

Abstract
Peripheral immune stimulation such as that provided by lipopolysaccharide (LPS) has been reported to increase brain levels of IL-1β mRNA, immunoreactivity, and bioactivity. Stressors produce many of the same neural and endocrine responses as those that follow LPS, but the impact of stressors on brain interleukin-1β (IL-1β) has not been systematically explored. An ELISA designed to detect IL-1β was used to measure levels of IL-1β protein in rat brain. Brain IL-1β was explored after exposure to inescapable shock (IS; 100 1.6 mA tail shocks for 5 sec each) and LPS (1 mg/kg) as a positive control. Rats were killed either immediately or 2, 7, 24, or 48 hr after IS. Brains were dissected into hypothalamus, hippocampus, cerebellum, posterior cortex, and nucleus tractus solitarius regions. LPS produced widespread increases in brain IL-1β, but IS did not. Adrenal glucocorticoids are known to suppress IL-1β production in both the periphery and brain. Thus, it was possible that the stressor did provide stimulus input to the brain IL-1β system(s), but that the production of IL-1β protein was suppressed by the rapid and prolonged high levels of glucocorticoids produced by IS. To test this possibility rats were adrenalectomized or given sham surgery, with half of the adrenalectomized rats receiving corticosterone replacement to maintain basal corticosterone levels. IS produced large increases in brain IL-1β protein in the adrenalectomized subjects 2 hr after stress, whether basal corticosterone levels had been maintained. Thus elimination of the stress-induced rise in corticosterone unmasked a robust and widespread increase in brain IL-1β.
Keywords: interleukin-1β, stress, brain, glucocorticoids, protein, rat
Interleukin-1 (IL-1) is a cytokine produced by a number of peripheral cell types and plays a variety of roles in immune and inflammatory responses (for review, see Fantuzzi and Dinarello, 1996; O’Neill, 1997). IL-1α and β, the IL-1 prohormone, IL-1-converting enzyme, the endogenous IL-1 receptor antagonist, and receptors for IL-1 have all been localized in the brain under varying circumstances (Cunningham and De Souza, 1993; Weiss et al., 1994). IL-1β has consistently been detected in CNS after injury to the brain or peripheral immune activation. For example, IL-1 bioactivity (Fontana et al., 1984; Coceani et al., 1988; Quan et al., 1994), immunoreactivity (Van Dam et al., 1992; Hagan et al., 1993;Hillhouse and Mosley, 1993), and mRNA (Ban et al., 1992; Laye et al., 1994; Buttini and Boddeke, 1995) have all been found to be present in brain after peripheral administration of lipopolysaccharide (LPS). However, detection of IL-1 in brain under basal or nonpathological conditions has been inconsistent. IL-1 bioactivity has sometimes been reported to be present (Nieto-Sampedro and Berman, 1987; Coceani et al., 1988; Quan et al., 1996) and sometimes not to be present (Fontana et al., 1984). Similarly, mRNA for IL-1β has been both detected (Bandtlow et al., 1990; Taishi et al., 1997) and not detected (Higgins and Olschowka, 1991; Gatti and Bartfai, 1993). IL-1 immunoreactivity has also been variable (Hagan et al., 1993; Hillhouse and Mosley, 1993).
Despite these inconsistencies, intracerebroventricular administration of IL-1β and the receptor antagonist to IL-1 (IL-1ra) have had quite clear and reliable effects. Central administration of IL-1 produces fever (Kluger, 1991), slow-wave sleep (Kreuger et al., 1984), hyperalgesia (Oka et al., 1993), reduced food and water intake (Plata-Salaman et al., 1988; Kent et al., 1994), reduced social interaction (Bluthe et al., 1996), increased plasma ACTH and glucocorticoids (Weiss et al., 1991), and alterations in peripheral immune parameters (Sundar et al., 1990; Brown et al., 1991). Correspondingly, intracerebroventricular IL-1ra has been reported to blunt or block several behavioral effects of peripheral immune stimulation (Kent et al., 1996). Although these effects of centrally administered IL-1 are robust, it has been unclear whether IL-1 plays a physiological role in the uninjured brain. However, recent data suggest that brain IL-1 is important in the regulation of sleep (Opp et al., 1991; Opp and Krueger, 1994).
Brain IL-1 may also play a role in mediating the neurochemical and behavioral consequences of stressors. The similarity of the neurochemical and behavioral sequelae of exposure to stressors and infectious agents has often been noted (Dunn, 1993; Zalcman et al., 1994), and brain IL-1 may be an important feature of this overlap. Indeed, central administration of IL-1 produces endocrine, neurochemical, and behavioral changes that are similar to those produced by stressors (Shintani et al., 1995a). Moreover, central injection of IL-1ra has been reported to block the brain monoamine and pituitary–adrenal response to immobilization (Shintani et al., 1995b) and the potentiation of fear conditioning and escape learning failure that follows exposure to inescapable tail shock (Maier and Watkins, 1995).
There has been little work directed at determining whether stressors actually alter brain IL-1. Immobilization has been reported to increase IL-1β mRNA (Minami et al., 1991) and bioactivity (Shintani et al., 1995b). However, the generality of these results is unknown, and factors that might determine these brain IL-1 changes have not been studied systematically. Here, we sought to determine whether a different stressor, inescapable tail shock, would increase brain protein levels of IL-1β.
MATERIALS AND METHODS
Subjects. Adult male pathogen-free Harlan Sprague Dawley (Indianapolis, IN) rats (300–350 gm) were individually housed in hanging metal cages at 25°C with a 12 hr light/dark cycle (lights on at 7:00 a.m.). Subjects were acclimated to the colony for 14 d before experimentation began. Standard rat chow and water were freely available. Care and use of the animals were in accordance with protocols approved by the University of Colorado Institutional Animal Care and Use Committee.
Inescapable tail shock treatment. All rats were handled and weighed 2 d before each study began. Animals either remained undisturbed in their home cages as controls (HCC) or were exposed to inescapable tail shock (IS). The stress protocol involved placing the rats in a Plexiglas restraining tube (23.4 cm long and 7 cm in diameter) and exposing them to 100 1.6 mA inescapable shocks for 5 sec each, with an average intertrial interval of 60 sec. The shocks were applied through electrodes taped to the tail. The animals were stressed between 8:00 and 10:00 a.m. and, after stressor termination, were returned to their home cages. Cages of stressed rats were placed on the opposite side of the room (14 × 12 ft) from the HCC animals.
Adrenalectomy. Bilateral adrenalectomies (ADX) were aseptically performed under halothane anesthesia (Halocarbon Laboratories, River Edge, NJ). All removed tissue was examined immediately to ensure complete removal of the adrenal gland. Sham-operated animals received the identical procedure, except that the adrenal glands were gently manipulated with forceps but not removed. Steroid replacement began for ADX animals immediately after surgery. ADX animals received corticosterone (CORT) replacement in their drinking water, because this method has been shown to mimic the normal circadian pattern of CORT secretion (Jacobson et al., 1988). Corticosterone (Sigma, St. Louis, MO) was initially dissolved in ethyl alcohol (ETOH) and diluted to a final concentration of 25 μg/ml in 0.2% ETOH. CORT-water also contained 0.5% saline. Sham animals received drinking water containing 0.2% ETOH. Animals were allowed 6 weeks to recover before any experiments were performed.
LPS treatment. LPS (Escherichia coli, 0111:B4; Sigma) was injected intraperitoneally at doses ranging from 1 μg/kg to 1 mg/kg dissolved in sterile, endotoxin-free 0.9% saline vehicle. Control injections were equivolume (1 ml/kg) vehicle, with all injections given between 7:30 and 8:00 a.m. Animals were returned to their home cage and killed 6–7 hr later. These time intervals were chosen because increases in both mRNA and bioactivity for IL-1 have been reported 2–6 hr after LPS (Fontana et al., 1984;Buttini and Boddeke, 1995; Goujon et al., 1995).
Perfusion. To verify whether the IL-1β measured was from blood or tissue in the CNS, animals were perfused with 0.9% saline before they were killed. In the same experiment, perfused rats were compared with nonperfused rats, and levels of IL-1β protein were measured in the hippocampus, hypothalamus, and posterior cortex.
Tissue collection. Animals were anesthetized with a brief exposure to ether, and brains were quickly removed after decapitation. All dissections were performed on a frosted glass plate placed on top of crushed ice, and brain structures were quickly frozen on dry ice. Brain samples, which included hypothalamus, hippocampus, cerebellum, posterior cortex, and nucleus tractus solitarius, were stored at −70°C until the time of sonication.
Tissue processing. Each tissue was added to 0.5–1.0 ml of Iscove’s culture medium containing 5% fetal calf serum and a cocktail enzyme inhibitor (in mm: 100 amino-n-caproic acid, 10 EDTA, 5 benzamidine-HCl, and 0.2 phenylmethylsulfonyl fluoride). Total protein was mechanically dissociated from tissue using an ultrasonic cell disruptor (Heat Systems, Inc., Farmingdale, NY). Sonication consisted of 10 sec of cell disruption at the setting 10. Sonicated samples were centrifuged at 10,000 rpm at 4°C for 10 min. Supernatants were removed and stored at 4°C until an ELISA was performed. Bradford protein assays were also performed to determine total protein concentrations in brain sonication samples.
ELISA. Sheep anti-rat IL-1β immunoaffinity-purified polyclonal antibody (Ab) was provided by Dr. Steven Poole (National Institute for Biological Standards and Control, Hertfordshire, England). The microtiter plates (96 flat-bottom wells, Immunoplate Maxisorp; Nunc, Roskilde, Denmark) were coated with this Ab overnight at 4°C. After washing the plates in assay buffer (0.01 mphosphate, 0.05 m NaCl, and 0.1% Tween 20, pH 7.2), 100 μl of rat IL-1β standards or samples was added to each well and incubated at room temperature for 4 hr. After washing the plates, 100 μl of biotinylated, immunoaffinity-purified polyclonal sheep anti-rat IL-1β Ab (1:2000) with 1% normal goat serum was added to each well and incubated at room temperature for 1 hr. The color was developed by use of avidin–HRP (Vector Laboratories, Burlingame, CA) and the chromogen orthophenylene diamine (Sigma). Plates were read at 490 nm, and results are expressed as picograms of IL-1/100 μg of total protein. The detection limit of this assay was ∼10 pg/ml. A commercially available rat IL-1β ELISA kit (R & D Systems, Minneapolis, MN) was also used to verify that similar results can be obtained using a different Ab to rat IL-1β. This ELISA kit uses a goat anti-rat IL-1β polyclonal Ab that can recognize both recombinant as well as natural rat IL-1β. No significant cross-reactivity was observed with the R & D Ab to recombinant human (r-human) IL-1RI, IL-1RII, and IL-1ra, r-rat IL-1α, IL-2, and IL-4, interferon-γ, tumor necrosis factor-α, and r-mouse IL-1α and IL-1ra. This R & D kit had a detection limit of ∼5 pg/ml.
Verification of rat IL-1β Ab specificity. The rat IL-1β Ab was tested for specificity to rat IL-1β by ECL Western blot analysis, ELISA on Sephadex column separation of brain sonicates, and immunoprecipitation. Western blot analysis consisted of loading ∼150 μg of total protein onto a 16% Tris-glycine polyacrylamide gel (Novex, San Diego, CA) and running for 90 min at 120 V. The proteins were then transferred onto a nitrocellulose membrane at 25 V for 90 min and blocked in 5% nonfat milk for 1 hr. The nitrocellulose blot was incubated in the sheep anti-rat IL-1β immunoaffinity-purified polyclonal Ab overnight at 4°C. After washing in Tris-buffered saline, the secondary Ab (rabbit anti-sheep IgG conjugated with HRP; Jackson ImmunoResearch, West Grove, PA) was added and incubated at room temperature for 1 hr. Enhanced chemiluminescence reagents (Amersham, Buckinghamshire, England) were added, and the nitrocellulose blot was exposed to x-ray film. Dark bands were detected on the film, showing where the secondary Ab was bound to the sheep anti-rat IL-1β Ab. A molecular weight standard ladder (Life Technologies, Gaithersburg, MD) ranging from 43 to 3 kDa was also used to verify the molecular weights of the protein bands that were detected by the rat IL-1β Ab. A detectable band occurred at ∼17 kDa, the approximate molecular weight of IL-1β. However, a second band was also present at 33 kDa, the approximate molecular weight of the IL-1 prohormone.
Total protein from the brain sonication supernatant was also separated according to the molecular weight of each protein on a Sephadex G-50 column. Sephadex G-50 was packed into a 10 ml serological pipette column and was equlibrated with 50 mm Tris-HCl and 100 mm potassium chloride, pH 7.5. Five hundred microliters of supernatant were loaded onto the column, and fractions were collected every 30 sec at a flow rate of 1 ml/min. Fractions were tested for IL-1β using the ELISA. Columns were calibrated using a calibration kit (Sigma) consisting of known protein standards ranging from 6.5 to 2000 kDa. ELISA detected IL-1β in the fifth and sixth fractions, which consisted of proteins in the range of 15–30 kDa.
The rat IL-1β Ab was also tested for its ability to recognize pure rat IL-1β by immunoprecipitation. Pure rat IL-1β (Biosource, Camarillo, CA) dissolved in 10% fetal calf serum (Life Technologies) at concentrations of 1 μg–10 pg was incubated with the rat IL-1β Ab overnight at 4°C. Protein G–Sepharose was added to the mixture and allowed to incubate at room temperature for 2 hr. Protein G–Sepharose bound to the IL-1–Ab complex was precipitated and washed three times in 50 mm Tris-HCl, pH 7.5. After heat denaturation at 80°C for 5 min, the IL-1–Ab protein complex was loaded onto a 16% SDS-PAGE gel for electrophoresis separation. The gel was silver-stained, and protein bands were visualized on the gel. The rat IL-1β Ab did precipitate pure rat IL-1β. The sensitivity of the immunoprecipitation was at 10 ng of IL-1β, according to the silver-stained proteins located at ∼17 kDa, and was located at the same point on the gel as the pure IL-1β control.
Verification of adrenalectomy. Whole trunk blood was collected by decapitation after a brief exposure to ether when rats were killed. Whole blood was allowed to clot overnight at 4°C, and serum was separated by centrifugation and frozen at −20°C until later analysis. Serum levels of corticosterone were measured using a modification of a radioimmunoassay procedure described by Keith et al. (1978) (intra- and inter-assay coefficients of variation < 10%). Corticosterone Ab was purchased from Sigma (catalog #C-8784). The limit of detection for this assay is 0.5 μg/ml.
Verification of corticosterone replacement. Thymus glands were removed when rats were killed to verify the corticosterone replacement in the drinking water. Thymus weights were divided by the total weights of their respective rat to determine the change in thymic hypertrophy resulting from adrenalectomy without CORT replacement.
Statistical analysis. A 2 × 2 × 3 ANOVA was performed on the IL-1 data from the perfusion study comparing saline versus LPS; 2 × 5 ANOVAs were performed on the IL-1 time course study; single-factor ANOVA was performed on the circadian, LPS dose–response, thymus, and corticosterone data; and 2 × 3 ANOVAs were performed on all IL-1 data in adrenalectomy studies.
RESULTS
Effects of LPS on brain IL-1β protein
The first series of studies determined the ability of the ELISA to detect increases in brain IL-1β after peripheral LPS administration. Figure 1 shows IL-1β protein in the posterior cortex, hippocampus, and hypothalamus from animals that were and were not perfused. Samples were collected 7 hr after 1 mg/kg LPS or vehicle. The Ab obtained from Dr. S. Poole was used in the assay. Clearly, LPS produced a large increase in IL-1β protein levels in each of the regions [F(1,18) = 35.02;p < 0.001]. The contribution of blood levels of IL-1β protein was also examined. There were no significant differences between subjects that were perfused with physiological saline at 7 hr after saline or 1 mg/kg LPS compared with their nonperfused controls [F(1,18) = 0.159;p > 0.05]. Perfusing with physiological saline did not appear to alter the levels of IL-1β in the tissues of either saline controls or LPS-stimulated rats, because there were no reliable interactions between treatment and procedure [F(1,18) = 0.0454; p > 0.05].
Fig. 1.

Effects of LPS (1 mg/kg) and perfusion on levels of IL-1β protein in the posterior cortex, hippocampus, and hypothalamus as measured by ELISA using the anti-rat IL-1β Ab from Dr. S. Poole. Eleven rats received an intraperitoneal injection of sterile 0.9% saline as controls, whereas another 11 rats received an intraperitoneal injection of LPS. From these groups, six of the animals were perfused with 0.9% saline before tissue collection. All animals were killed at 7 hr after injections. Saline nonperfused,n = 5; saline perfused, n = 6; LPS nonperfused, n = 5; LPS perfused,n = 6.
One milligram per kg is a very high dose of LPS. Figure2 shows IL-1β protein levels 6 hr after either 0, 1, 10, or 100 μg/kg LPS. Samples here were assayed with the R & D ELISA kit, and only hippocampus was examined to moderate cost. LPS produced a dose-dependent increase in IL-1β [F(3,15) = 73.128; p < 0.0001].
Fig. 2.

Effects of LPS on levels of IL-1β protein in the hippocampus as measured by the R & D ELISA kit. Rats were killed at 6 hr after intraperitoneal injections of either 0.9% saline or LPS. Saline, n = 4; LPS, 1 μg/kg,n = 4; LPS, 10 μg/kg, n = 4; LPS, 100 μg/kg, n = 4.
Effects of inescapable tail shock on brain IL-1β protein
Rats were exposed to IS and killed either immediately (n = 6) or 2 hr (n = 6), 7 hr (n = 6), 24 hr (n = 6), or 48 hr (n = 6) later. The immediate and 24 and 48 hr subjects were all killed at 10:00 a.m. because the stress session was held constant at 8:00–10:00 a.m. Thus, these groups required only a single home cage control group (HCC) killed at 10:00a.m. Separate controls were also killed at 12:00 and 5:00p.m. to match the times for the 2 and 7 hr IS groups. Only hypothalamus (Fig. 3A) and hippocampus (Fig. 3B) were examined. As can be seen, IS did not alter levels of IL-1β protein in either region [F(1,10) = 0.755; p > 0.05 for the hypothalamus; F(1,9) = 1.762;p > 0.05 for the hippocampus].
Fig. 3.
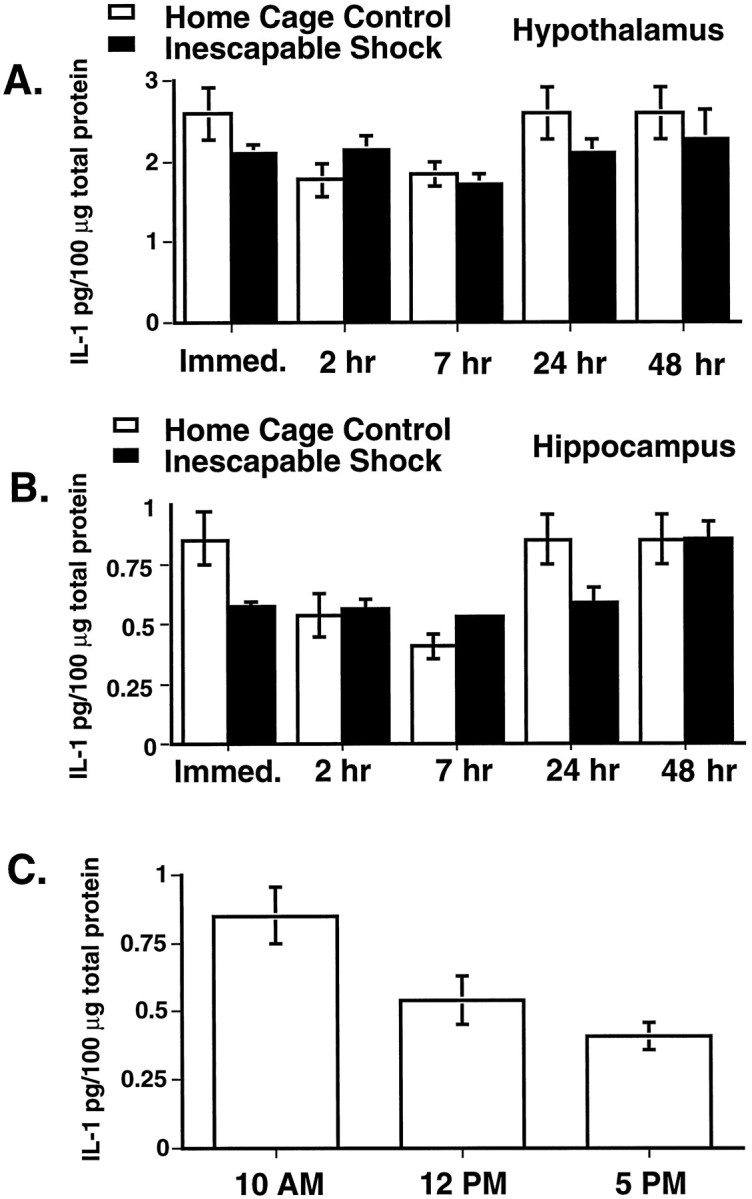
A, B, Levels of IL-1β protein in the hypothalamus (A) and hippocampus (B) as measured by ELISA using the anti-rat IL-1β polyclonal Ab from Dr. S. Poole. The immediate and 24 and 48 hr groups were killed at 10:00 a.m. The 2 and 7 hr groups were killed at 12:00 and 5:00 p.m., respectively. All groups contained six animals except the immediate and 24 and 48 hr HCC, which shared of the same six animals, because all were killed at the same time of the day. C, Circadian variation of rat IL-1β protein in the hippocampus. These data were taken from Bto clarify the potential circadian rhythmicity in HCC animals as measured throughout the sleep period. Each time point contained six animals.
Interestingly, IL-1β protein levels varied with time of day [F(4,40) = 4.45; p < 0.005 for the hypothalamus; F(4,36) = 8.492;p < 0.0001 for the hippocampus]. Figure 3Cshows this variation for HCC subjects in the hippocampus. Levels of IL-1β were highest at 10:00 a.m. near the beginning of the sleep cycle and lowest at 5:00 p.m. near the end of the sleep cycle [F(2,15) = 7.445; p< 0.01].
Effects of adrenalectomy on IS-induced increase in brain IL-1β protein
Adrenal corticosteroids inhibit IL-1β gene transcription (Lee et al., 1988; Amano et al., 1992). IS produces a rapid and prolonged rise in corticosteroids (Fleshner et al., 1995), and so it is possible that the corticosteroid response to IS masks possible alterations in IL-1β protein levels. To assess this possibility, rats were adrenalectomized or given sham surgery. Half of the adrenalectomized subjects received basal CORT replacement, and half did not. Figure4, A and B, shows brain IL-1β protein 2 hr after IS or HCC treatment in hypothalamus and hippocampus, respectively. With respect to hypothalamus, IS again had no effect on adrenal-intact subjects. However, IS produced a robust increase in IL-1β in adrenalectomized subjects, whether basal CORT was replaced [F(2,24) = 7.794;p < 0.005]. The pattern in hippocampus was similar, with the exception that the IL-1β increase produced by IS was only prominent in the adrenalectomized and basal CORT-replaced subjects [F(2,24) = 13.796; p < 0.0001].
Fig. 4.

A, B, Effects of adrenalectomy on levels of IL-1β protein in the hypothalamus (A) and hippocampus (B) at 2 hr after the end of the stress session. IL-1β protein results were obtained by ELISA using the anti-rat IL-1β polyclonal Ab from Dr. S. Poole. Sham HCC,n = 6; ADX HCC, n = 6; ADX-cort HCC, n = 6; sham IS, n = 3; ADX IS, n = 4; ADX-cort IS, n = 5.
These data were obtained using Ab from Dr. S. Poole. The implications of these data were sufficiently important that the experiment was repeated, and ELISAs were performed using the R & D kit. Furthermore, cerebellum, nucleus tractus solitarius, and posterior cortex were analyzed in addition to hypothalamus and hippocampus. Results from hypothalamus (Fig.5A) and hippocampus (Fig.5B) were similar to those obtained using Dr. Poole’s Ab. IS increased IL-1β protein in adrenalectomized but not sham subjects. Again this increase occurred in both CORT-replaced and nonreplaced subjects at 2 hr after IS. Large increases were also observed in cerebellum (Fig. 5C) and nucleus tractus solitarius (Fig.5D). In contrast, IS had no effect in posterior cortex (Fig.5E) under any of the conditions [F(2,38) = 5.777; p < 0.01 for hypothalamus; F(2,41) = 6.277; p< 0.005 for hippocampus; F(1,42) = 3.78;p < 0.05 for cerebellum;F(1,40) = 9.359; p < 0.0005 for nucleus tractus solitarius; and F(1,37) = 0.673;p > 0.05 for posterior cortex].
Fig. 5.

A–E, Effects of adrenalectomy on levels of IL-1β protein in the hypothalamus (A), hippocampus (B), cerebellum (C), nucleus tractus solitarius region (D), and posterior cortex (E) at 2 hr after the end of the stress session. IL-1β protein results were obtained using the R & D ELISA kit. Sham HCC, n = 8; sham IS, n = 6; ADX HCC, n = 6; ADX IS, n = 8; ADX-cort HCC, n = 8; ADX-cort IS,n = 8.
Verification of adrenalectomy and corticosterone replacement
Adrenalectomy was verified in the last IS experiment by measuring serum CORT after a brief ether exposure, which has been shown to rapidly activate the hypothalamo–pituitary–adrenal (HPA) response. Adrenalectomy eliminated the CORT response to ether [F(2,45) = 37.001; p < 0.0001] (data not shown). The CORT replacement procedure was verified by examining thymus weight relative to total body weight of each animal. Given that the thymus gland is very sensitive to levels of circulating CORT, adrenalectomy should cause thymic hypertrophy, because no corticosterone is present to influence lymphocyte maturation in the thymus. If the corticosterone replacement therapy in the adrenalectomized rats mimics natural basal CORT levels, a normalization of thymus weight would be expected. Adrenalectomy caused a significant increase in thymus weight, and this increase was normalized in the adrenalectomized group that received CORT replacement [F(2,45) = 22.517; p < 0.0001] compared with sham-operated controls (data not shown).
DISCUSSION
The purposes of the present experiments were to determine whether IL-1β protein could be detected in rat brain under basal nonstimulated conditions, and whether a stressor would alter these levels. The ELISA was adopted as the measurement technique, because Ab to rat IL-1β was available, and ELISAs are quite sensitive. Specificity of measurement is an issue with virtually all techniques designed to measure protein levels and is particularly problematic for Ab-based procedures. It is straightforward to determine whether an Ab recognizes the protein of interest, but it is not easy to prove that the epitope that it recognizes is not also contained on other proteins. To test Ab specificity for rat IL-1β, several approaches were used. First, Western blot showed a strong band at the molecular weight of IL-1β and only one other faint band. This band was at the molecular weight of the prohormone for IL-1β. Thus it may be that the data obtained partially reflect increases in the prohormone. This must simply be acknowledged, and the reader should bear this in mind. Second, the ELISA detected as much IL-1β in a fractionated sample as in the whole sample, suggesting that the predominant molecule measured was indeed mature IL-1β. Third, the Ab precipitated rat IL-1β in an immunoprecipitation procedure. Fourth, the Poole Ab has been studied for cross-reactivity with a large number of other proteins (Garabedian et al., 1995), and none was found. Finally, similar results were obtained with two different Abs.
To further address this issue, we next sought to determine whether the ELISA measured increases in brain IL-1β protein under conditions in which IL-1β increases have been detected with other measurement procedures. The peripheral administration of large doses of LPS has been reported to increase brain IL-1β bioactivity (Fontana et al., 1984; Coceani et al., 1988; Quan et al., 1994), immunoreactivity (Van Dam et al., 1992; Hagan et al., 1993; Hillhouse and Mosley, 1993), and mRNA (Ban et al., 1992; Buttini and Boddeke, 1995; Laye et al., 1994). A very large dose of LPS (1 mg/kg) was chosen as a positive control in the first experiment, not to test whether brain IL-1β is involved in mediating the usual effects of LPS. Lower doses of LPS (1–100 μg) were also examined using the R & D ELISA kit, which uses a different Ab specific to rat IL-1β. LPS did produce large increases in IL-1β protein in the brain, as measured by this ELISA as well. As with other measurement techniques, this increase appeared to be regionally nonspecific. In addition, brain IL-1β mRNA has recently been reported to have a circadian rhythm (Taishi et al., 1997) that is roughly the inverse of the glucocorticoid rhythm. IL-1β protein was therefore measured here at different times of the day, and a circadian rhythm was also obtained in which IL-1 protein levels were highest near the beginning of the light period and lowest near the dark period. This circadian rhythmicity is entirely in keeping with the proposal that brain IL-1 plays a physiological role in sleep regulation (Opp and Krueger, 1991).
In sum, all of the results were consistent with the proposition that the ELISA detected IL-1β protein and was sensitive to changes in IL-1β protein levels. The detection of IL-1β protein in the undisturbed animal is consistent with the careful work of Quan et al. (1996) showing IL-1β bioactivity in undisturbed animals. IL-1β specificity was verified in their bioassay system by demonstrating that the measured bioactivity was completely blocked by the IL-1 receptor antagonist. It must be noted that IL-1β mRNA has often been undetectable under basal conditions (Gatti and Bartfai, 1993), but IL-1β protein may be a more sensitive index of basal IL-1β in brain, because IL-1β mRNA may only be transcribed after IL-1β has been recently released or used. In addition, IL-1β mRNA may not be an abundant brain mRNA and therefore may be difficult to detect, relative to background, without amplification such as that provided by reverse transcription-PCR (Laye et al., 1994; Taishi et al., 1997). Furthermore, glucocorticoids specifically reduce IL-1β mRNA stability (Lee et al., 1988; Amano et al., 1992), thereby rendering IL-1β mRNA difficult to detect under a variety of conditions.
Inescapable shock had no detectable effect on IL-1β protein levels measured immediately and 2, 7, 24, or 48 hr after exposure to the stressor. However, glucocorticoids are known to potently suppress IL-1β transcription and mRNA stability (Lee et al., 1988), and IS produces a rapid increase in glucocorticoid levels that persist for roughly 1 hr after the termination of the stressor (Fleshner et al., 1995). Thus, the stressor might have provided input that would normally increase brain IL-1β signal, but any potential increases might have been inhibited by the glucocorticoids that are simultaneously stimulated. Indeed, Goujon et al. (1995) reported that adrenalectomy enhanced brain IL-1β mRNA increases produced by peripheral administration of LPS. Although an increase in IL-1β protein was not detected in adrenal-intact animals after stress, an effect of stress on brain IL-1β may nevertheless still exist. Stress could easily produce a small increase in IL-1β that is not detectable by ELISA, and small amounts of IL-1β may have biological effects given its potency. In addition, stress could modulate pro-IL-1 production or increase interleukin-1β-converting enzyme activity, changes that may occur at a later time than assayed here or be unmeasurable by the ELISA used in these studies.
The effects of IS on brain levels of IL-1β protein were therefore examined in adrenalectomized subjects. Indeed, adrenalectomy unmasked a stress-induced increase in brain IL-1β protein 2 hr after the stressor in both hypothalamus and hippocampus. This increase was also observed in adrenalectomized animals that were given basal corticosterone replacement, supporting the argument that it was the stress levels of glucocorticoids, rather than basal steroid, that inhibited the stress-induced production of brain IL-1β protein in the previous study. Interestingly, the IL-1β increase was regionally specific, with no detectable increase in the posterior cortex. The effects of immobilization observed by Minami et al. (1991) were also regionally specific, with increases in mRNA for IL-1β present in the hypothalamus but not cerebellum or midbrain. The brain regions examined in the present experiments were chosen based on previous observations implicating them in stress responses and peripheral immune challenge. The hypothalamus is a critical site mediating responses to IL-1β such as HPA activation and fever induction (Sapolsky et al., 1987; Klir et al., 1994). The cerebellum and the posterior cortex were chosen as control regions, largely because they have not been considered as important brain sites in the development of stress responses.
Although the source of brain IL-1β was not addressed in the present studies, several experiments have examined this issue. Human fetal microglia, as well as rat microglia, have been shown to express mRNA for IL-1β after peripheral LPS (Lee et al., 1993; Buttini and Boddeke, 1995). Immunoreactivity has also been found for IL-1β in ramified microglia and endothelial cells lining the venules (Van Dam et al., 1995). Furthermore, in situ hybridization has detected IL-1β mRNA for IL-1 in murine brain microvessel endothelial cells (Fabry et al., 1993). Although controversial, a neural source of IL-1β has also been reported (Molenaar et al., 1993; Pestarino et al., 1997).
The present data suggest a parallel between inflammatory stimuli and stressors, a parallel that has been noted often (Dunn, 1995). Similarities between the neurochemical, endocrine, and behavioral sequelae of immune challenge and stressors have been described frequently (Maier and Watkins, 1998). The physiological and psychological effects of immune challenge can resemble those caused by stressors. Influenza viral infection has been shown to increase plasma corticosterone and brain catecholamine metabolism (Dunn et al., 1989), effects that are also observed in response to stress (Shintani et al., 1995b). Yirmiya (1995) has recently reported depressive-like behaviors in rats after an endotoxin challenge that include reduced social interaction, food consumption, and locomotor activity, all of which have been implicated as responses to stressors (Johnson et al., 1992). The present data add one more parallel, namely, the production of increases in brain IL-1β. As with other sequelae of immune stimulation and stress, the IL-1β changes showed a family resemblance, not an identity. The regional distribution of IL-1β changes were not the same in the two cases, just as patterns of neural activation are not identical after stress and immune challenge (Rivest et al., 1995). However, patterns of neural activation differ between stressors (Li et al., 1996), so identity is not to be expected.
Finally, an activation of IL-1 usage by the IS stressor used in the present studies is entirely in keeping with documented similarities between the effects of IS and the intracerebroventricular administration of IL-1β. Intracerebroventricular IL-1 produces reductions in social interaction (Propes and Johnson, 1997), reductions in food and water intake (Kent et al., 1994), suppressed peripheral immune responses (Sundar et al., 1989), fever (Ovadia et al., 1989), and aspects of the acute phase response such as increases in acute phase proteins and decreases in negative reactants, such as carrier proteins (Morimoto et al., 1988; Morimoto et al., 1989). IS produces all of these (Laudenslager et al., 1988; Fleshner et al., 1992; Short and Maier, 1993), even prolonged fever, increases in circulating acute phase proteins, and decreases in circulating carrier proteins (Deak et al., 1997). It is important to note that all stressors do not produce these outcomes; they are specific to the stressor used. It may be that these outcomes are mediated by the induction of brain IL-1β, and that there will be selectivity in terms of which stressors do, and do not, alter brain IL-1β.
Footnotes
This work was supported by National Institutes of Health Grant MH45045, a National Institute of Mental Health minority student fellowship to K.T.N., and the Undergraduate Research Assistantship Program at the University of Colorado to S.M.O. We thank Dr. Steven Poole for the supply of anti-rat IL-1β antibody, Debra Berkelhammer for technical assistance on the bioassays, and Dr. Robert L. Spencer for assistance on the Western blot protein analysis.
Correspondence should be addressed to Dr. Kien T. Nguyen, Department of Psychology, Campus Box 345, University of Colorado, Boulder, CO 80309.
REFERENCES
- 1.Amano Y, Lee SW, Allison AC. Inhibition by glucocorticoids of the formation of interleukin-1α, interleukin-1β, and interleukin-6: mediation by decrease mRNA stability. Mol Pharmacol. 1992;43:176–182. [PubMed] [Google Scholar]
- 2.Ban E, Haour F, Lenstra R. Brain interleukin-1 gene expression induced by peripheral lipopolysaccharide administration. Cytokine. 1992;4:48–54. doi: 10.1016/1043-4666(92)90036-q. [DOI] [PubMed] [Google Scholar]
- 3.Bandtlow CE, Meyer M, Lindholm D, Spranger M, Heumann R, Thoenen H. Regional and cellular codistribution of interleukin-1β and nerve growth factor mRNA in the adult rat brain: possible relationship to the regulation of nerve growth factor synthesis. J Cell Biol. 1990;111:1701–1711. doi: 10.1083/jcb.111.4.1701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bluthe R-M, Michaud B, Kelley KW, Dantzer R. Vagotomy attenuates behavioural effects of interleukin-1 injected peripherally but not centrally. NeuroReport. 1996;7:1485–1488. doi: 10.1097/00001756-199606170-00008. [DOI] [PubMed] [Google Scholar]
- 5.Brown R, Li Z, Vriend CY, Nirula R, Janz L, Falk J, Nance DM, Dyck DG, Greenberg AH. Suppression of splenic macrophage interleukin-1 secretion following intracerebroventricular injection of interleukin-1β: evidence for pituitary-adrenal and sympathetic control. Cell Immunol. 1991;132:84–93. doi: 10.1016/0008-8749(91)90008-y. [DOI] [PubMed] [Google Scholar]
- 6.Buttini M, Boddeke H. Peripheral lipopolysaccharide stimulation induces interleukin-1β messenger RNA in rat brain microglia. Neuroscience. 1995;65:523–530. doi: 10.1016/0306-4522(94)00525-a. [DOI] [PubMed] [Google Scholar]
- 7.Coceani F, Lees J, Dinarello CA. Occurrence of interleukin-1 in cerebrospinal fluid of the conscious cat. Brain Res. 1988;446:245–250. doi: 10.1016/0006-8993(88)90883-9. [DOI] [PubMed] [Google Scholar]
- 8.Cunningham ET, De Souza EB. Interleukin-1 receptors in the brain and endocrine tissues. Immunol Today. 1993;14:171–176. doi: 10.1016/0167-5699(93)90281-o. [DOI] [PubMed] [Google Scholar]
- 9.Deak T, Meriwether JL, Fleshner M, Spencer RL, Abouhamze A, Moldawer LL, Grahn RE, Watkins LR, Maier SF. Evidence that brief stress may induce the acute phase response in rats. Am J Physiol. 1997;273:R1998–R2004. doi: 10.1152/ajpregu.1997.273.6.R1998. [DOI] [PubMed] [Google Scholar]
- 10.Dunn AJ. Role of cytokines in infection-induced stress. Ann NY Acad Sci. 1993;697:189–202. doi: 10.1111/j.1749-6632.1993.tb49932.x. [DOI] [PubMed] [Google Scholar]
- 11.Dunn AJ. Interactions between the nervous system and the immune system: implications for psychopharmacology. In: Bloom FE, Kupfer DJ, editors. Psychopharmacology: the fourth generation of progress. Raven; New York: 1995. pp. 719–733. [Google Scholar]
- 12.Dunn AJ, Powell ML, Meitin C, Small PA., Jr Virus infection as a stressor: influenza virus elevates plasma concentrations of corticosterone, and brain concentrations of MHPG and tryptophan. Physiol Behav. 1989;45:591–594. doi: 10.1016/0031-9384(89)90078-4. [DOI] [PubMed] [Google Scholar]
- 13.Fabry Z, Fitzsimmons KM, Herlein JA, Moninger TO, Dobbs MB, Hart MN. Production of the cytokines interleukin-1 and 6 by murine brain microvessel endothelium and smooth muscle pericytes. J Neuroimmunol. 1993;47:23–34. doi: 10.1016/0165-5728(93)90281-3. [DOI] [PubMed] [Google Scholar]
- 14.Fantuzzi G, Dinarello CA. The inflammatory response in interleukin-1 beta-deficient mice: comparison with other cytokine-related knock-out mice. J Leukoc Biol. 1996;59:489–493. doi: 10.1002/jlb.59.4.489. [DOI] [PubMed] [Google Scholar]
- 15.Fleshner M, Watkins LR, Lockwood LL, Bellgrau D, Laudenslager ML, Maier SF. Specific changes in lymphocyte subpopulation: a potential mechanism for stress-induced immunomodulation. J Neuroimmunol. 1992;41:131–142. doi: 10.1016/0165-5728(92)90063-q. [DOI] [PubMed] [Google Scholar]
- 16.Fleshner M, Deak T, Spencer RL, Laudenslager ML, Watkins LR, Maier SF. A long term increase in basal levels of corticosterone and a decrease in corticosteroid-binding globulin after acute stressor exposure. Endocrinology. 1995;136:5336–5342. doi: 10.1210/endo.136.12.7588279. [DOI] [PubMed] [Google Scholar]
- 17.Fontana A, Weber E, Dayer J-M. Synthesis of interleukin-1/endogenous pyrogen in the brain of endotoxin-treated mice: a step in fever induction? J Immunol. 1984;133:1696–1698. [PubMed] [Google Scholar]
- 18.Garabedian BS, Poole S, Allchorne A, Winter J, Woolf CJ. Interleukin-1β contributes to the inflammation-induced increase in nerve-growth factor levels and inflammatory hyperalgesia. Br J Pharmacol. 1995;115:1265–1275. doi: 10.1111/j.1476-5381.1995.tb15035.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gatti S, Bartfai T. Induction of tumor necrosis factor-α mRNA in the brain after peripheral endotoxin treatment: comparison with interleukin-1 family and interleukin-6. Brain Res. 1993;624:291–294. doi: 10.1016/0006-8993(93)90090-a. [DOI] [PubMed] [Google Scholar]
- 20.Goujon E, Parnet P, Laye S, Combe C, Dantzer R. Adrenalectomy enhances pro-inflammatory cytokines gene expression, in the spleen, pituitary and barin of mice in response to lipopolysacharide. Mol Brain Res. 1995;36:53–62. doi: 10.1016/0169-328x(95)00242-k. [DOI] [PubMed] [Google Scholar]
- 21.Hagan P, Poole S, Bristow AF. Endotoxin-stimulated production of rat hypothalamic interleukin-1β in vivo and in vitro, measured by specific immunoradiometric assay. J Mol Endocrinol. 1993;11:31–36. doi: 10.1677/jme.0.0110031. [DOI] [PubMed] [Google Scholar]
- 22.Higgins GA, Olschowka JA. Induction of interleukin-1β mRNA in adult rat brain. Mol Brain Res. 1991;9:143–148. doi: 10.1016/0169-328x(91)90139-o. [DOI] [PubMed] [Google Scholar]
- 23.Hillhouse EW, Mosley K. Peripheral endotoxin induces hypothalamic immunoreactive interleukin-1β in the rat. Br J Pharmacol. 1993;109:289–290. doi: 10.1111/j.1476-5381.1993.tb13567.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jacobson L, Akana SF, Cascio CS, Shinsako J, Dallman MF. Circadian variation in plasma corticosterone permit normal termination of adrenocorticotropin responses to stress. Endocrinology. 1988;122:1343–1348. doi: 10.1210/endo-122-4-1343. [DOI] [PubMed] [Google Scholar]
- 25.Johnson EO, Kamilaris TC, Chrousos GP, Gold PW. Mechanisms of stress: a dynamic overview of hormonal and behavioral homeostasis. Neurosci Biobehav Rev. 1992;16:115–130. doi: 10.1016/s0149-7634(05)80175-7. [DOI] [PubMed] [Google Scholar]
- 26.Keith LD, Winslow JR, Reynolds RW. A general procedure for estimation of corticosteroid response in individual rats. Steroids. 1978;31:523–531. doi: 10.1016/0039-128x(78)90034-x. [DOI] [PubMed] [Google Scholar]
- 27.Kent S, Rodriguez F, Kelley KW, Dantzer R. Anorexia induced by microinjection of interleukin-1β in the ventromedial hypothalamus of the rat. Physiol Behav. 1994;56:1031–1036. doi: 10.1016/0031-9384(94)90339-5. [DOI] [PubMed] [Google Scholar]
- 28.Kent S, Bret-Dibat JL, Kelley KW, Dantzer R. Mechanisms of sickness-induced decreases in food-motivated behavior. Neurosci Biobehav Rev. 1996;20:171–175. doi: 10.1016/0149-7634(95)00037-f. [DOI] [PubMed] [Google Scholar]
- 29.Klir JJ, McClellan JL, Kluger MJ. Interleukin-1β causes the increase in anterior hypothalamic interleukin-6 during LPS-induced fever in rats. Am J Physiol. 1994;266:R1845–R1848. doi: 10.1152/ajpregu.1994.266.6.R1845. [DOI] [PubMed] [Google Scholar]
- 30.Kluger MJ. Fever: the role of pyrogens and cryogens. Physiol Rev. 1991;71:93–127. doi: 10.1152/physrev.1991.71.1.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Krueger JM, Walter J, Dinarello CA, Wolf SM, Chedid L. Sleep promoting effects of endogenous pyrogen (interleukin-1). Am J Physiol. 1984;246:R994–R999. doi: 10.1152/ajpregu.1984.246.6.R994. [DOI] [PubMed] [Google Scholar]
- 32.Laudenslager ML, Fleshner M, Hofstadter P, Held PE, Simons L, Maier SF. Suppression of specific antibody production by inescapable shock: stability under varying conditions. Brain Behav Immun. 1988;2:92–101. doi: 10.1016/0889-1591(88)90010-4. [DOI] [PubMed] [Google Scholar]
- 33.Laye S, Parnet P, Goujon E, Dantzer R. Peripheral administration of lipopolysaccharide induces the expression of cytokine transcripts in the brain and pituitary of mice. Mol Brain Res. 1994;27:157–162. doi: 10.1016/0169-328x(94)90197-x. [DOI] [PubMed] [Google Scholar]
- 34.Lee SC, Liu W, Dickson DW, Brosman CF, Berman JW. Cytokine production by human microglia and astrocytes. Differential induction by lipopolysaccharide and IL-1β. J Immunol. 1993;150:2659–2667. [PubMed] [Google Scholar]
- 35.Lee SW, Tsou AP, Chan H, Thomas J, Petrie K, Eugui EM, Allison AC. Glucocorticoids selectively inhibit the transcription of the interleukin-1 beta gene and decrease the stability of interleukin-1 beta mRNA. Proc Natl Acad Sci USA. 1988;85:1204–1208. doi: 10.1073/pnas.85.4.1204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li HY, Ericsson A, Sawchenko PE. Distinct mechanisms underlie activation of hypothalamic neurosecretory neurons and their medullary catecholaminergic afferents in categorically different stress paradigms. Proc Natl Acad Sci USA. 1996;93:2359–2364. doi: 10.1073/pnas.93.6.2359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Maier SF, Watkins LR. Intracerebroventricular interleukin-1 receptor antagonist blocks the enhancement of fear conditioning and interference with escape produced by inescapble shock. Brain Res. 1995;695:279–282. doi: 10.1016/0006-8993(95)00930-o. [DOI] [PubMed] [Google Scholar]
- 38.Maier SF, Watkins LR (1998) Cytokines for Psychologist: Implication of bi-directional immune to brain communication for understanding behavior, mood, and cognition. Psychol Rev, in press. [DOI] [PubMed]
- 39.Minami M, Kuraishi Y, Yagaguchi T, Nakai S, Hirai Y, Satoh M. Immobilization stress induces interleukin-1β mRNA in the rat hypothalamus. Neurosci Lett. 1991;123:254–256. doi: 10.1016/0304-3940(91)90944-o. [DOI] [PubMed] [Google Scholar]
- 40.Molenaar GJ, Berkenbosch F, van Dam A-M, Lugard CM. Distribution of interleukin-1 beta immunoreactivity within the porcine hypothalamus. Brain Res. 1993;608:169–174. doi: 10.1016/0006-8993(93)90791-k. [DOI] [PubMed] [Google Scholar]
- 41.Morimoto A, Watanabe T, Sakata T, Murakami N. Leukocytosis induced by microinjection of endogenous pyrogen or interleukin-1 into the preoptic and anterior hypothalamus. Brain Res. 1988;475:345–348. doi: 10.1016/0006-8993(88)90623-3. [DOI] [PubMed] [Google Scholar]
- 42.Morimoto A, Murakami N, Yamaguchi T, Nakai S, Hirai Y. Brain regions involved in the development of acute phase responses accompanying fever in rabbits. J Physiol (Lond) 1989;416:645–657. doi: 10.1113/jphysiol.1989.sp017782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nieto-Sampedro M, Berman MA. Interleukin-1-like activity in rat brain: sources, targets, and effect of injury. J Neurosci Res. 1987;17:214–219. doi: 10.1002/jnr.490170303. [DOI] [PubMed] [Google Scholar]
- 44.O’Neill LA. Molecular mechanisms underlying the actions of the pro-inflammatory cytokine interleukin-1. Royal Irish Academy Medal Lecture. Biochem Soc Trans. 1997;25:295–302. doi: 10.1042/bst0250295. [DOI] [PubMed] [Google Scholar]
- 45.Oka T, Aou S, Hori T. Intracerebroventricular injection of interleukin-1β induces hyperalgesia in rats. Brain Res. 1993;624:61–68. doi: 10.1016/0006-8993(93)90060-z. [DOI] [PubMed] [Google Scholar]
- 46.Opp MR, Krueger JM. Anti-interleukin-1β reduces sleep and sleep rebound after sleep deprivation in rats. Am J Physiol. 1994;266:R688–R695. doi: 10.1152/ajpregu.1994.266.3.R688. [DOI] [PubMed] [Google Scholar]
- 47.Opp MR, Obal FJ, Krueger JM. Interleukin-1 alters rat sleep: temporal and dose-related effects. Am J Physiol. 1991;260:R52–R58. doi: 10.1152/ajpregu.1991.260.1.R52. [DOI] [PubMed] [Google Scholar]
- 48.Ovadia H, Abramsky O, Weidenfeld J. Evidence for the involvement of the central adrenergic system in the febrile response induced by interleukin-1 in rats. J Neuroimmunology. 1989;25:109–116. doi: 10.1016/0165-5728(89)90128-8. [DOI] [PubMed] [Google Scholar]
- 49.Pestarino M, De Anna E, Masini MA, Sturla M. Localization of interleukin-1 beta mRNA in the cerebral ganglion of the protochordate, Styela plicata. Neurosci Lett. 1997;222:151–154. doi: 10.1016/s0304-3940(97)13362-6. [DOI] [PubMed] [Google Scholar]
- 50.Plata-Salaman CR, Oomura Y, Kai Y. Tumor necrosis factor and interleukin-1β: suppression of food intake by direct action in the central nervous system. Brain Res. 1988;448:106–114. doi: 10.1016/0006-8993(88)91106-7. [DOI] [PubMed] [Google Scholar]
- 51.Propes MJ, Johnson RW. Role of corticosterone in the behavioral effects of central interleukin-1β. Physiol Behav. 1997;61:7–13. doi: 10.1016/s0031-9384(96)00350-2. [DOI] [PubMed] [Google Scholar]
- 52.Quan N, Sundar SK, Weiss JM. Induction of interleukin-1 in various brain regions after peripheral and central injections of lipopolysaccharide. J Neuroimmunol. 1994;49:125–134. doi: 10.1016/0165-5728(94)90188-0. [DOI] [PubMed] [Google Scholar]
- 53.Quan N, Zhang Z, Emery M, Bonsall R, Weiss JM. Detection of interleukin-1 bioactivity in various brain regions of normal healthy rats. Neuroimmunomodulation. 1996;3:47–55. doi: 10.1159/000097226. [DOI] [PubMed] [Google Scholar]
- 54.Rivest S, LaFlamme N, Nappi RE. Immune challenge and immobilization stress induce transcription of the gene encoding the CRF receptor in selective nuclei of the rat hypothalamus. J Neurosci. 1995;15:2680–2695. doi: 10.1523/JNEUROSCI.15-04-02680.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W. Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing Factor. Science. 1987;238:522–526. doi: 10.1126/science.2821621. [DOI] [PubMed] [Google Scholar]
- 56.Shintani F, Nakaki T, Kanba S, Kato R, Asai M. Role of interleukin-1 in stress responses: a putative neurotransmitter. Mol Neurobiol. 1995a;10:47–71. doi: 10.1007/BF02740837. [DOI] [PubMed] [Google Scholar]
- 57.Shintani F, Nakaki T, Kanba S, Sato K, Yagi G, Shiozawa M, Aiso S, Kato R, Asai M. Involvement of interleukin-1 in immobilization stress-induced increase in plasma adrenocorticotropic hormone and in release of hypothalamic monoamines in the rat. J Neurosci. 1995b;15:1961–1970. doi: 10.1523/JNEUROSCI.15-03-01961.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Short KR, Maier SF. Stressor controllability, social interaction, and benzodiazepine systems. Pharmacol Biochem Behav. 1993;45:827–835. doi: 10.1016/0091-3057(93)90128-g. [DOI] [PubMed] [Google Scholar]
- 59.Sundar SK, Becker KJ, Cierpial MA, Carpenter MD, Rankin LA, Fleener SL, Ritchie JC, Simson PE, Weiss JM. Intracerebroventricular infusion of interleukin-1 rapidly decreases peripheral cellular immune responses. Proc Natl Acad Sci USA. 1989;86:6398–6402. doi: 10.1073/pnas.86.16.6398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sundar SK, Cierpial MA, Kilts C, Ritchie JC, Weiss JM. Brain interleukin-1-induced immunosuppression occurs through activation of both pituitary-adrenal axis and sympathetic nervous system by corticotropin-releasing factor. J Neurosci. 1990;10:3701–3706. doi: 10.1523/JNEUROSCI.10-11-03701.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Taishi P, Bredow S, Guha-Thakurta N, Obal FJ, Kruger JM. Diurnal variations of interleukin-1β mRNA and β-actin mRNA in rat brain. J Neuroimmunol. 1997;75:69–74. doi: 10.1016/s0165-5728(97)00002-7. [DOI] [PubMed] [Google Scholar]
- 62.Van Dam A-M, Brouns M, Louisse S, Berkenbosch F. Appearance of interleukin-1 in macrophages and in ramified microglia in the brain of endotoxin-treated rats: a pathway for the induction of non-specific symptoms of sickness? Brain Res. 1992;588:291–296. doi: 10.1016/0006-8993(92)91588-6. [DOI] [PubMed] [Google Scholar]
- 63.Van Dam A-M, Bauer J, Tilders FJH, Berkenbosch F. Endotoxin-induced appearance of immunoreactive interleukin-1β in ramified microglia in rat brain: a light electron microscopic study. Neuroscience. 1995;65:815–826. doi: 10.1016/0306-4522(94)00549-k. [DOI] [PubMed] [Google Scholar]
- 64.Weiss JM, Sundar SK, Cierpial MA, Ritchie JC. Effects of interleukin-1 infused into brain are antagonized by α-MSH in a dose-dependent manner. Eur J Pharmacol. 1991;192:177–179. doi: 10.1016/0014-2999(91)90087-7. [DOI] [PubMed] [Google Scholar]
- 65.Weiss JM, Quan N, Sundar SK. Widespread activation and consequences of interleukin-1 in the brain. Ann NY Acad Sci. 1994;741:338–357. doi: 10.1111/j.1749-6632.1994.tb23117.x. [DOI] [PubMed] [Google Scholar]
- 66.Yirmiya R. Endotoxin produces a depressive-like episode in rats. Brain Res. 1996;711:163–174. doi: 10.1016/0006-8993(95)01415-2. [DOI] [PubMed] [Google Scholar]
- 67.Zalcman S, Green-Johnson JM, Murray L, Nance DM, Dyck D, Anisman H, Greenberg AH. Cytokine-specific central monoamine alterations induced by interleukin-1,-2, and -6. Brain Res. 1994;643:40–49. doi: 10.1016/0006-8993(94)90006-x. [DOI] [PubMed] [Google Scholar]