

Abstract
There is growing evidence that stressors occurring during pregnancy can impair biological and behavioral responses to stress in the adult offspring. For instance, prenatal stress enhances emotional reactivity, anxiety, and depressive-like behaviors associated with a prolonged stress-induced corticosterone secretion and a reduction in hippocampal corticosteroid receptors. Among the neurotransmitters involved in these hormonal and behavioral responses, acetylcholine may play a critical role. However, it is unknown whether prenatal stressful events also may influence the development of cholinergic systems. In the present study, hippocampal acetylcholine was measured, by in vivomicrodialysis, in both male and female adult prenatally stressed rats, under basal conditions, after a mild stress (saline injection) or after intracerebroventricular administration of corticotropin-releasing factor (CRF; 0.1 nm). No difference in basal release of acetylcholine was observed between control and prenatally stressed rats of both genders. Mild stress was found to increase hippocampal acetylcholine release to a greater extent in prenatally stressed rats than in controls. In males, the CRF-induced increase in hippocampal acetylcholine release was larger in prenatally stressed rats, as compared with controls, during the first hour after the injection and in females during the third hour after the injection. These data indicate that prenatal stress has long-term effects on the development of forebrain cholinergic systems. The augmented increase in hippocampal acetylcholine release after the mild stress and CRF injection in prenatally stressed rats may be involved in some of the hormonal and behavioral abnormalities found in prenatally stressed rats.
Keywords: prenatal stress, development, acetylcholine, ovine corticotropin-releasing factor, gender, hippocampus
Prenatal environment can influence an individual’s development profoundly, inducing changes lasting into adulthood (Weinstock, 1997). In humans, for example, the offspring of mothers experiencing stress during pregnancy have been reported to display long-term behavioral abnormalities (Stott, 1973; Shell, 1981;Meijer, 1985). However, because of limitations inherent to research on humans, this phenomenon has been examined most extensively by using an animal model of prenatally stressed (PS) rats. Among the behavioral attributes of adult PS rats, increased “emotionality” (Thompson, 1957; Fride et al., 1986; Wakshlak and Weinstock, 1990), “defensive behavior” (Takahashi et al., 1992), and “anxiety” (Weinstock et al., 1988; Vallée et al., 1997) have been shown. On the other hand, associated with those behavioral changes, prenatal stress can induce long-term changes in various neurobiological systems, including the hypothalamo–pituitary–adrenal (HPA) axis, mediating an animal’s hormonal response to stress. Indeed, increased basal and stress-induced plasma concentrations of adrenocorticotropin (ACTH) (McCormick et al., 1995), prolongation of stress-induced corticosterone secretion (Weinstock et al., 1992; Maccari et al., 1995; McCormick et al., 1995), and decreased binding capacity of hippocampal corticosteroid receptors (Maccari et al., 1995) have been reported in adult PS rats.
Prenatal stress also has been shown to affect various neurotransmitters, including serotonin (Peters, 1986, 1989, 1990) and catecholamines (Fride and Weinstock, 1989; Takahashi et al., 1992;Alonso et al., 1994). Another neurotransmitter system possibly involved in the mediation of prenatal stress-induced abnormalities is the septohippocampal cholinergic system. Given that several measures of activity in these neurons, including acetylcholine (ACh) release in the hippocampus, are increased by stress (Gilad, 1987; Imperato et al., 1991; Mark et al., 1996) and that cholinergic tone may be involved in emotional affect (Janowsky et al., 1994), it could be hypothesized that prenatal stress-induced changes in this neurotransmitter system may underlie some of the behavioral and neuroendocrine abnormalities of PS rats, as outlined above.
Another central neurotransmitter deserving further characterization with regard to prenatal stress-induced changes is corticotropin-releasing factor (CRF); whereas the median eminence CRF content of adult PS rats is unchanged, as compared with control animals (Smythe et al., 1996), prenatal stress has been reported to increase CRF content in amygdala (Cratty et al., 1995), a structure known to modulate emotional responses to stress (Gallagher and Chiba, 1996). Furthermore, CRF is known to act centrally to mediate stress-related behaviors (Menzaghi et al., 1993), which are altered by prenatal stress. Interestingly, we have observed recently that intracerebroventricular administration of CRF stimulates hippocampally projecting cholinergic neurons in a dose-dependent manner (our unpublished observations), suggesting that changes in the interactions between those two neurotransmitter systems might be involved in the behavioral characteristics of PS rats.
Experiments thus were undertaken to determine whether the CRF/ACh interaction indeed is altered by prenatal stress. To this end, the effects of intracerebroventricular (ICV) administration of CRF were assessed on hippocampal ACh release, using in vivomicrodialysis in control and PS rats of both genders. In addition, the effects of a mild stressor (an ICV injection of saline) on hippocampal ACh release were measured in those rats.
MATERIALS AND METHODS
Prenatal stress procedure
Adult virgin Sprague Dawley female rats (Iffa Credo, Lyon, France) weighing 240 gm were group-housed (10 per cage) for 10 d to coordinate their estrous cycle and then individually housed for a whole estrous cycle (4 d) in the presence of a sexually experienced male Sprague Dawley rat weighing 400 gm. Pregnant rats then were assigned randomly to prenatal stress or control groups, individually housed in plastic breeding cages, allowed ad libitum access to food and water, and maintained on a constant 12:12 light/dark cycle (lights on: 8:00 A.M.–8:00 P.M.) at constant room temperature (23°C) and humidity (60%). Stress was performed each day of the last week of pregnancy until delivery; pregnant females were restrained individually in plastic transparent cylinders (7 cm in diameter and 19 cm long) and exposed to bright light for 45 min, three times a day, at 9:00 A.M., 12:00 P.M., and 5:00 P.M. Control pregnant females were left undisturbed in their home cages. Male and female offspring were weaned 21 d after birth and housed in same-sex groups of four until the experiments started at 3 months of age. Only nine litters of 8–13 pups with similar numbers of males and females were kept for the study, all other litters having been eliminated to rule out extra stressors, such as removal of the pups. A maximum of two male and two female pups was used from each litter to remove any “litter effects” (Becker and Kowall, 1977; Chapman and Stern, 1979).
Experimental design
Transverse microdialysis probes were implanted into the dorsal hippocampi of prenatally stressed or control adult (male and female) rats. Two days later, these rats underwent intracerebral microdialysis perfusion and were injected ICV with saline (0.9%), considered as a mild stressor, and 60 min later with 0.5 μg/rat (0.1 nmol) of ovine CRF (oCRF; Sigma, Deisenhofen, Germany). oCRF was selected on the basis of its higher efficiency in stimulating hippocampal ACh release as compared to rat/human peptide (our unpublished observations).
Probe/guide cannula implantation surgery. Transverse dialysis probes were implanted stereotaxically into the dorsal hippocampi of anesthetized (pentobarbital 50–60 mg/kg, i.p.) rats, using the coordinates anterior (A) −3.9 and ventral (V) −3.3 mm measured from bregma, with the incisor bar set at −3.3 mm to imitate the location depicted in the Paxinos and Watson atlas (1986) as A −4.3 and V −3.3. The dialysis probes were made of acrylonitrile–sodium methallyl sulfonate fiber (inner diameter, 220 μm; outer diameter, 310 μm; molecular weight cutoff >60,000 Dalton; Filtral AN69, Vancouver Hospital Canada) and had an active surface length of 6.8 mm. In addition, a 10-mm-long 23 gauge guide cannula was implanted into the cortex above the lateral ventricle [A at 0, lateral (L) at +1.5 mm measured from bregma, and V at −2.5 mm measured from dura, with the incisor bar set at +5.0 mm] such that the 30 gauge injection cannula used during the experiments, which extended 1.50 mm beyond the guide, would cross the corpus callosum to reach the ventricle. The probe inlet and outlet were closed with caps, and the guide cannula was blocked with a stylette. On completion of each experiment, the ability of a liquid to flow by gravity from the injection cannula into the ventricle was tested; then the animals were killed, and the ICV guide cannula location was verified further, along with that of the probe, using standard histological procedures.
ICV injection of saline (mild stressor). After 1 hr of ACh collection under basal conditions, a 30 gauge stainless steel injection cannula was inserted into the guide in a freely moving rat, and 0.5 μl of saline was injected at 0.5 μl/min by a 50 μl Hamilton syringe driven by a syringe pump (BAS Bee). The saline was separated by a small bubble from the distilled water that filled the rest of the tubing. The cannula was left in place for 1 min after the end of the injection and thereafter was replaced by the stylette. Samples were collected every 10 min for 1 hr.
ICV injection of CRF. The frozen CRF stock solution (1 μg/μl) was thawed <30 min before administration, kept on ice, and drawn into an injection cannula built in the same way as that described for the saline injection. One hour after saline injection, 0.5 μl of the CRF solution was injected at 0.5 μl/min by a 50 μl Hamilton syringe driven by a syringe pump (BAS Bee). The cannula was left in place for 1 min after the end of the injection and thereafter was replaced by the stylette. Samples were collected every 10 min for 4 hr.
Microdialysis. Brain microdialysis was performed as described previously (Damsma and Westerink, 1991; Day and Fibiger, 1994). Rats were housed in a Plexiglas cage (31 × 32 × 35 cm) to which they had been habituated overnight, with free access to food and water. The dialysis probe was perfused at 5 μl/min, controlled by a syringe pump (BAS Bee). The syringe was connected to the probe inlet by polyethylene tubing (a length corresponding to 50 μl) as was the probe outlet connected to the sample loop (50 μl) of the analytical system. The sample valve (Rheodyne, Cotati, CA) was controlled by the internal programmable electronics of the analytical system (Antec, Leyden, The Netherlands; see below), and samples (50 μl) were collected and injected at 10 min intervals. The perfusion solution was an artificial CSF and contained (in mm) 125 NaCl, 3 KCl, 1.3 CaCl2, 1.0 MgCl2, and 23 NaHCO in aqueous phosphate buffer (1 mm, pH 7.4). To recover detectable dialysate concentrations of ACh, we included a reversible acetylcholinesterase inhibitor (neostigmine bromide, 0.1 μm; Sigma) in the perfusion solution. Thirty minutes of perfusion preceded the first sample collection to allow for equilibration of the brain with the perfusion solution.
Acetylcholine assay. ACh was assayed by HPLC with electrochemical detection in conjunction with an enzyme reactor (Damsma et al., 1987; Damsma and Westerink, 1991). ACh and choline were separated on a reverse-phase column (75 × 2.1 mm) pretreated with lauryl sulfate. The eluate from this analytical column then passed through an enzyme reactor (10 × 2.1 mm) containing acetylcholinesterase (EC 3.1.1.7; type VI-S, Sigma) and choline oxidase (1.1.3.17; Sigma) covalently bound to glutaraldehyde-activated Lichrosorb NH2 (10 μm; Merck, Darmstadt, Germany). The separated ACh and choline reacted to give a stoichiometric yield of hydrogen peroxide, which was detected electrochemically at a platinum electrode at a potential of +500 mV versus an Ag/AgCl reference electrode (Antec). In the mobile phase, 0.2 m aqueous potassium phosphate buffer pH 8.0, containing 1 mmtetramethylammonium hydroxide, was delivered by a pump (Shimadzu LC-10AD) at 0.35–0.45 ml/min. The best detection limit of the assay is ∼10 fmol/injection and averaged 50 fmol/injection during the experiments. The time required to complete a chromatogram was 4–5 min.
Statistical analyses. Biochemical data are presented first as averages of raw uncorrected dialysate ACh concentrations (in fmol/min). For statistical analysis of the effects of saline or CRF injections, data are calculated as absolute changes (in Δ fmol/min) from each animal’s average baseline concentration, this baseline being defined as the average of the last four values preceding the corresponding injection.
ANOVA with repeated measures were used to test for differences between groups (control/prenatally stressed) in both basal and CRF-induced ACh concentrations. ANOVA yielding significant differences were subjected to Dunnett post hoc analysis. The dialysate concentrations of ACh in response to the saline injection were analyzed for group differences by Student’s t test, using the first sample after injection.
RESULTS
A total of 24 rats in four different groups (control males,n = 6; PS males, n = 7; control females, n = 5; PS females, n = 6) were dialyzed. The effects of saline and CRF injections on ACh output (fmol/min) in males and females are depicted in Figure1, a and b, respectively.
Fig. 1.
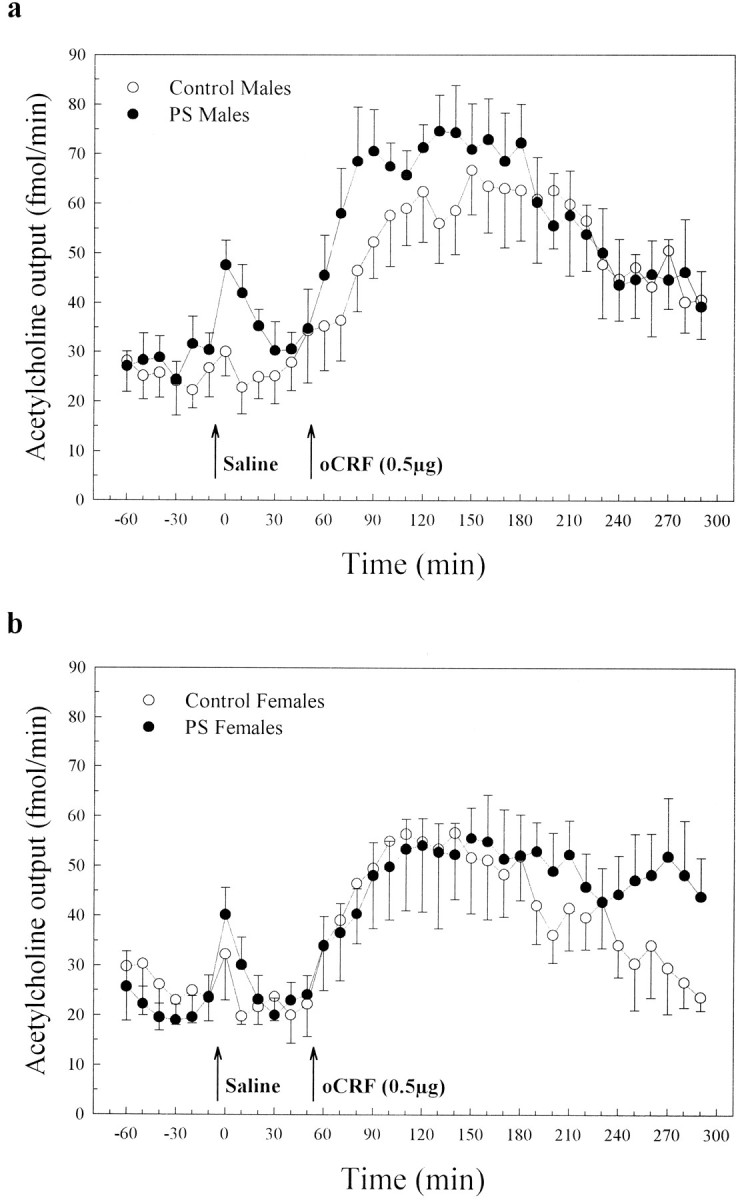
Effects of prenatal stress on basal, saline-stimulated, and CRF-stimulated hippocampal ACh output (fmol/min) in male (a) and female (b) adult rats. Each data point represents the group mean ± SEM of the dialysate ACh concentration in 10 min samples. The arrows indicate the times of saline and CRF injections.
Basal hippocampal ACh release in males
Prenatal stress did not modify basal concentrations of ACh, either before saline injection (group effect, F(1,11) = 0.471, p = 0.506; group × time interaction,F(3,33) = 0.918, p = 0.443) or before CRF injection (group effect, F(1,11) = 0.419, p = 0.539; group × time interaction,F(3,33) = 0.542, p = 0.657) in males (Fig. 1a). The mean ACh output of all rats was 26.9 ± 1.6 fmol/min before saline injection and 30.52 ± 2.1 fmol/min before CRF injection. Pre-CRF ACh concentrations had a tendency to be higher than pre-saline ACh concentrations in both control and PS rats (F(1,11) = 4.14,p = 0.066), this difference being similar in both groups (group × time interaction, F(1,11)= 0.026, p = 0.875).
Basal hippocampal ACh release in females
As in male rats, prenatal stress did not affect basal ACh concentrations, either before saline injection (group effect,F(1,9) = 0.373, p = 0.556; group × time interaction, F(3,27) = 0.568,p = 0.640) or before CRF injection (group effect,F(1,9) = 0.013, p = 0.911; group × time interaction, F(3,27) = 0.679,p = 0.572) in females (Fig. 1b). The mean ACh output of all rats was 22.20 ± 1.7 fmol/min before saline injection and 22.26 ± 1.4 fmol/min before CRF injection. In contrast to males, there was no tendency for basal ACh concentrations before CRF injection to differ from those preceding saline injection (F(1,9) = 0.02, p = 0.889).
Hippocampal ACh release after a mild stress (e.g., saline injection)
Effects of ICV saline injection on hippocampal dialysate concentrations of ACh (Δ fmol/min) in male and female adult offspring are shown in Figure 2, a andb. Saline injection produced a greater transient increase in hippocampal ACh release in PS males than in control males (t = −3.57, p = 0.004; Fig.2a). A nonsignificant similar tendency was observed in PS females as compared with their controls (t = −2.16,p = 0.059; Fig. 2b). Only PS rats responded significantly to this mild stress.
Fig. 2.
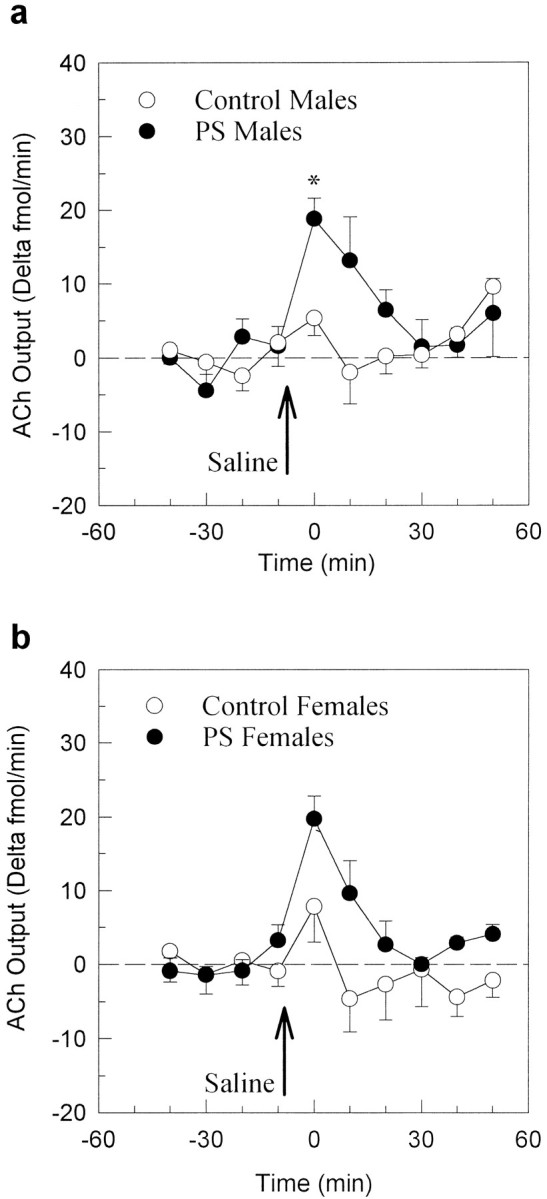
Effects of prenatal stress on saline-stimulated hippocampal ACh output (Δ fmol/min) in males (a) and females (b). Prenatal manipulation enhanced the transient increase of ACh release both in males (*p = 0.004) and, to a lower extent, in females (p = 0.059).
Hippocampal ACh release after CRF injection
ICV CRF increased hippocampal ACh release in all groups of rats, as shown in Figure 3 for males and in Figure 4 for females, to a much greater extent than did the saline injection. However, this increase was modified by prenatal stress in both males (group × time interaction, F(23,253) = 2.091,p = 0.003) and females (group × time interaction,F(23,207) = 1.71, p = 0.026) during the 4 hr after CRF injection. In males, prenatal stress increased CRF-induced ACh release over the first hour after the injection (Fig. 3inset; F(1,11) = 5.61, p = 0.037), whereas in females the increase of the CRF-induced ACh release was seen over the third hour after CRF injection (Fig. 4inset; F(1,9) = 6.37, p = 0.032).
Fig. 3.

Effects of prenatal stress on CRF-induced hippocampal ACh output (Δ fmol/min) in males. Prenatally stressed rats showed a higher ACh response to CRF than control rats over the first hour (inset; *p < 0.05). This difference was attributable to a higher ACh release 10, 20, 30, and 70 min after CRF injection (Dunnett post hoc analysis; *p < 0.05).
Fig. 4.

Effects of prenatal stress on CRF-induced hippocampal ACh output (Δ fmol/min) in females. Prenatally stressed rats showed a higher ACh response to CRF than control rats over the third hour (inset; *p < 0.05). This difference was attributable to higher ACh release 190, 210, 220, and 230 min after CRF injection (Dunnett post hoc analysis; *p < 0.05).
No significant differences in basal values, in saline-induced, or in CRF-induced hippocampal ACh release were found between male and female control groups (comparisons not shown). CRF treatment also was noted to increase locomotion and grooming behavior in all rats (data not shown).
DISCUSSION
Results of these experiments demonstrate that prenatal stress increases hippocampal acetylcholine release induced by a mild stressor (saline injection). Furthermore, the CRF-induced increase in hippocampal ACh release was larger in PS rats, as compared with controls, during the first hour after CRF injection in males and during the third hour after injection in females. However, no changes in basal ACh release were observed between the groups. The lack of statistically significant difference in basal ACh release between the treatment groups is quite common in microdialysis experiments, mainly because of large interindividual differences in this measure, and does not exclude the possibility that prenatal stress may induce changes in the cholinergic system, which could appear, using other measures, in resting conditions.
The increased hippocampal ACh release in response to mild stress in PS groups is in agreement with previous experiments showing augmented stress-induced effects in PS rats, using both neuroendocrine (Maccari et al., 1995; Weinstock, 1997) and behavioral measures (Fride et al., 1986; Vallée et al., 1997). Furthermore, the changes in the septohippocampal cholinergic system also could mediate the abnormalities in the activity of the HPA axis seen in PS rats, given that these cholinergic neurons may regulate the hippocampal glucocorticoid receptors (Yau et al., 1992; Alema et al., 1995) that are known to be involved in feedback inhibition of corticosterone secretion (McEwen et al., 1986; De Kloet and Reul, 1987). Although prenatal stress already has been shown to induce changes in other neurotransmitters, such as noradrenaline (Peters, 1982; Takahashi et al., 1992), dopamine (Alonso et al., 1994; Henry et al., 1995), and serotonin (Peters, 1986, 1989, 1990), this is the first report of an altered cholinergic functioning in PS rats. Given that CRF, the release of which occurs during various stressful events (Hauger et al., 1988;Owens and Nemeroff, 1991), has been shown to enhance hippocampal ACh release (our unpublished observations) and that prenatal stress increases central CRF content (Cratty et al., 1995), the increased release of ACh in PS rats reported here in response to the stress of a saline injection could result from a greater release or activity of CRF in these animals.
Prenatal stress augments the CRF-induced increase in hippocampal acetylcholine release, suggesting that the regulation of the hippocampal cholinergic system regulation by central CRF is sensitive to prenatal manipulations. We have shown previously that the CRF stimulation of hippocampal ACh release is independent of the CRF-induced corticosterone secretion and thus is centrally mediated. Indeed, subcutaneous injections of CRF increased plasma concentrations of corticosterone to the same levels as did the central injections, without affecting hippocampal ACh release (our unpublished observations). Taken together with reports of prenatal stress-induced increases in the content and release of CRF centrally (i.e., in the amygdala; Cratty et al., 1995), the observation that CRF-induced ACh release is independent of corticosterone secretion could suggest that prenatal stress may affect the central neurotransmitter role of CRF. Prenatal stress-induced changes could occur, for example, in the concentration or binding capacities of the CRF-binding protein or the CRF receptor(s), as well as in the second messenger systems that mediate this cholinergic effect and that as yet are undefined. This finding also suggests that the stress-induced release of endogenous CRF could affect hippocampal ACh release differently in PS rats than in control rats. Indeed, this theory is supported by the first finding concerning the response to saline injection that was discussed above.
Alteration in CRF-induced hippocampal ACh release is different in the male and female PS offspring. Indeed, the difference in the male rats represents a more rapid increase of hippocampal ACh release in the PS group to the same peak level as the controls. This anticipation of the peak response could be attributed to the augmented ACh release in the PS group after the mild stress of the injection itself (see Fig.2a), but it also may involve changes in the disposition or kinetics of CRF after injection, perhaps implicating modifications of the CRF-binding protein. The effect seen in the females is attributed to a prolonged effect of CRF in the PS group that is present over the third hour after injection. This effect may indicate abnormalities in the mechanisms responsible for returning the systems that are involved to their basal state. Sex differences in the effects of prenatal stress are, indeed, well known in the literature, the female offspring often showing larger changes in adulthood than male offspring both in neuroendocrine (Kinsley et al., 1989; Weinstock et al., 1992; McCormick et al., 1995) and behavioral measures (Fride and Weinstock, 1989; J.Alonso et al., 1991; S. Alonso, 1991).
An interesting implication of the prenatal stress-induced changes in CRF stimulation of hippocampally projecting cholinergic neurons demonstrated here relates to the suggestion that prenatal stress may represent an animal model of depression (S. Alonso et al., 1991, 1994). This theory is based on the fact that female PS rats exhibit “behavioral despair” (S. Alonso et al., 1991) in the forced swimming task (Porsolt et al., 1977) that has been proposed as a measure of “depression” and antidepressant efficacy in animal models. In addition, feedback inhibition of HPA axis activity by circulating glucocorticoids is impaired in both depressed patients and PS rats. Elevated amplitudes of cortisol and ACTH secretory episodes, as well as escape from the suppression of cortisol secretion induced by the glucocorticoid receptor agonist dexamethasone, are observed in depressed patients (Arana and Mossman, 1988); PS rats similarly escape from the feedback inhibition responsible for returning corticosteroid secretion to basal levels after a challenge-induced increase (Maccari et al., 1995; Barbazanges et al., 1996; Vallée et al., 1997). Altered circadian rhythms of cortisol/corticosterone secretion also have been reported both in depressed patients (Pfohl et al., 1985) and in PS rats (Koehl et al., 1997). Finally, elevated CRF levels in the CSF fluid (Nemeroff, 1988) and cholinergic hyperactivity have been found in depressed patients (for review, see Janowsky et al., 1994), and in the present report we show a cholinergic hypersensitivity of PS rats to a CRF challenge.
In conclusion, all of these data suggest that prenatal stress has long-term effects on the development of forebrain cholinergic systems. Cholinergic hypersensitivity, together with the well known abnormalities of HPA axis observed in PS rats, suggests that PS may represent an interesting animal model of latent or potential depression.
Footnotes
This study was supported by the Institut National de la Santé et de la Recherche Médicale, the Université de Bordeaux 2, and the Conseil Régional d’Aquitaine. J.C.D. was supported by a Human Frontiers Science Program long-term fellowship. We thank Josette Dulluc for technical assistance and Michela Marinelli and Michel Barrot for helpful comments.
J.C.D. and M.K. have participated equally in these experiments.
Correspondence should be addressed to Dr. Stefania Maccari, Institut National de la Santé et de la Recherche Médicale Unité 259, Université Victor Segalen Bordeaux 2, Domaine de Carreire, Rue Camille Saint Saëns, 33077 Bordeaux Cedex, France.
Dr. Day’s present address: Developmental Neuroendocrinology Laboratory, Lehmann Pavillion, Douglas Hospital Research Centre, 6875 Boulevard Lasalle, Verdun, Québec, H4H 1R3 Canada.
REFERENCES
- 1.Alema GS, Casolini P, Patacchioli FR, Angelucci L. Rat brain corticosteroid receptors are modulated by septo-hippocampal cholinergic innervation. NeuroReport. 1995;6:2461–2464. doi: 10.1097/00001756-199512150-00006. [DOI] [PubMed] [Google Scholar]
- 2.Alonso JA, Castellano MA, Rodriguez M. Behavioral lateralization in rats: prenatal stress effects on sex differences. Brain Res. 1991;539:45–50. doi: 10.1016/0006-8993(91)90684-n. [DOI] [PubMed] [Google Scholar]
- 3.Alonso SJ, Arevalo R, Afonso D, Rodriguez M. Effects of maternal stress during pregnancy on forced swimming test behavior of the offspring. Physiol Behav. 1991;50:511–517. doi: 10.1016/0031-9384(91)90538-y. [DOI] [PubMed] [Google Scholar]
- 4.Alonso SJ, Navarro E, Rodriguez M. Permanent dopaminergic alterations in the n. accumbens after prenatal stress. Pharmacol Biochem Behav. 1994;49:353–358. doi: 10.1016/0091-3057(94)90433-2. [DOI] [PubMed] [Google Scholar]
- 5.Arana GW, Mossman D. The dexamethasone suppression test and depression. Endocrinol Metab Clin North Am. 1988;17:21–39. [PubMed] [Google Scholar]
- 6.Barbazanges A, Piazza PV, Le Moal M, Maccari S. Maternal glucocorticoid secretion mediates long-term effects of prenatal stress. J Neurosci. 1996;16:3943–3949. doi: 10.1523/JNEUROSCI.16-12-03943.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Becker G, Kowall M. Crucial role of the postnatal maternal environment in the expression of prenatal stress effects in the male rats. J Comp Physiol Psychol. 1977;91:1432–1446. doi: 10.1037/h0077401. [DOI] [PubMed] [Google Scholar]
- 8.Chapman R, Stern J. Failure of severe maternal stress or ACTH during pregnancy to affect emotionality of male rat offspring: implications of litter effects for prenatal studies. Dev Psychobiol. 1979;12:255–269. doi: 10.1002/dev.420120309. [DOI] [PubMed] [Google Scholar]
- 9.Cratty MS, Ward HE, Johnson EA, Azzaro AJ, Birkle DL. Prenatal stress increases corticotropin-releasing factor (CRF) content and release in rat amygdala minces. Brain Res. 1995;675:297–302. doi: 10.1016/0006-8993(95)00087-7. [DOI] [PubMed] [Google Scholar]
- 10.Damsma G, Westerink BHC. A microdialysis and automated on-line analysis approach to study central cholinergic transmission in vivo. In: Robinson TE, Justice J, editors. Microdialysis in the neurosciences. Elsevier; Amsterdam: 1991. pp. 237–252. [Google Scholar]
- 11.Damsma G, Lammerts van Bueren D, Westerink BHC, Horn AS. Determination of acetylcholine and choline in the femtomole range by means of HPLC, a postcolumn enzyme reactor, and electrochemical detection. Chromatographia. 1987;24:827–831. [Google Scholar]
- 12.Day JC, Fibiger HC. Dopaminergic regulation of septohippocampal cholinergic neurons. J Neurochem. 1996;63:2086–2092. doi: 10.1046/j.1471-4159.1994.63062086.x. [DOI] [PubMed] [Google Scholar]
- 13.De Kloet ER, Reul JMHM. Feedback action and tonic influence of corticosteroids on brain function: a concept arising from the heterogeneity of brain receptor systems. Psychoneuroendocrinology. 1987;12:83–105. doi: 10.1016/0306-4530(87)90040-0. [DOI] [PubMed] [Google Scholar]
- 14.Fride E, Weinstock M. Alterations in behavioral and striatal dopamine asymmetries induced by prenatal stress. Pharmacol Biochem Behav. 1989;32:425–430. doi: 10.1016/0091-3057(89)90174-3. [DOI] [PubMed] [Google Scholar]
- 15.Fride E, Dan Y, Feldon J, Halevy G, Weinstock M. Effects of prenatal stress on vulnerability to stress in prepubertal and adult rats. Physiol Behav. 1986;37:681–687. doi: 10.1016/0031-9384(86)90172-1. [DOI] [PubMed] [Google Scholar]
- 16.Gallagher M, Chiba AA. The amygdala and emotion. Curr Opin Neurobiol. 1996;6:221–227. doi: 10.1016/s0959-4388(96)80076-6. [DOI] [PubMed] [Google Scholar]
- 17.Gilad G. The stress-induced response of the septo-hippocampal cholinergic system. A vectorial outcome of psychoneuroendocrinological interactions. Psychoneuroendocrinology. 1987;12:167–184. doi: 10.1016/0306-4530(87)90002-3. [DOI] [PubMed] [Google Scholar]
- 18.Hauger RL, Millan MA, Lorang M, Harwood JP, Aguilera G. Corticotropin-releasing factor receptors and pituitary adrenal responses during immobilization stress. Endocrinology. 1988;123:396–405. doi: 10.1210/endo-123-1-396. [DOI] [PubMed] [Google Scholar]
- 19.Henry C, Guegant G, Cador M, Arnauld E, Arsaut J, Le Moal M, Demotes-Mainard J. Prenatal stress in rats facilitates amphetamine-induced sensitization and induces long-lasting changes in dopamine receptors in the nucleus accumbens. Brain Res. 1995;685:179–186. doi: 10.1016/0006-8993(95)00430-x. [DOI] [PubMed] [Google Scholar]
- 20.Imperato A, Puglisi-Allegra S, Casolini P, Angelucci L. Changes in brain dopamine and acetylcholine release during and following stress are independent of the pituitary–adrenocortical axis. Brain Res. 1991;538:111–117. doi: 10.1016/0006-8993(91)90384-8. [DOI] [PubMed] [Google Scholar]
- 21.Janowsky DS, Overstreet DH, Nurnberger JI., Jr Is cholinergic sensitivity a genetic marker for the affective disorders? Am J Med Genet. 1994;54:335–344. doi: 10.1002/ajmg.1320540412. [DOI] [PubMed] [Google Scholar]
- 22.Kinsley CH, Mann PE, Bridges RS. Alterations in stress-induced prolactin release in adult female and male rats exposed to stress, in utero. Physiol Behav. 1989;45:1073–1076. doi: 10.1016/0031-9384(89)90240-0. [DOI] [PubMed] [Google Scholar]
- 23.Koehl M, Barbazanges A, Le Moal M, Maccari S. Prenatal stress induces a phase advance of circadian corticosterone rhythm in adult rats which is prevented by postnatal stress. Brain Res. 1997;759:317–320. doi: 10.1016/s0006-8993(97)00394-6. [DOI] [PubMed] [Google Scholar]
- 24.Maccari S, Piazza PV, Kabbaj M, Barbazanges A, Simon H, Le Moal M. Adoption reverses the long-term impairment in glucocorticoid feedback induced by prenatal stress. J Neurosci. 1995;15:110–116. doi: 10.1523/JNEUROSCI.15-01-00110.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mark GP, Rada PV, Shors TJ. Inescapable stress enhances extracellular acetylcholine in the rat hippocampus and prefrontal cortex but not the nucleus accumbens or amygdala. Neuroscience. 1996;74:767–774. doi: 10.1016/0306-4522(96)00211-4. [DOI] [PubMed] [Google Scholar]
- 26.McCormick CM, Smythe JW, Sharma S, Meaney MJ. Sex-specific effects of prenatal stress on hypothalamic-pituitary-adrenal responses to stress and brain glucocorticoid receptor density in adult rats. Dev Brain Res. 1995;84:55–61. doi: 10.1016/0165-3806(94)00153-q. [DOI] [PubMed] [Google Scholar]
- 27.McEwen BS, De Kloet ER, Rostene W. Adrenal steroid receptors and action in the nervous system. Physiol Rev. 1986;66:1121–1188. doi: 10.1152/physrev.1986.66.4.1121. [DOI] [PubMed] [Google Scholar]
- 28.Meijer A. Child psychiatric sequelae of maternal war stress. Acta Psychiatr Scand. 1985;72:505–511. doi: 10.1111/j.1600-0447.1985.tb02647.x. [DOI] [PubMed] [Google Scholar]
- 29.Menzaghi F, Heinrichs SC, Pich EM, Weiss F, Koob GF. The role of limbic and hypothalamic corticotropin-releasing factor in behavioral responses to stress. Ann NY Acad Sci. 1993;697:142–154. doi: 10.1111/j.1749-6632.1993.tb49929.x. [DOI] [PubMed] [Google Scholar]
- 30.Nemeroff CB. The role of corticotropin-releasing factor in the pathogenesis of major depression. Pharmacopsychiatry. 1988;21:76–82. doi: 10.1055/s-2007-1014652. [DOI] [PubMed] [Google Scholar]
- 31.Owens MJ, Nemeroff CB. Physiology and pharmacology of corticotropin releasing factor. Pharmacol Rev. 1991;43:425–473. [PubMed] [Google Scholar]
- 32.Paxinos G, Watson C. The rat brain in stereotaxic coordinates, 2nd Ed. Academic; London: 1986. [DOI] [PubMed] [Google Scholar]
- 33.Peters DAV. Prenatal stress: effects on brain biogenic amine and plasma corticosterone levels. Pharmacol Biochem Behav. 1982;17:721–725. doi: 10.1016/0091-3057(82)90353-7. [DOI] [PubMed] [Google Scholar]
- 34.Peters DAV. Prenatal stress increases the behavioral response to serotonin agonists and alters open field behavior in the rat. Pharmacol Biochem Behav. 1986;25:873–877. doi: 10.1016/0091-3057(86)90400-4. [DOI] [PubMed] [Google Scholar]
- 35.Peters DAV. Effects of maternal stress during different gestational periods on the serotonergic system in adult rat offspring. Pharmacol Biochem Behav. 1989;31:839–843. doi: 10.1016/0091-3057(88)90393-0. [DOI] [PubMed] [Google Scholar]
- 36.Peters DAV. Maternal stress increases fetal brain and neonatal cerebral cortex 5-hydroxytryptamine synthesis in rats: a possible mechanism by which stress influences brain development. Pharmacol Biochem Behav. 1990;35:943–947. doi: 10.1016/0091-3057(90)90383-s. [DOI] [PubMed] [Google Scholar]
- 37.Pfohl B, Sherman B, Schlechte J, Stone R. Pituitary–adrenal axis rhythm disturbances in psychiatric depression. Arch Gen Psychiatry. 1985;42:897–903. doi: 10.1001/archpsyc.1985.01790320069009. [DOI] [PubMed] [Google Scholar]
- 38.Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977;266:730–732. doi: 10.1038/266730a0. [DOI] [PubMed] [Google Scholar]
- 39.Shell LM. Environmental noise and human prenatal growth. Am J Physiol Anthropol. 1981;56:63–70. doi: 10.1002/ajpa.1330560107. [DOI] [PubMed] [Google Scholar]
- 40.Smythe JW, McCormick C, Meaney MJ. Median eminence corticotrophin-releasing hormone content following prenatal stress and neonatal handling. Brain Res Bull. 1996;40:195–199. doi: 10.1016/0361-9230(95)02146-9. [DOI] [PubMed] [Google Scholar]
- 41.Stott DN. Follow-up study from birth of the effects of prenatal stresses. Dev Med Child Neurol. 1973;15:770–787. doi: 10.1111/j.1469-8749.1973.tb04912.x. [DOI] [PubMed] [Google Scholar]
- 42.Takahashi LK, Turner JG, Kalin NH. Prenatal stress alters brain catecholaminergic activity and potentiates stress-induced behavior in adult rats. Brain Res. 1992;574:131–137. doi: 10.1016/0006-8993(92)90809-n. [DOI] [PubMed] [Google Scholar]
- 43.Thompson WR. Influence of prenatal maternal anxiety on emotionality in young rats. Science. 1957;125:698–699. doi: 10.1126/science.125.3250.698. [DOI] [PubMed] [Google Scholar]
- 44.Vallée M, Mayo W, Dellu F, Le Moal M, Simon H, Maccari S. Prenatal stress induces high anxiety and postnatal handling induces low anxiety in adult offspring: correlation with stress-induced corticosterone secretion. J Neurosci. 1997;17:2626–2636. doi: 10.1523/JNEUROSCI.17-07-02626.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wakshlak A, Weinstock M. Neonatal handling reverses behavioral abnormalities induced in rats by prenatal stress. Physiol Behav. 1990;48:289–292. doi: 10.1016/0031-9384(90)90315-u. [DOI] [PubMed] [Google Scholar]
- 46.Weinstock M. Does prenatal stress impair coping and regulation of hypothalamic-pituitary-adrenal axis? Neurosci Biobehav Rev. 1997;21:1–10. doi: 10.1016/s0149-7634(96)00014-0. [DOI] [PubMed] [Google Scholar]
- 47.Weinstock M, Fride E, Hertzberg R. Prenatal stress effects on functional development on the offspring. Prog Brain Res. 1988;73:319–331. doi: 10.1016/S0079-6123(08)60513-0. [DOI] [PubMed] [Google Scholar]
- 48.Weinstock M, Matlina E, Maor GI, Rosen H, McEwen B. Prenatal stress selectively alters the reactivity of the hypothalamic–pituitary adrenal system in the female rats. Brain Res. 1992;595:195–198. doi: 10.1016/0006-8993(92)91049-k. [DOI] [PubMed] [Google Scholar]
- 49.Yau JL, Dow RC, Fink G, Seckl JR. Medial septal cholinergic lesions increase hippocampal mineralocorticoid and glucocorticoid receptor messenger RNA expression. Brain Res. 1992;577:155–160. doi: 10.1016/0006-8993(92)90550-s. [DOI] [PubMed] [Google Scholar]