of segment polarity genes. However, the morphology that is under selection, and which we see varying in the diversity of arthropods and their extinct relatives, is manifested later. The differentiation of repeated units of different systems within the segmental precursor has hardly been explored. This significant gap can be closed by focusing attention on later stages in development. A research agenda aimed at understanding the later stages and their diversity that takes advantage of genomic datasets and that integrates with the fossil record, which is for the most part post-embryonic, has the potential to paint a much more complete and nuanced picture of the evolution of the arthropod segmented body plan.
Supplementary Material
Acknowledgements
For fossil images, we thank Allison Daley, Diego García-Bellido, Jakob Vinther and the Royal Ontario Museum. For embryo images, we thank Michael Akam and Hiroki Oda.
Data accessibility
This article has no additional data.
Authors' contributions
A.D.C. and G.D.E. conceived the study, wrote the manuscript and prepared the figures.
Competing interests
We declare no competing interests.
Funding
We received no funding for this study.
References
- 1.Chipman AD, Erwin DH. 2017. The evolution of arthropod body plans: integrating phylogeny, fossils, and development—an introduction to the symposium. Integr. Comp. Biol. 57, 450–454. ( 10.1093/icb/icx094) [DOI] [PubMed] [Google Scholar]
- 2.Edgecombe GD, Legg DA. 2013. The arthropod fossil record. In Arthropod biology and evolution (eds Minelli A, Boxshall G, Fusco G), pp. 393–415. Berlin, Germany: Springer. [Google Scholar]
- 3.Chipman AD. 2010. Parallel evolution of segmentation by co-option of ancestral gene regulatory networks. Bioessays 32, 60–70. ( 10.1002/bies.200900130) [DOI] [PubMed] [Google Scholar]
- 4.Chipman AD. 2019. Becoming segmented. In Perspectives on evolutionary developmental biology (ed. Fusco G.), pp. 235–244. Padova, Italy: Padova University Press. [Google Scholar]
- 5.Shubin N, Tabin C, Carroll S. 1997. Fossils, genes and the evolution of animal limbs. Nature 388, 639–648. ( 10.1038/41710) [DOI] [PubMed] [Google Scholar]
- 6.Coates MI. 1994. The origin of vertebrate limbs. Dev. Suppl. 1994, 169–180. [PubMed] [Google Scholar]
- 7.Coates MI, Cohn MJ. 1998. Fins, limbs and tails: outgrowths and axial patterning in vertebrate evolution. Bioessays 20, 371–381. ( 10.1002/(SICI)1521-1878(199805)20:5<371::AID-BIES4>3.0.CO;2-R) [DOI] [Google Scholar]
- 8.Patel NH, Kornberg TB, Goodman CS. 1989. Expression of engrailed during segmentation in grasshopper and crayfish. Development 107, 201–212. [DOI] [PubMed] [Google Scholar]
- 9.Patel NH, Ball EE, Goodman CS. 1992. Changing role of even-skipped during the evolution of insect pattern formation. Nature 357, 339–342. ( 10.1038/357339a0) [DOI] [PubMed] [Google Scholar]
- 10.Patel NH. 1994. The evolution of arthropod segmentation: insights from comparisons of gene expression patterns. Dev. Suppl. 1994, 201–207. ( 10.1016/0168-9525(89)90183-2) [DOI] [PubMed] [Google Scholar]
- 11.Akam M, Averof M, Castelli-Gair J, Dawes R, Falciani F, Ferrier D. 1994. The evolving role of Hox genes in arthropods. Dev. Suppl. 1994, 209–215. [PubMed] [Google Scholar]
- 12.Akam M. 1995. Hox genes and the evolution of diverse body plans. Phil. Trans. R. Soc. Lond. B 349, 313–319. ( 10.1098/rstb.1995.0119) [DOI] [PubMed] [Google Scholar]
- 13.Rogers BT, Kaufman TC. 1997. Structure of the insect head in ontogeny and phylogeny: a view from Drosophila. Int. Rev. Cytol. 174, 1–84. ( 10.1016/S0074-7696(08)62115-4) [DOI] [PubMed] [Google Scholar]
- 14.Rogers BT, Peterson MD, Kaufman TC. 1997. Evolution of the insect body plan as revealed by the Sex combs reduced expression pattern. Development 124, 149–157. [DOI] [PubMed] [Google Scholar]
- 15.Engel MS, Davis SR, Prokop J. 2013. Insect wings: the evolutionary development of nature's first fliers. In Arthropod biology and evolution (eds Minelli A, Boxshall G, Fusco G), pp. 269–298. Berlin, Germany: Springer. [Google Scholar]
- 16.Prokop J, Pecharova M, Nel A, Hornschemeyer T, Krzeminska E, Krzeminski W, Engel MS. 2017. Paleozoic nymphal wing pads support dual model of insect wing origins. Curr. Biol. 27, 263–269. ( 10.1016/j.cub.2016.11.021) [DOI] [PubMed] [Google Scholar]
- 17.Ortega-Hernández J, Janssen R, Budd GE. 2017. Origin and evolution of the panarthropod head: a palaeobiological and developmental perspective. Arthropod. Struct. Dev. 46, 354–379. ( 10.1016/j.asd.2016.10.011) [DOI] [PubMed] [Google Scholar]
- 18.Jockusch EL. 2017. Developmental and evolutionary perspectives on the origin and diversification of arthropod appendages. Integr. Comp. Biol. 57, 533–545. ( 10.1093/icb/icx063) [DOI] [PubMed] [Google Scholar]
- 19.Hughes NC, Hong PS, Hou JB, Fusco G. 2017. The development of the Silurian trilobite Aulacopleura koninckii reconstructed by applying inferred growth and segmentation dynamics: a case study in paleo-evo-devo. Front. Ecol. Evol. 5, 37 ( 10.3389/fevo.2017.00037) [DOI] [Google Scholar]
- 20.Fu DJ, et al. 2019. The Qingjiang biota-a Burgess Shale-type fossil Lagerstatte from the early Cambrian of South China. Science 363, 1338–1342. ( 10.1126/science.aau8800) [DOI] [PubMed] [Google Scholar]
- 21.Butterfield NJ, Harvey THP. 2012. Small carbonaceous fossils (SCFs): a new measure of early Paleozoic paleobiology. Geol. 40, 71–74. ( 10.1130/G32580.1) [DOI] [Google Scholar]
- 22.Maas A, et al. 2006. The ‘Orsten’—more than a Cambrian Konservat-Lagerstätte yielding exceptional preservation. Palaeoworld 15, 266–282. ( 10.1016/j.palwor.2006.10.005) [DOI] [Google Scholar]
- 23.Waloszek D, Maas A. 2005. The evolutionary history of crustacean segmentation: a fossil-based perspective. Evol. Dev. 7, 515–527. ( 10.1111/j.1525-142X.2005.05056.x) [DOI] [PubMed] [Google Scholar]
- 24.Haug JT, Maas A, Waloszek D. 2010. Henningsmoenicaris scutula, Sandtorpia vestrogothiensis gen. et sp nov and heterochronic events in early crustacean evolution. Earth Environ. Sci. Trans. R. Soc. Edinb. 100, 311–350. ( 10.1017/s1755691010008145) [DOI] [Google Scholar]
- 25.Chipman AD. 2017. Oncopeltus fasciatus as an evo-devo research organism. Genesis e23020 ( 10.1002/dvg.23020) [DOI] [PubMed]
- 26.Toubiana W, Khila A. 2016. The benefits of expanding studies of trait exaggeration to hemimetabolous insects and beyond morphology. Curr. Opin. Genet. Dev. 39, 14–20. ( 10.1016/j.gde.2016.05.015) [DOI] [PubMed] [Google Scholar]
- 27.McGregor AP, Hilbrant M, Pechmann M, Schwager EE, Prpic NM, Damen WGM. 2008. Cupiennius salei and Achaearanea tepidariorum: spider models for investigating evolution and development. Bioessays 30, 487–498. ( 10.1002/bies.20744) [DOI] [PubMed] [Google Scholar]
- 28.Leite DJ, McGregor AP. 2016. Arthropod evolution and development: recent insights from chelicerates and myriapods. Curr. Opin. Genet. Dev. 39, 93–100. ( 10.1016/j.gde.2016.06.002) [DOI] [PubMed] [Google Scholar]
- 29.Sharma PP, Schwager EE, Extavour CG, Giribet G. 2012. Hox gene expression in the harvestman Phalangium opilio reveals divergent patterning of the chelicerate opisthosoma. Evol. Dev. 14, 450–463. ( 10.1111/j.1525-142X.2012.00565.x) [DOI] [PubMed] [Google Scholar]
- 30.Sharma PP. 2017. Chelicerates and the conquest of land: a view of arachnid origins through an evo-devo spyglass. Integr. Comp. Biol. 57, 510–522. ( 10.1093/icb/icx078) [DOI] [PubMed] [Google Scholar]
- 31.Stauber M, Taubert H, Schmidt-Ott U. 2000. Function of bicoid and hunchback homologs in the basal cyclorrhaphan fly Megaselia (Phoridae). Proc. Natl Acad. Sci. USA 97, 10 844–10 849. ( 10.1073/pnas.190095397) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schmidt-Ott U, Lynch JA. 2016. Emerging developmental genetic model systems in holometabolous insects. Curr. Opin. Genet. Dev. 39, 116–128. ( 10.1016/j.gde.2016.06.004) [DOI] [PubMed] [Google Scholar]
- 33.García-Solache M, Jaeger J, Akam M. 2010. A systematic analysis of the gap gene system in the moth midge Clogmia albipunctata. Dev. Biol. 344, 306–318. ( 10.1016/j.ydbio.2010.04.019) [DOI] [PubMed] [Google Scholar]
- 34.Cicin-Sain D, Pulido AH, Crombach A, Wotton KR, Jiménez-Guri E, Taly JF, Roma G, Jaeger J. 2015. SuperFly: a comparative database for quantified spatio-temporal gene expression patterns in early dipteran embryos. Nuc. Acids Res. 43, D751–D755. ( 10.1093/nar/gku1142) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pavlopoulos A, Averof M. 2005. Establishing genetic transformation for comparative developmental studies in the crustacean Parhyale hawaiensis. Proc. Natl Acad. Sci. USA 102, 7888–7893. ( 10.1073/pnas.0501101102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Eriksson BJ, Ungerer P, Stollewerk A. 2013. The function of Notch signalling in segment formation in the crustacean Daphnia magna (Branchiopoda). Dev. Biol. 383, 321–330. ( 10.1016/j.ydbio.2013.09.021) [DOI] [PubMed] [Google Scholar]
- 37.Sun DA, Patel NH. 2019. The amphipod crustacean Parhyale hawaiensis: an emerging comparative model of arthropod development, evolution, and regeneration. Wiley Interdiscip. Rev. Dev. Biol. 8, e355 ( 10.1002/wdev.355) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Arthur W, Chipman AD. 2005. The centipede Strigamia maritima: what it can tell us about the development and evolution of segmentation. Bioessays 27, 653–660. ( 10.1002/bies.20234) [DOI] [PubMed] [Google Scholar]
- 39.Janssen R, Prpic N-M, Damen WGM. 2004. Gene expression suggests decoupled dorsal and ventral segmentation in the millipede Glomeris marginata (Myriapoda : Diplopoda). Dev. Biol. 268, 89–104. ( 10.1016/j.ydbio.2003.12.021) [DOI] [PubMed] [Google Scholar]
- 40.Medved V, Marden JH, Fescemyer HW, Der JP, Liu J, Mahfooz N, Popadic A. 2015. Origin and diversification of wings: insights from a neopteran insect. Proc. Natl Acad. Sci. USA 112, 15 946–15 951. ( 10.1073/pnas.1509517112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Clark-Hachtel CM, Tomoyasu Y. 2016. Exploring the origin of insect wings from an evo-devo perspective. Curr. Opin. Insect Sci. 13, 77–85. ( 10.1016/j.cois.2015.12.005) [DOI] [PubMed] [Google Scholar]
- 42.Thomas GWC, et al. 2019. The genomic basis of arthropod diversity. bioRxiv ( 10.1101/382945) [DOI]
- 43.Chipman AD, Arthur W, Akam M. 2004. Early development and segment formation in the centipede Strigamia maritima (Geophilomorpha). Evol. Dev. 6, 78–89. ( 10.1111/j.1525-142X.2004.04016.x) [DOI] [PubMed] [Google Scholar]
- 44.Choe CP, Miller SC, Brown SJ. 2006. A pair-rule gene circuit defines segments sequentially in the short-germ insect Tribolium castaneum. Proc. Natl Acad. Sci. USA 103, 6560–6564. ( 10.1073/pnas.0510440103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Janssen R. 2012. Segment polarity gene expression in a myriapod reveals conserved and diverged aspects of early head patterning in arthropods. Dev. Genes Evol. 222, 299–309. ( 10.1007/s00427-012-0413-9) [DOI] [PubMed] [Google Scholar]
- 46.Brena C, Akam M. 2013. An analysis of segmentation dynamics throughout embryogenesis in the centipede Strigamia maritima. BMC Biol. 11, 112 ( 10.1186/1741-7007-11-112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Schönauer A, Paese CL, Hilbrant M, Leite DJ, Schwager EE, Feitosa NM, Eibner C, Damen WG, McGregor AP. 2016. The Wnt and Delta-Notch signalling pathways interact to direct pair-rule gene expression via caudal during segment addition in the spider Parasteatoda tepidariorum. Development 143, 2455–2463. ( 10.1242/dev.131656) [DOI] [PubMed] [Google Scholar]
- 48.Stahi R, Chipman AD. 2016. Blastoderm segmentation in Oncopeltus fasciatus and the evolution of arthropod segmentation mechanisms. Proc. R. Soc. B 283, 20161745 ( 10.1098/rspb.2016.1745) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Auman T, Chipman AD. 2018. Growth zone segmentation in the milkweed bug Oncopeltus fasciatus sheds light on the evolution of insect segmentation. BMC Evol. Biol. 18, 178 ( 10.1186/s12862-018-1293-z) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Giribet G, Edgecombe GD. 2019. The phylogeny and evolutionary history of arthropods. Curr. Biol. 29, R592–R602. ( 10.1016/j.cub.2019.04.057) [DOI] [PubMed] [Google Scholar]
- 51.Misof B, et al. 2014. Phylogenomics resolves the timing and pattern of insect evolution. Science 346, 763–767. ( 10.1126/science.1257570) [DOI] [PubMed] [Google Scholar]
- 52.Schwentner M, Combosch DJ, Pakes Nelson J, Giribet G. 2017. A phylogenomic solution to the origin of insects by resolving crustacean-hexapod relationships. Curr. Biol. 27, 1818–1824. ( 10.1016/j.cub.2017.05.040) [DOI] [PubMed] [Google Scholar]
- 53.Lozano-Fernandez J, et al. 2019. Pancrustacean evolution illuminated by taxon-rich genomic-scale data sets with an expanded remipede sampling. Genome Biol. Evol. 11, 2055–2070. ( 10.1093/gbe/evz097) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fernández R, Laumer CE, Vahtera V, Libro S, Kaluziak S, Sharma PP, Perez-Porro AR, Edgecombe GD, Giribet G. 2014. Evaluating topological conflict in centipede phylogeny using transcriptomic data sets. Mol. Biol. Evol. 31, 1500–1513. ( 10.1093/molbev/msu108) [DOI] [PubMed] [Google Scholar]
- 55.Fernández R, Edgecombe GD, Giribet G. 2018. Phylogenomics illuminates the backbone of the Myriapoda Tree of Life and reconciles morphological and molecular phylogenies. Sci. Rep. 8, 83 ( 10.1038/s41598-017-18562-w) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lozano-Fernandez J, Tanner AR, Giacomelli M, Carton R, Vinther J, Edgecombe GD, Pisani D. 2019. Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida. Nat. Commun. 10, 2295 ( 10.1038/s41467-019-10244-7) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ballesteros JA, Sharma PP. In press A critical appraisal of the placement of Xiphosura (Chelicerata) with account of known sources of phylogenetic error. Syst. Biol. ( 10.1093/sysbio/syz011) [DOI] [PubMed] [Google Scholar]
- 58.Daley AC, Antcliffe JB, Drage HB, Pates S. 2018. Early fossil record of Euarthropoda and the Cambrian Explosion. Proc. Natl Acad. Sci. USA 115, 5323–5331. ( 10.1073/pnas.1719962115) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Legg DA, Vannier J. 2013. The affinities of the cosmopolitan arthropod Isoxys and its implications for the origin of arthropods. Lethaia 46, 540–550. ( 10.1111/let.12032) [DOI] [Google Scholar]
- 60.Yang J, Ortega-Hernández J, Legg DA, Lan T, Hou JB, Zhang XG. 2018. Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods. Nat. Commun. 9, 470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Aria C, Caron JB. 2017. Burgess Shale fossils illustrate the origin of the mandibulate body plan. Nature 545, 89–92. ( 10.1038/nature22080) [DOI] [PubMed] [Google Scholar]
- 62.Vannier J, Aria C, Taylor RS, Caron JB. 2018. Waptia fieldensis Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale. R. Soc. open sci. 5 ( 10.1098/rsos.172206) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Liu JN, Shu D, Han J, Zhang ZF, Zhang XL. 2006. A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstatte. Acta Palaeontol. Pol. 51, 215–222. [Google Scholar]
- 64.Liu JN, Shu DG, Han J, Zhang ZF, Zhang XL. 2007. Morpho-anatomy of the lobopod Magadictyon cf. haikouensis from the Early Cambrian Chengjiang Lagerstatte, South China. Acta Zool. 88, 279–288. ( 10.1111/j.1463-6395.2007.00281.x) [DOI] [Google Scholar]
- 65.Dzik J. 2011. The xenusian-to-anomalocaridid transition within the lobopodians. B. Soc. Paleontol. Ital. 50, 65–74. [Google Scholar]
- 66.Daley AC, Edgecombe GD. 2014. Morphology of Anomalocaris canadensis from the Burgess Shale. J. Paleont. 88, 68–91. ( 10.1666/13-067) [DOI] [Google Scholar]
- 67.Park TS, et al. 2018. Brain and eyes of Kerygmachela reveal protocerebral ancestry of the panarthropod head. Nat. Commun. 9, 1019 ( 10.1038/s41467-018-03464-w) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Daley AC, Budd GE, Caron JB, Edgecombe GD, Collins D. 2009. The Burgess Shale anomalocaridid Hurdia and its significance for early euarthropod evolution. Science 323, 1597–1600. ( 10.1126/science.1169514) [DOI] [PubMed] [Google Scholar]
- 69.Van Roy P, Daley AC, Briggs DE. 2015. Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps. Nature 522, 77–80. ( 10.1038/nature14256) [DOI] [PubMed] [Google Scholar]
- 70.Jockusch EL, Smith FW. 2015. Hexapoda: comparative aspects of later embryogenesis and development. In Evolutionary developmental biology of invertebrates (ed. Wanninger A.), pp. 111–208. Vienna, Austria: Springer. [Google Scholar]
- 71.Janssen R, Eriksson BJ, Budd GE, Akam M, Prpic NM. 2010. Gene expression patterns in an onychophoran reveal that regionalization predates limb segmentation in pan-arthropods. Evol. Dev. 12, 363–372. ( 10.1111/j.1525-142X.2010.00423.x) [DOI] [PubMed] [Google Scholar]
- 72.Janssen R, Jorgensen M, Prpic NM, Budd GE. 2015. Aspects of dorso-ventral and proximo-distal limb patterning in onychophorans. Evol. Dev. 17, 21–33. ( 10.1111/ede.12107) [DOI] [PubMed] [Google Scholar]
- 73.Ortega-Hernández J. 2016. Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848. Biol. Rev. Camb. Philos. Soc. 91, 255–273. ( 10.1111/brv.12168) [DOI] [PubMed] [Google Scholar]
- 74.Wagner GP. 2014. Homology, genes and evolutionary innovation. Princeton, NJ: Princeton University Press. [Google Scholar]
- 75.Arthur W, Jowett T, Panchen A. 1999. Segments, limbs, homology, and co-option. Evol. Dev. 1, 74–76. ( 10.1046/j.1525-142x.1999.98004.x) [DOI] [PubMed] [Google Scholar]
- 76.Ganfornina MD, Sanchez D. 1999. Generation of evolutionary novelty by functional shift. Bioessays 21, 432–439. ( 10.1002/(SICI)1521-1878(199905)21:5<432::AID-BIES10>3.0.CO;2-T) [DOI] [PubMed] [Google Scholar]
- 77.Shubin N, Tabin C, Carroll S. 2009. Deep homology and the origins of evolutionary novelty. Nature 457, 818–823. ( 10.1038/nature07891) [DOI] [PubMed] [Google Scholar]
- 78.Peterson T, Müller GB. 2013. What is evolutionary novelty? Process versus character based definitions. J. Exp. Zool. B 320, 345–350. ( 10.1002/jez.b.22508) [DOI] [PubMed] [Google Scholar]
- 79.Panganiban G, Nagy L, Carroll SB. 1994. The role of the Distal-less gene in the development and evolution of insect limbs. Curr. Biol. 4, 671–675. ( 10.1016/S0960-9822(00)00151-2) [DOI] [PubMed] [Google Scholar]
- 80.Hughes CL, Kaufman TC. 2002. Hox genes and the evolution of the arthropod body plan. Evol. Dev. 4, 459–499. ( 10.1046/j.1525-142X.2002.02034.x) [DOI] [PubMed] [Google Scholar]
- 81.Auman T, Chipman AD. 2017. The evolution of gene regulatory networks that define arthropod body plans. Integr. Comp. Biol. 57, 523–532. ( 10.1093/icb/icx035) [DOI] [PubMed] [Google Scholar]
- 82.Fu D, Ortega-Hernandez J, Daley AC, Zhang X, Shu D. 2018. Anamorphic development and extended parental care in a 520 million-year-old stem-group euarthropod from China. BMC Evol. Biol. 18, 147 ( 10.1186/s12862-018-1262-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Dunlop JA, Lamsdell JC. 2017. Segmentation and tagmosis in Chelicerata. Arthropod Struct. Dev. 46, 395–418. ( 10.1016/j.asd.2016.05.002) [DOI] [PubMed] [Google Scholar]
- 84.Miyawaki K, Mito T, Sarashina I, Zhang HJ, Shinmyo Y, Ohuchi H, Noji S. 2004. Involvement of Wingless/Armadillo signaling in the posterior sequential segmentation in the cricket, Gryllus bimaculatus (Orthoptera), as revealed by RNAi analysis. Mech. Dev. 121, 119–130. ( 10.1016/j.mod.2004.01.002) [DOI] [PubMed] [Google Scholar]
- 85.McGregor AP, Pechmann M, Schwager EE, Feitosa NM, Kruck S, Aranda M, Damen WG. 2008. Wnt8 is required for growth-zone establishment and development of opisthosomal segments in a spider. Curr. Biol. 18, 1619–1623. ( 10.1016/j.cub.2008.08.045) [DOI] [PubMed] [Google Scholar]
- 86.Mittmann B, Wolff C. 2012. Embryonic development and staging of the cobweb spider Parasteatoda tepidariorum C. L. Koch, 1841 (syn.: Achaearanea tepidariorum; Araneomorphae; Theridiidae). Dev. Genes Evol. 222, 189–216. ( 10.1007/s00427-012-0401-0) [DOI] [PubMed] [Google Scholar]
- 87.Dearden PK, Donly C, Grbic M. 2002. Expression of pair-rule gene homologues in a chelicerate: early patterning of the two-spotted spider mite Tetranychus urticae. Development 129, 5461–5472. ( 10.1242/dev.00099) [DOI] [PubMed] [Google Scholar]
- 88.Smith FW, Bartels PJ, Goldstein B. 2017. A hypothesis for the composition of the tardigrade brain and its implications for panarthropod brain evolution. Integr. Comp. Biol. 57, 546–559. ( 10.1093/icb/icx081) [DOI] [PubMed] [Google Scholar]
- 89.Mayer G, Kauschke S, Rüdiger J, Stevenson PA. 2013. Neural markers reveal a one-segmented head in tardigrades (water bears). PLoS ONE 8, e59090 ( 10.1371/journal.pone.0059090) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Cong PY, Edgecombe GD, Daley AC, Guo J, Pates S, Hou XG. 2018. New radiodonts with gnathobase-like structures from the Cambrian Chengjiang biota and implications for the systematics of Radiodonta. Pap. Palaeontol. 4, 605–621. ( 10.1002/spp2.1219) [DOI] [Google Scholar]
- 91.Chen JY, Edgecombe GD, Ramskold L, Zhou GQ. 1995. Head segmentation in Early Cambrian Fuxianhuia: implications for arthropod evolution. Science 268, 1339–1343. ( 10.1126/science.268.5215.1339) [DOI] [PubMed] [Google Scholar]
- 92.Budd GE. 2008. Head structure in upper stem-group euarthropods. Palaeontology 51, 561–573. ( 10.1111/j.1475-4983.2008.00752.x) [DOI] [Google Scholar]
- 93.Hunnekuhl VS, Akam M. 2017. Formation and subdivision of the head field in the centipede Strigamia maritima, as revealed by the expression of head gap gene orthologues and hedgehog dynamics. EvoDevo 8, 18 ( 10.1186/s13227-017-0082-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kanayama M, Akiyama-Oda Y, Nishimura O, Tarui H, Agata K, Oda H. 2011. Travelling and splitting of a wave of hedgehog expression involved in spider-head segmentation. Nat. Commun. 2, 500 ( 10.1038/ncomms1510) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Posnien N, Bucher G. 2010. Formation of the insect head involves lateral contribution of the intercalary segment, which depends on Tc-labial function. Dev. Biol. 338, 107–116. ( 10.1016/j.ydbio.2009.11.010) [DOI] [PubMed] [Google Scholar]
- 96.Posnien N, Schinko JB, Kittelmann S, Bucher G. 2010. Genetics, development and composition of the insect head: a beetle's view. Arthropod. Struct. Dev. 39, 399–410. ( 10.1016/j.asd.2010.08.002) [DOI] [PubMed] [Google Scholar]
- 97.Gallitano-Mendel A, Finkelstein R. 1997. Novel segment polarity gene interactions during embryonic head development in Drosophila. Dev. Biol. 192, 599–613. ( 10.1006/dbio.1997.8753) [DOI] [PubMed] [Google Scholar]
- 98.Vannier J, Liu J, Lerosey-Aubril R, Vinther J, Daley AC. 2014. Sophisticated digestive systems in early arthropods. Nat. Commun. 5, 3641 ( 10.1038/ncomms4641) [DOI] [PubMed] [Google Scholar]
- 99.Hannibal RL, Price AL, Patel NH. 2012. The functional relationship between ectodermal and mesodermal segmentation the crustacean, Parhyale hawaiensis. Dev. Biol. 361, 427–438. ( 10.1016/j.ydbio.2011.09.033) [DOI] [PubMed] [Google Scholar]
- 100.Azpiazu N, Lawrence PA, Vincent JP, Frasch M. 1996. Segmentation and specification of the Drosophila mesoderm. Gene. Dev. 10, 3183–3194. ( 10.1101/gad.10.24.3183) [DOI] [PubMed] [Google Scholar]
- 101.Ortega-Hernández J, Fu D, Zhang X, Shu D. 2018. Gut glands illuminate trunk segmentation in Cambrian fuxianhuiids. Curr. Biol. 28, R146–R147. ( 10.1016/j.cub.2018.01.040) [DOI] [PubMed] [Google Scholar]
- 102.Yang J, Ortega-Hernández J, Butterfield NJ, Liu Y, Boyan GS, Hou JB, Lan T, Zhang XG. 2016. Fuxianhuiid ventral nerve cord and early nervous system evolution in Panarthropoda. Proc. Natl Acad. Sci. USA 113, 2988–2993. ( 10.1073/pnas.1522434113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ortega-Hernández J, Brena C. 2012. Ancestral patterning of tergite formation in a centipede suggests derived mode of trunk segmentation in trilobites. PLoS ONE 7, e52623 ( 10.1371/journal.pone.0052623) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Martínez-Arias A, Lawrence PA. 1985. Parasegments and compartments in the Drosophila embryo. Nature 313, 639–642. ( 10.1038/313639a0) [DOI] [PubMed] [Google Scholar]
- 105.Janssen R, Damen WGM. 2006. The ten Hox genes of the millipede Glomeris marginata. Dev. Genes Evol. 216, 451–465. ( 10.1007/s00427-006-0092-5) [DOI] [PubMed] [Google Scholar]
- 106.Nulsen C, Nagy LM. 1999. The role of wingless in the development of multibranched crustacean limbs. Dev. Genes. Evol. 209, 340–348. ( 10.1007/s004270050262) [DOI] [PubMed] [Google Scholar]
- 107.Scholtz G. 2015. Heads and brains in arthropods: 40 years after the ‘endless dispute’. In Structure and evolution of invertebrate nervous systems (eds Schmidt-Rhaesa A, Harzsch S, Purschke G), pp. 402–410. Oxford, UK: Oxford University Press. [Google Scholar]
- 108.Chipman AD. 2015. An embryological perspective on the early arthropod fossil record. BMC Evol. Biol. 15, 285 ( 10.1186/s12862-015-0566-z) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Fusco G, Garland T Jr, Hunt G, Hughes NC. 2012. Developmental trait evolution in trilobites. Evolution 66, 314–329. ( 10.1111/j.1558-5646.2011.01447.x) [DOI] [PubMed] [Google Scholar]
- 110.Rota-Stabelli O, Daley AC, Pisani D. 2013. Molecular timetrees reveal a Cambrian colonization of land and a new scenario for ecdysozoan evolution. Curr. Biol. 23, 392–398. ( 10.1016/j.cub.2013.01.026) [DOI] [PubMed] [Google Scholar]
- 111.Lozano-Fernandez J, et al. 2016. A molecular palaeobiological exploration of arthropod terrestrialization. Phil. Trans. R. Soc. B 371, 20150133 ( 10.1098/rstb.2015.0133) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Wolfe JM, Daley AC, Legg DA, Edgecombe GD. 2016. Fossil calibrations for the arthropod Tree of Life. Earth-Sci. Rev. 160, 43–110. ( 10.1016/j.earscirev.2016.06.008) [DOI] [Google Scholar]
- 113.Bengtson S. 2000. Teasing fossils out of shales with cameras and computers. Palaeo. Electron. 3, 4. [Google Scholar]
- 114.Haug JT, et al. 2011. Autofluorescence imaging, an excellent tool for comparative morphology. J. Microsc. 244, 259–722. ( 10.1111/j.1365-2818.2011.03534.x) [DOI] [PubMed] [Google Scholar]
- 115.Zuber M, Laass M, Hamann E, Kretschmer S, Hauschke N, van de Kamp T, Baumbach T, Koenig T.. 2017. Augmented laminography, a correlative 3D imaging method for revealing the inner structure of compressed fossils. Sci. Rep. 7, 41413 ( 10.1038/srep41413) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Zhai D, Ortega-Hernández J, Wolfe JM, Hou X, Cao C, Liu Y. 2019. Three-dimensionally preserved appendages in an Early Cambrian stem-group pancrustacean. Curr. Biol. 29, 171–177. ( 10.1016/j.cub.2018.11.060) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
This article has no additional data.