

Summary
Transcription Activator‐Like effectors (TALes) represent the largest family of type III effectors among pathogenic bacteria and play a critical role in the process of infection. Strains of Xanthomonas oryzae pv. oryzae (Xoo) and some strains of other Xanthomonas pathogens contain large numbers of TALe genes. Previous techniques to clone individual or a complement of TALe genes through conventional strategies are inefficient and time‐consuming due to multiple genes (up to 29 copies) in a given genome, and technically challenging due to the repetitive sequences (up to 33 nearly identical 102‐nucleotide repeats) of individual TALe genes. Thus, only a limited number of TALe genes have been molecularly cloned and characterized, and the functions of most TALe genes remain unknown. Here, we present an easy and efficient cloning technique to clone TALe genes selectively through in vitro homologous recombination and single‐strand annealing, and demonstrate the feasibility of this approach with four different Xoo strains. Based on the Gibson assembly strategy, two complementary vectors with scaffolds that can preferentially capture all TALe genes from a pool of genomic fragments were designed. Both vector systems enabled cloning of a full complement of TALe genes from each of four Xoo strains and functional analysis of individual TALes in rice in approximately 1 month compared to 3 months by previously used methods. The results demonstrate a robust tool to advance TALe biology and a potential for broad usage of this approach to clone multiple copies of highly competitive DNA elements in any genome of interest.
Keywords: Xanthomonas, TAL effectors, Gibson assembly, rice
Introduction
Phytopathogenic bacteria of the genus Xanthomonas cause a variety of plant diseases, including those inflicting severe losses on economically important crop plants worldwide (Leyns et al., 1984). In rice, Xanthomonas oryzae pv. oryzae (Xoo) invades xylem to cause bacterial blight of vascular disease, while the closely related pathogen X. oryzae pv. oryzicola (Xoc) is a mesophyll colonizer and causes a disease known as bacterial leaf streak (Niño‐Liu et al., 2006). Individual strains of both Xoo and Xoc contain multiple (e.g. up to 29 in a typical Xoc strain) genes of the Transcription Activator‐Like effector (TALe) family, a few of which are known virulence or/and avirulence effectors depending on the genetic context of host plants (White, 2016). TALes depend on a type III secretion system for their translocation to host cells (Zhang et al., 2015). Typically, once internalized into nuclei of rice cells, TALes act upon the effector‐binding elements (EBEs) in the promoters of host genes (Boch et al., 2010). The general hypothesis is that TALes have evolved to target specific host genes, whose subsequent ectopic expression facilitates infection, and, indeed, a number of TALe‐targeted genes have been shown to enhance host susceptibility (Bart et al., 2012; Chakrabarty et al., 1997; Cohn et al., 2014; Cox et al., 2017; Hu et al., 2014; Schwartz et al., 2017; Sugio et al., 2007; Yang et al., 2006; Yu et al., 2011). However, relatively few TALe targets have been shown to be disease susceptibility (S) genes given the large number of TALe genes discovered. Plants have also evolved to turn TALe function against bacterial invasion, and a few TALes trigger host resistance response due to TALe‐mediated expression of the so‐called executor resistance (R) genes, which, in rice, includes Xa27, Xa10 and Xa23 (Gu et al., 2005; Tian et al., 2014; Wang et al., 2015). TALes are also recognized by members of the common NBS LRR family of R genes (e.g. Xa1) (Ji et al., 2016). A group of TALe variants or iTALes (interfering TALes or truncated TALes) from Xoo and Xoc can suppress NBS LRR‐mediated TALe recognition and resistance (Ji et al., 2016; Read et al., 2016). However, additional mechanisms by which a large number of TALes are involved in host–pathogen interaction remain largely unexplored, and a simple and efficient way to clone and characterize those TALe genes is required.
TALe genes encode three major functional domains. The N‐terminal domain contains the signal for bacterial type III secretion, followed by the central tandem repetitive region, which specifies the target nucleotide sequence of host genes, and the C‐terminal domain containing nuclear localization signals and transcription activation domain, the latter characteristic of features of eukaryotic transcription factors (Boch et al., 2010). The specificity of interaction between the TALes and their host target DNA is determined by the combination of the number of central repeats and composition of two amino acids at the 12th and 13th positions of repeats of TALes known as the repeat variable diamino (RVD) acid unit (Boch et al., 2009; Moscou and Bogdanove, 2009). Coding sequences of TALes and the SphI or BamHI restriction sites flanking the central or almost whole region across different Xanthomonas subspecies or pathovars are highly conserved. Often, the native TALe genes are not cloned, rather the repeat domains are cloned using the conserved SphI or BamHI sites of both ends of TALe genes. The cloned SphI or BamHI fragment fused with a common scaffold of sequences for N‐terminal and C‐terminal domains of TALe bestows specificity of the new TALe gene (Hopkins et al., 1992; Yang and White, 2004). The characterization of TALe genes from various Xanthomonas strains has primarily involved cloning by library construction followed by hybridization or PCR detection based on a conserved TALe sequence (De Feyter et al., 1993; Leach et al., 1992; Tran et al., 2018b; Yang and White, 2004; Yu et al., 2015). Cloning and screening for the right TALe genes are technically challenging due to the existence of many copies of TALe genes in a genome of Xanthomonas and excessive near‐identical tandem repeats in individual TALe genes. Sequencing through the whole central repetitive region of TALe genes is difficult. The whole cloning process is also time‐consuming and inefficient, taking more than 3 months to clone and test a TALe gene from a given Xanthomonas strain.
To improve the efficiency of cloning TALe genes from complex genomes, in the present study we adapted the Gibson assembly strategy using the conserved nature of TALe genes. The Gibson assembly method is a robust molecular cloning technology alternative to restriction/ligation subcloning (Gibson et al., 2009). The method depends on an exonuclease to excise the 5′ nucleotides of double‐stranded DNA (dsDNA) to produce a 3′ single‐strand overhang, which allows complementarity or annealing to the single strand overhang of an adjacent fragment also caused by the exonuclease. A DNA polymerase extends the nucleotides at the 3′ overhangs, and a DNA ligase is used to seal the nicks. The three reactions can be executed in a single tube using a programmed protocol in a thermocycler, and the reaction can be transferred directly into host cells for replication of the plasmids (Gibson et al., 2009). The strategy was applied to selectively clone the full complements of TALe genes from four strains of Xoo, representing the Asian and African lineages of the pathogen. The resulting TALe genes were then assessed for their abilities to restore virulence to a TALe‐defective mutant of Xoo.
Results
Similarity of TAL effector genes in Xanthomonas
Gibson assembly requires the overlapping sequences of approximately 20 bp between two adjacent DNA fragments, and the process is summarized in Fig. 1. The TALe gene sequences were retrieved from genome data of 33 Xoo, ten Xoc, two X. citri pv. vignicola and one X. citri pv. malvacearum strains in the NCBI (National Center for Biotechnology Information) database (Supplementary Table S1). The TALe gene content of 22 Asian Xoo strains and 11 African Xoo strains ranges from 10 to 20 and 9, respectively, in each strain. Xoc strains contain a range of 20–29 TALe genes. Most TALe genes contain two BamHI restriction sites (GGATCC), one at the ATG start codon and another approximately 150 bp upstream of the stop codon, and two SphI sites (GCATGC), one 33 bp upstream of the first repeat and the second one located about 450 bp downstream of the last repeat. Sequences around these restriction sites are also very conserved, almost 99% identical. However, some rare TALe genes contain single nucleotide polymorphism (SNP) within one of the SphI or BamHI recognition sequences. Only 40 out of 733 TALe genes (5.4%) contained a 1‐bp variation in one of the two BamHI restriction sites (GGATCC to GGATCT). Fifty‐seven of 733 TALe genes (7.8%) contain a SNP within one of the SphI restriction sequences (Supplementary Table S1). The loss of either BamHI or SphI sites prevents cloning of the BamHI or SphI fragment of corresponding TALe gene in a library made from either BamHI or SphI DNA fragments. The results indicate that the majority (>92%) of TALe genes can be cloned through capture of either BamHI or SphI fragments of full‐length genes (Supplementary Table S1), and rare TALe gene variants can be retrieved using other restriction enzyme combinations.
Figure 1.
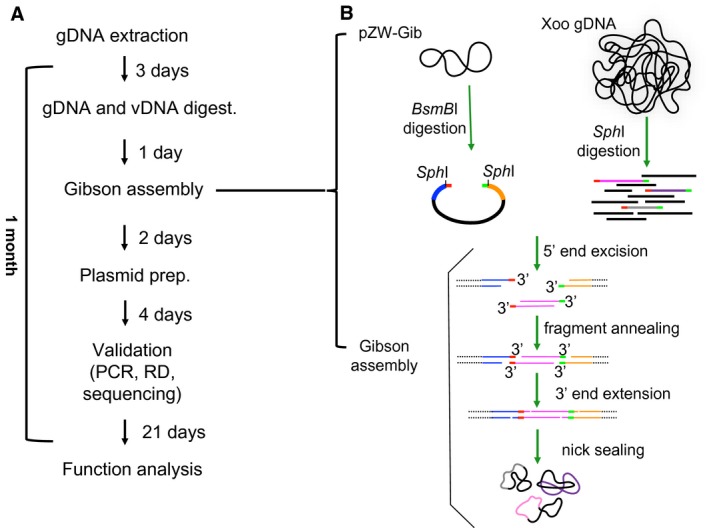
Flow chart and schematics of selectively isolating TALe genes using the Gibson assembly method. (A) Steps and timing of cloning and analysis of TALe genes from a given Xanthomonas oryzae pv. oryzae (Xoo) genome. (B) The selective cloning of SphI fragments of TALe genes. BsmBI digestion yields a cloning vector containing a homologous end (hook, coloured end) at each side beyond the SphI site; SphI digestion of genomic DNA of Xoo releases TALe fragments with both ends containing short sequences complementary to the hooks of cloning vector pZW‐Gib. Gibson assembly results in plasmids containing individual TALe genes. gDNA, genomic DNA; vDNA, vector DNA, RD, restriction enzume digestion.
Strategy to selectively clone TALe genes
We next developed a system to selectively isolate the genomic fragments, e.g. SphI or BamHI digested fragments, of TALe genes from a genome. The technique involves in vitro linking of a TALe fragment and a vector fragment with short (~20 bp) 5′ and 3′ end sequences matching to both ends of TALe gene fragments by annealing of the 3′ complementary overhangs produced by T5 exonuclease. The single‐stranded overhangs of the vector fragment function as sequence‐specific hooks to catch the corresponding overhangs of TALe fragments selectively from a pool of the genomic fragments. To make a cloning vector for selective cloning of the SphI fragments of TALe genes, the previously cloned TALe gene pthXo1, which encodes the major virulence effector for the Asian Xoo strain PXO99A (Yang and White, 2004), was used as the backbone of the TALe gene vector without the central SphI repetitive region. In addition, a counter selective gene was introduced to avoid undigested cloning vector or vector insert. First, a DNA fragment (gBlock) was synthesized that contained the ccdB gene, flanked by a sequence (5′‐GCATGCATGGCGCAATGCACTGACGGGTGCA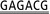 ‐3′) 23 nucleotides (nt) downstream of the first SphI (underlined) and a sequence (5′‐
‐3′) 23 nucleotides (nt) downstream of the first SphI (underlined) and a sequence (5′‐ AACGCCGGATCAGGCGTCTTTGCATGC‐3′) 20 nucleotides upstream of the second SphI of pthXo1. The two ccdB flanking sequences between SphI and BsmBI (double underlined) sites are highly conserved among the natural TALe genes (Supplementary Fig. S1A). The ccdB gene encodes a toxic protein (CcdB) to cause cell death in certain Escherichia coli strains (Bernard and Couturier, 1992). Inclusion of ccdB ensures loss of any clones that retains the region of ccdB when transferred to a strain lacking the antidote ccdA gene, in this case E. coil XL1‐Blue. Digestion of vector with BsmBI releases two ends (23 and 20 nt) that are homologous to the two ends of the SphI fragment of the TALe gene. The gBlock was cloned into pZW‐pthXo1 at SphI sites by replacing the central SphI region of pthXo1, resulting in pZW‐Gib. The backbone of TALe in pZW‐Gib contains the conserved ~800‐bp region upstream and the ~400‐bp region downstream of the SphI recognition sites of TALe genes (Fig. 2A). Insertion of the SphI central repeat region of any TALe gene through Gibson assembly should result in functional TALe genes.
AACGCCGGATCAGGCGTCTTTGCATGC‐3′) 20 nucleotides upstream of the second SphI of pthXo1. The two ccdB flanking sequences between SphI and BsmBI (double underlined) sites are highly conserved among the natural TALe genes (Supplementary Fig. S1A). The ccdB gene encodes a toxic protein (CcdB) to cause cell death in certain Escherichia coli strains (Bernard and Couturier, 1992). Inclusion of ccdB ensures loss of any clones that retains the region of ccdB when transferred to a strain lacking the antidote ccdA gene, in this case E. coil XL1‐Blue. Digestion of vector with BsmBI releases two ends (23 and 20 nt) that are homologous to the two ends of the SphI fragment of the TALe gene. The gBlock was cloned into pZW‐pthXo1 at SphI sites by replacing the central SphI region of pthXo1, resulting in pZW‐Gib. The backbone of TALe in pZW‐Gib contains the conserved ~800‐bp region upstream and the ~400‐bp region downstream of the SphI recognition sites of TALe genes (Fig. 2A). Insertion of the SphI central repeat region of any TALe gene through Gibson assembly should result in functional TALe genes.
Figure 2.
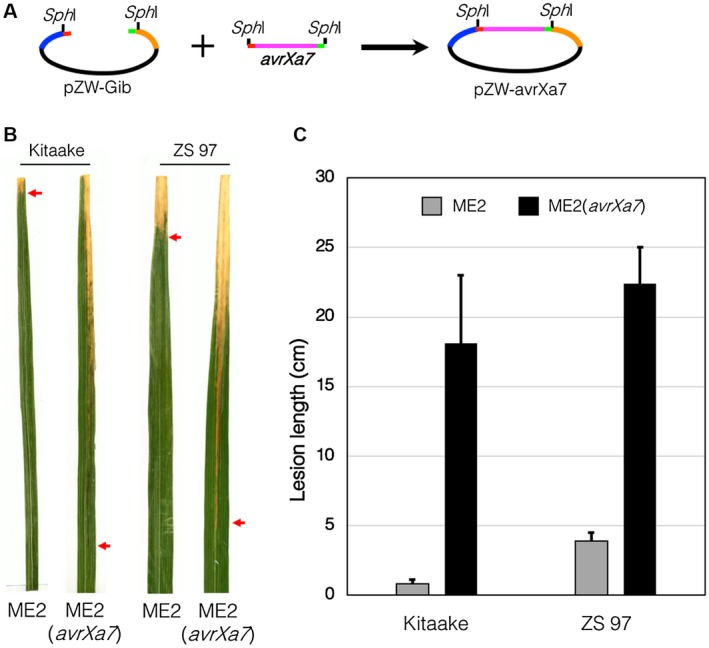
Validation of pZW‐Gib for cloning of a functional TALe gene. (A) The SphI fragment of avrXa7 was cloned into pZW‐Gib, resulting in pZW‐avrXa7. (B) Rice leaves of cultivars Kitaake and Zhenshan 97 (ZS 97) showing disease symptoms caused by Xanthomonas oryzae pv. oryzae (Xoo). (C) Lesion lengths in rice leaves caused by Xoo strain ME2 and its transformant ME2(avrXa7). Ten fully expanded young leaves were inoculated for lesion length measurements 14 days post‐inoculation. The experiment was repeated twice independently with similar results.
To test the system, pZW‐Gib was digested with BsmBI to remove the ccdB gene and used to clone the SphI fragment of the alternate major virulence TALe gene avrXa7, which was originally cloned from the Xoo strain PXO86 (Fig. 2A). The pZW‐Gib‐avrXa7 was introduced via subcloning into pHM1, a Xoo suitable plasmid, and transferred to ME2, a mutant derived from PXO99A with pthXo1 inactivated (Yang and White, 2004). Finally, the function of avrXa7 for virulence, as assembled in pZW‐Gib, was tested in the rice cultivars Kitaake and Zhenshan 97. The results showed that the Gibson‐cloned avrXa7 restored the virulence in ME2 (avrXa7) in terms of lesion lengths and disease symptoms compared to ME2 (Fig. 2B,C).
Isolation and functional test of TALe genes from PXO61
The PXO61 genome of Xoo (NCBI accession, CP033187.1) has over 3000 SphI sites and 18 TALe genes (Supplementary Fig. S3A). All TALe genes contain two SphI sites except one, Tal5b, which contains only one SphI restriction site with the second one missing due to a deletion in the 3′ region. Tal5b belongs to a group of iTALe genes that contribute virulence to Xoo by suppressing Xa1‐mediated disease resistance (Ji et al., 2016). After ligation of the SphI fragments of genomic DNA derived from PXO61 into the BsmBI‐digested pZW‐Gib using the Gibson assembly protocol, bacterial colonies were picked randomly to screen for the presence of SphI fragments of TALe genes (Fig. 3A,B). Twenty‐two out of 26 clones were positive for TALe genes based on PCR with specific primers, restriction enzyme digestion and Sanger sequencing (Fig. 3B). To clone the whole complement of TALe genes (n = 17 except Tal5b) of PXO61, a total of 45 clones were selected for sequencing based on the sizes of SphI fragments of clones and the predicted number of TALe genes in the PXO61 genome. Seventeen of the cloned TALe genes matched the corresponding annotated genes in the PXO61 genome (Fig. 4). The individual TALe genes in pZW‐Gib were inserted into the broad host range vector pHM1, and the constructs transferred to ME2 for functional analysis. One clone contained the previously identified major TALe gene pthXo3 (also known as Tal6C), which conferred disease susceptibility in rice Kitaake and Zhenshan 97 (Figs 5 and S4C), and another clone contained Tal7 (now pthXo2B), encoding a virulence TALe targeting the S gene SWEET13 and confers disease susceptibility only to Kitaake and not Zhenshan 97 (Figs 5 and S4C). The remaining 15 clones conferred no observable phenotype (Figs 5 and S4C).
Figure 3.
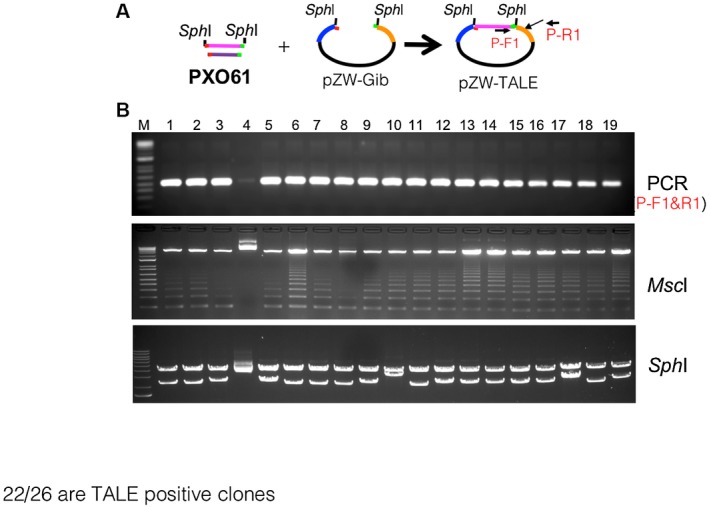
Validation of TALe clones through PCR, restriction enzyme digestions. (A) Schematic of selective isolation of SphI fragments TALe genes from genomic DNA of PXO61. (B) Validation of TALe clones through PCR with primers P‐F1 and P‐R1 of individual clones as indicated above lanes of upper gel image, digestion by MscI which cuts each of central repeats (the DNA band patterns resulted from partial digestion) and digestion by SphI.
Figure 4.

Seventeen of 18 annotated TALe genes from PXO61 were cloned using pZW‐Gib vector and Gibson assembly method. The repeat variable diamino (RVD) acid units of individual TALes are shown under numbers (1 to 29) indicating the order of 33–34 amino acid repeats. Asterisks (*) indicate that the amino acid at the 13th position missing.
Figure 5.
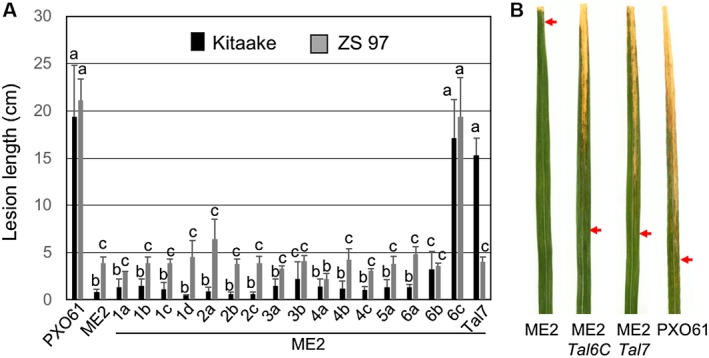
Virulence contribution of 17 TALes cloned from PXO61. (A) Lesion lengths caused in rice cultivars Kitaake and Zhenshan 97 (ZS 97) by different Xanthomonas oryzae pv. oryzae (Xoo) strains indicated below the paired columns. Different lower case letters indicate statistically significant differences (means ± SEM, n = 10, P < 0.05). (B) Blight symptom in Kitaake leaves caused by the Xoo strains as indicated below each leaf. Arrows indicate the edges of lesions.
Selective cloning and function test of TALe genes from the African strain AXO1947
AXO1947 has been sequenced previously and has nine TALe genes and no iTALe gene (Huguet‐Tapia et al., 2016). To further demonstrate the feasibility of pZW‐Gib to selectively clone TALe genes, a sub‐library of TALe genes was constructed by ligating the SphI fragments of genomic DNA from AXO1947 into the BsmBI‐digested pZW‐Gib. All nine TALe genes were retrieved that matched the annotated TALes in AXO1947 (Supplementary Fig. S4B). The nine TALe genes were individually subcloned into pHM1 and mobilized into ME2 for virulence tests. Only the gene corresponding to a homolog of the previously identified major TALe TalC, which targets the S gene SWEET14, conferred virulence to ME2 in Kitaake and Zhenshan 97 (Supplementary Fig. 4D).
Improved vector for cloning TALe genes
Certain TALes, e.g. iTal3a and iTal3b in PXO99A, require their unique N‐terminal and C‐terminal domains for functionality. Two deletions within the N‐termini of iTal3a and iTal3b coding sequences are required to suppress the Xa1‐mediated resistance triggered by TALes (Ji et al., 2016). Other TALe genes simply lack one SphI site in their DNA sequences due to DNA polymorphisms. The vector pZW‐Gib is not able to capture such sequences from the genomes. Furthermore, pZW‐Gib derived TALe clones need to be moved into a broad host range vector, pHM1 in this case, for further characterization of function in Xoo. To overcome such constraints, a cloning vector capable of capturing of the BamHI fragments and avoiding the lack of SphI restriction site of natural TALe genes and direct transfer to Xoo was devised, using pHM1 as the recipient vector for TALe BamHI fragments. Differing from pZW‐Gib, pHM1‐Gib contains the promoter sequence, the 3′ region downstream of the second BamHI site of pthXo1 from PXO99A, and the ccdB cassette. Additionally, a sequence of ColE1 (origin of replication from pBluescript) was integrated into pHM1‐Gib to increase the copy number of plasmids in E. coli, which facilitates DNA isolation and manipulation (Supplementary Fig. S1B). For validation, the BamHI fragment of pthXo2B was retrieved from the BamHI digestion of PXO61 genomic DNA in pHM1‐Gib through Gibson assembly. The resultant plasmids were directly transferred into ME2, and each transformant tested for the ability to cause disease in Kitaake. The newly assembled pthXo2B directed induction of SWEET13. To further demonstrate the improved capability of pHM1‐Gib, all nine TALe genes from two Africa strains CFBP7321 and CFBP7325 were cloned in the vector. Thirty‐nine out of 48 clones in CFBP7325 and 45 out of 48 in CFBP7321, respectively, were positive for TALe‐containing sequences (Supplementary Figs [Link], [Link], [Link]). A disease assay confirmed the presence of clones for TalC and TalF, both previously identified major TALe genes, which could direct disease in rice Kitaake (Supplementary Fig. S8).
Discussion
In the present study, we present two complementary vector systems for cloning TALe gene fragments selectively from a genomic pool of fragments through in vitro homologous recombination and single‐strand annealing, a protocol known as the Gibson assembly method. The strategy depends on exonuclease (e.g. T5 exonuclease) to generate the 3′ end overhangs by chewing back the 5′ nucleotides of vector fragments and genomic fragments, and the assembly of vector fragments and selectively the TALe gene fragments in a single reaction. Each vector system has been demonstrated as functional by reproducibly applying the vector to multiple Xoo strains. Approximately 87% (n = 122) of the clones are positive for TALe gene fragments, and a total of 44 TALe genes out of 45, with Tal5b of PXO61 missing, were retrieved from four Xoo genomes. The results indicate both plasmids are highly efficient and time saving compared to the conventional cloning methods for cloning of TALe genes from a given Xanthomonas genome. The strategy and even the specific vector system are applicable to other Xanthomonas species and pathovars (e.g. Xoc) due to the conserved nucleotide sequences around the SphI and BamHI sites of the TALe genes. Some species with a more divergent TALe sequence, including X. translucens, may require customization of the vector annealing regions (Peng et al., 2016).
The prior studies reported the difficult and time‐consuming task in cloning a complement of TALe gene fragments from a given genome largely due to a large number of TALe genes that were highly conserved in a given genome and the large number of central tandem repeats of individual TALe genes. The cloning was performed by making genomic libraries followed by Southern hybridization‐ or PCR‐based screening, which is laborious and inefficient. Due to this difficulty, only a few from a large number of sequenced and annotated Xanthomonas genomes have been subjected to systematic TALe gene cloning and characterization in contrast to Cernadas et al. (2014), Yang and White (2004) and Yu et al. (2015). For example, Yu et al. pulled out 115 positive clones (representing 18 unique TALe genes) from 3000 individual transformant colonies (c. 4%) through screening a library of BamHI fragments of Xoo K74 genome using dot‐ and Southern blotting (Yu et al., 2015). A similar but improved approach was developed to enrich the BamHI fragments of TALe genes from a given genome by applying two additional restriction enzymes (ApaLI and SfoI) to further cut the non‐TALe gene fragments of genomic DNA. Ligation and screening by PCR yielded 25.0% and 26.9% positive clones of TALe gene fragments for MAI1 and BAI3 strains of Xoo, respectively (Tran et al., 2018a). Both methods require further subcloning of the TALe BamHI fragments from the pUC‐based vector into TALe gene scaffold and Xanthomonas‐suitable vector for function analysis.
TALes represent the largest type III effector family of pathogenic bacteria, and some TALes have been found to be crucial to help bacteria to infect crop plants (Boch et al., 2010). Such important TALes represent attractive targets to create disease resistance in host crop plants by interfering with the disease mechanism (Schornack et al., 2013). Naturally occurring or gene editing‐derived alleles with EBE variations, if disruptive to TALe binding, in the promoters of S genes targeted by TALes confer genetically recessive resistance (Blanvillain‐Baufumé et al., 2017; Chu et al., 2006; Li et al., 2012; Liu et al., 2011). Multiplex genome editing produced single engineered rice lines that carried multiple mutations in three SWEET gene promoters and resulted in broad‐spectrum blight resistance (R. Oliva et al., unpublished data). Executor R genes along with promoters containing the synthetic EBEs have been used to trap the widely spread cognate TALes for induced resistance (Hummel et al., 2012; Römer et al., 2009). Deployment of such resistances (dominant, recessive, naturally occurring or artificially made) against TALes creates tremendous selection pressure on the dynamic populations of Xanthomonas pathogens. The repetitive and multigenic nature provides a repertoire of TALe genes in a given genome to evolve and potentially overcome the resistances. Broad and durable resistance depends on effectively monitoring the evolving pathogenic populations and quickly identifying newly evolving TALes, monitoring tools including a diagnostic kit comprising seven components based on TALe‐mediated disease mechanism (J. Eom et al., unpublished data) and the cloning methods described here. Additionally, the Gibson‐based vectors developed here have potential for high‐throughput profiling of complement TALe genes from multiple Xanthomonas strains when bar‐coded and combined with next‐generation sequencing technologies, including PacBio and Nanopore platforms. Development of a simple, cheap and efficient cloning system will also advance our understanding of TALe biology.
Experimental Procedures
Plant materials, bacterial strains and growth conditions
Rice (Oryza sativa) cultivar Kitaake and Zhenshan 97 were used in this study. Plasmids, Xoo and E. coli strains used in this study are listed in Supplementary Table S2. Rice plants were grown in a growth chamber at a temperature of 28 °C, relative humidity of 85% and a photoperiod of 12 h. Escherichia coli DB3.0 and Trans1‐T1 were grown in Luria–Bertani medium at 37 °C. Xoo strains were grown in either nutrient broth (NB: beef extract 3 g/L, peptone 5 g/L) or tryptone sucrose medium (TSA: tryptone 10 g/L, sucrose 10 g/L, glutamic acid 1 g/L) at 28 °C. Antibiotics used in this study include spectinomycin (100 mg/L) and ampicillin (100 mg/L) for bacterial strains containing appropriate plasmids.
DNA manipulation and plasmid construction
DNA manipulation and PCR were conducted according to standard protocols (Ausubel et al., 1998). Genomic DNA from Xoo was extracted from bacterial cells grown in NB medium using the MagAttract HMW DNA kit (Qiagen, Hilden, Germany) following the manufacturer’s manual. Plasmids were introduced by electroporation into X. oryzae and E. coli bacterial competent cells as described previously (Yang and White, 2004). Primers for PCR were synthesized by Integrated DNA Technologies (Coralville, IA, USA). PCR was performed using Taq DNA polymerase or Phusion High‐Fidelity DNA polymerase (New England BioLabs, Ipswich, MA, USA).
Construction of pZW‐Gib
The central repeat region of pthXo1was replaced by a gBlock, which contained a ccdB sequence, as follows. First the 3′ fragment of a TAL effector gene was PCR‐amplified using the primers TalAatII‐F and TalFlH3‐R along the genomic DNA of PXO99A. The PCR amplicon was cloned into pZW‐pthXo1 at AatII and HindIII sites through Gibson assembly. The ligation reaction was transferred into E. coli Trans1‐T1 cells. The positive bacterial colonies were screened by colony PCR using primers TalAatII‐F and TALFLH3‐R. Candidate clones were sequenced for confirmation of PthXo1 downstream sequence. A gBlock fragment (Supplementary Fig. S1A) synthesized by Integrated DNA Technologies was cloned into the new pZW‐pthXo1 at SphI sites by Gibson cloning. The ligation reaction was transferred into E. coil DB3.0 cells. Candidate clones were sequenced for confirmation of gBlock sequence.
Construct pZW‐Gib‐avrXa7 via Gibson cloning
The central repeat region of avrXa7 was obtained by digesting pZW‐avrXa7 with SphI and selecting a DNA fragment of right size through electrophoresis in 1% agarose gel. The SphI fragment was cloned into the backbone of pZW‐Gib that was derived from BsmBI‐digestion and purification from 1% agarose gel. The ligation was transferred into E. coli Trans1‐T1 cells; positive colonies were first screened via PCR using primer P‐F and P‐R, SphI digestion and finally sequencing for confirmation of avrXa7 insertion. Primer sequences are provided in Supplementary Table S3.
Construct TALe gene sublibraries of PXO61 and AXO1947
Genomic DNA of PXO61 and AXO1947 was digested with SphI and the fragments were separated through electrophoresis in 1% agarose gel. DNA fragments of about 2–5 kb were extracted and purified from the agarose gel and subcloned through Gibson assembly into the backbone of pZW‐Gib that was BsmBI‐digested and gel‐purified. The Gibson Assembly Cloning kit (New England Biolabs) was used following the manufacturer’s manual. The subsequent process was identical to the construction of pZW‐Gib‐avrXa7. Finally, a select set of clones was chosen based on the expected sizes of repeats as determined by SphI digestion and sequenced using primers Tal‐SphI‐F and P‐R.
Transform X. oryzae cells with plasmid DNA
Competent cells of Xoo ME2 were prepared and transformation was performed as previously described (Ji et al., 2016). Briefly, TSA + NB medium (NB 8 g/L, tryptone 10 g/L, sucrose 10 g/L, glutamic acid 1 g/L) was used to grow Xoo cells. An aliquot of 50 µL of bacterial competent cells was mixed with 10 ng (0.5 µL) of plasmid DNA for electroporation using a Bio‐Rad electroporation instrument (Bio‐Rad, Hercules, CA, USA). An electric field of 15 kV/cm with a resistance of 200 Ω and a capacitance of 25 µF was applied. After pulse delivery, cells were immediately transferred into 1 mL of SOC medium (20 g tryptone, 5 g yeast extract, 4.8 g MgSO4, 3.6 g dextrose, 0.5 g NaCl and 0.2 g KCl per L) in a 2 mL round‐bottom polypropylene tube. After incubation at 28 °C with constant shaking for 2–4 h, the electroporated cells were plated onto TSA‐NB medium containing the appropriate antibiotics and incubated at 28 °C for 3–5 days. Non‐electroporated competent cells were used as a control. Positive clones were picked for further analysis.
Disease assays
Disease assays to test the function of cloned TALe genes were conducted as follows. Briefly, Xoo strains were grown in TSA + NB with appropriate antibiotics at 28 °C. Bacterial cells were collected from culture through low‐speed (2300 g) centrifugation, washed twice and suspended in sterile water. The suspensions were adjusted to an optical density of 0.5 at 600 nm (OD600) and were used to inoculate into fully expanded leaves of 7–8‐week‐old rice plants using the leaf‐tip clipping method (Kauffman et al., 1973). Lesion lengths were measured about 12–14 days post‐inoculation. The disease assays were performed independently at least twice and on at least four plants each time. One‐way analysis of variance statistical analysis was performed on all measurements. The Tukey’s honestly significant difference test was used for post‐analysis of variance pairwise tests for significance, set at 5% (P < 0.05).
Sequence analysis of TALe genes in sequenced Xanthomonas genomes
Genomes of Xanthomonas strains were obtained from the NCBI under accession numbers Xoo OS198: CP031461.1, Xoo JL25: CP031457.1, Xoo PX086: CP031463.1, Xoo PX079: CP031462.1, Xoo YC11: CP031464.1, Xoo JL33: CP031459.1, Xoo JP01: CP031460.1, Xoo HuN37: CP031456.1, Xoo JL28: CP031458.1, Xoo BAI3: CP025610.1, Xoo MAI1: CP025609.1, Xoo MAI145: CP019092.1, Xoo MAI134: CP019091.1, Xoo MAI129: CP019090.1, Xoo MAI106: CP019089.1, Xoo MAI99: CP019088.1, Xoo MAI95: CP019087.1, Xoo MAI73: CP019086.1, Xoo MAI68: CP019085.1, Xoo PXO61: CP021789.1, Xoo PXO145: CP013961.1, Xoo AXO1947: CP013666.1, Xoo XF89b: CP011532.1, Xoo PXO602: CP013679.1, Xoo PXO563: CP013678.1, Xoo PXO524: CP013677.1, Xoo PXO282: CP013676.1, Xoo PXO236: CP013675.1, Xoo PXO211: CP013674.1, Xoo PXO71: CP013670.1, Xoo PXO83: CP012947.1, Xoo PXO99A: CP000967.2, Xoo PXO86: CP007166.1, Xoo MAFF 311018: NC_007705.1, Xoo KACC 10331: AE013598.1, Xoc CFBP7331: CP011958.1, Xoc CFBP2286: CP011962.1, Xoc RS105: CP011961.1, Xoc L8: CP011960.1, Xoc CFBP7341: CP011959.1, Xoc BXOR1: CP011957.1, Xoc BLS279: CP011956.1, Xoc B8‐12: CP011955.1, Xoc CFBP7342: CP007221.1, Xoc BLS256: CP003057.2, Xcv CFBP7113: CP022270.1, Xcv CFBP7112: CP022269.1, Xcm XcmH1005: CP013004.1.
Analysis of all TALe gene sequences was performed through NCBI blast and further confirmed by using SnapGene software. Sequences used for comparison were 5′‐GCATGCATGGCGCAATGCACTGACGGGTGC‐3′ and 5′‐ ACGCCGGATCAGGCGTCTTTGCATGC‐3′ around the two SphI restriction sites, while the pair of sequences 5′‐GGATCCCATTCGTTCGCGCACGCCAAGTCCTGCCCGCG‐3′ and 5′‐ACCAGGATCGGGGGCGGCCTCCCGGATCC‐3′ were used around the two BamHI sites of TALe genes.
Supporting information
Fig. S1 DNA sequences of gBlock fragments synthesized to make two Gibson cloning vectors. (A) gBlock1 was used to inserted into pZW‐pthXo1 at SphI sites through Gibson cloning. The two sequences shaded in yellow are homologous to the ends of SphI fragments of TALe genes. (B) gBlock2 was used to make pHM1‐Gib. The two sequences shaded in yellow are homologous to the ends of BamHI fragments of TALe genes.
Fig. S2 Nine TALe genes from AXO1947 were cloned using the pZW‐Gib vector and Gibson assembly method. The RVDs of individual TALes are shown under numbers 1 to 26, indicating the order of 33–34 amino acid repeats. Asterisks (*) indicate that the amino acid at the 13th position missing.
Fig. S3 Virulence contribution of nine TALes cloned from AXO1947. (A) Lesion lengths caused in rice Kitaake and Zhenshan 97 (ZS 97) by different Xoo strains indicated below the paired columns. Different lower letters indicate statistically significant differences (mean SEM, n = 10, P < 0.05). (B) Blight symptom in Kitaake leaves caused by the Xoo strains as indicated below each leaf. Arrows indicate the edges of lesions.
Fig. S4 TALe genes from PXO61 and AXO1947. (A) TALe genes clustered in the genome of PXO61. (B) TALe genes clustered in the genome of AXO1947. (C, D) Lesion length measurements (mean SEM, n = 10) in Kitaake caused by different strains.
Fig. S5 Validation of TALe clones through PCR, restriction enzyme digestions. (A) Schematics of selective isolation of BamHI fragment TALe genes from genomic DNA of CFBP7321. (B) Validation of TALe clones through PCR with primers P‐F and P‐R of individual clones as indicated above lanes of upper gel image, digestion by MscI which cuts each of central repeats (the DNA band patterns resulted from partial digestion) and digestion by BamHI.
Fig. S6 Validation of TALe clones through PCR, restriction enzyme digestions. (A) Schematics of selective isolation of BamHI fragments TALe genes from genomic DNA of CFBP7325. (B) Validation of TALe clones through PCR with primers P‐F1 and P‐R1 of individual clones as indicated above lanes of upper gel image, digestion by MscI which cuts each of central repeats (the DNA band patterns resulted from partial digestion) and digestion by BamHI.
Fig. S7 TALe gene distribution in two African Xoo genomes. (A, B) Nine TALe genes in each of two Xoo genomes are syntenic in their locations with the same colors denoting identical TALes at the amino acid level while different colors indicate different TALes at the amino acid level.
Fig. S8 Two TALe genes cloned with the pHM1‐Gib system were functional in virulence. The virulences of TalC from CFBP7321 and TalF from CFBP7325 were tested in Kitaake leaves. Different letters indicate statistically significant differences.
Table S1 Number of TALe genes from different Xanthomonas that can be selectively cloned with Gibson assembly.
Table S2 Plasmids and bacterial strains used in this study.
Table S3 Primers used in this study.
Acknowledgements
The work was partially supported by subawards to University of Missouri and University of Florida from the Heinrich Heine University of Dusseldorf funded by the Bill & Melinda Gates Foundation [OPP1155704] (B.Y., F.F.W) and the China Scholar Council (C.L., as a point PhD student). The authors are grateful to Wolf F. Frommer for critical reading and constructive comments on the manuscript.
Contributor Information
Hansong Dong, Email: hsdong@njau.edu.cn.
Bing Yang, Email: yangbi@missouri.edu.
References
- Ausubel, F. , Brent, R. , Kingston, R. , Moore, D. , Seidman, J. and Struhl, K. (1998) Current Protocols in Molecular Biology. New York: John Wiley and Sons. [Google Scholar]
- Bart, R. , Cohn, M. , Kassen, A. , McCallum, E.J. , Shybut, M. , Petriello, A. , Krasileva, K. , Dahlbeck, D. , Medina, C. , Alicai, T. , Kumar, L. , Moreira, L.M. , Rodrigues Neto, J. , Verdier, V. , Santana, M.A. , Kositcharoenkul, N. , Vanderschuren, H. , Gruissem, W. , Bernal, A. and Staskawicz, B.J. (2012) High‐throughput genomic sequencing of cassava bacterial blight strains identifies conserved effectors to target for durable resistance. Proc. Natl. Acad. Sci. USA. 109, E1972–E1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard, P. and Couturier, M. (1992) Cell killing by the F plasmid CcdB protein involves poisoning of DNA‐topoisomerase II complexes. J. Mol. Biol. 226, 735–745. [DOI] [PubMed] [Google Scholar]
- Blanvillain‐Baufumé, S. , Reschke, M. , Solé, M. , Auguy, F. , Doucoure, H. , Szurek, B. , Meynard, D. , Portefaix, M. , Cunnac, S. , Guiderdoni, E. , Boch, J. and Koebnik, R. (2017) Targeted promoter editing for rice resistance to Xanthomonas oryzae pv. oryzae reveals differential activities for SWEET14‐inducing TAL effectors. Plant Biotechnol. J. 15, 306–317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boch, J. , Scholze, H. , Schornack, S. , Landgraf, A. , Hahn, S. , Kay, S. , Lahaye, T. , Nickstadt, A. and Bonas, U. (2009) Breaking the code of DNA binding specificity of TAL‐type III effectors. Science, 326, 1509–1512. [DOI] [PubMed] [Google Scholar]
- Boch, J. , Bonas, U. , VanAlfen, N. , Bruening, G. and Leach, J. (2010) Xanthomonas AvrBs3 family‐type III effectors: Discovery and function. Annu. Rev. Phytopathol. 48, 419–436. [DOI] [PubMed] [Google Scholar]
- Cernadas, R.A. , Doyle, E.L. , Niño‐Liu, D.O. , Wilkins, K.E. , Bancroft, T. , Wang, L. , Schmidt, C.L. , Caldo, R. , Yang, B. , White, F.F. , Nettleton, D. , Wise, R.P. and Bogdanove, A.J. (2014) Code‐assisted discovery of TAL effector targets in bacterial leaf streak of rice reveals contrast with bacterial blight and a novel susceptibility gene. PLoS Pathog. 10, e1003972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakrabarty, P.K. , Duan, Y.P. and Gabriel, D.W. (1997) Cloning and characterization of a member of the Xanthomonas avr/pth gene family that evades all commercially utilized cotton R genes in the United States. Phytopathology, 87, 1160–1167. [DOI] [PubMed] [Google Scholar]
- Chu, Z. , Yuan, M. , Yao, J. , Ge, X. , Yuan, B. , Xu, C. , Li, X. , Fu, B. , Li, Z. , Bennetzen, J.L. , Zhang, Q. and Wang, S. (2006) Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Genes Dev. 20, 1250–1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohn, M. , Bart, R. , Shybut, M. , Dahlbeck, D. , Gomez, M. , Morbitzer, R. , Hou, B. , Frommer, W. , Lahaye, T. and Staskawicz, B. (2014) Xanthomonas axonopodis virulence is promoted by a transcription activator‐like effector mediated induction of a SWEET sugar transporter in cassava. Mol. Plant‐Microbe Interact. 27, 1186–1198. [DOI] [PubMed] [Google Scholar]
- Cox, K.L. , Meng, F. , Wilkins, K.E. , Li, F. , Wang, P. , Booher, N.J. , Carpenter, S.C.D. , Chen, L.Q. , Zheng, H. , Gao, X. , Zheng, Y. , Fei, Z. , Yu, J.Z. , Isakeit, T. , Wheeler, T. , Frommer, W.B. , He, P. , Bogdanove, A.J. and Shan, L. (2017) TAL effector driven induction of a SWEET gene confers susceptibility to bacterial blight of cotton. Nat. Commun. 8, 15588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Feyter, R. , Yang, Y. and Gabriel, D.W. (1993) Gene‐for‐genes interactions between cotton R genes and Xanthomonas campestris pv. malvacearum avr genes. Mol Plant‐Microbe Interact. 6, 225–237. [DOI] [PubMed] [Google Scholar]
- Gibson, D.G. , Young, L. , Chuang, R.Y. , Venter, J.C. , Hutchison, C.A. and Smith, H.O. (2009) Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods. 6, 343–345. [DOI] [PubMed] [Google Scholar]
- Gu, K. , Yang, B. , Tian, D. , Wu, L. , Wang, D. , Sreekala, C. , Yang, F. , Chu, Z. , Wang, G.L. , White, F.F. and Yin, Z. (2005) R gene expression induced by a type‐III effector triggers disease resistance in rice. Nature, 435, 1122–1125. [DOI] [PubMed] [Google Scholar]
- Hopkins, C.M. , White, F.F. , Choi, S.H. , Guo, A. and Leach, J.E. (1992) Identification of a family of avirulence genes from Xanthomonas oryzae pv. oryzae . Mol. Plant‐Microbe Interact. 5, 451–459. [DOI] [PubMed] [Google Scholar]
- Hu, Y. , Zhang, J. , Jia, H. , Sosso, D. , Li, T. , Frommer, W. , Yang, B. , White, F. , Wang, N. and Jones, J. (2014) Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc. Natl. Acad. Sci. USA. 111, E521–E529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huguet‐Tapia, J.C. , Peng, Z. , Yang, B. , Yin, Z. , Liu, S. and White, F.F. (2016) Complete genome sequence of the African strain AXO1947 of Xanthomonas oryzae pv. oryzae . Genome Announc. 4, e01730–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hummel, A.W. , Doyle, E.L. and Bogdanove, A.J. (2012) Addition of transcription activator‐like effector binding sites to a pathogen strain‐specific rice bacterial blight resistance gene makes it effective against additional strains and against bacterial leaf streak. New Phytol. 195, 883–893. [DOI] [PubMed] [Google Scholar]
- Ji, Z. , Ji, C. , Liu, B. , Zou, L. , Chen, G. and Yang, B. (2016) Interfering TAL effectors of Xanthomonas oryzae neutralize R‐gene‐mediated plant disease resistance. Nat. Commun. 7, 13435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kauffman, H.E. , Reddy, A.P.K. , Hsieh, S.P.Y. and Merca, S.D. (1973) An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae . Plant Dis. Report. 57, 5. [Google Scholar]
- Leach, J.E. , Rhoads, M.L. , Vera Cruz, C.M. , White, F.F. , Mew, T.W. and Leung, H. (1992) Assessment of genetic diversity and population structure of Xanthomonas oryzae pv. oryzae with a repetitive DNA element. Appl. Environ. Microbiol. 58, 2188–2195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leyns, F. , De Cleene, M. , Swings, J.G. and Ley, J.D. (1984) The Host Range of the Genus Xanthomonas. New York: Botanical Garden Press. [Google Scholar]
- Li, T. , Liu, B. , Spalding, M.H. , Weeks, D.P. and Yang, B. (2012) High‐efficiency TALEN‐based gene editing produces disease‐resistant rice. Nat. Biotechnol. 30, 390–392. [DOI] [PubMed] [Google Scholar]
- Liu, Q. , Yuan, M. , Zhou, Y. , Li, X. , Xiao, J. and Wang, S. (2011) A paralog of the MtN3/saliva family recessively confers race‐specific resistance to Xanthomonas oryzae in rice. Plant Cell Environ. 34, 1958–1969. [DOI] [PubMed] [Google Scholar]
- Moscou, M.J. and Bogdanove, A.J. (2009) A simple cipher governs DNA recognition by TAL effectors. Science, 326, 1501. [DOI] [PubMed] [Google Scholar]
- Niño‐Liu, D.O. , Ronald, P.C. and Bogdanove, A.J. (2006) Xanthomonas oryzae pathovars: model pathogens of a model crop. Mol. Plant Pathol. 7, 303–324. [DOI] [PubMed] [Google Scholar]
- Peng, Z. , Hu, Y. , Xie, J. , Potnis, N. , Akhunova, A. , Jones, J. , Liu, Z. , White, F.F. and Liu, S. (2016) Long read and single molecule DNA sequencing simplifies genome assembly and TAL effector gene analysis of Xanthomonas translucens . BMC Genom. 17, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Read, A.C. , Rinaldi, F.C. , Hutin, M. , He, Y.Q. , Triplett, L.R. and Bogdanove, A.J. (2016) Suppression of Xo1‐mediated disease resistance in rice by a truncated, non‐DNA‐binding TAL effector of Xanthomonas oryzae . Front. Plant Sci. 7, 1516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Römer, P. , Recht, S. and Lahaye, T. (2009) A single plant resistance gene promoter engineered to recognize multiple TAL effectors from disparate pathogens. Proc. Natl. Acad. Sci. USA. 106, 20526–20531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schornack, S. , Moscou, M. , Ward, E. , Horvath, D. and VanAlfen, N. (2013) Engineering plant disease resistance based on TAL effectors. Annu. Rev. Phytopathol. 51, 383–406. [DOI] [PubMed] [Google Scholar]
- Schwartz, A. , Morbitzer, R. , Lahaye, T. and Staskawicz, B. (2017) TALe‐induced bHLH transcription factors that activate a pectate lyase contribute to water soaking in bacterial spot of tomato. Proc. Natl. Acad. Sci. USA. 114, E897–E903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugio, A. , Yang, B. , Zhu, T. and White, F. (2007) Two type III effector genes of Xanthomonas oryzae pv. oryzae control the induction of the host genes OsTFIIA gamma 1 and OsTFX1 during bacterial blight of rice. Proc Natl Acad Sci USA. 104, 10720–10725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian, D. , Wang, J. , Zeng, X. , Gu, K. , Qiu, C. , Yang, X. , Zhou, Z. , Goh, M. , Luo, Y. , Murata‐Hori, M. , White, F. and Yin, Z. (2014) The rice TAL effector‐dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell, 26, 497–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran, T.T. , Doucouré, H. , Hutin, M. , Jaimes Niño, L.M. , Szurek, B. , Cunnac, S. and Koebnik, R. (2018a) Efficient enrichment cloning of TAL effector genes from Xanthomonas . MethodsX, 5, 1027–1032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran, T.T. , Perez‐Quintero, A.L. , Wonni, I. , Carpenter, S.C.D. , Yu, Y. , Wang, L. , Leach, J.E. , Verdier, V. , Cunnac, S. , Bogdanove, A.J. , Koebnik, R. , Hutin, M. and Szurek, B. (2018b) Functional analysis of African Xanthomonas oryzae pv. oryzae TALomes reveals a new susceptibility gene in bacterial leaf blight of rice. PLoS Pathog. 14, e1007092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, C. , Zhang, X. , Fan, Y. , Gao, Y. , Zhu, Q. , Zheng, C. , Qin, T. , Li, Y. , Che, J. , Zhang, M. , Yang, B. , Liu, Y. and Zhao, K. (2015) XA23 is an executor R protein and confers broad‐spectrum disease resistance in rice. Mol. Plant, 8, 290–302. [DOI] [PubMed] [Google Scholar]
- White, F. (2016) Xanthomonas and the TAL effectors: Nature's molecular biologist. Methods Mol Biol. 1338, 1–8. [DOI] [PubMed] [Google Scholar]
- Yang, B. and White, F.F. (2004) Diverse members of the AvrBs3/PthA family of type III effectors are major virulence determinants in bacterial blight disease of rice. Mol. Plant‐Microbe Interact. 17, 1192–1200. [DOI] [PubMed] [Google Scholar]
- Yang, B. , Sugio, A. and White, F. (2006) Os8N3 is a host disease‐susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA. 103, 10503–10508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, Y. , Streubel, J. , Balzergue, S. , Champion, A. , Boch, J. , Koebnik, R. , Feng, J. , Verdier, V. and Szurek, B. (2011) Colonization of rice leaf blades by an African strain of Xanthomonas oryzae pv. oryzae depends on a new TAL effector that induces the rice nodulin‐3 Os11N3 Gene. Mol. Plant‐Microbe Interact. 24, 1102–1113. [DOI] [PubMed] [Google Scholar]
- Yu, Y. , Lu, Y. , He, Y. , Huang, S. and Tang, J. (2015) Rapid and efficient genome‐wide characterization of Xanthomonas TAL effector genes. Sci. Rep. 5, 13162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, J. , Yin, Z. and White, F. (2015) TAL effectors and the executor R genes. Front. Plant Sci. 6, 641. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1 DNA sequences of gBlock fragments synthesized to make two Gibson cloning vectors. (A) gBlock1 was used to inserted into pZW‐pthXo1 at SphI sites through Gibson cloning. The two sequences shaded in yellow are homologous to the ends of SphI fragments of TALe genes. (B) gBlock2 was used to make pHM1‐Gib. The two sequences shaded in yellow are homologous to the ends of BamHI fragments of TALe genes.
Fig. S2 Nine TALe genes from AXO1947 were cloned using the pZW‐Gib vector and Gibson assembly method. The RVDs of individual TALes are shown under numbers 1 to 26, indicating the order of 33–34 amino acid repeats. Asterisks (*) indicate that the amino acid at the 13th position missing.
Fig. S3 Virulence contribution of nine TALes cloned from AXO1947. (A) Lesion lengths caused in rice Kitaake and Zhenshan 97 (ZS 97) by different Xoo strains indicated below the paired columns. Different lower letters indicate statistically significant differences (mean SEM, n = 10, P < 0.05). (B) Blight symptom in Kitaake leaves caused by the Xoo strains as indicated below each leaf. Arrows indicate the edges of lesions.
Fig. S4 TALe genes from PXO61 and AXO1947. (A) TALe genes clustered in the genome of PXO61. (B) TALe genes clustered in the genome of AXO1947. (C, D) Lesion length measurements (mean SEM, n = 10) in Kitaake caused by different strains.
Fig. S5 Validation of TALe clones through PCR, restriction enzyme digestions. (A) Schematics of selective isolation of BamHI fragment TALe genes from genomic DNA of CFBP7321. (B) Validation of TALe clones through PCR with primers P‐F and P‐R of individual clones as indicated above lanes of upper gel image, digestion by MscI which cuts each of central repeats (the DNA band patterns resulted from partial digestion) and digestion by BamHI.
Fig. S6 Validation of TALe clones through PCR, restriction enzyme digestions. (A) Schematics of selective isolation of BamHI fragments TALe genes from genomic DNA of CFBP7325. (B) Validation of TALe clones through PCR with primers P‐F1 and P‐R1 of individual clones as indicated above lanes of upper gel image, digestion by MscI which cuts each of central repeats (the DNA band patterns resulted from partial digestion) and digestion by BamHI.
Fig. S7 TALe gene distribution in two African Xoo genomes. (A, B) Nine TALe genes in each of two Xoo genomes are syntenic in their locations with the same colors denoting identical TALes at the amino acid level while different colors indicate different TALes at the amino acid level.
Fig. S8 Two TALe genes cloned with the pHM1‐Gib system were functional in virulence. The virulences of TalC from CFBP7321 and TalF from CFBP7325 were tested in Kitaake leaves. Different letters indicate statistically significant differences.
Table S1 Number of TALe genes from different Xanthomonas that can be selectively cloned with Gibson assembly.
Table S2 Plasmids and bacterial strains used in this study.
Table S3 Primers used in this study.