

Summary
Marine sponges represent one of the few eukaryotic groups that frequently harbour symbiotic members of the Thaumarchaeota, which are important chemoautotrophic ammonia‐oxidizers in many environments. However, in most studies, direct demonstration of ammonia‐oxidation by these archaea within sponges is lacking, and little is known about sponge‐specific adaptations of ammonia‐oxidizing archaea (AOA). Here, we characterized the thaumarchaeal symbiont of the marine sponge Ianthella basta using metaproteogenomics, fluorescence in situ hybridization, qPCR and isotope‐based functional assays. ‘Candidatus Nitrosospongia ianthellae’ is only distantly related to cultured AOA. It is an abundant symbiont that is solely responsible for nitrite formation from ammonia in I. basta that surprisingly does not harbour nitrite‐oxidizing microbes. Furthermore, this AOA is equipped with an expanded set of extracellular subtilisin‐like proteases, a metalloprotease unique among archaea, as well as a putative branched‐chain amino acid ABC transporter. This repertoire is strongly indicative of a mixotrophic lifestyle and is (with slight variations) also found in other sponge‐associated, but not in free‐living AOA. We predict that this feature as well as an expanded and unique set of secreted serpins (protease inhibitors), a unique array of eukaryotic‐like proteins, and a DNA‐phosporothioation system, represent important adaptations of AOA to life within these ancient filter‐feeding animals.
Introduction
Marine sponges (phylum Porifera) are among the most basal metazoan lineages (Simion et al., 2017), with fossil records suggesting their evolutionary emergence more than 600 million years ago (Love et al., 2009). Sponges form a major part of the marine benthic fauna across the world's oceans (Bell, 2008 ) where they mediate critical biogeochemical processes through their filtration of immense volumes of seawater (Maldonado et al., 2012; de Goeij et al., 2013). Many sponges harbour dense, diverse and species‐specific communities of microbes (Hentschel et al., 2012; Thomas et al., 2016), and these associations are often temporally and geographically stable (Luter et al., 2010; Schmitt et al., 2012; Astudillo‐García et al., 2017). Functional roles that have been assigned to specific sponge‐associated microorganisms include the provision of photosynthates (Wilkinson, 1983) and fixed N2 (Wilkinson and Fay, 1979) by cyanobacterial symbionts and the production of bioactive secondary metabolites (Wilson et al., 2014; Freeman et al., 2016; Agarwal et al., 2017). Furthermore, sponge symbionts can serve as an endogenous food source as exemplified by biomass transfer from a sponge‐associated sulfate‐reducing bacterial community to host cells (Hoffmann et al., 2005), and the ingestion of symbiotic methanotrophs in a deep‐sea carnivorous sponge (Vacelet et al., 1995). Whilst a vast array of additional putative symbiotic functions have been hypothesized from taxonomic or metagenomic data (Webster and Thomas, 2016), unequivocal evidence for specific sponge symbiont physiologies is comparably rare.
Nitrogen cycling in sponge holobionts has received considerable attention and sponge‐symbiont‐driven nitrification, denitrification and anaerobic ammonium oxidation has been described (Bayer et al., 2008; Southwell et al., 2008; Hoffmann et al., 2009; Schläppy et al., 2010; Radax et al., 2012a). Sponges are marine filter feeders and during decay of organic matter (from food and sponge cells) ammonia is released. Since ammonia is more toxic than nitrite and nitrate, for aquatic animals (Camargo and Alonso, 2006), nitrification that is performed by sponge‐associated microbes might be beneficial for the host. In addition, growth of nitrifier symbionts on waste products of the host (NH4 + and CO2) could also be a way of efficiently recycling those into new microbial food biomass. Consistently, many marine sponges harbour symbionts phylogenetically related to nitrifying microbes (Steger et al., 2008; Hoffman et al., 2009; Off et al., 2010), indicating that ammonia oxidation via nitrite to nitrate is a widely distributed process in these animals. More specifically, molecular signatures of proteobacterial as well as thaumarchaeal ammonia oxidizers are frequently detected in sponges, although these microbes rarely co‐occur in the same host (Bayer et al., 2008; Radax et al., 2012b). Most sponges that contain symbionts related to ammonia‐oxidizers also host bacteria affiliated with known nitrite‐oxidizers, particularly members of the genus Nitrospira (Hoffmann et al., 2009; Off et al., 2010; Reveillaud et al., 2014; Moitinho‐Silva et al., 2017a). Whilst many studies simply equate the molecular detection of microbes related to recognized nitrifiers with the occurrence of canonical nitrification in sponges (Mohamed et al., 2010; Fan et al., 2012), this assumption could be misleading due to the functional versatility of both Thaumarchaeota and Nitrospira (Mußmann et al., 2011; Koch et al., 2014; Daims et al., 2015; Palatinszky et al., 2015).
Thaumarchaeotes are important ammonia oxidizers in many environments (Pester et al., 2011). Marine sponges host a particularly high diversity of thaumarchaeotes, with many of the 16S rRNA gene sequences falling into phylogenetic clusters that exclusively contain sponge‐derived sequences (Simister et al., 2012). The detection of thaumarchaeotes in sponge larvae also suggests vertical transmission or early environmental acquisition of these symbionts (Sharp et al., 2007; Steger et al., 2008; Schmitt et al., 2012). The first genomic information of a member of this phylum was derived from Ca. Cenarchaeum symbiosum in the marine sponge Axinella mexicana (Schleper et al., 1998; Hallam et al., 2006), and subsequently thaumarchaeal genomes were recovered from the deep sea glass sponge Lophophysema eversa (Tian et al., 2016; referred to as DSGS‐AOA in this manuscript) and the temperate sponge Cymbastella concentrica, with the latter also shown to transcribe genes for ammonia oxidation (referred to as CCThau in Moitinho Silva et al., 2017a and in this manuscript). However, little information is available on the genomic plasticity and mechanisms of host adaptation in sponge thaumarchaeal symbionts. Furthermore, with the exception of a thaumarchaeal symbiont that inhabits the cold‐water sponge Geodia barretti (Radax et al., 2012a, b), direct evidence for the catalysis of ammonia oxidation by sponge‐associated thaumarchaeotes is lacking. Whilst transcription and translation of key functional genes like the amoA gene encoding a subunit of the ammonia monooxygenase of thaumarchaeotes has been detected across multiple sponge species (Liu et al., 2012; Fiore et al., 2015; Moitinho‐Silva et al., 2017a), experimental validation of their involvement in nitrification is required to confirm this activity in situ (Mußmann et al., 2011).
To better understand, the physiological capability of Thaumarchaeota in sponges, we used a metaproteogenomic approach to characterize the Thaumarchaeota symbiont of the marine sponge Ianthella basta. Ianthella basta is an abundant and ecologically important reef sponge found throughout the Indo‐Pacific (Bergquist and Kelly‐Borges, 1995). In contrast to many microbially diverse sponge species, 16S rRNA gene surveys have shown that I. basta harbours only three dominant microbial phylotypes which belong to a novel α‐proteobacterial lineage, a γ‐proteobacterial clade within the UBA10353 order and LS‐SOB family (Parks et al., 2018), and the Thaumarchaeota (Luter et al., 2010). This community structure is stable among different host colour morphotypes (Freckelton et al., 2012), between individuals sampled from different geographic regions (Luter et al., 2010), across different host health states (Luter et al., 2010) and in sponges exposed to different environmental stressors (Luter et al., 2012). Here we (i) quantify the abundance of the thaumarchaeal symbionts in I. basta, (ii) confirm their activity as ammonia oxidizers, (iii) document the expression of almost 100 thaumarchaeal genes in situ and (iv) reveal sponge‐specific adaptations of these archaea including a putative mixotrophic lifestyle. Furthermore, we demonstrate that nitrite oxidation surprisingly does not occur in I. basta.
Results and discussion
Quantification of the thaumarchaeal I. basta symbiont using FISH and qPCR
Fluorescence in situ hybridization (FISH) was performed on fixed cryosections from one sponge individual using the general archaeal probe Arch915 that is fully complementary to the 16S rRNA of the single archaeal phylotype known to inhabit I. basta (Fig. 1). Quantitative FISH across 10 images revealed that the thaumarchaeal symbiont comprised 24% ± 1.6% [standard error (SE)] of the total bacterial and archaeal cells detected with probes Arch915 and the probe set EUB338‐I‐III targeting most bacteria. Absolute quantification of the thaumarchaeal symbiont in five sponge individuals (including those used for metagenome sequencing) using specifically designed qPCR primers, revealed an average absolute abundance of 2.41 ± 0.7 (SE) × 1010 16S rRNA gene copies per gram wet weight of sponge tissue. Consistent with the relative abundances derived from FISH, the ratio of thaumarchaeal 16S rRNA sequences to the total 16S rRNA genes derived from qPCR assays targeting the additional α‐ and γ‐proteobacterial symbionts of I. basta (data not shown) was 22% ± 2.3% (SE). The qPCR experiments were performed with five sponge individuals, and sponge samples within an individual, were randomly selected, rendering it highly unlikely that the abundance pattern of the thaumarchaeal symbiont throughout a sponge varies significantly. Collectively, these data demonstrate that the I. basta thaumarchaeote is a dominant member of the I. basta microbiome, occurring at densities that exceed the total microbial biomass of many tropical sponge species (Taylor et al., 2007).
Figure 1.
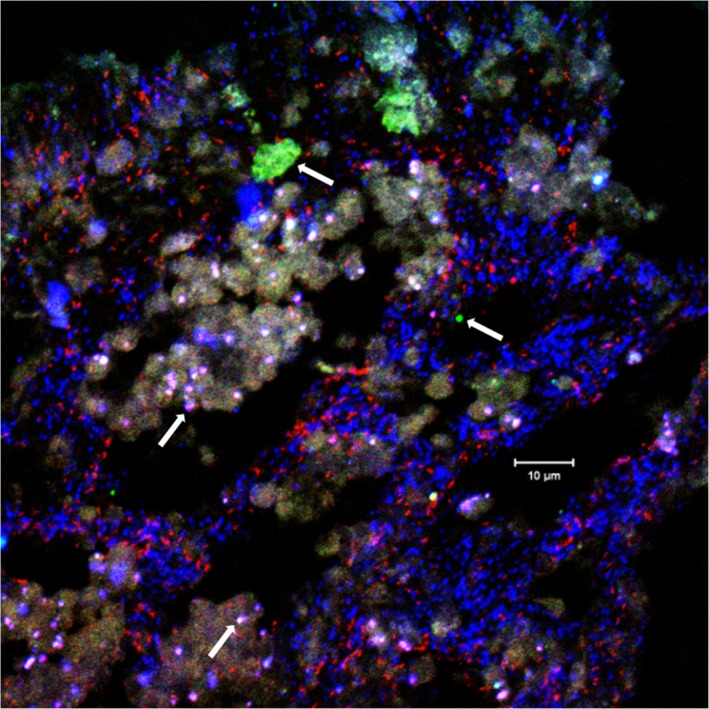
Fluorescence in situ hybridization of a 5 μm cryosection of I. basta using double‐labelled (Stoecker et al., 2010) probe Arch915 in red and the double‐labelled probe EUB338‐I‐III set in blue. Green and white/lila structures represent autofluorescence (see white arrows). As I. basta harbours ‘Ca. Nitrosospongia ianthellae’ as the only archaeon, all red signals represent this AOA. Blue signals represent bacterial symbionts.
Metaproteogenomic analyses of the thaumarchaeal symbiont of I. basta
An extensive metagenomic data set consisting of 25.8 Gbp of sequence information was obtained for an I. basta individual by Illumina sequencing. To facilitate the assembly of symbiont genomes, cell fractions were specifically concentrated for sponge symbionts and dissociated from sponge nuclei. As a result, we recovered a 1.99 Mbp metagenome‐assembled genome (MAG) consisting of 113 contigs, representing a nearly complete thaumarchaeal genome (99%) with very low contamination (Supporting Information Table S1). Of all genome‐sequenced thaumarchaeotes, the I. basta symbiont has the highest average genomic amino acid identity (AAI) of 58.1% with Ca. Nitrosopumilus piranensis. As this represents a new species within a new genus (60%–80% AAI is typical for organisms grouped at the genus level; Luo et al., 2014; Konstantinidis et al., 2017), we propose the name Ca. Nitrosospongia ianthellae. The name Nitrosospongia describes this organism's ability to oxidize ammonia to nitrite (nitrosus is the Latin adjective meaning full of natron; here intended to mean nitrous) whilst residing within a sponge (spongia is the feminine Latin noun for sponge). The species name ianthellae refers to its discovery and description as a symbiont of the marine sponge, I. basta. Furthermore, 16S rRNA gene, amoA and concatenated marker gene phylogenies suggest that Ca. N. ianthellae is a member of the family Ca. Nitrosopumilaceae (Qin et al., 2016) (Fig. 2 and Supporting Information Fig. S1). Based on the 16S rRNA gene sequence, Ca. N. ianthellae is a member of the sponge‐specific sequence cluster 174 (Simister et al., 2012) which does not include the three other sponge thaumarchaeal symbionts (C. symbiosum, CCThau and DSGS‐AOA, a putative deep‐sea glass sponge thaumarchaeal symbiont with the assembly accession number GCA_001541925.1) for which genome sequences are available (Hallam et al., 2006; Tian et al., 2016; Moitinho‐Silva et al., 2017a). The lack of a close relationship between Ca. N. ianthellae and the other sponge‐associated symbionts is consistent with the topology of the concatenated single copy conserved marker gene tree (Fig. 2B), whilst unexpectedly amoA phylogeny supported a clustering of Ca. N. ianthellae with C. symbiosum (Supporting Information Fig. S1). Whilst the amoA genes of C. symbiosum and Ca. N. ianthellae showed compositional bias (Li et al., 2014), it was not possible to determine whether this caused their monophyletic grouping in the amoA gene tree. Upon individual addition of C. symbiosum and Ca. N. ianthellae amoA gene sequences to an amoA data set, their phylogenetic position in the tree varied dependent on taxa selection (data not shown). Querying the Ca. N. ianthellae 16S rRNA gene against the sponge microbiome project (SMP) database (Moitinho‐Silva et al., 2017b) and against most other publicly available 16S rRNA gene amplicon data sets demonstrated that the habitat of Ca. N. ianthellae is restricted to a few sponge species (I. basta, Ancorina alata, Stellata maori, Stellata aremaria and Xestospongia exigua; see Supporting Information for more details).
Figure 2.
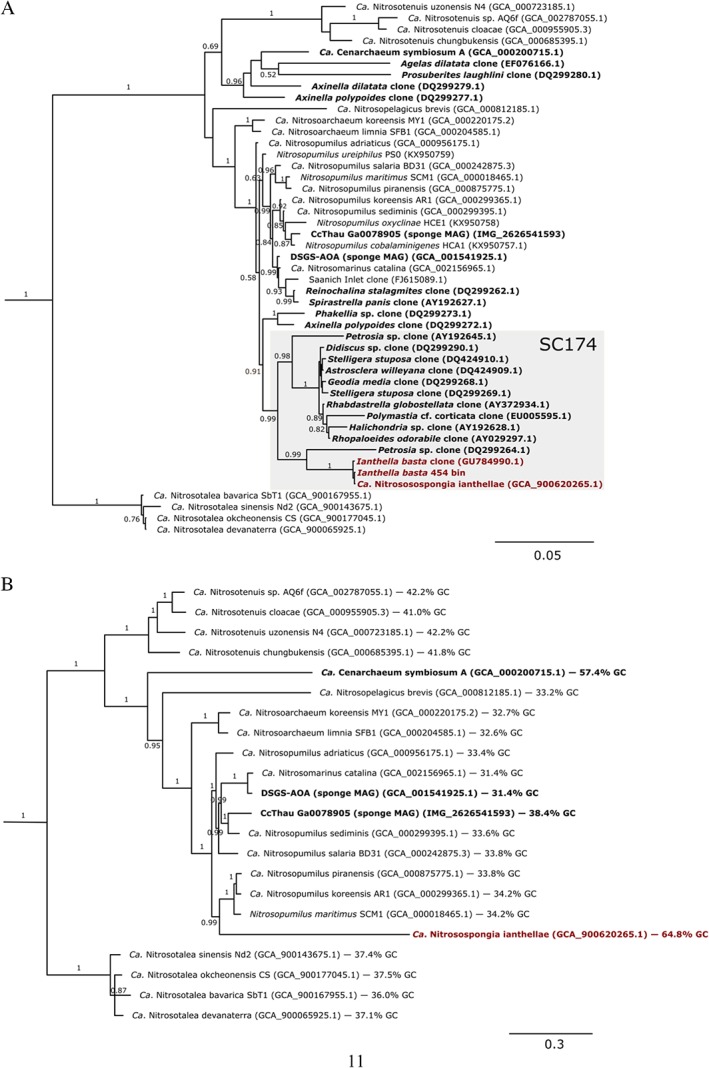
Phylogeny of Ca. Nitrosospongia ianthellae. A. Bayesian 16S rRNA gene tree. B. Bayesian phylogenomic tree based on 34 concatenated universal, single‐copy marker genes identified with CheckM (Parks et al., 2015). Bayesian posterior support values >0.5 are indicated for each branch. Outgroups for both trees consisted of all three genome‐sequenced members of the Nitrososphaera cluster, both members of the Nitrosocosmicus clade, and Ca. Nitrosocaldus icelandicus. In all trees, sequences obtained from sponges are depicted in bold.
An additional unpublished metagenomic data set (250 Mbp generated from pyrosequencing) derived from a different I. basta individual was also screened to confirm the presence of genes of interest (Supporting Information Table S2) and support the phylogenetic inferences displayed in Fig. 2. Whilst the shallow metagenome was excluded from detailed analyses, it confirmed that closely related thaumarchaeal symbionts inhabited both sponge individuals. The average nucleotide identity (ANI) between the two thaumarchaeote MAGs was 98.2%, with 82% coverage of the smaller (Illumina data set) MAG, confirming that members of the same thaumarchaeal species (Konstantinidis et al., 2017) reside in both sponge individuals. Both thaumarchaeal symbiont MAGs from I. basta possess the highest GC content (64.8%) of any genome‐sequenced thaumarchaeote (Fig. 2B) (see Supporting Information for a more detailed discussion of the high GC content).
Metaproteomic analysis was performed on the I. basta individual used for Illumina metagenomic sequencing. A total of 513 proteins was detected, 96 of which were specifically assigned to the I. basta thaumarchaeote (representing 5.4% of the genes in the MAG). When combining the normalized spectral abundance factor (NSAF; Florens et al., 2006) values for all samples and analyses, the 96 thaumarchaeal proteins comprised 10.5% of the 513 identified proteins (Supporting Information Table S3). Of the thaumarchaeal proteins, 60.2% (67.4% NSAF) were encoded by gene families shared by all thaumarchaeotes, and 7.1% (3.1% NSAF) were encoded by genes unique to the I. basta thaumarchaeote. Of the 96 expressed proteins, 88 were also encoded at a predicted average amino acid identity of 99.4% ± 1.4% (SD) in the second MAG recovered by pyrosequencing. The other eight protein homologues were also found in the second MAG but at much lower predicted amino acid identities (from 31.4% to 89%).
Core metabolism of Ca. N. ianthellae
AOA oxidize ammonia to conserve energy. Like their bacterial counterparts they activate ammonia with the help of an ammonia monooygenase (AMO). In bacterial ammonia oxidizers AMO forms hydroxylamine that is oxidized by hydroxylamine dehydrogenase to NO (Caronto and Lancaster, 2017) that is further oxidized to nitrite by a yet unknown enzyme. Ca. N. ianthellae encodes the recognized repertoire of AOA for ammonia oxidation. All genes encoding the subunits of the ammonia monooxygenase enzyme (amoA, amoB and amoC), including the hypothetical gene amoX were identified. AmoB and C were also detected as proteins (Supporting Information Table S3). The amo gene arrangement [amoA‐amoX‐amoC‐amoB] is syntenic to most analysed members of Ca. Nitrosopumilaceae (Lehtovirta‐Morley et al., 2011; Park et al., 2014). Consistent with other AOAs, no canonical hydroxylamine dehydrogenase was found but Ca. N. ianthellae encodes lineage one multicopper oxidases (MCO), which have been suggested as candidates for archaeal hydroxylamine dehydrogenases (Kerou et al., 2016). Like most other AOA (except for Ca. C. symbiosum), Ca. N. ianthellae encodes the putative NO‐forming nitrite reductase (nirK; found to be highly expressed as protein; Supporting Information Table S3) that has been suggested to play an important role for archaeal ammonia oxidation (Kozlowski et al., 2016; Carini et al., 2018) a hypothesis that nicely explains the inhibitory effect of the NO scavenger PTIO used in our incubation experiments described below. Interestingly however, the purple cupredoxin Nmar_1307 from Nitrosopumilus maritimus that is capable of oxidizing NO to NO2 − (Hosseinzadeh et al., 2016) is absent in Ca. N. ianthellae and other sponge‐associated AOA (and also many other AOA), but there are several mononuclear cupredoxins encoded by Ca. N. ianthellae that could have the same function. Consistent with all other AOA, no canonical NO‐ or N2O‐reductases were found suggesting that Ca. N. ianthellae cannot perform denitrification, although – like in many other thaumarchaeotes (Liu et al., 2012, Zhalnina et al., 2014; Santoro et al., 2015) – the putative nitric oxide reductase accessory proteins, NorQ and NorD are encoded in the genome (with the NorD subunit being expressed).
Urea and cyanate can be used by some ammonia oxidizers as ammonia source for energy conservation and assimilation. Consistent with marine and non‐marine thaumarchaeota (Hallam et al., 2006; Tourna et al., 2011; Spang et al., 2012; Park et al., 2014; Bayer et al., 2016), Ca. N. ianthellae possesses a complete urease gene cluster in addition to a gene encoding a urea active transporter (DUR3; 86% homologous to Ca. Nitrosopumilus piranensis) and is thus capable of taking up (or using internally produced) urea and converting it to ammonia and CO2. The sponge symbionts C. symbiosum and CCThau also encode urea transporters and urease gene clusters. Furthermore, urease subunit gene transcripts from CCThau were found in the metatranscriptomes from C. concentrica (Moitinho‐Silva et al., 2017a). Thus, it seems likely that urea is not only used by some free‐living marine AOA (Alonso‐Sáez et al., 2012; Kitzinger et al. 2019) and a few cultured AOA (Fig. 3), but also appears to be a common substrate for group I.1a Thaumarchaeota residing in marine sponges. In this context it is also interesting to note that some sponges excrete urea (Morley et al., 2016). In contrast to Nitrososphaera gargensis (Palatinszky et al., 2015), Ca. N. ianthellae does not contain a cyanase for ammonia generation from cyanate, but like all other AOA, it encodes a protein with modest homology to a creatinine‐amidohydrolase indicating that it could utilize sponge‐derived creatinine and convert it to creatine. Whilst all genome‐sequenced AOA symbionts lack a canonical creatinase that would form urea from creatine, AOA including Ca. N. ianthellae do possess a Xaa‐Pro aminopeptidase that has been hypothesized as a functional analog (Moitinho‐Silva et al., 2017a).
Figure 3.
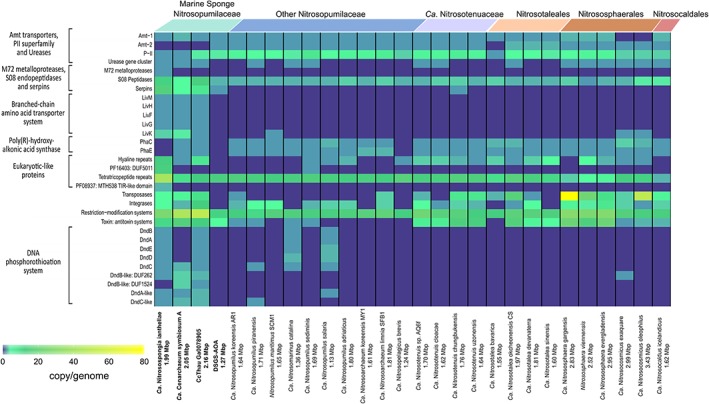
Heat map showing the distribution and gene copy number per genome of selected genes and gene classes among genome‐sequenced AOA. The colour scale indicates copies per genome and MAG respectively. Sponge‐derived MAGs start on the left and are depicted in bold, followed by members of Ca. Nitrosopumilaceae, Ca. Nitrosotenuaceae, Ca. Nitrosotaleales, the Nitrososphaerales, and Ca. Nitrosocaldales. Genome sizes are listed next to each species name. An extended version of this Figure is available as Supporting Information Fig. S6.
As expected, Ca. N. ianthellae encodes the thaumarchaeal 3‐hydroxypropionate/4‐hydroxybutyrate pathway for autotrophic CO2 fixation (see Supporting Information for more details). Regarding nitrogen assimilation, Ca. N. ianthellae encodes and expresses an amt transporter of the amt‐2 lineage which has been hypothesized to be a high affinity ammonia transporter based on indirect evidence from transcriptomic data (Nakagawa and Stahl, 2013; Offre et al., 2014). The amt‐2 gene of Ca. N. ianthellae is nested within the complex V ATP synthase operon (of which four of the encoded subunits were found to be expressed). Interestingly, in contrast to most other AOA, no putative low affinity amt transporter gene was found in Ca. N. ianthellae, a feature shared with the sponge symbionts Ca. C. symbiosum (Hallam et al., 2006) and CCThau (Fig. 3; Moitinho‐Silva et al., 2017a). Under the assumption that amt affinity groups can reliably be inferred from annotation, this finding might indicate that the dense thaumarchaeal populations in some marine sponges including I. basta (see also Radax et al., 2012a) may be perpetually ammonia limited and thus might rely on a high affinity ammonia transporter in combination with the uptake of organic nitrogen sources like urea or amino acids. Regarding nitrogen assimilation, it is also noteworthy, that the almost complete bin of Ca. N. ianthellae contains only a single gene encoding a member of the nitrogen regulatory protein PII superfamily, whereas, with the exception of Ca. C. symbiosum, higher copy numbers of these genes are generally found in other AOA (Fig. 3; Kerou et al., 2016). This observation might indicate that Ca. N. ianthellae lives in a rather stable environment in regard to nitrogen availability compared to other AOA. Furthermore, in contrast to most other AOA, the two key genes coding for subunits of the polyhydroxyalkanoate (PHA) synthase phaC and phaE are apparently lacking in Ca. N. ianthellae. We also found no indications for use of alternative carbon storage compounds such as starch or glycogen in Ca. N. ianthellae. The apparent lack of carbon storage would be consistent with balanced growth in a chemically stable environment.
The negatively charged surface of the Thaumarchaeota S‐layer was recently proposed to help concentrate the charged solute ammonium into the pseudo‐periplasmic space (Li et al., 2018). Intriguingly, Ca. N. ianthellae encodes many putative surface‐layer (S‐layer) proteins (Supporting Information Fig. S2), with six of them being expressed at combined NSAF values of 6.78% (Supporting Information Table S3). The theoretical isoelectric points of all Ca. N. ianthellae S‐layer proteins (3.62–4.92) were in a similar range to those calculated for N. maritimus and Nitrosoarchaeum limnia SFB1 S‐layer proteins (3.4–4.08), consistent with the proposed mechanism for charged solute acquisition. In addition to its ammonium concentration effect, the expanded group of S‐layer proteins in Ca. N. ianthellae (and Ca. C. symbiosum) may be involved in adhesion, surface recognition or other types of interactions within the extracellular matrix of the sponge environment.
Shared gene families of sponge AOA
During genome annotation of Ca. N. ianthellae, we particularly focused on gene families unique to this organism or to Ca. N. ianthellae and other thaumarchaeal sponge symbionts (among all genome‐sequenced thaumarchaeotes) as these gene families likely represent adaptations to a sponge‐associated lifestyle. Whilst 40% (represented by 681 gene families) of the Ca. N. ianthellae genes were members of core gene families represented in every sequenced thaumarchaeote, 35% (represented by 616 gene families) had no close orthologues among other thaumarchaeotes. Additional pairwise comparisons between Ca. N. ianthellae and all other thaumarchaeote genomes were performed in order to identify shared gene families of species pairs that are absent in all other thaumarchaeotes. Interestingly, the highest number occurred with CCThau (n = 18) and Ca. C. symbiosum (n = 11) (Supporting Information Fig. S3). Furthermore, 14 gene families were found to be exclusively shared among all three thaumarchaeal sponge symbionts, whilst only one gene family was found to be shared with DSGS‐AOA and C. symbiosum. In total, 44 gene families were exclusively shared with at least one other thaumarchaeal sponge symbiont representing 2.4% of the Ca. N. ianthellae genes. These genes are likely candidates for adaptations of AOA to life within sponges.
Ca. N. ianthellae and other sponge AOA possess proteases and their inhibitors
Among the genes that were unique to Ca. N. ianthellae or exclusively shared with other sponge AOA were several genes predicted to be involved in degradation of extracellular protein and inactivation of extracellular proteases, which may also be host‐derived. Ca. N. ianthellae possesses a putatively exported metalloprotease of the M72 family (Drapeau, 1980; Passmore et al., 2015) that was also detected in the metaproteome. This large protease (2027 AA) contains hyaline repeat domains suggestive of an additional adhesive property. The M72 family of metalloendopeptidases have so far not been found in any Archaea (Trame et al., 2014) and most characterized members of this enzyme family are peptidyl‐Asp metalloendopeptidases that hydrolyze bonds on the NH2‐terminal side of aspartic acid and cysteic acid residues (Drapeau, 1980). Furthermore, the I. basta thaumarchaeal symbiont contained 11 genes affiliated with four gene families that encode subtilisin‐like serine protease domains (S08A family endopeptidases) (Supporting Information Fig. 4A). Although serine endopeptidases of the S08A family are also found in other thaumarchaeotes, except for C. symbiosum (four copies), and CCThau (13 copies), most contain only two copies. Furthermore, it is noteworthy that two of the four S08A family endopeptidase gene families were either exclusively found in Ca. N. ianthellae or shared with CCThau (Fig. 4, Supporting Information Fig. 4A). Interestingly, several of the Ca. N. ianthellae S08A endopeptidases are predicted to be exported without a membrane anchor, whilst most other thaumarchaeal S08A serine endopeptidases seem to be membrane anchored. This suggests that the Ca. N. ianthellae S08A endopeptidases act at larger distances outside of the pseudoperiplasmic space where host‐ or food‐derived proteins that cannot pass through the S‐layer will be available.
Figure 4.
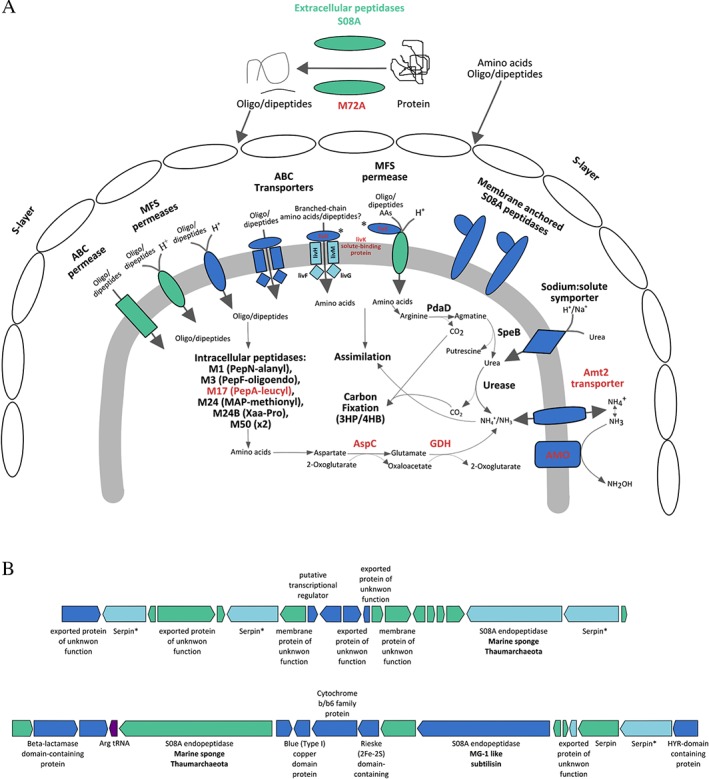
A. Reconstructed metabolic pathways of genes and their expression (red) detected in Ca. N. ianthellae proposed to be involved in extracellular (and intracellular) protein degradation as well as amino acid transport and assimilation. Predicted proteins (and their respective subunits when relevant) are colour coded to denote the degree of homology among all sequenced Thaumarchaeota: green – Ca. N. ianthellae unique gene families; light blue – shared exclusively among thaumarchaeal sponge symbionts; dark blue – ubiquitously found in Thaumarchaeota. Extracellular proteins derived from the marine environment as well as the sponge mesohyl may be degraded extracellularly or in the thaumarchaeal pseudo‐periplasmic space. These resultant oligo/dipeptides and amino acids which can also be derived from the environment can then be transported by a suite of ABC transporters and major facilitator superfamily (MFS; Newstead, 2015 and references therein) permeases into the cytoplasm to be further degraded by intracellular peptidases or assimilated. Amino acids such as arginine and aspartate can be further degraded to form NH3/NH4 + for assimilation or export to the pseudo‐periplasmic space for ammonia oxidation. Arginine can be decarboxylated by arginine decarboxylase (PdaD) to agmatine which can then be degraded to urea by agmatinase (SpeB). Both proteins are ubiquitously distributed among the Thaumarchaeota. All but one of the branched‐chain amino acid transporter subunits (LivFGHMK operon) are exclusively found among thaumarchaeal sponge symbionts, whilst the periplasmic solute binding subunit (LivK – found to be expressed and denoted by an asterisk) can be found not only among sponge symbionts but also in N. maritimus and members of the genus Nitrosocosmicus (see also Fig. 3). B. Genetic context map depicting examples of colocalized S08A endopeptidases and serine protease inhibitors (serpins). Colour coding as in panel (A), except purple denotes the presence of a tRNA, sites at which gene insertions are common. The asterisk next to serpins colour coded as sponge‐specific (light‐blue) denotes that Ca. N. chungbukensis is the sole non‐sponge symbiont encoding a serpin and belonging to this orthologous group (see Supporting Information Fig. S4B). AspC, aspartate aminotransferase; GDH, glutamate dehydrogenase.
Ca. N. ianthellae also encodes 15 genes for the I4 family of serpins, which represent serine protease inhibitors. Some of these cluster with serpins from C. symbiosum (Hallam et al., 2006) and CCThau, although among the non‐host associated AOA, only Nitrosotenuis chungkubensis encodes a serpin (Supporting Information Fig. 4B). Nine of the Ca. N. ianthellae serpins are predicted to be extracellular and are frequently found adjacent to S08A family endopeptidases in the genome (Fig. 4B). Serpins belong to a large family of irreversible inhibitory substrates of proteases, often but not exclusively of the serine class, that has been widely characterized in mammals, insects, plants as well as some viruses, and which can mediate host–microbe interactions (Ventura et al., 2012). These findings suggest that AOA living as sponge symbionts might use serpins to regulate their own secreted endopeptidases and/or sponge serine proteases found to be highly expressed in other sponges (Riesgo et al., 2014), including within the host of C. symbiosum (Zaikova, E., PhD thesis, 2007).
Peptide and amino acid uptake by Ca. N. ianthellae and other sponge AOA
Ca. N. ianthellae, like many other AOA, is well‐equipped for uptake of oligo‐ and dipeptides as well as amino acids by encoding a set of transporters widely distributed in this clade. After uptake, Ca. N. ianthellae, like other AOA, has the genomic repertoire to degrade these peptides and amino acids and release ammonia (Fig. 4). In addition to features common to many AOA, Ca. N. ianthellae also contains the livFGHMK operon which in proteobacteria and cyanobacteria encodes a high‐affinity branched‐chain amino acid transport system (which can also transport other amino acids) belonging to the ATP binding cassette (ABC) superfamily of transporters (Hoshino, 1979; Adams et al., 1990; Hosie et al., 2002; Picossi et al., 2005) (Fig. 4). Among all other thaumarchaeote genomes, only C. symbiosum, CCThau, and a thaumarchaeote MAG from the Caspian Sea (Mehrshad et al., 2016) encode this transporter (Supporting Information Fig. S5). The periplasmic subunit LivK from Ca. N. ianthellae was expressed as protein (Supporting Information Table S3) and this subunit was also reported to be expressed in CCThau (Moitinho‐Silva et al., 2017a). Adjacent to the expressed livK in Ca. N. ianthellae is a gene encoding three consecutive livK domains (along with the requisite ligand binding sites) which is congruent with C. symbiosum containing 4 copies of the livK solute binding component (Supporting Information Fig. S5A). In C. symbiosum, the transporter encoding the livFGHMK operon is located next to several extracellular trypsin‐like serine protease encoding genes, suggesting that the transporter is involved in amino acid uptake in this organism. However, given the substrate promiscuity of this transporter family (Adams et al., 1990; Valladares et al., 2002; Beckers et al., 2004; Picossi et al., 2005) (Supporting Information Fig. S5), experimental validation will be required before more specific predictions on its function in sponge AOA can be made. Such investigations would be particularly important, as multiple livFGHMK operons are also present in two γ‐proteobacterial sponge symbionts (Gauthier et al., 2016), suggesting that amino‐acid transport and utilization mediated by this transporter‐type could be an important feature of sponge symbionts and may even contribute to sponge‐mediated dissolved organic matter transfer to higher trophic levels (de Goeij et al., 2013). Proteases are also important for sponge metabolism and a body of research has focused on sponge proteases and their inhibitors from a biodiscovery context (Arreguín et al., 1993; Wilkesman and Schröder, 2002; Wilkesman and Schröder, 2007; Tabares et al., 2011). In these analyses generally, sponge holobiont samples are used for enzyme or inhibitor purification and characterization. Thus, our observation that abundant archaeal sponge symbionts encode and express putatively exported proteases and serpins, demonstrate that in such assays, it will remain unclear whether it is the microbial symbionts, or the host animals that are producing the analysed biomolecules.
Mixotrophy of Thaumarchaeota
A mixotrophic lifestyle for Thaumarchaeota has been inferred since the earliest reports on planktonic thaumarchaeotes described incorporation of organic carbon into signature lipids (Ingalls et al., 2006) and incorporation of labelled amino acids leucine (Ouverney and Fuhrman, 2000; Herndl et al., 2005) and aspartate (Teira et al., 2006) into single planktonic cells. Whilst initial findings – that pure cultures of Thaumarchaeota in group I.1a (Qin et al., 2014) and group I.1b (Tourna et al., 2011) could have increased growth rates in the presence of α‐keto acids and ammonia – were subsequently proven to be a result of the H2O2 scavenging ability of α‐keto acids (Kim et al., 2016; Qin et al., 2017), abundant genes and transcripts of S08A family serine proteases and other metallopeptidases in MG‐1 deep‐sea thaumarchaea have also been attributed to mixotrophic activity (Li et al., 2015). A deep sea Thaumarchaeota has also been found to encode the livFGHMK operon (Mehrshad et al., 2016), which clusters with the two sponge thaumarchaeotes (Supporting Information Fig. S5), highlighting both environments as potential ecological reservoirs of mixotrophic thaumarchaea. Furthermore, growth of thaumarchaeotes in an industrial wastewater treatment plant that was uncoupled to ammonia‐oxidation has been described (Mußmann et al., 2011). Interestingly, many members of the Aigarchaeota, a sister group to the Thaumarchaeota (Guy and Ettema, 2011), also encode the full livFGHMK operon, and a single cell aigarchaeal‐like genome from cold marine sediments encodes additional extracellular proteases, di‐ or tripeptide transporters and aminotransferases (Lloyd et al., 2013). It is therefore tempting to speculate that the thaumarchaeal ancestor was a mixotroph or even strict heterotroph (as also indicated by genomic and experimental data that deep branching clade I1c and d members in the Thaumarchaeota lack genes required for ammonia oxidation; Beam et al., 2014; Lin et al., 2015; Weber et al., 2015), and that sponge Thaumarchaeota along with a few other members of this clade retained the capability to use amino acids due to specialized environmental conditions. Future experiments would need to demonstrate uptake of amino acids by Ca. N. ianthellae to reveal whether it fuels assimilation or heterotrophic growth and/or is used for sequential intracellular generation and oxidation of ammonia (de Boer and Laanbroek, 1989; Burton and Prosser, 2001).
Eukaryotic‐like proteins (ELPs) in Ca. N. ianthellae
Ca. N. ianthellae encodes a number of genes containing domains postulated to have an evolutionary origin within the eukaryotes and which are thought to be important for modulating interactions between bacteria and eukaryotic hosts (Callebaut et al., 2000; Lurie‐Weinberger et al., 2010; Patterson et al., 2014). Recent metagenomic analyses revealed an abundance of genes encoding such eukaryotic‐like proteins (ELPs) in the bacterial symbionts of sponges (Fan et al., 2012; Reynolds and Thomas, 2016; Díez‐Vives et al., 2017), with many of these being expressed (Díez‐Vives et al., 2017). Four types of ELPs are found in Ca. N. ianthellae: Proteins with tetratricopeptide repeats (TPR), the Toll‐interleukin‐1 receptor (TIR)–like domain PF08937 (DUF1863; Cort et al., 2000; Essuman et al., 2018), immunoglobulin‐like (Ig‐like) domains (DUF5011; Shigeno‐Nakazawa et al., 2016), and hyaline repeats (HYR; Callebaut et al., 2000) and an extensive discussion of these ELPs is provided in the Supporting Information.
Mobile and selfish genetic elements in Ca. N. ianthellae
In contrast to other members of Ca. Nitrosopumilaceae, Ca. N. ianthellae along with the other two sponge AOA, C. symbiosum and CcThau (but not DSGS‐AOA) are enriched in transposases, restriction‐modification (RM) systems (including a Type II restriction endonuclease, PF13156, which is exclusively found in sponge AOA – Supporting Information Fig. S6), toxin‐antitoxin (T‐A) systems, as well as genes putatively involved in DNA phosphorothioation (Fig. 3, Supporting Information Fig. S6). However, no differential enrichment of integrases was detected within sponge‐associated thaumarchaotes. The abundance of transposases and other mobile/selfish genetic elements (MGEs/SGEs) in sponge AOA is consistent with what has been reported for other sponge‐associated microbes (Fan et al., 2012, Horn et al., 2016), suggesting that evolution of AOA sponge symbionts compared to free‐living marine AOA (see also Fig. 3) is more heavily shaped by horizontal gene transfer in the concentrated milieu of environmental bacteria and viruses resulting from sponge feeding and pumping activity.
Among the above‐mentioned genetic elements, the complete genetic repertoire for DNA phosporothioation (PT) (Wang et al., 2007; You et al., 2007) in Ca. N. ianthellae and CcThau (dndA, B, C, D, E) is particularly noteworthy as among cultured AOA, only Ca. Nitrosomarinus catalina (Ahlgren et al., 2017) and Ca. Nitrosopumilus salaria also encode this DNA modification system. These genes possibly encode a primitive immune system by enabling discrimination between self and non‐self DNA (but can also epigenetically affect the transcriptome of bacteria; Tong et al. 2018) and might thus – in concert with the better understood canonical RM systems (Fig. S6) – be particularly useful for sponge symbionts living in a system where they are prone to genetic exchanges. In addition, Ca. N. ianthellae, CcThau and C. symbiosum have genes with low similarity to dndA, B and C respectively. The dndB‐like genes contain domains (DUF262, DUF1524) previously identified as components of R‐M systems (Miller, 2012; Machnika et al., 2015) which are known to be enriched in sponge microbiomes (Fan et al., 2012, Horn et al., 2016).
Ca. N. ianthellae also encodes most enzymes necessary for the production of archaeosine, a highly modified tRNA nucleoside (Phillips et al., 2012) which putatively confers structural stability that is in some archaea important for growth at low temperature (Blaby et al. 2010). Interestingly, among all analysed Thaumarchaeota, the critical enzyme (aTGT) of this pathway is missing the RNA binding site (PUA domain) whilst maintaining the conserved substrate binding pocket found in Crenarchaeota (Phillips et al., 2012). Although the PUA domain is dispensable for archaeosine formation (Sabina and Söll, 2006) it is as yet unknown whether Ca. N. ianthellae can use this for DNA modification, as recently described for another restriction‐modification system variant in a Salmonella species (Thiaville et al., 2016).
Ammonia‐oxidation is exclusively mediated by Ca. N. bastadiens in I. basta and is coupled to carbon fixation
The presence of amoA‐encoding and expressing thaumarchaea does not prove that these microbes actually perform ammonia‐oxidation in a system (Mußmann et al., 2011). Consequently, nitrification rates of I. basta harbouring the thaumarchaeal symbiont were experimentally determined to verify ammonia‐oxidizing activity and to obtain insights into the mean nitrifying activity per symbiont cell. Interpretation of these data were facilitated by the fact that I. basta according to previous 16S rRNA gene based surveys (Webster et al., 2009; Luter et al., 2010; Freckelton et al., 2012; Luter et al., 2012) and our metagenomic and metaproteomic data contains a single AOA symbiont species and does not harbour bacterial ammonia‐oxidizers or comammox organisms (Daims et al., 2015). Incubation experiments with freshly collected sponge clones were performed in the presence of different ammonium concentrations ranging from ambient seawater (0.29 ± 0.1 μM) via 25 to 100 μM. Gross and net nitrification rates as well as net fluxes of ammonium, nitrite and nitrate were determined in 24 h laboratory incubation experiments that were repeated several times over 7 days (Supporting Information Fig. S7). Net nitrification rates were inferred from the measurable increase of the nitrification products nitrite and nitrate, but can underestimate the actual nitrification rate if these products are concurrently consumed in the system. Gross nitrification rates were determined by the isotope pool dilution technique that directly measures the total gross nitrite and nitrate formation and is not affected by concurrent consumption of these compounds (for details see experimental procedures). Across all treatments, gross and net nitrification rates were similar in magnitude and highly correlated (gross rates = 1.13 × net rates +0.6; R2 = 0.94; p < 0.01), suggesting that nitrate removal (e.g. via assimilation or denitrification) did not occur at significant rates, with the exception of sponges in the 100 μM NH4 + treatment (Fig. 5A). Gross nitrification rates of sponges incubated with 25 or 100 μM NH4 + were significantly greater than in ambient seawater (p < 0.05; ANOVA followed by Tukey HSD Test) (Fig. 5A). The stimulation of nitrification by increased ammonium availability, which has also been observed for other sponge species (Corredor et al., 1988; Bayer et al., 2007; Bayer et al., 2008; Schläppy et al., 2010), indicates ammonium limitation of the symbiotic nitrifiers under ambient conditions. Almost no net ammonium release was observed from the I. basta holobiont in unamended seawater, indicating similar in situ rates of ammonium production and consumption (Fig. 5B). Net nitrification rates of the I. basta holobiont under ambient experimental conditions were slightly higher but generally comparable to those reported for other sponge species (Supporting Information Table S4; to the best of our knowledge no gross nitrifications reports for sponges have been published). These data likely reflect nitrification rates under natural field conditions, as NH4 + concentrations in the unamended treatment in the aquaria experiments were consistent with those reported for Orpheus Island, where I. basta was collected (Jompa and McCook, 2002).
Figure 5.

Nitrification activity of the I. basta holobiont during 7‐day incubation experiments at ambient conditions and with added ammonium (25 or 100 μM) and with/without the AOA inhibitor PTIO. A schematic overview of the corresponding experimental setup is given in Supporting Information Fig. S7. A. Depicts net nitrification (as calculated from the addition of net NO2 − and NO3 − flux) and gross nitrification rates (estimated using the 15NO3 − isotope pool dilution method). B. Net fluxes of all DIN species for the ambient, +25 μM NH4 + and +100 μM NH4 + treatments. Rates of the different days in the ambient and +25 μM NH4+ incubations were averaged as they were not statistically different. C. The relationship between gross nitrification rates of the I. basta holobiont and the carbon fixation rate of sponge clones sampled on day 7. The carbon fixation rates derived from the I. basta holobiont nitrification experiments displayed a positive and significant correlation with gross nitrification rates (R = 0.665, p < 0.005). Error bars in panel C reflect the standard error of the sample mean, where n = 3 for the carbon fixation rates and for all weighted gross nitrification rates in all treatment conditions. PTIO additions started 2 days after the experiment was commenced; hence, fixation of 13C‐labelled bicarbonate during the first 2 days was not influenced by inhibition.
Net nitrification rates in sponges subjected to 100 μM NH4 + for 24 h were significantly lower than gross nitrification rates (p < 0.05; two‐tailed t‐test, Fig. 5A), indicating increased nitrate consumption most likely by denitrification by other sponge holobiont members (in this context it should be kept in mind that oxygen concentration will decrease if nitrification is stimulated by ammonia addition). Gross and net nitrification rates also became significantly depressed after prolonged exposure (> 3 days) to 100 μM NH4 +, being significantly lower than the 24 h incubations at 100 μM NH4 + and the ambient seawater controls (p < 0.01, Mann–Whitney U‐test) (Fig. 5A). From AOA pure culture studies, there are no indications that 100 μM NH4 + can be inhibitory for members of this clade, with concentrations of 2 to 20 mM NH4 + needed for inhibition (Hatzenpichler et al., 2008; Martens‐Habbena et al., 2009; Tourna et al., 2011; Li et al., 2016; Sauder et al., 2017; Sauder et al., 2018). Whilst we cannot exclude that Ca. N. ianthellae is particularly sensitive to ammonium, it is also possible that the inhibition was caused by indirect effects. For example, if the sponge host was adversely affected at 100 μM NH4 +, decay of sponge cells could cause stimulation of heterotrophic microbes and thus oxygen limitation. Typical acute toxicities in freshwater fish and invertebrates (96‐h LC50) are reached at unionized ammonia (NH3) concentrations between 4.7 and 38 μM (with salinity typically having a slight ameliorating effect and tests on marine invertebrates very rare; Boardman et al., 2004) and susceptibility to ammonia toxicity can be increased by low O2 concentrations (Camargo and Alonso, 2006). At ambient incubation conditions about 4.5% of total ammonia was present as NH3, hence I. basta may have experienced physiological stress and intermittent hypoxic conditions in the 100 μM NH4 + treatments.
PTIO, a scavenger of free radical nitric oxide (NO) (Amano and Noda, 1995; Ellis et al., 2001), has been described as a specific inhibitor of AOA (Yan et al., 2012; Martens‐Habbena et al., 2015) and was therefore used to demonstrate that Ca. N. ianthellae is responsible for ammonia‐oxidation in I. basta. As expected, the addition of PTIO to sponge incubations in the ambient and 25 μM NH4 + amended treatments, resulted in significantly depressed gross and net nitrification rates and increased ammonium accumulation when compared to the non‐PTIO treated sponge incubations (p < 0.05; except for gross nitrification rates under ambient conditions, p = 0.885; Mann–Whitney U‐test; Fig. 5A,B). In the case of the 100 μM NH4 + amendment, gross and net nitrification was already strongly reduced in the absence of PTIO after 3 days of incubation, so further inhibition by PTIO could not be demonstrated under these conditions (Fig. 5A). Residual ammonia oxidizing activity in some of the experiments in the presence of 75 μM PTIO was unexpected as pure AOA cultures are known to be fully or almost completely inhibited by this concentration of PTIO (Shen et al., 2013; Martens‐Habbena et al., 2015). NO production by I. basta (many sponges express NO synthases, Riesgo et al., 2014 ) may have contributed to PTIO inactivation thereby lowering its inhibitory effect on the AOA. To our knowledge, PTIO has not previously been used in sponge microbiome research (but is commonly applied to eukaryotic tissues including sponges; Müller et al., 2006; Ueda et al., 2016) and further optimization of the concentration is recommended for future applications in these animals. Study design of such experiments should also take into consideration that PTIO has been shown to be cytotoxic for some microbes (Kits et al., 2019).
In 29 of the 33 sponge incubations (incubations with added PTIO or inhibited by addition of 100 μM NH4 + were excluded) (Fig. 5B), accumulation rates of NO2 − exceeded those of NO3 − by an average factor of 2.8 and this difference was found to be significant in the ambient and 25 μM NH4 + treatments (both p < 0.01, Mann–Whitney U‐test). In contrast, no significant difference between NO2 − and NO3 − production was observed in the PTIO‐amended incubations nor in those experiments where prolonged exposure to 100 μM NH4 + negatively affected net nitrification rates (all: p > 0.05, Mann–Whitney U‐test).
In addition, several control experiments were performed in order to better understand the influence of the experimental setup and the seawater microbial community on our results. All nitrification experiments discussed above were performed in intermittently closed aquaria (Supporting Information Fig. S7) to prevent loss of labelled CO2 but to provide sufficient oxygen exchange to maintain sponge health. Control experiments were performed at ambient and 100 μM NH4 +, to compare DIN fluxes and inferred net nitrification rates using this setup, to incubations performed in constantly open containers. Interestingly, slightly but significantly higher NO2 − fluxes and net nitrification rates were observed with ambient seawater in the intermittently closed aquaria (both p < 0.01, two‐tailed t‐test; Supporting Information Fig. S8), indicating that ammonia‐oxidizers in the closed system either benefited from the minimized loss of the added bicarbonate (via CO2 off gassing) or from potentially increased ammonia production from stressed sponge clones. Furthermore, we determined the contribution of the seawater microbial community to net nitrification by using aquaria seawater without sponges. These experiments revealed minimal nitrification with net rates of only 0.06 ± 0.3 μM N d−1 (n = 12).
To calculate cell specific ammonia oxidation rates for Ca. N. ianthellae, its 16S rRNA genes were quantified at the end of the 7 day sponge incubation under ambient and +25 μM NH4 + treatments. Sponges subjected to ambient and +25 μM NH4 + treatments contained thaumarchaeal symbiont gene copy numbers of 8.58 ± 4.9 × 1010 and 1.64 ± 0.12 × 1010 per g wet weight (SE, n = 3, for both) respectively. By dividing the net nitrification rate on day 7 by the 16S rRNA gene copy number of Ca. N. ianthellae on this day, average cell specific rates were estimated to be 0.11 ± 0.08 and 0.66 ± 0.09 fmol NH4 + oxidized per cell per day (SE, n = 3, for both) for the ambient and +25 μM NH4 + incubations respectively. Inferred cell specific ammonia oxidation rates for Ca. N. ianthellae were ~20–120 times lower than what has been reported for N. maritimus (Martens‐Habbena et al., 2009) and ~6–65 times lower than coastal marine seawater (Wuchter et al., 2006), whilst the highest ammonia oxidation rate of Ca. N. ianthellae was comparable to the lowest rates detected in the sponge Phakellia ventilabrum (Radax et al., 2012a). The relatively low cell specific ammonia oxidation rate might reflect that not all I. basta AOA symbiont cells are physiologically active (or even in the process of digestion by the host) or that the sponge host exerts some control over the ammonia oxidation of the symbiont.
Furthermore, as 13C‐bicarbonate was added during all experiments (Supporting Information Fig. S7), we were able to measure the δ13C values of sponge tissues and calculate inorganic carbon fixation rates to compare with the gross nitrification rates. PTIO was found to reduce the δ13C value in both the ambient and + 25 μM NH4 + treatments (Supporting Information Fig. S9; both: p < 0.05, one‐tailed t‐test), whilst no significant differences were observed with the +100 μM NH4 + treatments. In addition, gross nitrification rates and carbon fixation were significantly positively correlated (Fig. 5C). These data suggest that Ca. N. ianthellae contributes significantly to carbon fixation by the sponge holobiont. This finding is consistent with (i) the detection of key genes for CO2 fixation in the I. basta thaumarchaeote metagenome bin, (ii) the detection of some of the respective proteins in the metaproteome, and (iii) the absence of autotrophic CO2 fixation pathways in the metagenomic bins of the alpha‐ and gamma‐symbionts (data not shown). In addition, the slope of the positive linear relationship between the gross nitrification rate and the carbon fixation rate (Fig. 5C; 43.0), reflects the gross nitrification: carbon fixation ratio (N:C ratio), which is most likely over‐estimated since the experimental setup of the intermittently closed aquaria still allowed for the loss of added 13C‐bicarbonate. In addition, CO2 production derived from respiratory activities within the I. basta holobiont were not accounted for, which may also lead to an underestimation of carbon fixation rates. Despite these possible biases the N:C ratio determined for I. basta is within the range typically found for aquatic environmental samples (2–60) (Andersson et al., 2006), but higher than the values we have inferred for N. maritimus (~7–29; Könneke et al., 2005; Martens‐Habbena et al., 2009). The relatively high N:C ratio along with the low cell‐specific ammonia‐oxidation rates within the I. basta holobiont are consistent with the proteogenomic‐derived hypothesis that Ca. N. ianthellae is not sustaining its population only by chemolithoautotrophic growth on ammonia, but more likely grows as a mixotroph.
Surprisingly, nitrite‐oxidizing bacteria (NOB) could not be detected in I. basta by amplicon sequencing, metagenomic sequencing, or with FISH. Consistent with the absence of NOB, NO2 − accumulated to high concentrations ranging from 8 to 21 μM in ambient treatments containing I. basta (Fig. 5B, Supporting Information Fig. S8), making this sponge one of the few natural systems in which greater NO2 − than NO3 − concentrations occur (Brezonik and Lee, 1968; Lam et al., 2011; Schaefer and Hollibaugh, 2017 and refs. therein). Whilst nitrite accumulation might have been more pronounced in the closed aquaria system than in the open ocean, we have shown before that strong nitrite gradients do occur in nitrifying open biofilm systems and possibly affect microbial community structure (Maixner et al. 2006). It will be interesting to explore in future studies how high the nitrite concentration is within I. basta and whether nitrite production contributes (via nitrite toxicity; Camargo and Alonso, 2006 and references therein) to protection from predators or to the unusually low microbial diversity in I. basta.
Given the apparent absence of NOB in I. basta, the observed production of nitrate (Fig. 5) is difficult to explain. One possibility is that some NO released by either the host or a host‐associated microorganism is detoxified to nitrate by another member of the sponge holobiont. However, we did not detect genes with homology to those encoding the NO detoxifying enzyme, flavohemoglobin‐NO‐dioxygenase (Hmp), which converts NO together with O2 to NO3 − (Gardner, 2012 and references therein), in our microbial metagenomic datasets. Still, NO dioxygenases (NODs), could be encoded in the sponge genome since neuroglobin‐like sequences are encoded in other sponges (Lechauve et al., 2013) and members of this enzyme family have been shown to have NOD activity in vitro (Brunori et al., 2005). Furthermore, a NADH‐cytochrome b5 reductase, which belongs to the same family as Hmp, was found to be highly expressed at all life stages of the sponge Amphimedon queenslandica (Conaco et al., 2012). Alternatively, partial nitrite oxidation may be catalysed by free‐living NOB in the seawater, although nitrification rates in the control were negligible.
Conclusions and outlook
In this study, we combined proteogenomic and experimental analyses to show that Ca. N. ianthellae, the first characterized but yet uncultured representative of a new genus within the thaumarchaeotes, is responsible for ammonia oxidation in the widespread marine sponge I. basta. Whilst Ca. N. ianthellae is equipped with the typical genetic repertoire of free‐living AOA, it also exhibits a number of putative adaptations to a host‐associated lifestyle. Several of these adaptive features were unique to Ca. N. ianthellae, whereas others were shared exclusively with the previously described sponge symbionts Ca. C. symbiosum (Hallam et al., 2006) and the AOA symbiont of the sponge C. concentrica (Moitinho‐Silva et al., 2017a). Many of these putative adaptations to a sponge‐associated lifestyle were not encoded in the recently sequenced genome of an AOA from a deep‐sea glass sponge (DSGS‐AOA; Tian et al., 2016), indicating that this archaeon is not an obligate symbiont or that thriving in a deep‐sea glass sponge requires very different traits. Our results confirm an emerging view that marine sponge microbiomes tend to converge on a few shared functional traits, a process shaped by the environmental niche provided by the sponge host and governed by specific ecological factors such as high dissolved nutrient loads and frequent contact with resident and transient microorganisms (Liu et al., 2012). Whereas other studies have focused on this phenomenon by comparing functional convergence across entire sponge microbiomes (Fan et al., 2012; Liu et al., 2012; Horn et al., 2016), we demonstrate that in symbiotic marine sponge Thaumarchaeota, a similar evolutionary convergence is achieved that stands in contrast to their strictly chemoautotrophic non‐symbiotic free‐living relatives and that this process is likely not solely achieved by gene acquisition but rather by selective gene retention and gene family expansion.
With I. basta emerging as a model species for sponge symbiosis research, additional work should be undertaken to address the conspicuous absence of nitrite‐oxidizing bacteria and ascertain why, in contrast to most tropical sponge species, I. basta hosts such a low diversity of microbial symbionts. Furthermore, hypotheses about the interaction of Ca. N. ianthellae with other members of the sponge holobiont should be experimentally confirmed. For example, assimilation of amino acids and peptides should be tested using stable isotope probing and the function of unique serine proteases, serpins and ELPs should be analysed via heterologous gene expression to reveal mechanistic insights into the interaction of this archaeon with its host. Finally, the mechanism for symbiont acquisition should be assessed via screening for Ca. N. ianthellae in gametes and larvae.
Experimental procedures
Sponge collection
Large adult specimens (n = 4) of the sponge Ianthella basta were collected from Orpheus Island (18°36.878′S, 146°29.990′E), Queensland, Australia, cut into 10 cm x 10 cm explants and transferred to racks on the reef. After a 12 week healing period in the field, sponge clones were collected in two separate sampling trips by scuba diving between September and October 2011 and transported to the indoor temperature‐controlled aquarium at the Australian Institute of Marine Science (AIMS), Townsville, where they were acclimated at ambient temperature (25°C) for 48 h and then randomly assigned to experimental treatments. The two adult I. basta specimens used for metaproteogenomic analyses were collected from Orpheus Island in October 2010 and 2011 at Orpheus Island. Upon sample collection, specimens were cut into small strips, immediately snap‐frozen in liquid N2 and subsequently stored at −80°C until DNA extraction.
Sponge incubations and nitrification rate measurements
To infer nitrification activity of the I. basta holobiont, a 7‐day incubation experiment was performed using triplicate sponge clones under different ammonium concentrations: ambient (0.29 ± 0.1 μM), 25 and 100 μM NH4 +. A schematic representation of the experimental design is given in Supporting Information Fig. S7. Specifically, sponge clones (1.5–13.0 g post‐experimental wet weight; mean = 5.1; standard deviation = 2.2) were incubated at 25°C in the dark in 1.5 l acid‐washed glass containers completely filled with 5 μm filtered seawater or with seawater amended with 25 or 100 μM NH4Cl. Additionally, abiotic control experiments were performed with ambient seawater without sponge clones that were incubated for 24 h under identical conditions. Sponge clones were transferred to a new container with fresh seawater every 24 h during the 7‐day incubation, in order to reduce the effects of O2 depletion and NH4 + accumulation. To assess carbon fixation in concert with nitrification, NaH13CO3 − (100 μM; 99% 13C) was added every 12 h to all experimental treatments. To prevent loss of 13C‐labelled CO2, containers were closed for 6 h after the NaH13CO3 − addition and subsequently opened until the next NaH13CO3 − addition to avoid oxygen depletion. The effect of such intermittently closed containers on nitrification activity was assessed in a control experiment by comparing ambient seawater and 100 μM NH4Cl amended seawater in constantly open containers (Supporting Information Fig. S8). Nitrification activity was determined as gross nitrification rates using a 15N isotope pool dilution technique (Inselsbacher et al., 2007), and net fluxes of individual dissolved inorganic N species (DIN), namely NH4 +, NO2 −, NO3 −. Gross nitrification rates were also measured on day 1, 3 and 5, where at the beginning of each day K15NO3 was added to a final 15NO3 − concentration of 1% – 10% of the nitrate pool (Murphy et al., 2003) in the filtered ambient seawater. Gross nitrification rates were calculated from seawater samples collected after a short equilibration following 15N‐label addition (0 h) and after 20 h. Net fluxes of NH4 +, NO2 − and NO3 − were measured on day 1, 3, 5 and 7 and were calculated as change in concentration over time (i.e., between 6 and 18 h after 15N‐label addition to avoid biased net NO3 − fluxes through a stimulation of consumptive processes from the added 15NO3 −). Net nitrification refers to the change in concentration of NO2 − + NO3 −. Furthermore, we used the AOA‐specific inhibitor PTIO (2‐phenyl‐4,4,5,5‐tetramethylimidazoline‐1‐oxyl 3‐oxide; Tokyo Chemical Industry) (Martens‐Habbena et al., 2015) in additional parallel incubations, which was added at a concentration of 75 μM at the beginning of day 3 and 7.
Seawater samples (10 ml) were taken from each aquarium and filtered using 0.45 μm Sartorius Minisart cellulose acetate filters (Göttingen, Germany). Duplicate samples for dissolved inorganic nitrogen (NH4 +, NO2 −, NO3 −) were measured on a Seal AA3 segmented flow analyser and referenced against OSIL standards and in‐house reference samples. For 15N‐analysis of NO2 − + NO3 −, sample water was filtered through pre‐combusted GF/Fs (Whatman International; treated for 4 h at 450°C), and subsequently through 0.2 μm filters (Sartorius). All samples were immediately frozen at −20°C for later analysis. Prior to shipment to the University of Vienna, samples were thawed at room temperature and the microbial inhibitor pheynylmercuric acetate was added (to a final concentration of 10 μM). Upon arrival in Vienna the samples were promptly stored at −80°C. Nitrite and nitrate were isolated together from seawater by sequential microdiffusion (Sørensen and Jensen, 1991). To remove ammonium from sample water, 100 mg MgO, and an acid trap (acidified cellulose filter disc enclosed in a semi‐permeable Teflon membrane) was added to 9 ml of sample and 1.5 ml of 3 M KCl. After 5 days shaking at 35°C, the acid traps were removed, and 50 mg of Devarda's alloy was added along with a new acid trap, and shaken at 35°C for 7 days. Devarda's alloy is a reducing catalyst converting both NO2 − and NO3 − to NH4 + and the subsequently formed NH3 was collected in the acid trap. Acid traps were dried over concentrated sulfuric acid and analysed for 15N by an elemental analyser (EA 1110, CE Instruments, Milan, Italy) coupled to an isotope ratio mass spectrometer (IRMS; Finnigan MAT DeltaPlus IRMS with a Finnigan MAT ConFlo III interface). Gross nitrification rates were calculated based on Wanek et al. (2010). Gross and net rates are expressed as μmol nitrogen species per gram wet weight sponge per day (μmol N g−1 d−1).
For the determination of 13C enrichment in whole sponge tissue at the end of the incubation, sponge tissue was freeze‐dried, ground to a fine powder, and stored at dry conditions prior to analysis. The δ 13C values of sponge tissue were determined using an EA‐IRMS system as described above. Carbon fixation rates were calculated based on the δ 13C values treatments amended with 100 μM NaH13CO3 −. For the calculations, we used a background (natural abundance) bicarbonate concentration of 1975 μM.
DNA extraction from whole sponge tissue and qPCR for symbiont quantification
Between 80 and 150 mg of I. basta tissue was thawed, rinsed successively (3x) in 1X calcium‐ and magnesium‐free artificial seawater (CMF‐ASW) and immediately ground into a paste with a mortar and pestle in liquid N2. After resuspension in TE buffer (10 mM Tris–HCL, pH 8.0, 1 mMEDTA), DNA was extracted from the suspension using an adapted SDS‐based isolation method (Zhou et al., 1996) and using 1% polyvinylpyrrolidone. DNA was extracted from the two individuals used for metaproteogenomics (see below) as well as three healthy individuals from a previous study (Luter et al., 2010). Additionally, DNA was extracted from a subset of sponge clones that were subjected to nitrification incubations. Quantitative PCR (qPCR) was used to estimate the number of specific thaumarchaeal, as well as α‐ and γ‐proteobacterial symbionts by quantifying the 16S rRNA gene using specific primers designed for each symbiont phylotype. The following primer sets were thus used for the dominant I. basta thaumarchaeal, and α‐, and γ‐proteobacterial symbionts respectively; IBthaum16S_523F, 5′‐CCG TAG CCT GCC CTG TTA G‐3′, IBthaum16S_727R, 5′‐GCT TTC ATC CCT CAC CGT‐3′; IBalpha16S_1010F, 5′‐CGG AGA CGC TTC CTT CG −3′, IBalpha16S_1206R, 5’‐GCC CAG CCC ATA AAT GC‐3′; IBgamma16S_466F, 5′‐TAC CCY TGY GTT TTG ACG‐3′, IBgamma16S_655R, 5′‐ CCR CTT CTC TCT RCC ATA C‐3′. An iCycler real‐time PCR system (Bio‐Rad, Hercules, CA) was used to measure all samples, in duplicate wells per reaction and reactions were performed in a 25 μl volume with 1 μl of DNA template. All symbiont 16S rRNA gene qPCR assays used SYBR®Green reaction mixtures containing 12.5 μl iQ SYBR®Green Supermix (Bio‐Rad), and optimized concentrations of 400 nM primer as well as 0.25 mg ml−1 BSA. Cycling conditions were 95°C for 5 min followed by 40 cycles of 95°C for 40 s, 58°C for 30 s, and 72°C for 40 s. Fluorescence intensities were measured after each cycle, and a final elongation at 72°C was followed by a melting curve analysis from 55–95°C in 10 s increments of 0.5°C.
Standard curves were generated for each primer set using serial dilutions of a standard containing a known number of the target sequences. Standards used the M13 primer set to amplify 16S rRNA gene clones derived from the sponge symbionts. PCR products were visualized on an agarose gel, purified separately using the QIAquick PCR Purification Kit (Qiagen), followed by fluorometric quantification of DNA concentrations using PicoGreen (Molecular Probes, Eugene, OR) and a NanoDrop ND‐3300 Fluorospectrometer (NanoDrop). Gene abundance was calculated based on DNA concentration and product size. Dilution series ranging from 106 to 100 copies μl−1 were used to generate standard curves. Final 16S rRNA gene abundances for each microbial symbiont were then normalized by the wet weight of the sponge tissue used for DNA extraction.
Cryosectioning and FISH for symbiont quantification
The I. basta individual collected for metagenomic sequencing in October 2010 was also assessed using FISH. Briefly, after sample collection, the I. basta specimen was cut into tissue strips of ~2 mm3, fixed in 4% PFA for 1 h at room temperature and stored in ethyl alcohol (EtOH)‐phosphate‐buffered saline (PBS) at −20°C. For FISH, PFA‐fixed samples of I. basta were embedded in Neg‐50 (Richard‐Allan Scientific), and cut to 5‐μm sections (Leica CM3050 S). A double‐labelled Arch915 probe in Cy3 (Thermo Fisher Scientific, Waltham, MA, USA) was used for the microscopic visualization and quantification of the thaumarchaeal symbiont of I. basta. To calculate the relative abundance of the thaumarchaeal symbiont, equimolar amounts of the double‐labelled probes EUB338‐I, EUB338‐II, and EUB338‐III (Fluos and Cy5) (Stoecker et al., 2010) were used for quantification of most bacteria. Hybridizations were prepared with an equimolar mixture of both probes and using 25% formamide in the hybridization buffer, with the stringency of the washing buffer adjusted accordingly (Daims et al., 2001). As a negative control, the non‐EUB338‐I (reverse complementary probe to EUB338‐I) was applied on one section per well per slide hybridized (Wallner et al., 1993). All hybridized samples were analysed with a confocal laser scanning microscope (CLSM) (LSM 510 Meta; Zeiss, Oberkochen, Germany). Archaeal and bacterial cells were counted by eye on 10 randomly selected images derived from multiple tissue sections obtained from a single I. basta individual, and the proportion of archaeal cells to total prokaryotic cells was calculated.
Microbial cell enrichment for metaproteogenomics
To separate symbiont cells from host tissue prior to DNA extraction, ~15.7 g wet weight of I. basta was rinsed successively (3x) in 1X CMF‐ASW. Sponge tissue was cut into small pieces (<1 cm3), ground on ice in 1X CMF‐ASW with a mortar and pestle, transferred into a glass douncer on ice, and sponge tissue was dissociated through shear force and vortexing. The supernatant was transferred into multiple Eppendorf microcentrifuge tubes, centrifuged at 39 x g for 15 min at 4°C to pellet larger sponge particles, filtered through a 5 μm Sartorius filter and centrifuged at 10,844 × g for 15 min at 4°C to pellet microbial cells. Microscopic examination of the pellet revealed an enrichment of microbial cells and an absence of sponge nuclei. For the sponge individual sampled in October 2010, DNA was extracted from this pellet using the procedure referenced above. For the individual sampled in October 2011, the pellets were resuspended in 1X CMF‐ASW and the suspension was layered in 1.8 ml amounts onto 6.5 ml cushions of 30% Gastrografin dissolved in 1X CMF‐ASW + 0.2 M EDTA, and centrifuged at 40,008 x g for 1 h at 4°C in a Beckman L‐100 XP ultracentrifuge with the SW 41 Ti swing rotor (Beckman Coulter). Following centrifugation, a cell‐rich layer above the 30% Gastrografin cushion (‘Fraction A’) and the cell‐rich pellet (‘Fraction B’) were carefully removed, resuspended in 10 mM Tris, and re‐pelleted at 24,400 × g for 15 min at 4°C in a new solution of 10 mM Tris.
The same I. basta individual sampled in October 2011 was used for proteomic analyses. However, in order to minimize protein degradation, biomass preparation methods were shortened and modified. The first cell fraction used for proteomic analysis involved rinsing in 1X CMF‐ASW and tissue homogenization using a mortar/pestle and glass douncer as described above. However, the tissue homogenization steps occurred in 1X TE buffer with Roche Complete Protease Inhibitor (Roche). Supernatant from the glass douncer was immediately collected into 2 ml Eppendorf tubes and frozen at −80°C before shipment on dry ice to Greifswald, Germany, for protein extraction and downstream analyses. A second cell fraction comprised a 5 μm filtrate from the supernatant in the glass douncer. Finally, a crude sponge homogenate sample was obtained by direct grinding of sponge tissue (without rinsing in 1X CMF‐ASW) in liquid N2 followed by freezing at −80°C in 1X TE with Roche Complete. These three fractions could thus be characterized as a sponge homogenate without sponge skeleton, a sponge homogenate without sponge nuclei, and a direct sponge homogenate respectively.
DNA extraction, library preparation and sequencing
DNA was extracted from the individual sampled in October 2011 using the FastDNA spin kit for soil (MP Biomedicals, Solon, OH, USA) from the cell‐rich layers above the 30% Gastrografin cushion and in the ultracentrifuged pellet according to the manufacturer's instructions. Sequencing libraries were prepared using the Nextera kit (Illumina) according to the manufacturer's instructions and concentrations measured using the QuantIT kit (Molecular Probes, Life Technologies, Naerum, Denmark). The libraries were paired‐end (2 × 150 bp) sequenced on an Illumina HiSeq2000 using the TruSeq PE Cluster Kit v3‐cBot‐HS and TruSeq SBS kit v.3‐HS sequencing kit and on an Illumina MiSeq using v3 2 × 300 bp kits.
For the individual sampled in October 2010, cell pellets were pooled from the 5 μm filtration step and DNA was extracted using the modified protocol of Zhou et al. (1996) described above. Metagenomic sequences were then generated at the Ramaciotti Sequencing Centre (Sydney, Australia) using the GS FLX instrument using Titanium chemistry (Roche) on a 454 half‐sequencing‐plate (454 Life Sciences, Branford, CT, USA).
Metagenome assembly and genome binning
Metagenome reads in fastq format, obtained from the Illumina sequencing runs, were end‐trimmed at a minimum phred score of 15, a minimum length of 50 bp, allowing no ambiguous nucleotides and Illumina sequencing adaptors removed. Trimmed reads from each dataset were assembled using Spades version 3.11.0 (Bankevich et al., 2012), using default parameters and genomes were binned using Metabat v 2.12.0 (Kang et al., 2015). MAGs from multiple Illumina data sets were dereplicated using dRep (Olm et al., 2017) and provisionally classified using CheckM (Parks et al., 2015). A single high‐quality archaeal MAG was recovered after dereplication and uploaded to MaGe (Vallenet et al., 2009) for annotation. This MAG has been submitted to the European Nucleotide Archive with the accession number UYNY01000000 under BioProject PRJEB29556.
For the 454‐pyrosequencing run, artificially amplified reads were dereplicated using CD‐HIT (Li and Godzik, 2006), assembled with MIRA (Chevreux et al., [Link]) and binned with a genome‐specific Phymm model (Brady and Salzberg, 2009), which was trained from the contigs that contained phylogenetic marker genes. The MAG derived from this pyrosequencing run was similarly GC‐rich (64.2%), but considerably larger (6.58 Mb), slightly less complete (98.06%), and considerably more fragmented (2508 scaffolds). This MAG is available on the MaGe platform as ‘Ianthella basta symbiont thaum’ at http://www.genoscope.cns.fr/agc/microscope/home/index.php.
Comparative genomics
The annotated archaeal MAG was downloaded from MaGe and compared to published thaumarchaeotal genomes (Supporting Information Table S5) using genomic average nucleotide identity (gANI), average amino acid identity (AAI), and through construction of orthologous gene families. For all analyses, annotated genes were supplemented with additional gene calls predicted by Prodigal (Hyatt et al., 2010). gANI was calculated with MiSI (Varghese et al., 2015). AAI (Konstantinidis and Tiedje, 2005) was calculated using bidirectional best blastp hits (Camacho et al., 2009) that aligned over at least 70% of gene length with average identity values weighted according to gene length. Orthologous gene families were constructed using Orthofinder (Emms and Kelly, 2015). For functional annotation of eukaryotic‐like proteins (ELPs) and mobile/selfish genetic elements, predicted genes from all sequenced thaumarchaeal genomes were searched against the Protein Family A (Pfam‐A) database (v31.0) (Finn et al., 2014) using Hmmer 3 (http://hmmer.org.) and the gathering threshold option (−cutga). Results were screened for the presence of domains associated with ELPs and mobile/selfish genetic elements.
Phylogenetic analyses
Bayesian trees were constructed using Phylobayes v 4.1c (Lartillot and Philippe, 2004) using the best model identified for each dataset by ModelFinder (Kalyaanamoorthy et al., 2017). Phylogenomic reconstruction was based on a concatenated amino‐acid alignment of 34 marker genes constructed with CheckM (Parks et al., 2015) with 10 independent runs of 11,000 generations under the LG4 model. Six thousand generations of each independent run were discarded as burn‐in and the remaining trees from each run were pooled for calculation of a consensus tree and for determining posterior branch support. For the 16S rRNA gene phylogenies, top representative hits in a blastn query against the Genbank nr database of the full‐length sequence from Ca. N. ianthellae and Ca. C. symbiosum, along with sequences from sequenced thaumarchaeal genomes were aligned with SINA (Pruesse et al., 2012) and analysed further as described above. Ten independent runs of 30,000 generations under the GTR model were used. Then, 7500 generations of each independent run were discarded as burn‐ins and the remaining trees from each run were pooled for calculation of a consensus tree and for determining posterior branch support.
Protein identification and proteome analyses
1D PAGE followed by liquid chromatography‐based mass spectrometry (1D‐PAGE‐LC–MS/MS) were used for protein and peptide separation and identification as described previously (Washburn et al., 2001; Otto et al., 2010), with slight modifications. MS spectra and MS/MS spectra were acquired from eluting peptides ionized with electrospray ionization (ESI) and analysed in a LTQ Orbitrap Velos hybrid mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA), as described previously (Verberkmoes et al., 2009; Otto et al., 2010), with minor modifications. Samples of the sponge homogenate without sponge skeleton and crude sponge homogenate processed in liquid N2 were analysed in technical duplicates, whereas the sponge homogenate without sponge nuclei was analysed only once. All MS/MS spectra were searched against predicted protein sequence databases composed of the I. basta symbiont‐enriched metagenome bins and common laboratory contaminants using the Sorcerer SEQUEST (v.27, rev. 11) algorithm. The CD‐HIT software (Li and Godzik, 2006) was used to remove redundancies from the database due to the potential for strain‐level redundancies. Protein identifications were filtered with Scaffold version 3.5.1 applying the ‘sequest’ filter criteria described previously (Heinz et al. 2012). For protein identification only peptides identified with high mass accuracy (maximum ±10 ppm difference between calculated and observed mass) were considered and at least two exclusively unique peptides were required to identify a protein. False‐discovery rates (FDRs) were estimated with searches against a target‐decoy database as described previously (Peng et al., 2003; Elias and Gygi, 2007). Peptide FDRs were between 2.5% and 3.1%, and protein FDRs were below 0.4% throughout all samples. For relative quantitation of proteins, normalized spectral abundance factor were calculated for each sample according to the method of Florens et al. (2006) and averaged for all replicates and samples. The proteomics raw data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the data set identifier PXD012484 and 10.6019/PXD012484.
Supporting information
Appendix. Supporting Information.
Acknowledgements
This work was supported by the Marie Curie Initial Training Network ‐ SYMBIOMICS. In addition, F.U.M., C.W.H, M.M. and M.W. were supported by the European Research Council Advanced Grant project NITRICARE 294343. M.A. and P.H.N. were supported by research grants (15510 and 16578) from VILLUM FONDEN. Many thanks to Christian Hentschker for Orbitrap Velos measurements and database searches. We thank Bernhard Schink for valuable advice regarding provisional naming of the I. basta AOA symbiont.
References
- Adams, M.D. , Wagner, L.M. , Graddis, T.J. , Landick, R. , Antonucci, T.K. , Gibson, A.L. , and Oxender, D.L. (1990) Nucleotide sequence and genetic characterization reveal six essential genes for the LIV‐I and LS transport systems of Escherichia coli . J Biol Chem 265: 11436–11443. [PubMed] [Google Scholar]
- Agarwal, V. , Blanton, J.M. , Podell, S. , Taton, A. , Schorn, M.A. , Busch, J. , et al (2017) Metagenomic discovery of polybrominated diphenyl ether biosynthesis by marine sponges. Nat Chem Biol 13: 537–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahlgren, N.A. , Chen, Y. , Needham, D.M. , Parada, A.E. , Sachdeva, R. , Trinh, V. , et al (2017) Genome and epigenome of a novel marine Thaumarchaeota strain suggest viral infection, phosphorothioation DNA modification and multiple restriction systems: genome and epigenome of a new marine thaumarchaeon. Environ Microbiol 19: 2434–2452. [DOI] [PubMed] [Google Scholar]
- Alonso‐Saez, L. , Waller, A.S. , Mende, D.R. , Bakker, K. , Farnelid, H. , Yager, P.L. , et al (2012) Role for urea in nitrification by polar marine Archaea. Proc Natl Acad Sci U S A 109: 17989–17994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amano, F. , and Noda, T. (1995) Improved detection of nitric oxide radical (NO.) production in an activated macrophage culture with a radical scavenger, carboxy PTIO and Griess reagent. FEBS Lett 368: 425–428. [DOI] [PubMed] [Google Scholar]
- Andersson, M. , Brion, N. , and Middelburg, J. (2006) Comparison of nitrifier activity versus growth in the Scheldt estuarya turbid, tidal estuary in northern Europe. Aquat Microb Ecol 42: 149–158. [Google Scholar]
- Arreguín, R. , Arreguín, B. , Soriano‐García, M. , Hernández‐Arana, A. , and Rodríguez‐Romero, A. (1993) Isolation and characterization of a protease from the marine sponge Spheciospongia vesparia . FEBS Lett 320: 235–238. [DOI] [PubMed] [Google Scholar]
- Astudillo‐García, C. , Bell, J.J. , Webster, N.S. , Glasl, B. , Jompa, J. , Montoya, J.M. , and Taylor, M.W. (2017) Evaluating the core microbiota in complex communities: a systematic investigation. Environ Microbiol 19: 1450–1462. [DOI] [PubMed] [Google Scholar]
- Bankevich, A. , Nurk, S. , Antipov, D. , Gurevich, A.A. , Dvorkin, M. , Kulikov, A.S. , et al (2012) SPAdes: a new genome assembly algorithm and its applications to single‐cell sequencing. J Comput Biol 19: 455–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer, K. , Schmitt, S. , and Hentschel, U. (2007) Microbial nitrification in Mediterranean sponges: possible involvement of ammonium‐oxidizing Betaproteobacteria In Porifera Research: Biodiversity, Innovation, Sustainability, Custódio M., Lôbo‐Hajdu G., Hajdu E., and Muricy G. (eds). Brazil: Museu Nacional: Série Livros. Rio de Janeiro, pp. 165–171. [Google Scholar]
- Bayer, K. , Schmitt, S. , and Hentschel, U. (2008) Physiology, phylogeny and in situ evidence for bacterial and archaeal nitrifiers in the marine sponge Aplysina aerophoba . Environ Microbiol 10: 2942–2955. [DOI] [PubMed] [Google Scholar]
- Bayer, B. , Vojvoda, J. , Offre, P. , Alves, R.J.E. , Elisabeth, N.H. , Garcia, J.A. , et al (2016) Physiological and genomic characterization of two novel marine thaumarchaeal strains indicates niche differentiation. ISME J 10: 1051–1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beam, J.P. , Jay, Z.J. , Kozubal, M.A. , and Inskeep, W.P. (2014) Niche specialization of novel Thaumarchaeota to oxic and hypoxic acidic geothermal springs of Yellowstone National Park. ISME J 8: 938–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beckers, G. , Bendt, A.K. , Kramer, R. , and Burkovski, A. (2004) Molecular identification of the urea uptake system and transcriptional analysis of urea transporter‐ and urease‐encoding genes in Corynebacterium glutamicum . J Bacteriol 186: 7645–7652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell, J.J. (2008) The functional roles of marine sponges. Estuar Coast Shelf Sci 79: 341–353. [Google Scholar]
- Bergquist, P.R. , and Kelly‐Borges, M. (1995) Systematics and biogeography of the genus Ianthella (Demospongiae: Verongida: Ianthellidae) in the south‐west Pacific. The Beagle, Records of the Museums and Art Galleries of the Northern Territory 12: 151–176. [Google Scholar]
- Boardman, G.D. , Starbuck, S.M. , Hudgins, D.B. , Li, X. , and Kuhn, D.D. (2004) Toxicity of ammonia to three marine fish and three marine invertebrates. Environ Toxicol 19: 134–142. [DOI] [PubMed] [Google Scholar]
- de Boer, W. , and Laanbroek, H.J. (1989) Ureolytic nitrification at low pH by Nitrosospira spec. Arch Microbiol 152: 178–181. [Google Scholar]
- Brady, A. , and Salzberg, S.L. (2009) Phymm and PhymmBL: metagenomic phylogenetic classification with interpolated Markov models. Nat Methods 6: 673–676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brezonik, P.L. , and Lee, G.F. (1968) Dentrification as a nitrogen sink in Lake Mendota, Wisconsin. Environ Sci Technol 2: 120–125. [Google Scholar]
- Brunori, M. , Giuffre, A. , Nienhaus, K. , Nienhaus, G.U. , Scandurra, F.M. , and Vallone, B. (2005) Neuroglobin, nitric oxide, and oxygen: functional pathways and conformational changes. Proc Natl Acad Sci U S A 102: 8483–8488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton, S.A.Q. , and Prosser, J.I. (2001) Autotrophic ammonia oxidation at low pH through urea hydrolysis. Appl Environ Microbiol 67: 2952–2957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callebaut, I. , Mornon, J.‐P. , Gilgès, D. , and Vigon, I. (2000) HYR, an extracellular module involved in cellular adhesion and related to the immunoglobulin‐like fold. Protein Sci 9: 1382–1390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camacho, C. , Coulouris, G. , Avagyan, V. , Ma, N. , Papadopoulos, J. , Bealer, K. , and Madden, T.L. (2009) BLAST+: architecture and applications. BMC Bioinformatics 10: 421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camargo, J.A. , and Alonso, Á. (2006) Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment. Environ Int 32: 831–849. [DOI] [PubMed] [Google Scholar]
- Carini, P. , Dupont, C.L. , and Santoro, A.E. (2018) Patterns of thaumarchaeal gene expression in culture and diverse marine environments. Environ Microbiol 20: 2112–2124. [DOI] [PubMed] [Google Scholar]
- Caranto, J.D. , and Lancaster, K.M. (2017) Nitric oxide is an obligate bacterial nitrification intermediate produced by hydroxylamine oxidoreductase. Proc Natl Acad Sci U S A 114: 8217–8222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevreux, B., Wetter, T., and Suhai, S. (1999) Genome sequence assembly using trace signals and additional sequence information. German Conference on Bioinformatics GCB'99 GCB. Comput Sci Biol Proc pp. 45–56.
- Conaco, C. , Neveu, P. , Zhou, H. , Arcila, M. , Degnan, S.M. , Degnan, B.M. , and Kosik, K.S. (2012) Transcriptome profiling of the demosponge Amphimedon queenslandica reveals genome‐wide events that accompany major life cycle transitions. BMC Genomics 13: 209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corredor, J.E. , Wilkinson, C.R. , Vicente, V.P. , Morell, J.M. , and Otero, E. (1988) Nitrate release by Caribbean reef sponges. Limnol Oceanogr 33: 114–120. [Google Scholar]
- Cort, J.R. , Yee, A. , Edwards, A.M. , Arrowsmith, C.H. , and Kennedy, M.A. (2000) Structure‐based functional classification of hypothetical protein MTH538 from Methanobacterium thermoautotrophicum . J Mol Biol 302: 189–203. [DOI] [PubMed] [Google Scholar]
- Daims, H. , Ramsing, N.B. , Schleifer, K.‐H. , and Wagner, M. (2001) Cultivation‐independent, semiautomatic determination of absolute bacterial cell numbers in environmental samples by fluorescence in situ hybridization. Appl Environ Microbiol 67: 5810–5818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daims, H. , Lebedeva, E.V. , Pjevac, P. , Han, P. , Herbold, C. , Albertsen, M. , et al (2015) Complete nitrification by Nitrospira bacteria. Nature 528: 504–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Díez‐Vives, C. , Moitinho‐Silva, L. , Nielsen, S. , Reynolds, D. , and Thomas, T. (2017) Expression of eukaryotic‐like protein in the microbiome of sponges. Mol Ecol 26: 1432–1451. [DOI] [PubMed] [Google Scholar]
- Drapeau, G.R. (1980) Substrate specificity of a proteolytic enzyme isolated from a mutant of Pseudomonas fragi . J Biol Chem 255: 839–840. [PubMed] [Google Scholar]
- Elias, J.E. , and Gygi, S.P. (2007) Target‐decoy search strategy for increased confidence in large‐scale protein identifications by mass spectrometry. Nat Methods 4: 207–214. [DOI] [PubMed] [Google Scholar]
- Ellis, A. , Lu, H. , Li, C.G. , and Rand, M.J. (2001) Effects of agents that inactivate free radical NO (NO center dot) on nitroxyl anion‐mediated relaxations, and on the detection of NO center dot released from the nitroxyl anion donor Angeli's salt. Brit J Pharmacol 134: 521–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emms, D.M. , and Kelly, S. (2015) OrthoFinder: solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol 16: 157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Essuman, K. , Summers, D.W. , Sasaki, Y. , Mao, X. , Yim, A.K.Y. , DiAntonio, A. , and Milbrandt, J. (2018) TIR domain proteins are an ancient family of NAD(+)‐consuming enzymes. Curr Biol 28: 421–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan, L. , Reynolds, D. , Liu, M. , Stark, M. , Kjelleberg, S. , Webster, N.S. , and Thomas, T. (2012) Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc Natl Acad Sci U S A 109: E1878–E1887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finn, R.D. , Bateman, A. , Clements, J. , Coggill, P. , Eberhardt, R.Y. , Eddy, S.R. , et al (2014) Pfam: the protein families database. Nucleic Acids Res 42: D222–D230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiore, C.L. , Labrie, M. , Jarett, J.K. , and Lesser, M.P. (2015) Transcriptional activity of the giant barrel sponge, Xestospongia muta Holobiont: molecular evidence for metabolic interchange. Front Microbiol 6: 364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Florens, L. , Carozza, M. , Swanson, S. , Fournier, M. , Coleman, M. , Workman, J. , and Washburn, M. (2006) Analyzing chromatin remodeling complexes using shotgun proteomics and normalized spectral abundance factors. Methods 40: 303–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freckelton, M.L. , Luter, H.M. , Andreakis, N. , Webster, N.S. , and Motti, C.A. (2012) Qualitative variation in colour morphotypes of Ianthella basta (Porifera: Verongida). Hydrobiologia 687: 191–203. [Google Scholar]
- Freeman, M.F. , Vagstad, A.L. , and Piel, J. (2016) Polytheonamide biosynthesis showcasing the metabolic potential of sponge‐associated uncultivated ‘Entotheonella’ bacteria. Curr Opin Chem Biol 31: 8–14. [DOI] [PubMed] [Google Scholar]
- Gardner, P.R. (2012) Hemoglobin: a nitric‐oxide dioxygenase. Scientifica 2012, 1–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gauthier, M.‐E.A. , Watson, J.R. , and Degnan, S.M. (2016) Draft genomes shed light on the dual bacterial symbiosis that dominates the microbiome of the coral reef sponge Amphimedon queenslandica . Front Mar Sci 3: 196.1–196.18. [Google Scholar]
- de Goeij, J.M. , van Oevelen, D. , Vermeij, M.J.A. , Osinga, R. , Middelburg, J.J. , de Goeij, A.F.P.M. , and Admiraal, W. (2013) Surviving in a marine desert: the sponge loop retains resources within coral reefs. Science 342: 108–110. [DOI] [PubMed] [Google Scholar]
- Guy, L. , and Ettema, T.J.G. (2011) The archaeal ‘TACK’ superphylum and the origin of eukaryotes. Trends Microbiol 19: 580–587. [DOI] [PubMed] [Google Scholar]
- Hallam, S.J. , Konstantinidis, K.T. , Putnam, N. , Schleper, C. , Watanabe, Y. , Sugahara, J. , et al (2006) Genomic analysis of the uncultivated marine crenarchaeote Cenarchaeum symbiosum . Proc Natl Acad Sci U S A 103: 18296–18301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatzenpichler, R. , Lebedeva, E.V. , Spieck, E. , Stoecker, K. , Richter, A. , Daims, H. , and Wagner, M. (2008) A moderately thermophilic ammonia‐oxidizing crenarchaeote from a hot spring. Proc Natl Acad Sci U S A 105: 2134–2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinz, E. , Williams, T.A. , Nakjang, S. , Noël, C.J. , Swan, D.C. , Goldberg, A.V. , et al (2012) The genome of the obligate intracellular parasite Trachipleistophora hominis: new insights into microsporidian genome dynamics and reductive evolution. PLoS Pathog 8: e1002979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hentschel, U. , Piel, J. , Degnan, S.M. , and Taylor, M.W. (2012) Genomic insights into the marine sponge microbiome. Nat Rev Microbiol 10: 641–654. [DOI] [PubMed] [Google Scholar]
- Herndl, G.J. , Reinthaler, T. , Teira, E. , van Aken, H. , Veth, C. , Pernthaler, A. , and Pernthaler, J. (2005) Contribution of Archaea to total prokaryotic production in the deep Atlantic Ocean. Appl Environ Microbiol 71: 2303–2309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann, F. , Larsen, O. , Thiel, V. , Rapp, H.T. , Pape, T. , Michaelis, W. , and Reitner, J. (2005) An anaerobic world in sponges. Geomicrobiol J 22: 1–10. [Google Scholar]
- Hoffmann, F. , Radax, R. , Woebken, D. , Holtappels, M. , Lavik, G. , Rapp, H.T. , et al (2009) Complex nitrogen cycling in the sponge Geodia barretti . Environ Microbiol 11: 2228–2243. [DOI] [PubMed] [Google Scholar]
- Horn, H. , Slaby, B.M. , Jahn, M.T. , Bayer, K. , Moitinho‐Silva, L. , Förster, F. , et al (2016) An enrichment of CRISPR and other defense‐related features in marine sponge‐associated microbial metagenomes. Front Microbiol 7: 1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoshino, T. (1979) Transport systems for branched‐chain amino acids in Pseudomonas aeruginosa . J Bacteriol 139: 705–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosie, A.H.F. , Allaway, D. , Galloway, C.S. , Dunsby, H.A. , and Poole, P.S. (2002) Rhizobium leguminosarum has a second general amino acid permease with unusually broad substrate specificity and high similarity to branched‐chain amino acid transporters (Bra/LIV) of the ABC family. J Bacteriol 184: 4071–4080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseinzadeh, P. , Tian, S. , Marshall, N.M. , Hemp, J. , Mullen, T. , Nilges, M.J. , et al (2016) A purple cupredoxin from Nitrosopumilus maritimus containing a mononuclear type 1 copper center with an open binding site. J Am Chem Soc 138: 6324–6327. [DOI] [PubMed] [Google Scholar]
- Hyatt, D. , Chen, G.‐L. , LoCascio, P.F. , Land, M.L. , Larimer, F.W. , and Hauser, L.J. (2010) Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 11: 119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ingalls, A.E. , Shah, S.R. , Hansman, R.L. , Aluwihare, L.I. , Santos, G.M. , Druffel, E.R.M. , and Pearson, A. (2006) Quantifying archaeal community autotrophy in the mesopelagic ocean using natural radiocarbon. Proc Natl Acad Sci U S A 103: 6442–6447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inselsbacher, E. , Cambui, C.A. , Richter, A. , Stange, C.F. , Mercier, H. , and Wanek, W. (2007) Microbial activities and foliar uptake of nitrogen in the epiphytic bromeliad Vriesea gigantea . New Phytol 175: 311–320. [DOI] [PubMed] [Google Scholar]
- Jompa, J. , and McCook, L.J. (2002) The effects of nutrients and herbivory on competition between a hard coral (Porites cylindrica) and a brown alga (Lobophora variegata). Limnol Oceanogr 47: 527–534. [Google Scholar]
- Kalyaanamoorthy, S. , Minh, B.Q. , Wong, T.K.F. , von Haeseler, A. , and Jermiin, L.S. (2017) ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods 14: 587–589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang, D.D. , Froula, J. , Egan, R. , and Wang, Z. (2015) MetaBAT, an efficient tool for accurately reconstructing single genomes from complex microbial communities. PeerJ 3: e1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerou, M. , Offre, P. , Valledor, L. , Abby, S.S. , Melcher, M. , Nagler, M. , et al (2016) Proteomics and comparative genomics of Nitrososphaera viennensis reveal the core genome and adaptations of archaeal ammonia oxidizers. Proc Natl Acad Sci U S A 113: E7937–E7946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, J.‐G. , Park, S.‐J. , Sinninghe Damsté, J.S. , Schouten, S. , Rijpstra, W.I.C. , Jung, M.‐Y. , et al (2016) Hydrogen peroxide detoxification is a key mechanism for growth of ammonia‐oxidizing archaea. Proc Natl Acad Sci U S A 113: 7888–7893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kits, K.D. , Jung, M.Y. , Vierheilig, J. , Pjevac, P. , Sedlacek, C.J. , Liu, S. , et al (2019) Low yield and abiotic origin of N2O formed by the complete nitrifier Nitrospira inopinata . Nat Commun 10: 1836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitzinger, K. , Padilla, C.C. , Marchant, H.K. , Hach, P.F. , Herbold, C.W. , Kidane, A.T. , et al (2019) Cyanate and urea are substrates for nitrification by Thaumarchaeota in the marine environment. Nat Microbiol 4: 234–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch, H. , Galushko, A. , Albertsen, M. , Schintlmeister, A. , Gruber‐Dorninger, C. , Lucker, S. , et al (2014) Growth of nitrite‐oxidizing bacteria by aerobic hydrogen oxidation. Science 345: 1052–1054. [DOI] [PubMed] [Google Scholar]
- Könneke, M. , Bernhard, A.E. , de la Torre, J.R. , Walker, C.B. , Waterbury, J.B. , and Stahl, D.A. (2005) Isolation of an autotrophic ammonia‐oxidizing marine archaeon. Nature 437: 543–546. [DOI] [PubMed] [Google Scholar]
- Konstantinidis, K.T. , and Tiedje, J.M. (2005) Towards a genome‐based taxonomy for prokaryotes. J Bacteriol 187: 6258–6264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konstantinidis, K.T. , Rosselló‐Móra, R. , and Amann, R. (2017) Uncultivated microbes in need of their own taxonomy. ISME J 11: 2399–2406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozlowski, J.A. , Stieglmeier, M. , Schleper, C. , Klotz, M.G. , and Stein, L.Y. (2016) Pathways and key intermediates required for obligate aerobic ammonia‐dependent chemolithotrophy in bacteria and Thaumarchaeota. ISME J 10: 1836–1845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam, P. , Jensen, M.M. , Kock, A. , Lettmann, K.A. , Plancherel, Y. , Lavik, G. , et al (2011) Origin and fate of the secondary nitrite maximum in the Arabian Sea. Biogeosciences 8: 1565–1577. [Google Scholar]
- Lartillot, N. , and Philippe, H. (2004) A Bayesian mixture model for across‐site heterogeneities in the amino‐acid replacement process. Mol Biol Evol 21: 1095–1109. [DOI] [PubMed] [Google Scholar]
- Lechauve, C. , Jager, M. , Laguerre, L. , Kiger, L. , Correc, G. , Leroux, C. , et al (2013) Neuroglobins, pivotal proteins associated with emerging neural systems and precursors of metazoan globin diversity. J Biol Chem 288: 6957–6967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehtovirta‐Morley, L.E. , Stoecker, K. , Vilcinskas, A. , Prosser, J.I. , and Nicol, G.W. (2011) Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil. Proc Natl Acad Sci U S A 108: 15892–15897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, B. , Lopes, J.S. , Foster, P.G. , Embley, T.M. , and Cox, C.J. (2014) Compositional biases among synonymous substitutions cause conflict between gene and protein trees for plastid origins. Mol Biol Evol 31: 1697–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, M. , Baker, B.J. , Anantharaman, K. , Jain, S. , Breier, J.A. , and Dick, G.J. (2015) Genomic and transcriptomic evidence for scavenging of diverse organic compounds by widespread deep‐sea archaea. Nat Commun 6: 8933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , Ding, K. , Wen, X. , Zhang, B. , Shen, B. , and Yang, Y. (2016) A novel ammonia‐oxidizing archaeon from wastewater treatment plant: its enrichment, physiological and genomic characteristics. Sci Rep‐UK 6: 23747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, P.‐N. , Herrmann, J. , Tolar, B.B. , Poitevin, F. , Ramdasi, R. , Bargar, J.R. , et al (2018) Nutrient transport suggests an evolutionary basis for charged archaeal surface layer proteins. ISME J 12: 2389–2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, X. , Handley, K.M. , Gilbert, J.A. , and Kostka, J.E. (2015) Metabolic potential of fatty acid oxidation and anaerobic respiration by abundant members of Thaumarchaeota and Thermoplasmata in deep anoxic peat. ISME J 9: 2740–2744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, M. , Fan, L. , Zhong, L. , Kjelleberg, S. , and Thomas, T. (2012) Metaproteogenomic analysis of a community of sponge symbionts. ISME J 6: 1515–1525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd, K.G. , Schreiber, L. , Petersen, D.G. , Kjeldsen, K.U. , Lever, M.A. , Steen, A.D. , et al (2013) Predominant archaea in marine sediments degrade detrital proteins. Nature 496: 215–218. [DOI] [PubMed] [Google Scholar]
- Love, G.D. , Grosjean, E. , Stalvies, C. , Fike, D.A. , Grotzinger, J.P. , Bradley, A.S. , et al (2009) Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 457: 718–721. [DOI] [PubMed] [Google Scholar]
- Luo, C. , Rodriguez‐R, L.M. , and Konstantinidis, K.T. (2014) MyTaxa: an advanced taxonomic classifier for genomic and metagenomic sequences. Nucleic Acids Res 42: e73#x2013;e73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lurie‐Weinberger, M.N. , Gomez‐Valero, L. , Merault, N. , Glöckner, G. , Buchrieser, C. , and Gophna, U. (2010) The origins of eukaryotic‐like proteins in Legionella pneumophila . Int J Med Microbiol 300: 470–481. [DOI] [PubMed] [Google Scholar]
- Luter, H.M. , Whalan, S. , and Webster, N.S. (2010) Exploring the role of microorganisms in the disease‐like syndrome affecting the sponge Ianthella basta . Appl Environ Microbiol 76: 5736–5744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luter, H.M. , Whalan, S. , and Webster, N.S. (2012) Thermal and sedimentation stress are unlikely causes of brown spot syndrome in the coral reef sponge, Ianthella basta . PLoS ONE 7: e39779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maldonado, M. , Ribes, M. , and van Duyl, F.C. (2012) Nutrient fluxes through sponges: biology, budgets, and ecological implications. Adv Mar Biol 62: 113–182. [DOI] [PubMed] [Google Scholar]
- Maixner, F. , Noguera, D.R. , Anneser, B. , Stoecker, K. , Wegl, G. , Wagner, M. , and Daims, H. (2006) Nitrite concentration influences the population structure of Nitrospira‐like bacteria. Environ Microbiol 8: 1487–1495. [DOI] [PubMed] [Google Scholar]
- Martens‐Habbena, W. , Berube, P.M. , Urakawa, H. , de la Torre, J.R. , and Stahl, D.A. (2009) Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 461: 976–979. [DOI] [PubMed] [Google Scholar]
- Martens‐Habbena, W. , Qin, W. , Horak, R.E.A. , Urakawa, H. , Schauer, A.J. , Moffett, J.W. , et al (2015) The production of nitric oxide by marine ammonia‐oxidizing archaea and inhibition of archaeal ammonia oxidation by a nitric oxide scavenger: marine AOA produce NO and are inhibited by NO scavenger. Environ Microbiol 17: 2261–2274. [DOI] [PubMed] [Google Scholar]
- Mehrshad, M. , Amoozegar, M.A. , Ghai, R. , Shahzadeh Fazeli, S.A. , and Rodriguez‐Valera, F. (2016) Genome reconstruction from metagenomic data sets reveals novel microbes in the brackish waters of the Caspian Sea. Appl Environ Microbiol 82: 1599–1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, W.T. (2012) Tyrosine kinase signaling and the emergence of multicellularity. BBA‐Mol Cell Res 1823: 1053–1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohamed, N.M. , Saito, K. , Tal, Y. , and Hill, R.T. (2010) Diversity of aerobic and anaerobic ammonia‐oxidizing bacteria in marine sponges. ISME J 4: 38–48. [DOI] [PubMed] [Google Scholar]
- Moitinho‐Silva, L. , Díez‐Vives, C. , Batani, G. , Esteves, A.I. , Jahn, M.T. , and Thomas, T. (2017a) Integrated metabolism in sponge–microbe symbiosis revealed by genome‐centered metatranscriptomics. ISME J 11: 1651–1666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moitinho‐Silva, L. , Nielsen, S. , Amir, A. , Gonzalez, A. , Ackermann, G.L. , Cerrano, C. , et al (2017b) The sponge microbiome project. GigaScience 6: 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morley, S.A. , Berman, J. , Barnes, D.K.A. , de Juan Carbonell, C. , Downey, R.V. , and Peck, L.S. (2016) Extreme phenotypic plasticity in metabolic physiology of Antarctic demosponges. Front Ecol Evol 3: 157. [Google Scholar]
- Müller, W.E.G. , Ushijima, H. , Batel, R. , Krasko, A. , Borejko, A. , Müller, I.M. , and Schröder, H.‐C. (2006) Novel mechanism for the radiation‐induced bystander effect: nitric oxide and ethylene determine the response in sponge cells. Mutat Res 597: 62–72. [DOI] [PubMed] [Google Scholar]
- Murphy, D. , Recous, S. , Stockdale, E. , Fillery, I.R. , Jensen, L. , Hatch, D. , and Goulding, K.W. (2003) Gross nitrogen fluxes in soil: theory, measurement and application of 15N pool dilution techniques. Adv Agron 79: 69–118. [Google Scholar]
- Mußmann, M. , Brito, I. , Pitcher, A. , Sinninghe Damste, J.S. , Hatzenpichler, R. , Richter, A. , et al (2011) Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but are not obligate autotrophic ammonia oxidizers. Proc Natl Acad Sci U S A 108: 16771–16776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa, T. , and Stahl, D.A. (2013) Transcriptional response of the archaeal ammonia oxidizer Nitrosopumilus maritimus to low and environmentally relevant ammonia concentrations. Appl Environ Microbiol 79: 6911–6916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newstead, S. (2015) Molecular insights into proton coupled peptide transport in the PTR family of oligopeptide transporters. BBA ‐ Gen Subjects 1850: 488–499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Off, S. , Alawi, M. , and Spieck, E. (2010) Enrichment and physiological characterization of a novel Nitrospira‐like bacterium obtained from a marine sponge. Appl Environ Microbiol 76: 4640–4646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Offre, P. , Kerou, M. , Spang, A. , and Schleper, C. (2014) Variability of the transporter gene complement in ammonia‐oxidizing archaea. Trends Microbiol 22: 665–675. [DOI] [PubMed] [Google Scholar]
- Olm, M.R. , Brown, C.T. , Brooks, B. , and Banfield, J.F. (2017) dRep: a tool for fast and accurate genomic comparisons that enables improved genome recovery from metagenomes through de‐replication. ISME J 11: 2864–2868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto, A. , Bernhardt, J. , Meyer, H. , Schaffer, M. , Herbst, F.‐A. , Siebourg, J. , et al (2010) Systems‐wide temporal proteomic profiling in glucose‐starved Bacillus subtilis . Nat Commun 1: 137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ouverney, C.C. , and Fuhrman, J.A. (2000) Marine planktonic archaea take up amino acids. Appl Environ Microbiol 66: 4829–4833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palatinszky, M. , Herbold, C. , Jehmlich, N. , Pogoda, M. , Han, P. , von Bergen, M. , et al (2015) Cyanate as an energy source for nitrifiers. Nature 524: 105–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, S.‐J. , Ghai, R. , Martín‐Cuadrado, A.‐B. , Rodríguez‐Valera, F. , Chung, W.‐H. , Kwon, K. , et al (2014) Genomes of two new ammonia‐oxidizing archaea enriched from deep marine sediments. PLoS ONE 9: e96449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks, D.H. , Imelfort, M. , Skennerton, C.T. , Hugenholtz, P. , and Tyson, G.W. (2015) CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res 25: 1043–1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks, D.H. , Chuvochina, M. , Waite, D.W. , Rinke, C. , Skarshewski, A. , Chaumeil, P.‐A. , and Hugenholtz, P. (2018) A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat Biotechnol 36: 996–1004. [DOI] [PubMed] [Google Scholar]
- Passmore, I.J. , Nishikawa, K. , Lilley, K.S. , Bowden, S.D. , Chung, J.C.S. , and Welch, M. (2015) Mep72, a metzincin protease that is preferentially secreted by biofilms of Pseudomonas aeruginosa . J Bacteriol 197: 762–773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson, N.J. , Günther, J. , Gibson, A.J. , Offord, V. , Coffey, T.J. , Splitter, G. , et al (2014) Two TIR‐like domain containing proteins in a newly emerging zoonotic Staphylococcus aureus strain sequence type 398 are potential virulence factors by impacting on the host innate immune response. Front Microbiol 5: 662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng, J. , Elias, J.E. , Thoreen, C.C. , Licklider, L.J. , and Gygi, S.P. (2003) Evaluation of multidimensional chromatography coupled with tandem mass spectrometry (LC/LC‐MS/MS) for large‐scale protein analysis: the yeast proteome. J Proteome Res 2: 43–50. [DOI] [PubMed] [Google Scholar]
- Pester, M. , Schleper, C. , and Wagner, M. (2011) The Thaumarchaeota: an emerging view of their phylogeny and ecophysiology. Curr Opin Microbiol 14: 300–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips, G. , Swairjo, M.A. , Gaston, K.W. , Bailly, M. , Limbach, P.A. , Iwata‐Reuyl, D. , and de Crécy‐Lagard, V. (2012) Diversity of archaeosine synthesis in crenarchaeota. ACS Chem Biol 7: 300–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picossi, S. , Montesinos, M.L. , Pernil, R. , Lichtlé, C. , Herrero, A. , and Flores, E. (2005) ABC‐type neutral amino acid permease N‐I is required for optimal diazotrophic growth and is repressed in the heterocysts of Anabaena sp. strain PCC 7120. Mol Microbiol 57: 1582–1592. [DOI] [PubMed] [Google Scholar]
- Pruesse, E. , Peplies, J. , and Glöckner, F.O. (2012) SINA: accurate high‐throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28: 1823–1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin, W. , Martens‐Habbena, W. , Kobelt, J.N. , and Stahl, D.A. (2016) Candidatus Nitrosopumilaceae In Bergey's Manual of Systematics of Archaea and Bacteria, Whitman W.B., Rainey F., Kämpfer P., Trujillo M., Chun J., DeVos P., et al (eds). Chichester, UK: John Wiley & Sons, pp. 1–2. [Google Scholar]
- Qin, W. , Meinhardt, K.A. , Moffett, J.W. , Devol, A.H. , Virginia Armbrust, E. , Ingalls, A.E. , and Stahl, D.A. (2017) Influence of oxygen availability on the activities of ammonia‐oxidizing archaea: influence of oxygen availability. Envirol Microbiol Rep 9: 250–256. [DOI] [PubMed] [Google Scholar]
- Qin, W. , Amin, S.A. , Martens‐Habbena, W. , Walker, C.B. , Urakawa, H. , Devol, A.H. , et al (2014) Marine ammonia‐oxidizing archaeal isolates display obligate mixotrophy and wide ecotypic variation. Proc Natl Acad Sci U S A 111: 12504–12509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radax, R. , Hoffmann, F. , Rapp, H.T. , Leininger, S. , and Schleper, C. (2012a) Ammonia‐oxidizing archaea as main drivers of nitrification in cold‐water sponges: Archaeal nitrification in cold‐water sponges. Environ Microbiol 14: 909–923. [DOI] [PubMed] [Google Scholar]
- Radax, R. , Rattei, T. , Lanzen, A. , Bayer, C. , Rapp, H.T. , Urich, T. , and Schleper, C. (2012b) Metatranscriptomics of the marine sponge Geodia barretti: tackling phylogeny and function of its microbial community. Environ Microbiol 14: 1308–1324. [DOI] [PubMed] [Google Scholar]
- Reveillaud, J. , Maignien, L. , Eren, A.M. , Huber, J.A. , Apprill, A. , Sogin, M.L. , and Vanreusel, A. (2014) Host‐specificity among abundant and rare taxa in the sponge microbiome. ISME J 8: 1198–1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds, D. , and Thomas, T. (2016) Evolution and function of eukaryotic‐like proteins from sponge symbionts. Mol Ecol 25: 5242–5253. [DOI] [PubMed] [Google Scholar]
- Riesgo, A. , Farrar, N. , Windsor, P.J. , Giribet, G. , and Leys, S.P. (2014) The analysis of eight transcriptomes from all poriferan classes reveals surprising genetic complexity in sponges. Mol Biol Evol 31: 1102–1120. [DOI] [PubMed] [Google Scholar]
- Sabina, J. , and Söll, D. (2006) The RNA‐binding PUA domain of archaeal tRNA‐guanine transglycosylase is not required for archaeosine formation. J Biol Chem 281: 6993–7001. [DOI] [PubMed] [Google Scholar]
- Santoro, A.E. , Dupont, C.L. , Richter, R.A. , Craig, M.T. , Carini, P. , McIlvin, M.R. , et al (2015) Genomic and proteomic characterization of “ Candidatus Nitrosopelagicus brevis”: an ammonia‐oxidizing archaeon from the open ocean. Proc Natl Acad Sci U S A 112: 1173–1178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauder, L.A. , Engel, K. , Lo, C.‐C. , Chain, P. , and Neufeld, J.D. (2018) “Candidatus Nitrosotenuis aquarius,” an ammonia‐oxidizing archaeon from a freshwater aquarium biofilter. Appl Environ Microbiol 84: e01430#x2013;e01418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauder, L.A. , Albertsen, M. , Engel, K. , Schwarz, J. , Nielsen, P.H. , Wagner, M. , and Neufeld, J.D. (2017) Cultivation and characterization of Candidatus Nitrosocosmicus exaquare, an ammonia‐oxidizing archaeon from a municipal wastewater treatment system. ISME J 11: 1142–1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaefer, S.C. , and Hollibaugh, J.T. (2017) Temperature decouples ammonium and nitrite oxidation in coastal waters. Environ Sci Technol 51: 3157–3164. [DOI] [PubMed] [Google Scholar]
- Schläppy, M.‐L. , Schöttner, S.I. , Lavik, G. , Kuypers, M.M.M. , de Beer, D. , and Hoffmann, F. (2010) Evidence of nitrification and denitrification in high and low microbial abundance sponges. Mar Biol 157: 593–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schleper, C. , DeLong, E.F. , Preston, C.M. , Feldman, R.A. , Wu, K.Y. , and Swanson, R.V. (1998) Genomic analysis reveals chromosomal variation in natural populations of the uncultured psychrophilic archaeon Cenarchaeum symbiosum . J Bacteriol 180: 5003–5009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt, S. , Tsai, P. , Bell, J. , Fromont, J. , Ilan, M. , Lindquist, N. , et al (2012) Assessing the complex sponge microbiota: core, variable and species‐specific bacterial communities in marine sponges. ISME J 6: 564–576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp, K.H. , Eam, B. , Faulkner, D.J. , and Haygood, M.G. (2007) Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl Environ Microbiol 73: 622–629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen, T. , Stieglmeier, M. , Dai, J. , Urich, T. , and Schleper, C. (2013) Responses of the terrestrial ammonia‐oxidizing archaeon Ca. Nitrososphaera viennensis and the ammonia‐oxidizing bacterium Nitrosospira multiformis to nitrification inhibitors. FEMS Microbiol Let 344: 121–129. [DOI] [PubMed] [Google Scholar]
- Shigeno‐Nakazawa, Y. , Kasai, T. , Ki, S. , Kostyanovskaya, E. , Pawlak, J. , Yamagishi, J. , et al (2016) A pre‐metazoan origin of the CRK gene family and co‐opted signaling network. Sci Rep‐UK 6: 3439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simion, P. , Philippe, H. , Baurain, D. , Jager, M. , Richter, D.J. , Di Franco, A. , et al (2017) A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals. Curr Biol 27: 958–967. [DOI] [PubMed] [Google Scholar]
- Simister, R.L. , Deines, P. , Botté, E.S. , Webster, N.S. , and Taylor, M.W. (2012) Sponge‐specific clusters revisited: a comprehensive phylogeny of sponge‐associated microorganisms. Environ Microbiol 14: 517–524. [DOI] [PubMed] [Google Scholar]
- Sørensen, P. , and Jensen, E.S. (1991) Sequential diffusion of ammonium and nitrate from soil extracts to a polytetrafluoroethylene trap for 15N determination. Anal Chim Acta 252: 201–203. [Google Scholar]
- Southwell, M.W. , Popp, B.N. , and Martens, C.S. (2008) Nitrification controls on fluxes and isotopic composition of nitrate from Florida keys sponges. Mar Chem 108: 96–108. [Google Scholar]
- Spang, A. , Poehlein, A. , Offre, P. , Zumbrägel, S. , Haider, S. , Rychlik, N. , et al (2012) The genome of the ammonia‐oxidizing Candidatus Nitrososphaera gargensis: insights into metabolic versatility and environmental adaptations. Environ Microbiol 14: 3122–3145. [DOI] [PubMed] [Google Scholar]
- Steger, D. , Ettinger‐Epstein, P. , Whalan, S. , Hentschel, U. , de Nys, R. , Wagner, M. , and Taylor, M.W. (2008) Diversity and mode of transmission of ammonia‐oxidizing archaea in marine sponges. Environ Microbiol 10: 1087–1094. [DOI] [PubMed] [Google Scholar]
- Stoecker, K. , Dorninger, C. , Daims, H. , and Wagner, M. (2010) Double labeling of oligonucleotide probes for fluorescence in situ hybridization (DOPE‐FISH) improves signal intensity and increases rRNA accessibility. Appl Environ Microbiol 76: 922–926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabares, P. , Pimentel‐Elardo, S.M. , Schirmeister, T. , Hünig, T. , and Hentschel, U. (2011) Anti‐protease and immunomodulatory activities of bacteria associated with Caribbean sponges. Marine Biotechnol 13: 883–892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor, M.W. , Radax, R. , Steger, D. , and Wagner, M. (2007) Sponge‐associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol Mol Biol Rev 1: 295–347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teira, E. , van Aken, H. , Veth, C. , and Herndl, G.J. (2006) Archaeal uptake of enantiomeric amino acids in the meso‐ and bathypelagic waters of the North Atlantic. Limnol Oceanogr 51: 60–69. [Google Scholar]
- Thiaville, J.J. , Kellner, S.M. , Yuan, Y. , Hutinet, G. , Thiaville, P.C. , Jumpathong, W. , et al (2016) Novel genomic Island modifies DNA with 7‐deazaguanine derivatives. Proc Natl Acad Sci U S A 113: E1452–E1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, T. , Moitinho‐Silva, L. , Lurgi, M. , Björk, J.R. , Easson, C. , Astudillo‐García, C. , et al (2016) Diversity, structure and convergent evolution of the global sponge microbiome. Nat Commun 7: 11870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian, R.‐M. , Sun, J. , Cai, L. , Zhang, W.‐P. , Zhou, G.‐W. , Qiu, J.‐W. , and Qian, P.‐Y. (2016) The deep‐sea glass sponge Lophophysema eversa harbours potential symbionts responsible for the nutrient conversions of carbon, nitrogen and sulfur. Environ Microbiol 18: 2481–2494. [DOI] [PubMed] [Google Scholar]
- Tong, T. , Chen, S. , Wang, L. , Tang, Y. , Ryu, J.Y. , Jiang, S. , et al (2018) Occurrence, evolution, and functions of DNA phosphorothioate epigenetics in bacteria. Proc Natl Acad Sci U S A 115: E2988–E2996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tourna, M. , Stieglmeier, M. , Spang, A. , Könneke, M. , Schintlmeister, A. , Urich, T. , et al (2011) Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc Natl Acad Sci U S A 108: 8420–8425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trame, C.B. , Chang, Y. , Axelrod, H.L. , Eberhardt, R.Y. , Coggill, P. , Punta, M. , and Rawlings, N.D. (2014) New mini‐ zincin structures provide a minimal scaffold for members of this metallopeptidase superfamily. BMC Bioinformatics 15: 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ueda, N. , Richards, G.S. , Degnan, B.M. , Kranz, A. , Adamska, M. , Croll, R.P. , and Degnan, S.M. (2016) An ancient role for nitric oxide in regulating the animal pelagobenthic life cycle: evidence from a marine sponge. Sci Rep‐UK 6: 37546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vacelet, J. , Boury‐Esnault, N. , Fiala‐Medioni, A. , and Fisher, C.R. (1995) A methanotrophic carnivorous sponge. Nature 377: 296–296. [Google Scholar]
- Valladares, A. , Montesinos, M.L. , Herrero, A. , and Flores, E. (2002) An ABC‐type, high‐affinity urea permease identified in cyanobacteria. Mol Microbiol 43: 703–715. [DOI] [PubMed] [Google Scholar]
- Vallenet, D. , Engelen, S. , Mornico, D. , Cruveiller, S. , Fleury, L. , Lajus, A. , et al (2009) MicroScope: a platform for microbial genome annotation and comparative genomics. Database 2009: bap021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varghese, N.J. , Mukherjee, S. , Ivanova, N. , Konstantinidis, K.T. , Mavrommatis, K. , Kyrpides, N.C. , and Pati, A. (2015) Microbial species delineation using whole genome sequences. Nucleic Acids Res 43: 6761–6771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventura, M. , Turroni, F. , Motherway, M.O. , MacSharry, J. , and van Sinderen, D. (2012) Host–microbe interactions that facilitate gut colonization by commensal bifidobacteria. Trends Microbiol 20: 467–476. [DOI] [PubMed] [Google Scholar]
- Verberkmoes, N.C. , Russell, A.L. , Shah, M. , Godzik, A. , Rosenquist, M. , Halfvarson, J. , et al (2009) Shotgun metaproteomics of the human distal gut microbiota. ISME J 3: 179–189. [DOI] [PubMed] [Google Scholar]
- Wallner, G. , Amann, R. , and Beisker, W. (1993) Optimizing fluorescent in situ hybridization with rRNA‐targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry 14: 136–143. [DOI] [PubMed] [Google Scholar]
- Wanek, W. , Mooshammer, M. , Blöchl, A. , Hanreich, A. , and Richter, A. (2010) Determination of gross rates of amino acid production and immobilization in decomposing leaf litter by a novel 15N isotope pool dilution technique. Soil Biol Biochem 42: 1293–1302. [Google Scholar]
- Wang, L. , Chen, S. , Xu, T. , Taghizadeh, K. , Wishnok, J.S. , Zhou, X. , et al (2007) Phosphorothioation of DNA in bacteria by dnd genes. Nat Chem Biol 3: 709–710. [DOI] [PubMed] [Google Scholar]
- Washburn, M.P. , Wolters, D. , and Yates, J.R. (2001) Large‐scale analysis of the yeast proteome by multidimensional protein identification technology. Nat Biotechnol 19: 242–247. [DOI] [PubMed] [Google Scholar]
- Weber, E.B. , Lehtovirta‐Morley, L.E. , Prosser, J.I. , and Gubry‐Rangin, C. (2015) Ammonia oxidation is not required for growth of group 1.1c soil Thaumarchaeota. FEMS Microbiol Ecol 91: fiv001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster, N.S. , and Thomas, T. (2016) The sponge Hologenome. MBio 7: e00135#x2013;e00116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster, N.S. , Taylor, M.W. , Behnam, F. , Lücker, S. , Rattei, T. , Whalan, S. , et al (2009) Deep sequencing reveals exceptional diversity and modes of transmission for bacterial sponge symbionts. Environ Microbiol 12: 2070–2082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkesman, J. , and Schröder, H.C. (2002) Heat‐stable protease from the marine sponge Geodia cydonium . Cell Mol Bio (Noisy‐le‐grand) 48: 379–383. [PubMed] [Google Scholar]
- Wilkesman, J.G. , and Schröder, H.C. (2007) Analysis of serine proteases from marine sponges by 2‐D zymography. Electrophoresis 28: 429–436. [DOI] [PubMed] [Google Scholar]
- Wilkinson, C.R. (1983) Net primary productivity in coral reef sponges. Science 219: 410–412. [DOI] [PubMed] [Google Scholar]
- Wilkinson, C.R. , and Fay, P. (1979) Nitrogen fixation in coral reef sponges with symbiotic cyanobacteria. Nature 279: 527–529. [Google Scholar]
- Wilson, M.C. , Mori, T. , Rückert, C. , Uria, A.R. , Helf, M.J. , Takada, K. , et al (2014) An environmental bacterial taxon with a large and distinct metabolic repertoire. Nature 506: 58–62. [DOI] [PubMed] [Google Scholar]
- Wuchter, C. , Abbas, B. , Coolen, M.J.L. , Herfort, L. , van Bleijswijk, J. , Timmers, P. , et al (2006) Archaeal nitrification in the ocean. Proc Natl Acad Sci U S A 103: 12317–12322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan, J. , Haaijer, S.C.M. , Op den Camp, H.J.M. , van Niftrik, L. , Stahl, D.A. , Könneke, M. , et al (2012) Mimicking the oxygen minimum zones: stimulating interaction of aerobic archaeal and anaerobic bacterial ammonia oxidizers in a laboratory‐scale model system. Environ Microbiol 14: 3146–3158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- You, D. , Wang, L. , Yao, F. , Zhou, X. , and Deng, Z. (2007) A novel DNA modification by sulfur: DndA is a NifS‐like cysteine desulfurase capable of assembling DndC as an iron‐sulfur cluster protein in Streptomyces lividans . Biochemistry 46: 6126–6133. [DOI] [PubMed] [Google Scholar]
- Zhalnina, K.V. , Dias, R. , Leonard, M.T. , Dorr de Quadros, P. , Camargo, F.A.O. , Drew, J.C. , et al (2014) Genome sequence of Candidatus Nitrososphaera evergladensis from group I.1b enriched from Everglades soil reveals novel genomic features of the ammonia‐oxidizing archaea. PLoS ONE 9: e101648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaikova, E. (2014) Molecular Mechanisms of Host‐Symbiont Recognition in a Highly Specific Sponge‐Archaeal Symbiosis. PhD Thesis, University of British Columbia; [Google Scholar]
- Zhou, J. , Bruns, M.A. , and Tiedje, J.M. (1996) DNA recovery from soils of diverse composition. Appl Environ Microbiol 62: 316–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix. Supporting Information.