

Abstract
Nitric oxide (NO) has been implicated in the establishment of precise synaptic connectivity throughout the neuroaxis in several species. To determine the contribution of NO to NMDA receptor-dependent dendritic growth in motor neurons, we administered the NMDA antagonist MK-801 to wild-type mice and neuronal nitric oxide synthase (nNOS) knock-out mice between postnatal days 7 and 14. Compared to saline-treated wild-type animals the number of dendritic bifurcations was significantly reduced in nNOS knock-out animals and MK-801-treated wild-type animals. There was no significant difference in dendritic bifurcation between MK-801-treated wild-type, MK-801-treated nNOS knock-out, and saline-treated nNOS knock-out animals, suggesting that nNOS knock-out and NMDA receptor block had similar effects. The path of the longest dendrite and the number of primary dendrites was the same in all treatment groups, indicating an effect specific to bifurcation. Sholl analysis revealed that differences in bifurcation numbers occurred between 160 and 320 μm from the cell body, the distance at which second, third, and fourth order dendrites are most prevalent. Dendrite order analyses confirmed a significant reduction in numbers, but not lengths, of third and fourth order dendrites in nNOS knock-out and drug-treatment groups. Finally, immunohistochemical examination of the developing spinal cord indicated that NMDA receptors and nNOS are colocalized within interneurons surrounding the motor neuron pool. These results support the view that at least part of NMDA receptor-dependent arborization of motor neuron dendrites is mediated by the local production of NO within the developing spinal cord.
Keywords: nitric oxide, nitric oxide synthase, NMDA receptor, development, motor neuron, spinal cord, dendrite, synaptic plasticity
Activity-dependent control of synaptic rearrangements during development involves the dynamic interplay between axons and dendrites during a critical period in early postnatal life. During this period, it is believed that axonal and dendritic arbors with stable synapses are preserved (or growth is promoted) whereas those portions of arbor not harboring stable synapses are eliminated (Cline et al., 1987; Constantine-Paton et al., 1990;Shatz, 1990; Schilling et al., 1991; Fox and Daw, 1993; Goodman and Shatz, 1993; Seil and Drake-Baumann, 1994). Studies of the developing visual system of frogs and mammals have led to the view that repeatedly coactivated presynaptic and postsynaptic elements are a necessary prelude to synaptic stabilization (Kleinschmidt et al., 1987; Bear et al., 1990; Cline and Constantine-Paton, 1990; Hahm et al., 1991; Simon et al., 1992; Yen et al., 1993, 1995). In these systems, activation of NMDA receptors has been implicated as the molecular coincidence detector for correlated presynaptic and postsynaptic activation. The participation of the NMDA receptor in developmental synaptic stabilization and long-term potentiation (LTP) suggests that the two phenomena may share biochemical mechanisms (Kandel and O’Dell, 1992;Crair and Malenka, 1995; Fox, 1995; Kirkwood et al., 1995, 1996;Stryker, 1995; Cramer et al., 1996). It is believed that through these cellular and molecular interactions, the activity-dependent control of axonal and dendritic architecture regulates how patterns of synaptic connections emerge during development.
The development of the dendritic tree of rodent motor neurons has proven to be a useful model for the study of activity-dependent development. Establishment of mature motor neuron dendritic geometry, for example, depends on afferent input during a sensitive period in early postnatal life (O’Hanlon and Lowrie, 1996). During this time window, NMDA and non-NMDA receptors are expressed in particularly great abundance in the ventral horn of the developing spinal cord (Watanabe et al., 1994; Jakowec et al., 1995a,b) and LTP can be induced (Pockett and Figurov, 1993). Administration of NMDA receptor antagonists to neonatal but not adult animals blocks the molecular development and establishment of mature dendritic architecture of motor neurons (Kalb and Hockfield, 1990; Kalb and Agostini, 1993; Kalb, 1994). Given the demonstrated plasticity of immature motor systems (Kalverboer et al., 1993; Bloedel et al., 1996), it is likely that activity-dependent development of spinal cord circuitry subserves behaviorally relevant adaptation of motor function.
Activation of NMDA receptors leads to a transmembrane flux of Ca2+ through the receptor channel, and it is thought that this rise in intracellular Ca2+ plays a central role in activity-dependent development (Collins et al., 1991; Koike and Tanaka, 1991; Spitzer, 1994). Three major Ca2+-dependent effector molecules tied to synaptic plasticity through their functional link with NMDA receptors are calcium calmodulin-dependent kinase 2 (CaMKII), protein kinase C γ (PKCγ), and neuronal nitric oxide synthase (nNOS). Genetic and pharmacological studies have demonstrated the key role played by these effector molecules in LTP and long-term depression (Malenka et al., 1986; Linden et al., 1987; Malinow et al., 1989; Linden and Connor, 1991; Shibuki and Okada, 1991; Haley et al., 1992; Silva et al., 1992a; Abeliovich et al., 1993a; O’Dell et al., 1994; Kantor et al., 1996; Otmakhov et al., 1997), learning and memory (Silva et al., 1992b; Abeliovich et al., 1993b; Böhme et al., 1993; Cho et al., 1998), formation of olfactory memories (Kendrick et al., 1997), elimination of transient projections during development (Wu et al., 1994), stabilization of axonal growth cones at appropriate targets, and the maturation of dendritic arbor structure (Wu and Cline, 1998). However, the Ca2+-dependent effector system subserving NMDA receptor-dependent motor neuron dendrite growth has not been established. A leading candidate is nNOS since previous work has demonstrated a physical link (Brenman et al., 1996a,b) and functional coupling (Garthwaite et al., 1988) of nNOS to the NMDA receptor. In addition, we have found that NOS antagonists (Kalb and Agostini, 1993) are as effective as NMDA receptor antagonists (Kalb and Hockfield, 1990) in blocking some aspects of the molecular development of motor neurons. In the present study, we used mice in which the nNOS gene has been knocked out (Huang et al., 1993) to investigate the participation of nitric oxide (NO) signaling in NMDA receptor-mediated development of motor neuron dendrites. A comparison of the effects of application of NMDA receptor antagonists to wild-type and nNOS knock-out animals indicates that nNOS plays an essential role in NMDA receptor-mediated motor neuron dendrite growth.
MATERIALS AND METHODS
Mice. Mice in which nNOS has been knocked out were provided by Dr. Paul Huang, Harvard University, Boston, MA (Huang et al., 1993). Knock-out mice were from a background of 129/Sv × C57BL/6, back-crossed with C57BL/6, and were bred with wild-type C57BL/6 mice (National Institutes of Health, Bethesda, MD) to produce F1 heterozygotes. The heterozygotes were cross-bred to produce litters (F2) of mixed genotypes, with a ratio of 25% wild-type (+/+), 50% heterozygote (+/−), and 25% nNOS knock-out (−/−) mice. All experimental measurements were performed on F2 littermates. This ensured that variability within the genetic background of the mice was randomly distributed between (+/+) and (−/−) animals (Banbury conference on genetic background in mice, 1997). All experiments were performed in accordance with Yale Animal Care and Use Committee guidelines.
Drug treatment and tissue processing. Each animal in a litter was randomly assigned to receive daily intraperitoneal injection of MK-801 diluted in saline (5 mg/kg; 1 μl/gm body weight) or saline (3 μl), from postnatal day 7 (P7) to P14. Twenty-four hours after the last injection, mice were anesthetized deeply with chloral hydrate and perfused transcardially with 0.1 m PBS, pH 7.4, followed by 4% paraformaldehyde in 0.1 m phosphate buffer, pH 7.4 (PFA). The spinal cord, with ventral roots intact, was dissected and stored in PFA. Before perfusion, a 1 cm portion of tail was removed and used for genotyping.
Genotyping. Mouse tails were digested with proteinase K (0.5 mg/ml; Boehringer Mannheim, Indianapolis, IN), and the genomic DNA was extracted by phenol–chloroform and ethanol precipitation, and amplified by PCR. The PCR conditions used were 30 cycles of 94°C (30 sec), 60°C (30 sec), and 72°C (60 sec), followed by a final 5 min incubation at 72°C for chain elongation. Two sets of primers were used in each reaction tube, one for amplification of a 404 base pair region of the nNOS gene, and the other for amplification of a 603 base pair region of the neomycin resistance gene, inserted in the locus of the nNOS gene in knock-out mice (Huang et al., 1993). Primer sequences for detection of the nNOS gene were: 5′ CCT TTG AGA GTA AGG AAG GGG GCG GG 3′ (B1 primer) and 5′ GGG CCG ATC GTT GAC TGC GAG AAT GAT G 3′ (B2 primer); and for detection of the neomycin resistance gene were: 5′ ATG AAC TGC AGG ACG AGG CAG CG 3′ (CF 13 primer) and 5′ GGC GAT AGA AGG CGA TGC GCT G 3′ (CF 14 primer).
Retrograde labeling of motor neurons. To label the dendritic tree of motor neurons in the lumbar spinal cord, the fluorescent tracer DiI (Molecular Probes, Eugene, OR) was applied to the ventral roots of the lumbar enlargement of fixed spinal cord. Approximately four roots were labeled, corresponding to a region between L2 and L5. Cords were maintained in PFA at 37°C for 10–14 d. Each cord was then cut on a vibrating microtome (Electron Microscopy Sciences, Fort Washington, PA) into 80 μm transverse sections. The sections were mounted on glass slides and examined using epifluorescent rhodamine optics. Between five and ten motor neurons were analyzed in each spinal cord. Although this method of identifying neonatal motor neuron dendrites does not delineate the entire dendritic arbor of each neuron, it provides a reliable measure of dendritic arbor morphology that can be used to compare groups of animals.
Data collection and analysis. Fluorescently labeled motor neurons were traced using a computer-assisted camera lucida program, Neurolucida (Microbrightfield, Colchester, VT). The following primary measurements were made: cell body area, number of primary dendrites originating from the cell body, number of bifurcations, total arborization, and longest dendritic path per cell. Statistical comparisons of variance between treatment groups were performed using ANOVA (SAS).
To investigate whether alterations in dendritic parameters were localized to a specific portion of the dendritic tree, we used a modified Sholl analysis (Sholl, 1953) in which the amount of dendritic arbor was calculated within concentric radii drawn at 20 μm intervals, originating at the center of the cell body. Statistical comparisons of groups were made using repeated measures ANOVA, with radial distance as the repeated measure. We also performed analysis of dendrites according to their order, considering a dendrite emanating directly from the cell body as the primary dendrite; once it bifurcates, two secondary dendrites are formed and so on. The average number of dendrites in each order and an average length of dendrites of a particular order were calculated per cell, and treatment groups were compared using repeated measures ANOVA.
All post hoc comparisons between groups were performed using Student-Newman–Keuls test, with significance set atp < 0.05.
Immunohistochemistry. In addition to genotyping neonatal animals using tail DNA and PCR, we confirmed the presence or absence of nNOS by immunohistological staining of 50-μm-thick slices of PFA-fixed P7 spinal cord using an affinity-purified polyclonal rabbit antibody against nNOS (gift of Dr. David Bredt, University of California at San Francisco, San Francisco, CA). Tissue sections were incubated overnight with anti-nNOS antibodies (0.25 μg/ml), then for 2 hr in biotinylated goat anti-rabbit antibodies (Amersham, Arlington, IL). Immunoreactivity was visualized with a peroxidase–diaminobenzidine reaction (Vectastain ABC system; Vector laboratories, Burlingame, CA), and sections were mounted on glass slides and coverslipped.
Colocalization of nNOS and NMDA receptor subunits was accomplished by incubating P7 spinal cord slices simultaneously with a mouse monoclonal antibody to anti-nNOS (Sigma, St. Louis, MO; 1:1000) and a variety of different rabbit antibodies to NMDA receptor subunits. Dr. Robert Wenthold (National Institutes of Health, Bethesda, MD) provided antibodies to the NR1 subunit (used at 2 μg/ml), and Dr. Masahiko Watanabe (Hokkaido University, Hokkaido, Japan) provided antibodies to the NR1 and NR2A subunits (used at 0.5 μg/ml). After overnight incubation in primary antibody, tissue sections were incubated with Cy3-conjugated anti-rabbit antibody (Jackson ImmunoResearch, West Grove, PA) and fluorescein isothiocyanate-conjugated anti-mouse antibodies (Sigma) for 4 hr, mounted on slides with Vectashield (Vector Laboratories) and viewed with epifluorescent illumination. Colocalization of nNOS immunoreactivity and NMDA receptor subunit immunoreactivity at the cellular level was accomplished by viewing stained tissue sections alternatively with rhodamine and fluorescein optics. To control for fluorochrome cross-talk, some tissue sections were incubated with both primary antibodies but only one of the fluorescently conjugated secondary antibodies. In the absence of the Cy3 anti-rabbit antibody, no specific fluorescent signal was seen with rhodamine optics and, in the absence of the FITC anti-mouse antibodies, no fluorescent signal was seen with fluorescein optics.
RESULTS
In accordance with previous observations (Huang et al., 1993), it was not possible to distinguish (−/−) animals from (+/+) animals by gross inspection alone. Both (+/+) and (−/−) groups had similar body weights before and after saline treatment for 7 d (Table1). Comparison of animal weights using ANOVA indicated no significant group effect before drug or saline treatment (F(3,20) = 0.30; p = 0.99). Administration of MK-801 for 7 d resulted in lower body weights in both (+/+) and (−/−) groups, compared with saline-treated animals (Table 1). However, ANOVA revealed that although drug-treated animals tended to be lower in body weight than saline-treated animals after 7 d treatment, this trend did not reach significance (F(3,20) = 3.01; p = 0.054).
Table 1.
Effects of drug treatment on weight of animal
| +/+ saline (n = 7) | −/− saline (n = 5) | +/+ MK-801 (n = 5) | −/− MK-801 (n = 7) | |
|---|---|---|---|---|
| Weight before treatment | 4.0 ± 0.2 | 4.1 ± 0.4 | 4.0 ± 0.3 | 4.0 ± 0.2 |
| Weight after last treatment | 6.6 ± 0.4 | 6.1 ± 0.4 | 5.1 ± 0.4 | 5.1 ± 0.8 |
Results are mean ± SEM.
Effects of NMDA receptor antagonism on dendrite branching and arbor size in wild-type and nNOS knock-out animals
Neurons positive for DiI (Fig. 1) were analyzed using computer-assisted camera lucida tracing techniques. Figure 2 illustrates representative neurons drawn from (+/+) and (−/−) animals. The effects of nNOS knock-out and of MK-801 administration on various indices of dendritic growth are shown in Table 2. The number of primary dendrites emanating from the cell body was the same in each treatment group (F(3,164) = 0.54;p < 0.65), and the longest dendritic path from a cell, an indication of the maximum length to which a dendrite may grow, was unaltered by any treatment (F(3,164) = 1.67;p < 0.17). Thus, some basic features of dendritic architecture were not affected by drug treatment or genotype.
Fig. 1.

DiI labeling of motor neuron dendrites. The lipophilic dye DiI was applied to the ventral roots of fixed P14 spinal cords and, after a 10 d waiting period, horizontal tissue slices were prepared and viewed with rhodamine optics. A, In this low-power view of a hemicord, dorsal is up and lateral is right. Film exposure was set to enable visualization of distal dendrites into widespread regions of the spinal gray matter. At these settings, the fluorescent signal in the ventral horn is overexposed, making it impossible to distinguish individual motor neuron cell bodies. Scale bar, 25 μm. B, In this higher power view of the ventral horn, dorsal is up and lateral is right. Film exposure was set to enable visualization of motor neuron cell bodies and proximal portions of dendrites within the ventral horn. Scale bar, 45 μm. Figures were made by assembling scanned photographic images using Abode Photoshop.
Fig. 2.

Motor neurons from saline-treated animals expressing nNOS have a more complex dendritic tree than motor neurons from animals that do not express nNOS or have received MK-801. Composite camera lucida drawings of representative motor neurons from P14 animals that express (+/+) or do not express (−/−) the nNOS gene. Animals received saline or MK-801 (5 mg/mg) daily. The major effect of genotype is on the number of dendritic bifurcations and the number of third and higher order dendrites. No measurable differences were found in the number of primary dendrites, segment lengths per order, or the length of the longest dendrite. Scale bar, 100 μm.
Table 2.
Effects of nNOS knock-out and MK-801 administration on parameters of dendritic growth
| +/+ saline (n = 48) | −/− saline (n = 33) | +/+ MK-801 (n = 43) | −/− MK-801 (n = 45) | |
|---|---|---|---|---|
| Number of primary dendrites (ns) | 3.7 ± 0.2 | 3.8 ± 0.2 | 3.6 ± 0.1 | 3.8 ± 0.2 |
| Number of bifurcations (p < 0.001) | 10.3 ± 0.5 | 8.6 ± 0.6* | 7.4 ± 0.5* | 7.1 ± 0.4* |
| Total arborization (μm) (p < 0.001) | 2668 ± 108 | 2372 ± 111 | 2186 ± 111* | 2080 ± 119* |
| Longest dendrite (μm) (ns) | 446 ± 12 | 401 ± 12 | 424 ± 11 | 425 ± 18 |
| Cell body area (μm2) (p < 0.001) | 617 ± 47 | 719 ± 54 | 464 ± 35* | 546 ± 41** |
Results are mean ± SEM. Results of group comparisons made using ANOVA are indicated in parentheses after parameter. ns indicates no significant difference between groups. *indicates significant difference from +/+ saline, ** indicates significant difference from −/− saline (p < 0.05; Student-Newman–Keuls).
In contrast to these results, there were significant differences across the groups in the number of bifurcations as indicated by ANOVA (F(3,164) = 8.85; p < 0.001).Post hoc analysis within groups (Student-Newman–Keuls; p < 0.05) revealed that compared with (+/+) saline animals, there were significantly fewer bifurcations in the (−/−) saline group, a reduction of 17%. In wild-type animals MK-801 caused a statistically significant 28% reduction in the number of dendritic bifurcations, whereas in (−/−) animals, MK-801 caused a nonsignificant reduction in the number of dendritic bifurcations. No statistically significant differences existed between the number of bifurcations in the (−/−) saline, (+/+) MK-801, or (−/−) MK-801 treatment groups. Given that the amount of branching in the dendritic tree of the (−/−) animals is significantly reduced below that of (+/+) animals to begin with, it is noteworthy that MK-801 does not reduce the number of bifurcations in these animals below the value seen in the MK-801-treated (+/+) animals. This would suggest that the effect of MK-801 in (−/−) animals is not distinct from the effect of the gene knock-out itself on dendrite branching. This analysis implies that the presence of nNOS is necessary for the normal pattern of dendritic branching and indicates that the most prominent effect of NMDA receptor block on reducing motor neuron dendrite branching occurs in animals expressing the nNOS gene.
A reduction in dendrite branching in MK-801-treated (+/+) animals in comparison with all other groups of animals might be expected to reduce the size of the overall dendritic tree per cell, considering that the number of primary dendrites and longest dendrite does not differ between any treatment group. This suggestion is supported by the observation of significant differences across treatment groups in total amount of arborization (F(3,164) = 5.70;p < 0.001). Post hoc analysis within groups revealed that compared with (+/+) saline group there was a significantly smaller total arborization in the (+/+) MK-801 and (−/−) MK-801 treatment groups, reductions of 18 and 22%, respectively. The total arborization in the (−/−) saline-treated groups was 11% smaller than the (+/+) saline group, but this did not meet statistical significance. There were no significant differences between (−/−) saline-treated animals and either drug-treated group. The inherently greater variability in the total arborization measure may account for the lack of statistical significance. Alternatively, this result might suggest that in the absence of the correct number of branches in (−/−) animals, dendrites lengthen existing segments of the tree. However, this suggestion is at odds with the observation that the longest dendrite per cell is unaltered by any treatment.
Finally, there was a significant difference in cell body size, as measured by ANOVA (F(3,164) = 5.44;p < 0.001). Whereas the size of the cell body was not significantly different between (+/+) and (−/−) saline groups, MK-801 treatment resulted in a decrease in cell body area. The similarity in cell body size in both (+/+) and (−/−) saline-treated groups suggests that the effects we observed in (−/−) saline-treated animals are restricted to the dendritic tree, rather than representing simply an effect of NO on the overall growth of the neuron.
Effects of NMDA receptor block and nNOS on dendrites as a function of distance from cell body: Sholl analysis
Sholl analysis of the amount of dendritic arbor within concentric radii (20 μm) from the cell body revealed differences between groups (Fig. 3), suggesting that alterations in the dendritic tree might occur at specific distances from the cell body. At short distances from the cell body, the total amount of arbor per cell is the same for each group. This is confirmed by the observation that the number of primary dendrites does not differ between treatment groups. However, at a distance of ∼100 μm from the cell body, there is a divergence in the amount of arbor with respect to treatment group. This difference is maximal between ∼160 and 260 μm. At distances >260 μm, the amounts of arbor within each group begin to converge, such that at a distance of 440 μm from the cell body, there is little difference between groups. Statistical analysis confirmed differences in the magnitude of total dendrite arbor among the four treatment groups (F(3,164) = 5.955; p < 0.001). There was also a significant group × distance interaction (F(93,5084) = 4.548; p < 0.001), indicating that the pattern of arborization differs across treatment group. These observations are consistent with a decreased number of bifurcations in (−/−) animals and drug-treated animals, which would result in progressively smaller amounts of total arbor as the distance from the cell body increases.Post hoc analysis indicated that between 160 and 320 μm from the cell body in (+/+) animals MK-801 significantly reduced the amount of dendritic arbor. In addition, MK-801 reduced the amount of dendritic arbor between 200 and 240 μm from the cell body in the (−/−) animals. Although there was a trend for the amount of arbor to be lower in the (−/−) saline group compared with (+/+) animals, this trend was not significant at any radial distance from the cell body. These observations indicate that (1) MK-801 causes a larger reduction in bifurcation number than the absence of nNOS, and (2) MK-801 might have some effects on dendritic branching that are not mediated through nNOS.
Fig. 3.

Sholl analysis of dendritic length. Total dendritic length was measured in concentric radii (20 μm) from the cell body. Open circles represent saline-treated (+/+);filled circles, MK-801-treated (+/+); open squares, saline-treated (−/−); and filled squares, MK-801-treated (−/−) animals. * represents significant difference between saline- and MK-801-treated (+/+) animals; † represents significant difference between saline- and MK-801-treated (−/−) animals (p < 0.05; Student-Newman–Keuls).
Effects of NMDA receptor block and nNOS on the number and length of dendrite per order
It is possible that neurons might compensate for a reduction in the number of bifurcations and, consequently, fewer dendritic segments by lengthening existing segments of the dendritic tree. Such an observation might explain our failure to observe significantly reduced total arbor in the saline-treated (−/−) animals compared with the (+/+) animals, despite significant decreases in bifurcations (Table 2). To determine whether neurons would compensate for a reduction in bifurcations by extending segment length, we examined dendritic length as a function of order and calculated the number of dendrites of a particular order (i.e., primary, secondary, etc.). When dendrites were analyzed accordingly we found no treatment effect on the length of dendrites (Fig.4A) within any order (F(3,1172) = 0.04;p = 0.99). This suggests that the length of dendrite between bifurcations does not increase in the absence of nNOS or NMDA activity. When we examined the number of dendrites within each order, however (Fig. 4B), we found significant variation within treatment groups (F(3,1172) = 5.226;p < 0.01). Post hoc analysis revealed that saline-treated (−/−) animals and both MK-801-treated groups had significantly fewer dendrites of third and fourth orders compared with the saline-treated (+/+) animals, indicating that the absence of the nNOS gene or antagonism of the NMDA receptor could reduce the number of branches formed. In contrast, there were no differences in the number of primary or secondary dendrites between any of the treatment groups, confirming our initial finding that the number of primary dendrites is unaltered by either treatment. These results confirm that both nNOS and functional NMDA receptors are required for the normal branching of motor neuron dendrites and indicate that this effect is most prominent within a restricted part of the dendritic tree. There was also a smaller, but significant, effect of MK-801 in (−/−) animals. This effect was limited to third order dendrites, suggesting that a small portion of the effects of MK-801 are mediated through a nNOS-independent mechanism.
Fig. 4.
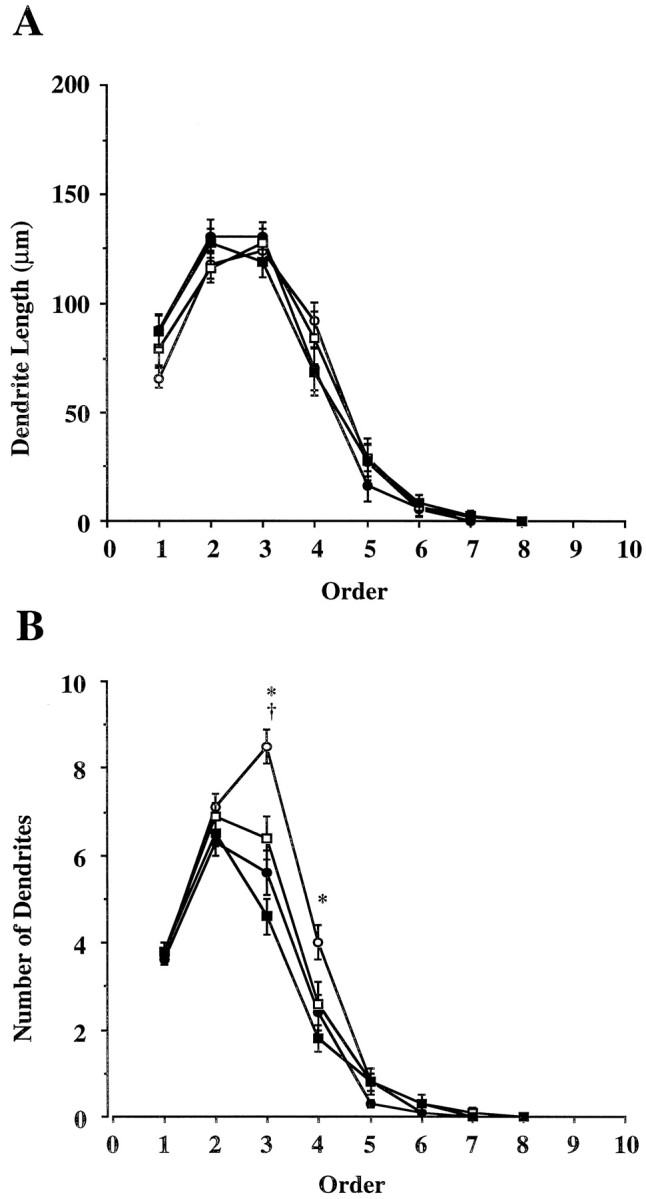
Analysis of dendrites according to dendrite order.Open circles represent saline-treated (+/+);filled circles, MK-801-treated (+/+); open squares, saline-treated (−/−); and filled squares, MK-801-treated (−/−) animals. A, Average length of dendrites in each order. B, Average number of dendrites in each order. * represents significant difference between saline-treated (+/+) and all other treatment groups; † represents significant difference between saline- and MK-801-treated (−/−) groups (p < 0.05; Student-Newman–Keuls).
Interneurons express both nNOS and NMDA receptor subunits
In neonatal wild-type rodents, nNOS is found in a subpopulation of interneurons adjacent to the motor neuron pool in the ventral horn (Kalb and Agostini, 1993) (Fig. 5). To determine whether these cells express NMDA receptor subunits, we undertook a series of double-labeling experiments using anti-nNOS antibodies and antibodies to NMDA receptor subunits NR1 and NR2A. We find that all nNOS-expressing interneurons within the P7 spinal cord ventral horn express immunoreactivity for NR1 (Fig.6) and NR2A (data not shown). These colocalization studies indicate that nNOS interneurons are capable of expressing functional NMDA receptors.
Fig. 5.
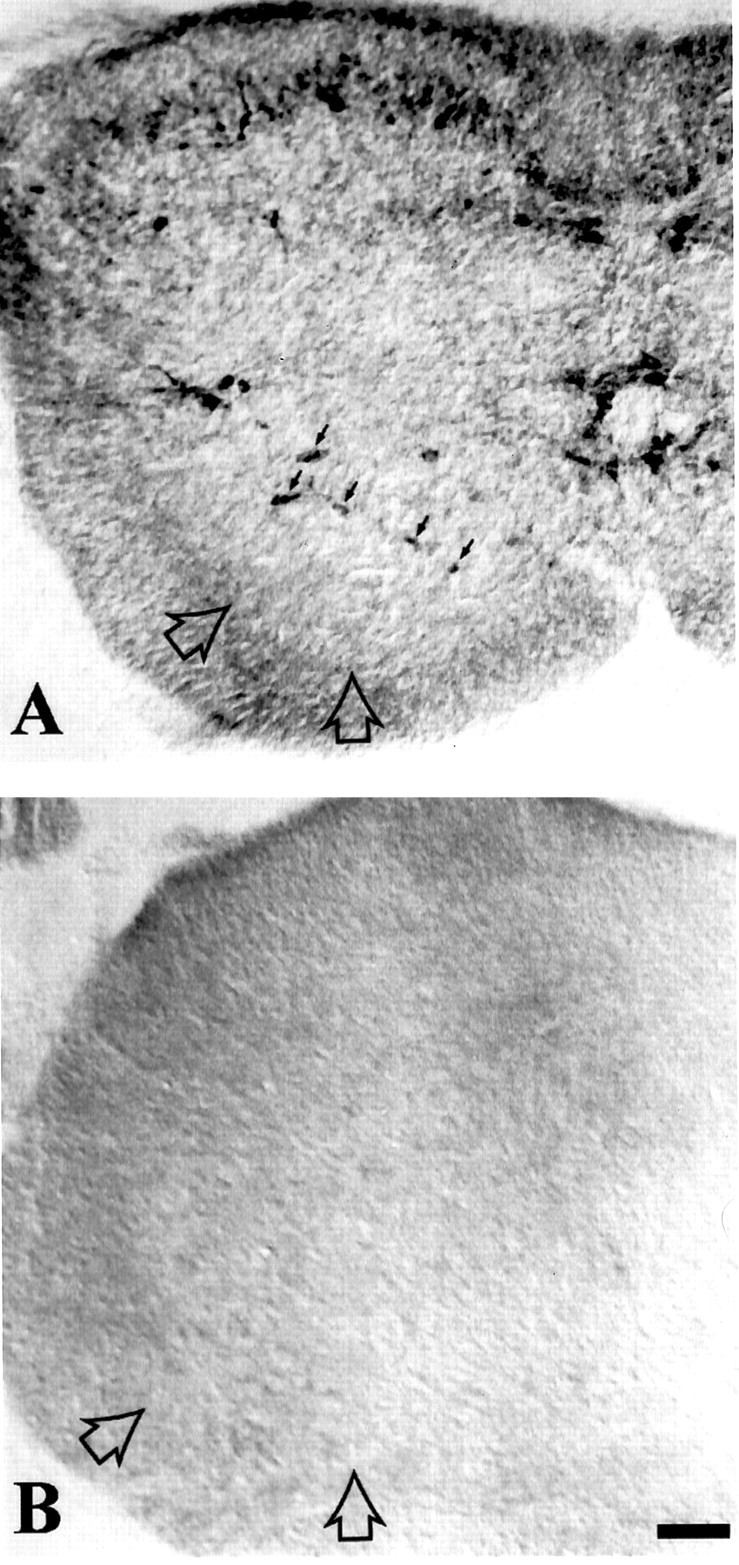
Neuronal NOS-expressing cells are present in the spinal cord of wild-type but not nNOS knock-out neonatal mice. Postnatal day 7 spinal cords were immunohistologically stained using a rabbit anti-nNOS serum and an HRP-conjugated anti-rabbit IgG secondary antibody followed by reaction with diaminobenzidine. Sections were mounted on gelatin-coated slides, dehydrated, and delipidated with xylene before coverslipping in permount. The sections were viewed with Nomarski optics on a Zeiss Axioskop microscope. Dorsal is up.A, In the spinal cord of wild-type animals, nNOS-immunoreactive neurons are seen in a ring (small arrows) surrounding the motor neuron pool (large open arrows). Neuronal NOS-immunoreactive neurons are also found in the substantia gelatinosa of dorsal horn and adjacent to the central canal. B, In the spinal cord of nNOS knock-out mice, no nNOS immunoreactivity is evident. The location of the motor neuron pool is noted (open arrows). Scale bar, 90 μm.
Fig. 6.
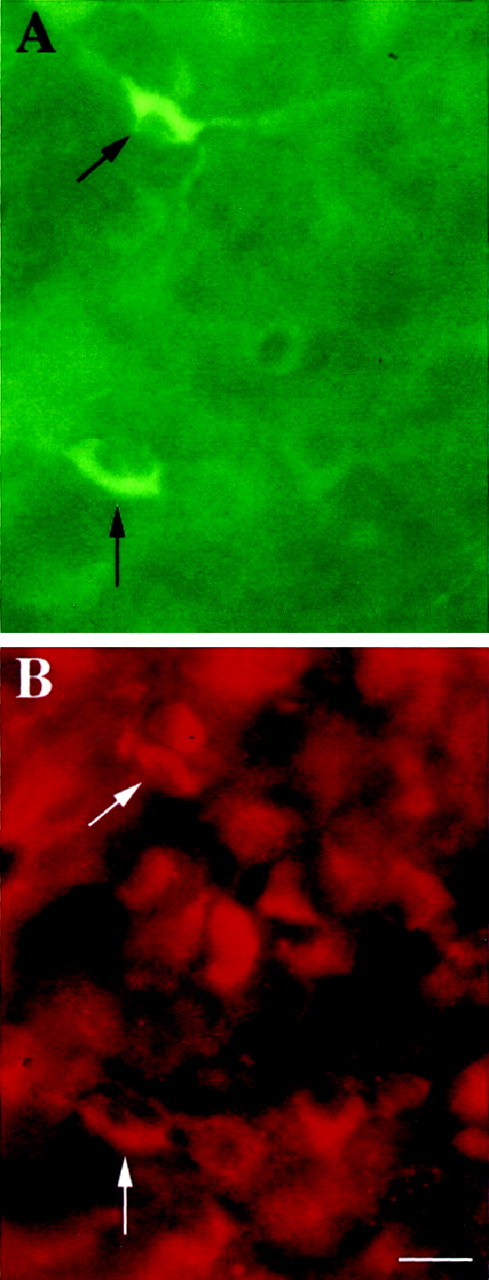
Spinal cord nNOS-containing interneurons express NMDA receptor subunit NR1. Postnatal day 7 spinal cords were immunohistologically stained using a mouse anti-nNOS monoclonal antibody followed by FITC-conjugated anti-mouse IgG antibody (A) and a rabbit anti-mouse NR1 serum (B) followed by Cy3-conjugated anti-rabbit IgG antibody. Controls to ensure specific staining are described in Materials and Methods. Tissue sections were mounted on gelatinized slides, and coverslips were mounted in Vectashield (Vector Laboratories) and viewed alternately with FITC and Rhodamine optics.A, Two nNOS-immunoreactive neurons (arrows) are visible among a field of nonreactive cells.B, The same field as in A now viewed with rhodamine optics reveals numerous NR1-immunoreactive neurons. The nNOS-immunoreactive cells seen in A also express NR1 immunoreactivity (arrows). Scale bar, 28 μm.
DISCUSSION
Investigation of the role of nNOS signaling in NMDA receptor-dependent development of the rodent spinal motor neuron dendritic tree has led to a number of insights into the molecular mechanisms governing activity-dependent dendrite growth. First, administration of the NMDA antagonist MK-801 reduces bifurcations in the dendritic tree of developing wild-type motor neurons to a value similar to the number of bifurcations found in untreated nNOS knock-out animals. Second, Sholl analysis of dendrites and analysis according to dendrite order indicated that block of NMDA receptors in wild-type animals has its effects on third or greater order dendrites, beginning at a distance of ∼160 μm from the cell body. There is also a smaller but statistically significant effect of NMDA receptor block on the number of third order dendrites in nNOS knock-out animals. Third, nNOS-containing interneurons adjacent to the motor neuron pool (Kalb and Agostini, 1993) express the appropriate subunits of the NMDA-type glutamate receptor to generate functional cell surface receptors. The most likely explanation for these results is that NMDA receptor activation leads to the local production of NO within the ventral horn, which acts as a intercellular messenger to promote the growth of motor neuron dendrites. This formulation is consistent with the recent demonstration of anterograde NO signaling between identified neurons inLymnaea stagnalis (Park et al., 1998).
In previous work we found that the ability of NMDA receptor antagonists to reduce motor neuron dendrite branching was the same when antagonists were administered systemically (by daily intraperitoneal injection of MK-801) or supplied locally to the lumbar enlargement (using the slow release plastic elvax impregnated with aminophosphonovaleric acid) (Kalb, 1994). Thus, we believe that in the present study, the locus of action of MK-801 is primarily on NMDA receptors in the segmental spinal cord. It should be noted, however, that MK-801 is a potent antagonist of NMDA receptors (Wong et al., 1986), that, at high doses, can induce locomotion, catalepsy, akinesia, and impaired food intake (Koek et al., 1988; Wishaw and Auer, 1989). Although it is conceivable that these actions in neonatal mice could generally depress neuronal growth, for several reasons this is not likely. The longest dendritic path in MK-801-treated animals was similar to that of saline-treated (+/+) animals, implying that MK-801 does not simply stunt the growth of motor neurons. In addition, the NMDA receptor-mediated NO-dependent effects are restricted to a distinct portion of the motor neuron dendritic tree (third and higher order dendrites) that largely corresponds to the region of overlap between nNOS-expressing interneurons and higher order motor neuron dendrites in Rexed layer VII. The simplest explanation for our findings is that MK-801 is acting to block NMDA receptors on spinal ventral horn nNOS interneurons.
Although there was clearly a large NMDA receptor-mediated effect that requires nNOS, the Sholl analysis and analysis of the number of dendrites per order also indicated that there was a smaller, but significant, drug effect in (−/−) animals. We consider several possibilities for this effect. First, MK-801 might exert some of its effect through inhibition of NMDA-dependent production of NO by another form of NOS, such as the endothelial form of NOS (eNOS), in nNOS (−/−) animals. Recent work indicates that eNOS is expressed in embryonic motor neurons (Estévez et al., 1998), and we have found that it is expressed by motor neurons throughout life (R. Kalb and W. Sessa, personal communication). It is also worth noting that a small amount of residual nNOS activity has been detected in the (−/−) animals because of the presence of an isoform of nNOS generated by alternative splicing that skips the targeted second exon of nNOS (Brenman et al., 1996b). The nNOS remaining in (−/−) animals could be functionally significant and subject to regulation by the activity of the NMDA receptor. Another consideration is that some NMDA receptors might mediate motor neuron growth independently of NO. If so, one role of NO might be to increase the number of synapses at which NMDA receptors can stimulate dendrite growth.
NO has been implicated in a variety of aspects of nervous system development including neurogenesis, synaptic efficacy, and axon motility (Roskams et al., 1994; Peunova and Enikolopov, 1995; Wang et al., 1995; Kuzin et al., 1996), and in some, but not all, aspects of activity-dependent synaptic rearrangements during postnatal development. Pharmacological antagonism of NOS blocks the ON/OFF sublamination of retinogeniculate afferents in the lateral geniculate nucleus (LGN) and retraction of the normally eliminated ipsilateral retinotectal projection in chick embryos (Wu et al., 1994; Cramer et al., 1996). On the other hand, NOS inhibitors have no demonstrable effect on the formation of ocular dominance columns or segregation of retinogeniculate fibers into eye-specific layers in the LGN (Reid et al., 1996; Ruthazer et al., 1996). All of these events in visual system development have previously been shown to occur in an NMDA receptor-dependent manner. These observations and our results support the principle that NMDA receptor-mediated development is a complex multistep process with both NO-dependent and NO-independent components.
What are the cellular interactions whereby activity-dependent, local production of NO within the ventral horn might regulate motor neuron dendrite growth? One possibility is that NO operates primarily on axon behavior and, thus, influences motor neuron dendrite structure indirectly. Gibbs and Truman (1998) have shown that blockade of NOS activity in Drosophila leads to disorganization of the projection of photoreceptor axons into the optic lobe. Since NO has been demonstrated to mediate growth cone collapse (Hess et al., 1993;Rentería and Constantine-Paton, 1995), it has been suggested that an NO “stop signal” may subserve the maintenance of initial contacts between ingrowing axons and their postsynaptic targets. An analogous situation might exist in the developing spinal cord because the establishment of patterned afferent input into motor neurons from segmental interneuronal and suprasegmental sources in early postnatal life is coincident with the period of major remodeling of the motor neuron dendritic tree (Curfs et al., 1993;Núñez-Abades et al., 1994). Activity-dependent release of NO within the ventral horn might participate in the signaling needed for ingrowing axons to cease growth near motor neuron dendrites. In the absence of NO either because of NMDA receptor block or the knock-out of the nNOS gene, some presynaptic inputs may fail to innervate motor neurons. Reduced growth of motor neuron dendrites would result from the deprivation of the tropic effect of synapses as postulated in the synaptotropic hypothesis of Vaughn et al. (1988). Such a scenario is supported by in vitro investigations: the branching of motor neuron dendrites is promoted by coculture with interneurons that form synapses on the cell body and dendrites of motor neurons (O’Brien and Fischbach, 1986).
NO may participate in a second, perhaps related, physiological process at the level of the synapse that is relevant to dendrite maturation. Current views of activity-dependent development postulate that changes in synaptic efficacy assayed electrophysiologically precede morphological alterations (Mooney et al., 1993; Crair and Malenka, 1995; Kirkwood et al., 1995). The induction of increases in synaptic gain, possibly involving an LTP-like mechanism, would be followed by synaptic stabilization, and the maintenance, during development, of otherwise transient axonal and dendritic arbors. In this regard it is noteworthy that some forms of LTP are likely to involve the NMDA receptor-dependent activation of NOS with NO presumed to act as an intercellular messenger initiating the presynaptic changes observed in LTP (Haley et al., 1992; Schuman and Madison, 1993; Doyle et al., 1996). These observations are germane to our findings in that motor neuron dendrite remodelling occurs at precisely the time during development at which NMDA receptors and nNOS are colocalized in the ventral horn (Kalb et al., 1992; Kalb and Agostini, 1993) and LTP can be induced (Pockett and Figurov, 1993). Thus the activity-dependent development of motor neuron dendrites might occur prominently at those portions of the tree where NO-dependent LTP occurs. In the absence of NMDA receptor activation or nNOS, such activity-dependent refinement of the dendrite tree would be limited.
One attractive feature of this formulation is that it can explain in part why activity-dependent development occurs exclusively in early postnatal life (Kalb, 1994). nNOS is only transiently expressed in the interneurons that surround the motor neuron pool (Kalb and Agostini, 1993), and high levels of NMDA (Kalb et al., 1992) and non-NMDA (Jakowec et al., 1995a,b) subtypes of glutamate receptors are present throughout the spinal cord for only the first few weeks of postnatal life. Thus, the molecular conditions required for the generation of an activity-dependent growth-promoting substance such as NO are only present for a brief window in early postnatal life.
It is clear that glutamatergic synaptic transmission during early postnatal life regulates the geometric features of the dendrite tree, with important consequences on the quantitative and qualitative aspects of synaptic input received by motor neurons (Hume and Purves, 1981;Purves and Hume, 1981). The complexity of the dendritic tree will also have important effects on neuronal computational capabilities because recent work indicates that branch points may operate as switches that control the back propagation of action potential from the cell body to dendrites (Spruston et al., 1995). Synapses located equidistant from the cell body but on different branches of the dendritic tree can experience disparate voltage and Ca2+ signals during repetitive action potential firing. The functional consequences at the cellular and network level of a more highly branched dendritic tree are under active exploration (Agmon-Snir et al., 1998; Segev, 1998).
Footnotes
This study was supported by U.S. Public Health Service Grants NS29837 and NS33437. The authors thank Paul Huang (Harvard University, Boston, MA) for the gift of mice, David Bredt (University of California at San Francisco, San Francisco, CA), Robert Wenthold (National Institutes of Health, Bethesda, MD), and Masahiko Watanabe (Hokkaido University, Hokkaido, Japan) for gifts of antibodies, Juliana Pakes and David Jentsch (Yale University) for their help with statistical analyses, and Ken Wikler for his help with photographic reproductions.
Correspondence should be addressed to Dr. Robert G. Kalb, Department of Neurology, Yale University School of Medicine, P.O. Box 208018, 333 Cedar Street, New Haven, CT 06520-8018.
REFERENCES
- 1.Abeliovich A, Chen C, Goda Y, Silva AJ, Stevens CF, Tonegawa S. Modified hippocampal long-term potentiation in PKCγ-mutant mice. Cell. 1993a;75:1253–1262. doi: 10.1016/0092-8674(93)90613-u. [DOI] [PubMed] [Google Scholar]
- 2.Abeliovich A, Paylor R, Chen C, Kim JJ, Wehner JM, Tonegawa S. PKCγ mutant mice exhibit mild deficits in spatial and contextual learning. Cell. 1993b;75:1263–1271. doi: 10.1016/0092-8674(93)90614-v. [DOI] [PubMed] [Google Scholar]
- 3.Agmon-Snir H, Carr CE, Rinzel J. The role of dendrites in auditory coincidence detection. Nature. 1998;393:268–272. doi: 10.1038/30505. [DOI] [PubMed] [Google Scholar]
- 4.Banbury Conference on Genetic background in Mice. Mutant mice and neuroscience: recommendations concerning genetic background. Neuron. 1997;18:755–759. doi: 10.1016/s0896-6273(00)80958-7. [DOI] [PubMed] [Google Scholar]
- 5.Bear MF, Kleinschmidt A, Gu Q, Singer W. Disruption of experience-dependent synaptic modifications in striate cortex by infusion of an NMDA receptor antagonist. J Neurosci. 1990;10:909–925. doi: 10.1523/JNEUROSCI.10-03-00909.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bloedel JR, Ebner TJ, Wise SP. The acquisition of motor behavior in vertebrates. MIT; Cambridge, MA: 1996. [Google Scholar]
- 7.Böhme GA, Bon C, Lemaire M, Reibaud M, Piot O, Stutzmann J-M, Doble A, Blanchard J-C. Altered synaptic plasticity and memory formation in nitric oxide synthase inhibitor-treated rats. Proc Natl Acad Sci USA. 1993;90:9191–9194. doi: 10.1073/pnas.90.19.9191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brenman JE, Christopherson KS, Craven SE, McGee AW, Bredt DS. Cloning and characterization of postsynaptic density 93, a nitric oxide synthase interacting protein. J Neurosci. 1996a;16:7407–7415. doi: 10.1523/JNEUROSCI.16-23-07407.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brenman JE, Chao DS, Gee Sh, McGee AW, Craven SE, Santillano DR, Wu Z, Huang F, Xia H, Peters MF, Froehner SC, Bredt DS. Interaction of nitric oxide synthase with the postsynaptic density protein PSD-95 and α1-syntrophin mediated by PDZ domains. Cell. 1996b;84:757–767. doi: 10.1016/s0092-8674(00)81053-3. [DOI] [PubMed] [Google Scholar]
- 10.Cho YH, Giese KP, Tanila H, Silva AJ, Eichenbaum H. Abnormal hippocampal spatial representations in α-CaMKIIT286A and CREBαΔ− mice. Science. 1998;279:867–869. doi: 10.1126/science.279.5352.867. [DOI] [PubMed] [Google Scholar]
- 11.Cline HT, Constantine-Paton M. NMDA receptor agonist and antagonists alter retinal ganglion cell arbor structure in the developing frog retinotectal projection. J Neurosci. 1990;10:1197–1216. doi: 10.1523/JNEUROSCI.10-04-01197.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cline HT, Debski EA, Constantine-Paton M. NMDA receptor antagonist desegregates eye-specific stripes. Proc Natl Acad Sci USA. 1987;84:4342–4345. doi: 10.1073/pnas.84.12.4342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Collins F, Schmidt MF, Guthrie PB, Kater SB. Sustained increase in intracellular calcium promotes neuronal survival. J Neurosci. 1991;11:2582–2587. doi: 10.1523/JNEUROSCI.11-08-02582.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Constantine-Paton M, Cline HT, Debski E. Patterned activity, synaptic convergence, and the NMDA receptor in developing visual pathways. Annu Rev Neurosci. 1990;13:129–154. doi: 10.1146/annurev.ne.13.030190.001021. [DOI] [PubMed] [Google Scholar]
- 15.Crair MC, Malenka RC. A critical period for long-term potentiation at thalamocortical synapses. Nature. 1995;375:325–328. doi: 10.1038/375325a0. [DOI] [PubMed] [Google Scholar]
- 16.Cramer KS, Angelucci A, Hahm J-O, Bogdanov MB, Sur M. A role for nitric oxide in the development of the ferret retinogeniculate projection. J Neurosci. 1996;16:7995–8004. doi: 10.1523/JNEUROSCI.16-24-07995.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Curfs MHJM, Gribnau AAM, Dereden PJWC. Postnatal maturation of the dendritic fields of motoneuron pools supplying flexor and extensor muscles of the distal forelimb in the rat. Development. 1993;117:535–541. doi: 10.1242/dev.117.2.535. [DOI] [PubMed] [Google Scholar]
- 18.Doyle C, Hölscher C, Rowan MJ, Anwyl R. The selective neuronal NO synthase inhibitor 7-nitro-indazole blocks both long-term potentiation and depotentiation of field EPSPs in rat hippocampal CA1 in vivo. J Neurosci. 1996;16:418–424. doi: 10.1523/JNEUROSCI.16-01-00418.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Estévez AG, Spear N, Thompson JA, Cornwell TL, Radi R, Barbeito L, Beckman JS. Nitric oxide-dependent production of cGMP supports the survival of rat embryonic motor neurons cultured with brain-derived neurotropic factor. J Neurosci. 1998;18:3708–3714. doi: 10.1523/JNEUROSCI.18-10-03708.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fox K. The critical period for long-term potentiation in primary sensory cortex. Neuron. 1995;15:485–488. doi: 10.1016/0896-6273(95)90136-1. [DOI] [PubMed] [Google Scholar]
- 21.Fox K, Daw NW. Do NMDA receptors have a critical function in visual cortical plasticity? Trends Neurosci. 1993;16:116–122. doi: 10.1016/0166-2236(93)90136-a. [DOI] [PubMed] [Google Scholar]
- 22.Garthwaite J, Charles SL, Chess-Williams R. Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain. Nature. 1988;336:385–388. doi: 10.1038/336385a0. [DOI] [PubMed] [Google Scholar]
- 23.Gibbs SM, Truman JW. Nitric oxide and cyclic GMP regulate retinal patterning in the optic lobe of Drosophila. Neuron. 1998;20:83–93. doi: 10.1016/s0896-6273(00)80436-5. [DOI] [PubMed] [Google Scholar]
- 24.Goodman CS, Shatz CJ. Developmental mechanisms that generate precise patterns of neuronal connectivity. Cell. 1993;72:77–98. doi: 10.1016/s0092-8674(05)80030-3. [DOI] [PubMed] [Google Scholar]
- 25.Hahm J-O, Langdon RB, Sur M. Disruption of retinogeniculate afferent segregation by antagonists to NMDA receptors. Nature. 1991;351:568–570. doi: 10.1038/351568a0. [DOI] [PubMed] [Google Scholar]
- 26.Haley JE, Wilcox GL, Chapman PF. The role of nitric oxide in hippocampal long-term potentiation. Neuron. 1992;8:211–216. doi: 10.1016/0896-6273(92)90288-o. [DOI] [PubMed] [Google Scholar]
- 27.Hess DT, Patterson SI, Smith DS, Skene JHP. Neuronal growth cone collapse and inhibition of protein fatty acetylation. Nature. 1993;366:562–565. doi: 10.1038/366562a0. [DOI] [PubMed] [Google Scholar]
- 28.Huang PL, Dawson TM, Bredt DS, Snyder SH, Fishman MC. Targeted disruption of the neuronal nitric oxide synthase gene. Cell. 1993;75:1273–1286. doi: 10.1016/0092-8674(93)90615-w. [DOI] [PubMed] [Google Scholar]
- 29.Hume RI, Purves D. Geometry of neonatal neurones and the regulation of synapse elimination. Nature. 1981;293:469–471. doi: 10.1038/293469a0. [DOI] [PubMed] [Google Scholar]
- 30.Jakowec MW, Fox AJ, Martin LJ, Kalb RG. Quantitative and qualitative changes in AMPA receptor expression during spinal cord development. Neuroscience. 1995a;67:893–907. doi: 10.1016/0306-4522(95)00026-f. [DOI] [PubMed] [Google Scholar]
- 31.Jakowec MW, Yen L, Kalb RG. In situ hybridization analysis of AMPA receptor subunit gene expression in the developing rat spinal cord. Neuroscience. 1995b;67:909–920. doi: 10.1016/0306-4522(95)00094-y. [DOI] [PubMed] [Google Scholar]
- 32.Kalb RG. Regulation of motor neuron dendrite growth by NMDA receptor activation. Development. 1994;120:3063–3071. doi: 10.1242/dev.120.11.3063. [DOI] [PubMed] [Google Scholar]
- 33.Kalb RG, Agostini J. Molecular evidence for nitric oxide-mediated motor neuron development. Neuroscience. 1993;57:1–8. doi: 10.1016/0306-4522(93)90107-q. [DOI] [PubMed] [Google Scholar]
- 34.Kalb RG, Hockfield S. Induction of a neuronal proteoglycan by the NMDA receptor in the developing spinal cord. Science. 1990;250:294–296. doi: 10.1126/science.2145629. [DOI] [PubMed] [Google Scholar]
- 35.Kalb RG, Lidow MS, Halsted MJ, Hockfield S. N-methyl-d-aspartate receptors are transiently expressed in the developing spinal cord ventral horn. Proc Natl Acad Sci USA. 1992;89:8502–8506. doi: 10.1073/pnas.89.18.8502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kalverboer AF, Hopkins B, Geuze R. Motor Development in early and later childhood: longitudinal approaches. Cambridge UP; Cambridge, UK: 1993. [Google Scholar]
- 37.Kandel ER, O’Dell TJ. Are adult learning mechanisms also used for development? Science. 1992;258:243–245. doi: 10.1126/science.1411522. [DOI] [PubMed] [Google Scholar]
- 38.Kantor DB, Lanzrein M, Stary SJ, Sandoval GM, Smith WB, Sullivan BM, Davidson N, Schuman EM. A role for endothelial NO synthase in LTP revealed by adenovirus-mediated inhibition and rescue. Science. 1996;274:1744–1748. doi: 10.1126/science.274.5293.1744. [DOI] [PubMed] [Google Scholar]
- 39.Kendrick KM, Guevara-Guzman R, Zorilla J, Hinton MR, Broad KD, Mimmack M, Ohkura S. Formation of olfactory memories mediated by nitric oxide. Nature. 1997;388:670–674. doi: 10.1038/41765. [DOI] [PubMed] [Google Scholar]
- 40.Kirkwood A, Lee H-K, Bear MF. Co-regulation of long-term potentiation and experience-dependent synaptic plasticity in visual cortex by age and experience. Nature. 1995;375:328–331. doi: 10.1038/375328a0. [DOI] [PubMed] [Google Scholar]
- 41.Kirkwood A, Rioult MG, Bear MF. Experience-dependent modification of synaptic plasticity in visual cortex. Nature. 1996;381:526–528. doi: 10.1038/381526a0. [DOI] [PubMed] [Google Scholar]
- 42.Kleinschmidt A, Bear MF, Singer W. Blockade of “NMDA” receptors disrupts experience-dependent plasticity of kitten striate cortex. Science. 1987;238:355–358. doi: 10.1126/science.2443978. [DOI] [PubMed] [Google Scholar]
- 43.Koek W, Woods JH, Winger GD. MK-801, a proposed noncompetitive antagonist of excitatory amino acid neurotransmission, produces phencyclidine-like behavioral effects in pigeons, rats and rhesus monkeys. J Pharmacol Exp Ther. 1988;245:969–974. [PubMed] [Google Scholar]
- 44.Koike T, Tanaka S. Evidence that nerve growth factor dependence of sympathetic neurons for survival in vitro may be determined by the levels of cytoplasmic free Ca2+. Proc Natl Acad Sci USA. 1991;88:3892–3896. doi: 10.1073/pnas.88.9.3892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kuzin B, Roberts I, Peunova N, Enikolopov G. Nitric oxide regulates cell proliferation during Drosophila development. Cell. 1996;87:639–649. doi: 10.1016/s0092-8674(00)81384-7. [DOI] [PubMed] [Google Scholar]
- 46.Linden DJ, Connor JA. Participation of postsynaptic PKC in cerebellar long-term depression in culture. Science. 1991;254:1656–1659. doi: 10.1126/science.1721243. [DOI] [PubMed] [Google Scholar]
- 47.Linden DJ, Sheu F-S, Murakami K, Routtenberg A. Enhancement of long-term potentiation by cis-unsaturated fatty acid: relation to protein kinase C and phospholipase A2. J Neurosci. 1987;7:3783–3792. doi: 10.1523/JNEUROSCI.07-11-03783.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Malenka RC, Madison DV, Nicoll RA. Potentiation of synaptic transmission in the hippocampus by phorbol esters. Nature. 1986;321:175–177. doi: 10.1038/321175a0. [DOI] [PubMed] [Google Scholar]
- 49.Malinow R, Schulman H, Tsien RW. Inhibition of post-synaptic PKC or CaMKII blocks induction but not expression of LTP. Science. 1989;245:862–866. doi: 10.1126/science.2549638. [DOI] [PubMed] [Google Scholar]
- 50.Mooney R, Madison DV, Shatz CJ. Enhancement of transmission at the developing retinogeniculate synapse. Neuron. 1993;10:815–825. doi: 10.1016/0896-6273(93)90198-z. [DOI] [PubMed] [Google Scholar]
- 51.Núñez-Abades PA, He F, Barrionuevo G, Cameron WE. Morphology of developing rat genioglossal motoneurons studied in vitro: changes in length, branching pattern, and spatial distribution of dendrites. J Comp Neurol. 1994;339:401–420. doi: 10.1002/cne.903390308. [DOI] [PubMed] [Google Scholar]
- 52.O’Brien RJ, Fishbach GD. Isolation of embryonic chick motoneurons and their survival in vitro. J Neurosci. 1986;6:3265–3274. doi: 10.1523/JNEUROSCI.06-11-03265.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.O’Dell TJ, Huang PL, Dawson TM, Dinerman JL, Snyder SH, Kandel ER, Fishman MC. Endothelial NOS and the blockade of LTP by NOS inhibitors in mice lacking neuronal NOS. Science. 1994;265:542–546. doi: 10.1126/science.7518615. [DOI] [PubMed] [Google Scholar]
- 54.O’Hanlon GM, Lowrie MB. The effects of neonatal dorsal root section on the survival and dendritic development of lumbar motoneurons in the rat. Eur J Neurosci. 1996;8:1072–1077. doi: 10.1111/j.1460-9568.1996.tb01274.x. [DOI] [PubMed] [Google Scholar]
- 55.Otmakhov N, Griffith LC, Lisman JE. Postsynaptic inhibitors of calcium/calmodulin-dependent protein kinase type II block induction but not maintenance of pairing-induced long-term potentiation. J Neurosci. 1997;17:5357–5365. doi: 10.1523/JNEUROSCI.17-14-05357.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Park J-H, Straub VA, O’Shea M. Anterograde signaling by nitric oxide: characterization and in vitro reconstitution of an identified nitrergic synapse. J Neurosci. 1998;18:5464–5476. doi: 10.1523/JNEUROSCI.18-14-05463.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Peunova N, Enikolopov G. Nitric oxide triggers a switch to growth arrest during differentiation of neuronal cells. Nature. 1995;375:68–73. doi: 10.1038/375068a0. [DOI] [PubMed] [Google Scholar]
- 58.Pockett S, Figurov A. Long-term potentiation and depression in the ventral horn of rat spinal cord in vitro. NeuroReport. 1993;4:97–99. doi: 10.1097/00001756-199301000-00025. [DOI] [PubMed] [Google Scholar]
- 59.Purves RI, Hume RI. The relation of post-synaptic geometry to the number of presynaptic axons that innervate autonomic ganglion cells. J Neurosci. 1981;1:441–452. doi: 10.1523/JNEUROSCI.01-05-00441.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Reid SNM, Daw NW, Czepita D, Flavin HJ, Sessa WC. Inhibition of nitric oxide synthase does not alter ocular dominance shifts in kitten visual cortex. J Physiol (Lond) 1996;494.2:511–517. doi: 10.1113/jphysiol.1996.sp021509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rentería RC, Constantine-Paton M. Exogenous nitric oxide causes collapse of retinal ganglion cell axonal growth cones in vitro. J Neurobiology. 1995;29:415–428. doi: 10.1002/(SICI)1097-4695(199604)29:4<415::AID-NEU1>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- 62.Roskams AJ, Bredt DS, Dawson TM, Ronnett GV. Nitric oxide mediates the formation of synaptic connections in developing and regenerating olfactory receptor neurons. Neuron. 1994;13:289–299. doi: 10.1016/0896-6273(94)90347-6. [DOI] [PubMed] [Google Scholar]
- 63.Ruthazer ES, Gillespie DC, Dawson TM, Snyder SH, Stryker MP. Inhibition of nitric oxide synthase does not prevent ocular dominance plasticity in kitten visual cortex. J Physiol (Lond) 1996;494.2:519–527. doi: 10.1113/jphysiol.1996.sp021510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schilling K, Dickinson MH, Connor JA, Morgan JI. Electrical activity in cerebellar cultures determines purkinje cell dendritic growth patterns. Neuron. 1991;7:891–902. doi: 10.1016/0896-6273(91)90335-w. [DOI] [PubMed] [Google Scholar]
- 65.Schuman EM, Madison DV. A requirement for the intercellular messenger nitric oxide in long-term potentiation. Science. 1993;254:1503–1506. doi: 10.1126/science.1720572. [DOI] [PubMed] [Google Scholar]
- 66.Segev I. Sound grounds for computing dendrites. Nature. 1998;393:207–208. doi: 10.1038/30340. [DOI] [PubMed] [Google Scholar]
- 67.Seil FJ, Drake-Baumann R. Reduced cortical inhibitory synaptogenesis in organotypic cerebellar cultures developing in the absence of neuronal activity. J Comp Neurol. 1994;342:366–377. doi: 10.1002/cne.903420305. [DOI] [PubMed] [Google Scholar]
- 68.Shatz CJ. Impulse activity and the patterning of connections during CNS development. Neuron. 1990;5:745–756. doi: 10.1016/0896-6273(90)90333-b. [DOI] [PubMed] [Google Scholar]
- 69.Shibuki K, Okada D. Endogenous nitric oxide release required for long-term synaptic depression in the cerebellum. Nature. 1991;349:326–328. doi: 10.1038/349326a0. [DOI] [PubMed] [Google Scholar]
- 70.Sholl DA. Dendritic organization in the neurons of the visual and motor cortices of the cat. J Anat (Lond) 1953;87:387–406. [PMC free article] [PubMed] [Google Scholar]
- 71.Silva AJ, Stevens CF, Tonegawa S, Wang Y. Deficient hippocampal long-term potentiation in α-calcium-calmodulin kinase II mutant mice. Science. 1992a;257:201–206. doi: 10.1126/science.1378648. [DOI] [PubMed] [Google Scholar]
- 72.Silva AJ, Stevens CF, Tonegawa S, Wang Y. Impaired spatial learning in α-calcium-calmodulin kinase II mutant mice. Science. 1992b;257:206–211. doi: 10.1126/science.1321493. [DOI] [PubMed] [Google Scholar]
- 73.Simon DK, Prusky GT, O’Leary DD, Constantine-Paton M. N-methyl-d-aspartate receptor antagonists disrupt the formation of a mammalian neural map. Proc Natl Acad Sci USA. 1992;89:10593–10597. doi: 10.1073/pnas.89.22.10593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Spitzer NC. Spontaneous Ca2+ spikes and waves in embryonic neurons: signaling systems for differentiation. Trends Neurosci. 1994;17:115–118. doi: 10.1016/0166-2236(94)90120-1. [DOI] [PubMed] [Google Scholar]
- 75.Spruston N, Schiller Y, Stuart G, Sakmann B. Activity-dependent action potential invasion and calcium influx into hippocampal CA1 dendrites. Science. 1995;268:297–300. doi: 10.1126/science.7716524. [DOI] [PubMed] [Google Scholar]
- 76.Stryker MP. Growth through learning. Nature. 1995;375:277–278. doi: 10.1038/375277a0. [DOI] [PubMed] [Google Scholar]
- 77.Vaughn JE, Barber RP, Sims TJ. Dendritic development and preferential growth into synaptic fields: a quantitative study of Golgi-impregnated spinal motor neurons. Synapse. 1988;2:69–78. doi: 10.1002/syn.890020110. [DOI] [PubMed] [Google Scholar]
- 78.Wang T, Xie Z, Lu B. Nitric oxide mediates activity-dependent synaptic suppression at developing neuromuscular synapses. Nature. 1995;374:262–266. doi: 10.1038/374262a0. [DOI] [PubMed] [Google Scholar]
- 79.Watanabe M, Mishina M, Inuoe Y. Distinct spatiotemporal distributions of the N-methyl-d-aspartate receptor channel subunit mRNAs in the mouse cervical cord. J Comp Neurol. 1994;345:314–319. doi: 10.1002/cne.903450212. [DOI] [PubMed] [Google Scholar]
- 80.Wishaw IQ, Auer RN. Immediate and long-lasting effects of MK-801 on motor activity, spatial navigation in a swimming pool and EEG in the rat. Psychopharmacology. 1989;98:500–507. doi: 10.1007/BF00441949. [DOI] [PubMed] [Google Scholar]
- 81.Wong EHF, Kemp JA, Priestley T, Knight AR, Woodruff G, Iversen LL. The anticonvulsant MK-801 is a potent N-methyl-d-aspartate antagonist. Proc Natl Acad Sci USA. 1986;83:7104–7108. doi: 10.1073/pnas.83.18.7104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wu HH, Williams CV, McLoon SC. Involvement of nitric oxide in the elimination of a transient retinotectal projection in development. Science. 1994;265:1593–1596. doi: 10.1126/science.7521541. [DOI] [PubMed] [Google Scholar]
- 83.Wu G-Y, Cline HT. Stabilization of dendritic arbor structure in vivo by CaMKII. Science. 1998;279:222–226. doi: 10.1126/science.279.5348.222. [DOI] [PubMed] [Google Scholar]
- 84.Yen L-H, Sibley JT, Constantine-Paton M. Fine-structural alterations and clustering of developing synapses after chronic treatments low levels of NMDA. J Neurosci. 1993;13:4949–4960. doi: 10.1523/JNEUROSCI.13-11-04949.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yen L, Sibley JT, Constantine-Paton M. Analysis of synaptic distribution within single retinal axonal arbors after chronic NMDA treatment. J Neurosci. 1995;15:4712–4725. doi: 10.1523/JNEUROSCI.15-06-04712.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]