

Abstract
Background/Aims
The aim of the present study was to determine the changes on the small intestine in mice during pregnancy using histological, enzyme histochemical, and immunohistochemical methods.
Materials and Methods
A total of 24 Swiss albino female mice were divided as non-pregnant/control, first week, second week, and third week of pregnancy (n=6). Tissue samples obtained from the duodenum, jejunum, and ileum were processed by means of routine histological techniques and stained with Crossmon’s triple staining. Alkaline phosphatase (ALP) was demonstrated with the simultaneous azo-coupling method. Proliferating cell nuclear antigen (PCNA) was demonstrated with the streptavidin-biotin-peroxidase complex method. The numerical data of the parameters were obtained and analyzed statistically.
Results
Villus height, villus width, and the rate of villus height/crypt depth were decreased in the duodenum, jejunum, and ileum in the last week of pregnancy compared with the control group. Changes in the crypt depth of the duodenum, jejunum, and ileum in pregnancy were found. The muscle width increased in pregnancy. It was identified that the ALP reactivity statistically significantly increased in the duodenum, jejunum, and ileum in pregnancy. The percentage of PCNA-positive cells in the duodenum, jejunum, and ileum increased in the first and second weeks of pregnancy, whereas it decreased in the third week of pregnancy compared with non-pregnant control animals.
Conclusion
In conclusion, villus parameters, ALP reactivity, and percentage of PCNA-positive cells in the small intestine were affected during pregnancy.
Keywords: ALP, mice, PCNA, pregnancy, villus
INTRODUCTION
Pregnancy is a process influencing other systems, together with the digestive system. Some changes occur in the digestive system organs, such as the stomach, intestines, pancreas, and liver, during pregnancy (1–4). Pregnancy is characterized by structural and functional changes in some organs and regulated by the estrogen and progesterone hormones (5). In this process, an increase in progesterone levels is responsible for the changes observed in the digestive system. Progesterone loosens up the smooth muscle in the digestive system and slows down the digestion. During pregnancy, morphological and histological changes are observed in the small intestine. These changes are characterized by an increase in parameters, such as the length of the small intestine and the height of the villus depending on mother’s consuming and her general physiological condition (6). The length of the small intestine and the height of the villus are at maximum level in rats giving multiple births and at minimum level in rats giving no births (5).
Alkaline phosphatase (ALP) is activated by magnesium ions and shows maximum activity at a pH of approximately 10 (7). The main sources of ALP, an enzyme having the structure of a glycoprotein, are the liver, small intestine, pancreas, bone, thyroid gland, placenta, tooth enamel, and testis (8–12). Intestinal ALP localized in the brush border membrane plays an important role in the absorption of cholesterol, lipid, vitamin D, calcium, amino acids, and glucose (13). Additionally, intestinal ALP detoxifies a variety of bacterial pro-inflammatory factors and functions to preserve gut barrier function (14). ALP activity, which is the most stable, has been accepted as a signal of the condition of the mucosal epithelial surface (15,16). During pregnancy, ALP is involved in cytotrophoblast development, is present in the placental and maternal circulation, and serves as the source of food for the fetus in the endometrium (17,18). The ALP enzyme is responsible for regulating the high proliferation potential of cytotrophoblastic cells in the early stages of pregnancy, together with many growth factors, such as epidermal growth factor and insulin-like growth factor 1 (19). This enzyme is more concerned with maternal circulation than with fetal circulation. A slight increase in the level of serum ALP is noted in the third trimester of pregnancy. It has been thought that ALP serves the function of supplying the maximum needs of the fetus (12,20). ALP completely disappeared in the placenta brush border membrane during miscarriage (21,22).
Proliferating cell nuclear antigen (PCNA) is a processing factor for DNA polymerase delta auxiliary protein and plays essential roles in the replication and repair of damaged DNA (23–25). The rate of PCNA release increases rapidly from the middle of the G1 phase of the cell cycle. It maintains a high level through the S phase and starts decreasing in the G2/M phase (26,27). Therefore, the PCNA is used as an indicator for proliferating cells (28–30). PCNA immunohistochemistry has been used as a potential tool for the study of proliferative activity of tissues (31). PCNA is expressed with high levels in nearly all proliferating tissues, such as thymus, bone marrow, fetal liver, certain cells of the small intestine, and colon (32).
The pregnancy period in humans is approximately 270 days which occurs in three trimesters, whereas this period in mice is approximately 21 days. In humans, the first trimester (0–90 days) of pregnancy corresponds to 0.5–10.5 days in mice, whereas the 2nd trimester (91–180 days) and 3rd trimester (181–270 days) of pregnancy correspond to 11–21 days in mice (33). Despite some differences in pregnancy physiology, mice have been widely used for studying human pregnancy disorders.
The aim of the present study was to determine the changes in the small intestinal tissue in mice at different stages of pregnancy with histological, enzyme histochemical, and immunohistochemical methods.
MATERIALS AND METHODS
Study groups design
A total of 24 female Swiss albino mice 12–14 weeks of age and weighing 20–25 g were used in the present study. During the study, the animals were fed daily without restriction of forage and water under standard conditions (room temperature 20°C±1°C, relative humidity 50%±10%, and 12/12-hour light-dark period). All procedures were approved by the ethical committee of Selçuk University School of Medicine, Konya, Turkey (2009/32). The female mice leaving for mating at night were controlled daily with respect to vaginal plug formation. The mice whose vaginal plug formation was completed were accepted on day 0 of their pregnancies. Mice were divided into four groups as non-pregnant/control, first week (on day 3 of early pregnancy which occurs after implantation), second week (on day 10 of midpregnancy which occurs in decidualization), and third week (on day 17 of late pregnancy which is equivalent to the third trimester in human pregnancy) of pregnancy (n=6 for each group). The mice were weighed and sacrificed after anesthetizing by ether inhalation. Then, small intestinal tissue samples (proximal portion of the duodenum and middle portion of both the jejunum and ileum) were obtained from the sacrificed animals.
Histological procedure
The samples were fixed in 10% buffered-formal saline (pH 7.4) for 24 h at room temperature. Then, the samples were dehydrated, cleared, and embedded in paraffin. Sections of 6 μm thickness were stained using the Crossmon’s triple staining technique for determining the general histological structure and for histomorphologic analysis of the villus (villus height, villus width, crypt depth, villus height/crypt depth, and muscle width) (34). For ALP demonstration, samples were fixed in a formal-calcium solution (+4°C) for 24 h and then kept in Holt’s solution (+4°C) for an additional 24 h. Enzyme histochemical reactions were ascertained on 12 μm frozen sections (Leica, Germany). The demonstration of ALP was assessed using the simultaneous azo-coupling method (35). All specimens were examined using a light microscope with a digital camera (Nikon Eclipse, E-400 equipped with Nikon DS Camera Control Unit DS-L1 and DS Camera Head DS-5M; Nikon, Japan), and the digital images of necessary areas were saved. The images were analyzed by the digital imaging analysis program (BS200 PRO, 2005) using the measured color intensity, and the numerical data of parameters were obtained.
The PCNA protein activity was assessed using the immunohistochemical method to determine the rate of cell proliferation (36). The immunohistochemical staining of tissue sections was performed using the streptavidin-biotin-peroxidase complex procedure. The sections were deparaffinized in xylene series and rehydrated. The sections were placed into citrate buffer (pH 6) and heated in a microwave oven (700 W for 5 min) to unmask the antigen. The sections were left for 20 min in 3% hydrogen peroxide solution for the inhibition of the activation of endogenous peroxidase. Non-specific binding sites were blocked by incubating the sections in blocking solutions. The sections were incubated with mouse anti-PCNA monoclonal antibody (GeneTex GTX71945, 1:100 dilution) and then with biotinylated goat anti-mouse secondary antibody (IgG) (ScyTek UHP 125, USA) for 20 min. Next, they were incubated with horseradish peroxidase-streptavidin (ScyTek UHP 125, USA) at room temperature for 20 min. Color reaction was developed with 3,3′-diaminobenzidine (ScyTek ACK 125, USA). The slides were counterstained with Mayer’s hematoxylin and then mounted in synthetic resin (Entellan, Merck, Darmstadt, Germany). PCNA-positive cells were counted in a total of 100 cells in crypts, and positivity percentage (%) was determined.
Statistical analysis
Data were analyzed using Statistical Package for Social Sciences 10 program (SPSS Inc.; Chicago, IL, USA). One-way analysis of variance test, followed by post hoc Duncan multiple comparison tests, was used to evaluate data. The significant levels of the differences among the average values of different groups were determined. A p<0.05 was considered significant.
RESULTS
Body weights and intestine weights
Mean body weights and relative intestine weights during different periods of pregnancy are shown in Table 1.
Table 1.
Mean body weights (g) and relative intestine weights (%) during different periods of pregnancy.
| Groups (n=6) | Body weight | Relative intestine weight |
|---|---|---|
| Control | 23.06±0.47c | 11.77±0.61 |
| First week | 23.13±0.44c | 12.64±0.56 |
| Second week | 28.89±2.17b | 12.66±1.23 |
| Third week | 39.58±1.66a | 14.65±0.59 |
Differences between mean values written in the same column with different letters are significant (p<0.05).
A significant increase was observed in the mean values of body weights during pregnancy compared with the control group (p<0.05). Although an increase was also observed in the mean relative intestine weights, it was not statistically significant (p>0.05).
Villus parameters
Measurement results obtained from the duodenal tissue during different periods of pregnancy are given in Table 2.
Table 2.
Data obtained from the duodenum during different periods of pregnancy (μm) (X±SE).
| Groups (n=6) | Villus height | Villus width | Crypt depth | Muscle width | Villus height/crypt depth |
|---|---|---|---|---|---|
| Control | 467.82±35.66 | 92.85±2.00 | 112.200±5.72c | 23.81±1.59c | 4.22±0.32a |
| First week | 501.68±8.87 | 89.63±8.60 | 126.81±4.81bc | 27.53±1.76bc | 3.84±0.25ab |
| Second week | 414.11±26.13 | 100.30±4.89 | 148.31±4.44a | 38.18±1.80a | 2.51±0.20c |
| Third week | 399.61±33.30 | 98.31±8.09 | 134.26±9.43ab | 32.79±2.79ab | 3.22±0.40bc |
Differences between mean values written in the same column with different letters are significant (p<0.05).
Although an increase was observed in the villus height values in the first week of pregnancy compared with the control group, a decrease was noted in the second and third weeks of pregnancy. However, this change was not statistically significant (p>0.05). The ratio of villus height/crypt depth also decreased gradually in the pregnancy groups compared with the control group (p<0.05).
Data obtained from the jejunum are given in Table 3.
Table 3.
Data obtained from the jejunum during different periods of pregnancy (μm) (X±SE).
| Groups (n=6) | Villus height | Villus width | Crypt depth | Muscle width | Villus height/crypt depth |
|---|---|---|---|---|---|
| Control | 400±36.13b | 84.20±4.86 | 132.44±10.44 | 27.43±1.85c | 3.07±0.18 |
| First week | 365.79±19.55b | 81.90±7.25 | 145.37±14.17 | 49.40±1.70a | 2.66±0.34 |
| Second week | 474.55±20.08a | 89.11±4.42 | 141.61± 8.32 | 41.62±2.90b | 3.20±0.13 |
| Third week | 309.77±27.48b | 69.63±5.45 | 133.59± 8.96 | 40.01±3.03b | 2.38±0.29 |
Differences between mean values written in the same column with different letters are significant (p<0.05).
It was observed that villus height decreased in the first week of pregnancy, increased in the second week of pregnancy, and was at the lowest level in the third week of pregnancy (p<0.05). Furthermore, the crypt depth values increased in the first and second weeks of pregnancy and decreased in the third week of pregnancy compared with the control group. However, the difference was not statistically significant (p>0.05). The muscle width values showed a statistically significant increase in the pregnancy groups compared with the control group (p<0.05). No statistically significant difference was found in the ratio of villus height/crypt depth (p>0.05).
Data obtained from the ileal tissue during different periods of pregnancy are given in Table 4.
Table 4.
Data obtained from the ileum tissue during different periods of pregnancy (μm) (X±SE).
| Groups (n=6) | Villus height | Villus width | Crypt depth | Muscle width | Villus height/crypt depth |
|---|---|---|---|---|---|
| Control | 281.24±34.50a | 80.50±4.03ab | 140.42±5.58b | 30.57±3.01c | 2.32±0.27 |
| First week | 301.98±8.56a | 90.35±7.07a | 141.70±7.86b | 39.41±2.12b | 2.15±0.13 |
| Second week | 255.76±7.87a | 80.26±4.27ab | 165.66±3.90a | 55.58±2.31a | 1.83±0.14 |
| Third week | 181.23±4.00b | 63.55±6.02b | 108.67±7.75c | 33.23±3.62bc | 1.70±0.11 |
Differences between mean values written in the same column with different letters are significant (p<0.05).
It was observed that the villus height was at the lowest level in the third week of pregnancy compared with the control group and the first and second weeks of pregnancy (p<0.05). In addition, the villus width values were similar to the villus height values (p<0.05). The crypt depth increased in the second week of pregnancy compared with the control group and first week and decreased in the third week of pregnancy (p<0.05). It was found that the muscle width was at the highest level in the second week of pregnancy compared with the other groups (p<0.05). A decrease was observed in the ratio of villus height/crypt depth in the pregnancy groups compared with the control group; however, it was not statistically significant (p>0.05).
ALP reactivity
Alkaline phosphatase reactivity was observed on the enterocyte brush border membrane. ALP reactivity was not seen in the crypt epithelium. The ALP reactivity determined during different periods of pregnancy is given in Table 5.
Table 5.
ALP reactivity determined during different periods of pregnancy (%).
| Groups (n=6) | Duodenum | Jejunum | Ileum |
|---|---|---|---|
| Control | 5.24±0.20c | 4.20±0.54c | 4.10±0.21c |
| First week | 6.61±0.84bc | 6.83±0.54b | 6.85±0.33b |
| Second week | 8.19±0.39ab | 8.58±0.33a | 8.22±0.46a |
| Third week | 9.61±0.60a | 8.61±0.27a | 8.81±0.44a |
Differences between mean values written in the same column with different letters are significant (p<0.05).
A gradual increase was found in the ALP reactivity on the brush border membrane of the duodenum, jejunum, and ileum during pregnancy compared with the control group (p<0.05) (Figure 1).
Figure 1. A–D.
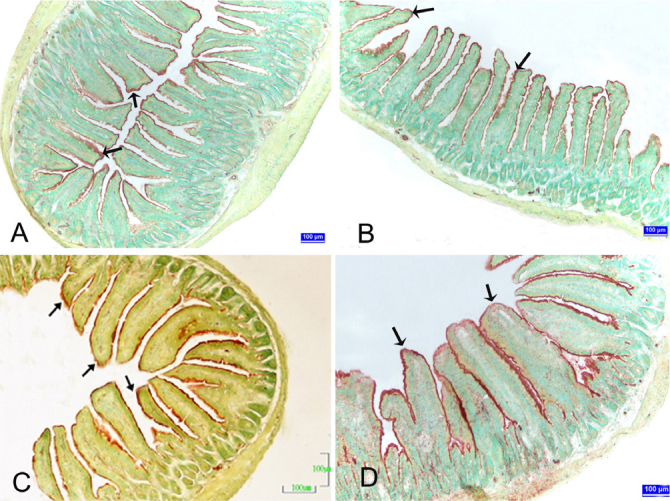
Jejunum sections of the control and pregnant groups. (A) Control group, (B) first week of pregnancy, (C) second week of pregnancy, (D) third week of pregnancy Arrows: ALP reactivity. ALP demonstration. Bar: 100 μm.
PCNA immunohistochemistry
The cells with a brown-stained nucleus were evaluated as PCNA-positive cells. The percentages of PCNA-positive cells determined during different periods of pregnancy are given in Table 6.
Table 6.
PCNA-positive cell percentages of the different periods of pregnancy (%).
| Groups (n=6) | Duodenum | Jejunum | Ileum |
|---|---|---|---|
| Control | 55.37±3.10b | 47.70±2.82b | 50.57±3.73c |
| First week | 65.17±0.91a | 64.92±1.29a | 59.26±1.73ba |
| Second week | 66.71±1.14a | 68.61±1.39a | 62.55±1.45a |
| Third week | 50.78±2.93b | 53.37±2.59b | 52.79±1.75bc |
Differences between mean values written in the same column with different letters are significant (p<0.05).
The percentage of PCNA-positive cells increased in the first and second weeks of pregnancy, whereas it decreased in the third week of pregnancy compared with the control group in all sections of the small intestine (Figure 2, p<0.05).
Figure 2. A–D.
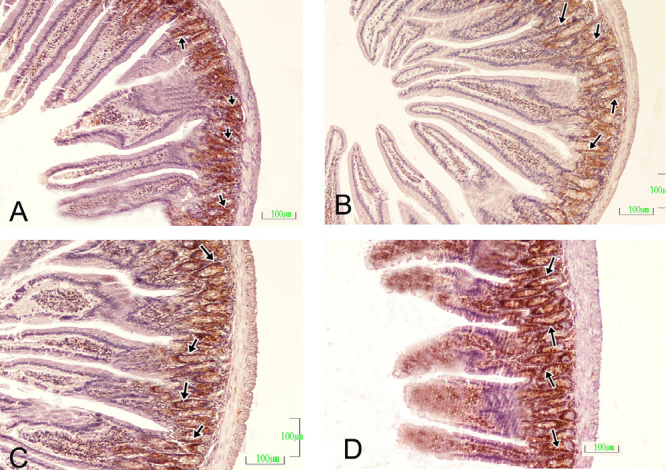
Ileum sections of the control and pregnant groups. (A) Control group, (B) first week of pregnancy, (C) second week of pregnancy, (D) third week of pregnancy Arrows: PCNA-positive cells. PCNA immunohistochemical staining. Bar: 100 μm.
DISCUSSION
Food intake increases in females because of increasing energy needs during pregnancy. Changes are seen in the body systems, particularly the digestive system and structures of the organs (37). During the pregnancy and lactation periods, some certain metabolic diseases and gastrointestinal tract problems have more serious clinical importance. In addition, some histological changes during pregnancy have been reported in the liver including increased cytoplasmic fat vacuoles in centrilobular hepatocytes and Kupffer cell hypertrophy. Liver adenomas may enlarge during pregnancy in humans (38). A gradual increase in basal insulin concentration with the progression of pregnancy has been reported (39,40). Nalbant observed hyperplasia in pancreatic beta cells and increased insulin secretion during pregnancy (2). In addition, there are many studies about the structure of the small intestine flora changes in the literature (3,41,42).
There are some clinical symptoms related with the gastrointestinal system and digestive system organs. An increase in progesterone levels reduces motility and causes an increase in gastric acid secretion in the gastrointestinal system (43). Increases in live body weight, food intake, anatomical dimensions of the gastrointestinal system, and absorption capacity of the small intestine were found in albino rats during pregnancy and lactation (44). Food intake increased 60% during pregnancy and 250% during lactation (45). Statistically significant increases were found in body weights of rats during pregnancy and lactation (46). A 40% increase was reported in the small intestine weights of rats in the last week of pregnancy and the first week of lactation (47). Different investigators reported a 50% increase in the small intestine weights of rats during pregnancy and lactation (48). The change found in the mean body weights of rats in the present study was compatible with the other researchers’ findings. Although a statistically significant increase was observed in the mean body weights compared with the control group during the mid- and late gestational periods (p<0.05), it might be related to the developing fetuses. In addition, an increase was observed in the mean relative intestine weights during pregnancy compared with the control group, and this increase was not statistically significant (p>0.05).
An increase was observed in the villus heights in albino rats during pregnancy (46,47). The villus heights of pregnant mouse were found to be more than 50% than those of non-pregnant mouse (49). An increase was reported in the villus heights in rats during pregnancy and lactation (37). An increase was found in the villus heights of the small intestine and the total villus area in rats during pregnancy and lactation (37). An increase was demonstrated in the villus height and the area of the jejunum in rats during pregnancy, and this increase reached maximum level during lactation (47). However, there were studies which showed that the villus height decreased in pregnant rats (50,51). In the present study, it was found that the villus height of the duodenum increased in the first week of pregnancy and decreased in the second and third weeks of pregnancy compared with the control group (p>0.05). In addition, the villus height of the jejunum decreased in the first week of pregnancy, increased in the second week, and was at the lowest level in the third week of pregnancy (p<0.05). The villus height of the ileum decreased in the third week of pregnancy compared with the control group in the first and second weeks of pregnancy (p<0.05). The reason of this difference might be the ages of the animals, number of offsprings, pregnancy number of animals, and sampling method.
Intestinal ALP activity is accepted as a marker of the functional status of the small intestine epithelium, because it is quite stable (10,52). The ALP level in the mammary tissue of pregnant rats reached maximum level at birth by increasing rapidly during pregnancy and remained at the same level during lactation (52). ALP increased in the pregnant horn from days 5 to 8, whereas it remained unaltered in the non-pregnant horn (53). The relationship between ALP value and uterine weight in rats during pregnancy showed that ALP activity declined on days 1 and 3 of pregnancy, started to increase on day 7, reached maximum value on day 14, and maintained its high level until birth (54). Activities and changes in ALP isoenzymes (bone, liver, intestine, and placental ALP) during pregnancy in humans showed an increase in the levels of all ALP isoenzymes; the increase in ALP in the placenta was more than the increase in other locations (55). The present study found that the intensity of relative ALP reactivity observed in the duodenum, jejunum, and ileum gradually increased during pregnancy compared with the control group (p<0.05). These findings were consistent with the findings of other researchers and showed that ALP release began during early pregnancy and its level increased in time.
PCNA has been used as a marker for proliferating cells. The level of PCNA in the placental tissue in humans during early pregnancy was at maximum level and decreased in parallel with placental development (56). The levels of placental PCNA increased during pregnancy in humans with preeclampsia and in normal pregnancy (57). However, the number of PCNA-positive trophoblast cells declined during later pregnancy in some studies performed on rats and mice (58,59). PCNA immunoreactivity declined from the second day of pregnancy and disappeared from the fourth day of pregnancy in the uterine luminal epithelium and glandular epithelium during early pregnancy in rats (60). During a healthy pregnancy, PCNA is expressed at the highest level in the early period (4–5 weeks) in cytotrophoblastic cell nuclei, and the level of PCNA is decreased progressively in the late period (19,61). Acar et al. (62) stated that while PCNA is highest on days 11 and 13, it decreases with progression of pregnancy in rats. Ozaydın et al. (63) reported that the percentage of PCNA positivity in trophoblast and decidual cells increases at the mid-gestational stage and decreases at the late stage in mouse. PCNA-positive cell numbers in the small intestine crypts increased during lactation despite a decline during pregnancy in non-pregnant, 3-week pregnant, and lactating rats (37). The present study found that the percentages of PCNA-positive cells in the duodenum, jejunum, and ileum increased in the first and second weeks of pregnancy and then decreased in the third week of pregnancy compared with the control group (p<0.05).
The changes in the small intestine during pregnancy occur according to the needs of the developing fetus. Investigating the changes in the small intestine during different periods of pregnancy using histological, enzyme histochemical, and immunohistochemical methods can help in diagnosing and treating the complications that may occur in the digestive system disorders during pregnancy. Although there are some differences in pregnancy physiology between mouse and human, animal studies are critical for understanding the mechanisms of gastrointestinal discomfort of pregnancy. However, further studies are needed to evaluate gastrointestinal motility and some certain blood parameters, in addition to histological findings, to improve the clinical approach of pregnant women suffering from gastrointestinal dysfunction.
Footnotes
Ethics Committee Approval: Ethics committee approval was received for this study from the Ethics Committee of Selcuk University Experimental Medical Research and Application Center Experimental Animal Ethics Committee (2009/32).
Informed Consent: N/A.
Peer-review: Externally peer-reviewed.
Author Contributions: Concept - E.Ş., Y.Ö; Design - E.Ş., Y.Ö; Supervision - E.Ş., Y.Ö; Resources - E.Ş., Y.Ö; Materials - E.Ş., Y.Ö; Data Collection and/or Processing - E.Ş., Y.Ö; Analysis and/or Interpretation - E.Ş., Y.Ö; Literature Review - E.Ş., Y.Ö; Writer - E.Ş., Y.Ö; Critical Review - E.Ş., Y.Ö.
Conflict of Interest: The authors have no conflict of interest to declare.
Financial Disclosure: Provision of materials of our study has been afforded by Selçuk University Scientific Research Projects (BAP) Coordinating Office (Project No: 09202056).
REFERENCES
- 1.Ch’ng CL, Morgan M, Hainsworth I, Kingham JG. Prospective study of liver dysfunction in pregnancy in Southwest Wales. Gut. 2002;51:876–80. doi: 10.1136/gut.51.6.876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nalbant S. Gebelikte gelişen fizyolojik değişiklikler. Onuncu Ulusal İç Hastalıkları Kongresi Antalya. 2008:39–40. [Google Scholar]
- 3.Chung SY, Ravel J, Regan M. Clinical relevance of gastrointestinal microbiota during pregnancy: A primer for nurses. Biol Res Nurs. 2018;20:84–102. doi: 10.1177/1099800417732412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sur E, Oznurlu Y, Ozaydın T. The determination of Ghrelin immunoreactivity in gastric mucosa of fundus during pregnancy in mice. Int J Biomed Res. 2017;8:672–6. [Google Scholar]
- 5.Casirola DM, Ferraris RP. Role of the small intestine in postpartum weight retention in mice. Am J Clin Nutr. 2003;78:1178–87. doi: 10.1093/ajcn/78.6.1178. [DOI] [PubMed] [Google Scholar]
- 6.Meyer AM, Caton JS. Role of the small ıntestine in developmental programming: Impact of maternal nutrition on the dam and offspring. Adv Nutr January. 2016;7:169–78. doi: 10.3945/an.115.010405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kumar V, Gill KD. To estimate the activity of alkaline phosphatase in serum in basic concepts in clinical biochemistry: A practical guide. Springer; Singapore: 2018. pp. 107–9. [DOI] [Google Scholar]
- 8.Sakharov IYU, Makasova I, Ermolin GA. Purification and characterisation of intestinal alkaline phosphatase from harp seal comp. Biochem Physiol. 1988;90:709–14. doi: 10.1016/0305-0491(88)90324-0. [DOI] [PubMed] [Google Scholar]
- 9.Klumpp S, Schultz JE. Alkaline phosphatase from pramercium cilia and cell bodies purification and characterisation. Biochem Biophys Acta. 1990;1037:233–9. doi: 10.1016/0167-4838(90)90173-D. [DOI] [PubMed] [Google Scholar]
- 10.Kumandaş S, Kurtoğlu S. Çocukluk döneminde alkalen fosfataz enziminin değerlendirilmesi. Yeni Tıp Drg. 1992;9:68–70. [Google Scholar]
- 11.Kaplan LA, Pesce AJ. Clinical chemistry theory analysis and correlation. Mosby Third Edition Baltimore Boston Chicago Newyork London Madrid Mexico City Singapore Tokyo Toronto. 1996:514–5. [Google Scholar]
- 12.Uzunoğlu N. Alkalen fosfataz enziminin fizikokimyasal özellikleri. T Klin J Med Sci. 1998;18:69–75. [Google Scholar]
- 13.Uysal O. İnsan plasental alkalen fosfatazın histokimyasal lokalizasyonu. Ankara Ün Dikimevi MYO Drg. 2003;4:13–21. doi: 10.1501/Ashd_0000000042. [DOI] [Google Scholar]
- 14.Hamarneh SR, Kim BM, Kaliannan K, Morrison SA. Intestinal alkaline phosphatase attenuates alcohol-induced hepatosteatosis in mice. Dig Dis Sci. 2017;62:2021–34. doi: 10.1007/s10620-017-4576-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gu X, Li D, She R. Effect of weaning on small intestinal structure and function in the piglet. Arch Anim Nutr. 2002;56:275–86. doi: 10.1080/00039420214345. [DOI] [PubMed] [Google Scholar]
- 16.Özaydın T. Kuluçkada deneysel olarak oluşturulan ısı stresinin boylerlerde ince bağırsağın embriyonik gelişimi üzerindeki etkilerinin histokimyasal, immünohistokimyasal ve histometrik metotlarla belirlenmesi. Selçuk Ün Sağlık Bil Ens Dr Tezi Konya. 2009 [Google Scholar]
- 17.Posen S. Alkaline phosphate. Ann Int Med. 1967;67:183–203. doi: 10.7326/0003-4819-67-1-183. [DOI] [PubMed] [Google Scholar]
- 18.Shane JM, Suzuki K. Placenta alkaline phosphate. A review and re-evaluation of its applicability in monitoring fetoplacental function. Obst Gynec Survey. 1974;29:97–105. doi: 10.1097/00006254-197402000-00001. [DOI] [PubMed] [Google Scholar]
- 19.Ishıhara N, Matsuo H, Murakoshı H, Laoag-Fernandez J, Samoto T, Maruo T. Changes in proliferative potential, apoptosis and Bcl-2 protein expression in cytotrophoblasts and syncytiotrophoblast in human placenta over the course of pregnancy. Endocr J. 2000;47:317–27. doi: 10.1507/endocrj.47.317. [DOI] [PubMed] [Google Scholar]
- 20.Okamoto T, Seo H, Mano H, et al. Expression of human placental alkaline phosphatase in placenta during pregnanc. Placenta. 1990;2:319–27. doi: 10.1016/S0143-4004(05)80223-1. [DOI] [PubMed] [Google Scholar]
- 21.Levine B. The early trophoblast a review including theoretical considerations. Obst Gynec. 1961;17:769–78. [PubMed] [Google Scholar]
- 22.Jeacock MK, Monis NF, Plester JA. The activity of alkaline and acid phospatase in the human placenta. J Obst Gynaeo Brit. 1963;70:267–73. doi: 10.1111/j.1471-0528.1963.tb04214.x. [DOI] [Google Scholar]
- 23.Dong Y, Liu G, Wang Z, Li J, Cao J, Chen Y. Effects of catecholaminergic nerve lesion on endometrial development during early pregnancy in mice. Histol Histopathol. 2015;11:1–23. doi: 10.14670/HH-11-684. [DOI] [PubMed] [Google Scholar]
- 24.Hall P, Levison D, Woods A. Proliferating cell nuclear antigen PCNA immunolocalization in parafin sections an index of cell proliferation with evidence of deregulated expression in some neoplasms. J Pathol. 1990;62:285–94. doi: 10.1002/path.1711620403. [DOI] [PubMed] [Google Scholar]
- 25.Soyuer I, Canöz Ö, Er Ö, Deniz K, Soyuer S. Malign mezotelyomada (proliferating nuclear cell antijen) PCNA immünreaktivitesinin ve mitotik indeksin prognoza etkisi. Erciyes Ün Tıp Fk Drg. 2002;24:115–9. [Google Scholar]
- 26.Maga G, Hübscher U. Proliferating cell nuclear antigen-PCNA a dancer with many partners. Journ of Cell Sci. 2003;116:3051–60. doi: 10.1242/jcs.00653. [DOI] [PubMed] [Google Scholar]
- 27.Kelman Z. PCNA: Structure, function and interactions. Oncogene. 1997;14:629–40. doi: 10.1038/sj.onc.1200886. [DOI] [PubMed] [Google Scholar]
- 28.Foley JF, Dietrich DR, Swenberg JA, Maronpot RR. Detection and evaluation of proliferating cell nuclear antigen PCNA in rat tissue by an improved immunohistochemical procedure. J Histotech. 1991;14:237–41. doi: 10.1179/his.1991.14.4.237. [DOI] [Google Scholar]
- 29.Şen O, Kayaselçuk F, Zorludemir S, Aydın MV, Erdogan B. Meningiomlarda histopatolojik tanının flovsitometrik DNA analizi, PCNA ve KI-67 ile korelasyonu. Türk Nöroşürurji Drg. 2002;12:48–53. [Google Scholar]
- 30.Özaydın T, Çelik İ. Histological, histochemical and immunohistochemical investigations on the developing small intestines of broiler embryos. Journ of Ani and Vet Adv. 2012;11:2936–44. doi: 10.3923/javaa.2012.2936.2944. [DOI] [Google Scholar]
- 31.Banlunara W, Bintvihok A, Kumagai S. Immunohistochemical study of proliferating cell nuclear antigen (PCNA) in duckling liver fed with aflatoxin B1 and esterified glucomannan. Toxicon. 2005;46:954–7. doi: 10.1016/j.toxicon.2005.04.019. [DOI] [PubMed] [Google Scholar]
- 32.Stoimenov I, Helleday T. Atlas of genetics and cytogenetics in oncology and haematology. Vol. 106. Department of Genetics Microbiology, Toxicology, Stockholm University, Stockholm, Sweden (IS, TH); Gray Institute for Radiation Oncology & Biology, University of Oxford; Oxford: 2011. p. 91. [Google Scholar]
- 33.Sones JL, Davisson RL. Preeclampsia, of mice and women. Physiol Genomics. 2016;48:565–72. doi: 10.1152/physiolgenomics.00125.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Culling FA, Allison RT, Barr WT. Cellular pathology technique. Butterworts and Co Ltd; London: 1985. pp. 25–9. [DOI] [Google Scholar]
- 35.Lojda Z, Grossrau R, Schibler TH. Enzyme histochemistry. Springer-Verlag; Berlin: 1979. pp. 59–70. [DOI] [Google Scholar]
- 36.Uni Z, Geyra A, Ben-Hur H, Sklan D. Small intestinal development in the young chick crypt formation and enterocyte proliferation and migration. Br Poult Sci. 2000;41:544–51. doi: 10.1080/00071660020009054. [DOI] [PubMed] [Google Scholar]
- 37.Mohammed SA, Ibrahim SH. Histological and immunohistochemical study on the effect of pregnancy and lactation on the jejunal mucosa of albino rat. J Egypt Hist. 2004;27:375–88. [Google Scholar]
- 38.Van Dyke RW. The liver in pregnancy. In: Boyer TD, Wright Tl, editors. Zakim and Boyer’s Hepatology: A textbook of liver disease. 5th ed. USA: Sounders Elseiver; 2006. pp. 1003–29. [DOI] [Google Scholar]
- 39.Solomon EP, Berg LR, Martin DW. Biology. Saunders Collage Publishing, electronic resource. 9th ed. Vol. 32. Belmont. CA: Brooks/Cole; 2011. pp. 48–52. [Google Scholar]
- 40.Erhan F, Ergün L. Kanatlı ve memeli karaciğerinde karbonhidrat ve yağ metabolizmasının karşılaştırılması. MAKÜ Sağ Bil Enst Derg. 2018;6:33–42. doi: 10.24998/maeusabed.356417. [DOI] [Google Scholar]
- 41.Collado MC, Isolauri E, Laitinen K, Salminen S. Distinct composition of gut microbiota during pregnancy in overweight and normal weight women. Am J Clin Nutr. 2008;88:894–9. doi: 10.1093/ajcn/88.4.894. [DOI] [PubMed] [Google Scholar]
- 42.Gohir W, Whelan FJ, Surette MG, Moore C, Jonathan D. Pregnancy-related changes in the maternal gut microbiota are dependent upon the mother’s periconceptional diet. Gut Microbes. 2015;6:310–20. doi: 10.1080/19490976.2015.1086056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Güven S, Türkay C. Gebelik ve karaciğer hastalıkları. Güncel Gastroenteroloji. 2011;15:107–13. [Google Scholar]
- 44.Cripps AW, Williams VJ. The effect of pregnancy and lactation on food intake, gastrointestinal anatomy and the absorptive capacity of the small intestine in the albino rat. Br J Nutr. 1975;33:17–32. doi: 10.1079/BJN19750005. [DOI] [PubMed] [Google Scholar]
- 45.Williamson DH. Regulation of metabolism during lactation in the rat. Reprod Nutr Develop. 1986;26:597–603. doi: 10.1051/rnd:19860409. [DOI] [PubMed] [Google Scholar]
- 46.Boass A, Lovdal JA, Toverud SU. Pregnancy-and lactation-induced changes in active intestinal calcium transport in rats. Am J Gastrointest Liver Physiol. 1992;263:127–34. doi: 10.1152/ajpgi.1992.263.1.G127. [DOI] [PubMed] [Google Scholar]
- 47.Burdett K, Reek C. Adaptation of the small intestine during pregnancy and lactation in the rat. Biochem J. 1979;184:245–51. doi: 10.1042/bj1840245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Clarke RM. The effects of age on mucosal morphology and epithelial cell production in rat small intestine. J Anat. 1977;123:805–11. [PMC free article] [PubMed] [Google Scholar]
- 49.Harmatz P, Carrington P, Barry V, Hatz R, Bloch K. Intestinal adaptation during lactation in the mouse altered intestinal processing of a dietary protein. Am J Phys. 1993;264:1126–34. doi: 10.1152/ajpgi.1993.264.6.G1126. [DOI] [PubMed] [Google Scholar]
- 50.Cairnie AB, Bentley RE. Cell proliferation studies in the intestinal epithelium of the rat: Hyperplasia during lactation. Exp Cell Res. 1967;46:428–40. doi: 10.1016/0014-4827(67)90079-1. [DOI] [PubMed] [Google Scholar]
- 51.Firmansyah A, Suwandito L, Penn D, Lebenthal E. Biochemical and morphological changes in the digestive tract of rats after prenatal and postnatal malnutrition. Am J Clin Nutr. 1989;50:216–8. doi: 10.1093/ajcn/50.2.261. [DOI] [PubMed] [Google Scholar]
- 52.Folley SJ, Greenbaum AL. Changes in the arginase and alkaline phosphatase contents of the mammary gland and liver of the rat during pregnancy, lactation and mammary involution. Nat Inst for Res in Dairying. 1946;41:261–8. doi: 10.1042/bj0410261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Manning JP, Meli A, Steinetz BG. Alkaline phosphatase and β-glucuronidase activity in the rat uterus during early pregnancy. J Endocrinol. 1966;35:385–91. doi: 10.1677/joe.0.0350385. [DOI] [PubMed] [Google Scholar]
- 54.Murdoch RN, Kay DJ, Cross M. Activity and subcellular distribution of mouse uterine alkaline phosphatase during pregnancy and pseudopregnancy. J Reprod Fert. 1978;54:293–300. doi: 10.1530/jrf.0.0540293. [DOI] [PubMed] [Google Scholar]
- 55.Valenzuela GJ, Munson LA, Tarbaux NM, Farley JR. Time-dependent changes in bone, placental, intestinal, and hepatic alkaline phosphatase activities in serum during human pregnancy. Clin Chem. 1987;33:1801–6. [PubMed] [Google Scholar]
- 56.Maruo T, Ishihara N, Samoto T, Murakoshi H, Laoag-Fernandez JB, Matsuo H. Regulation of human trophoblast proliferation and apoptosis during pregnancy. Early Pregn. 2001;5:28–9. [PubMed] [Google Scholar]
- 57.Elpek GÖ, Karaveli Ş, Keleş N. Preeklampsili olguların term plasentalarında villöz trofoblast preliferasyonunun incelenmesi. Türk Pathol Derg. 2000;16:10–2. [Google Scholar]
- 58.Erboga M, Kanter M. Trophoblast cell proliferation and apoptosis in placental development during early gestation period in rats. Anal Quant Cytopathol Histpathol. 2015;37:286–94. [PubMed] [Google Scholar]
- 59.Ozaydın T, Sur E, Oznurlu Y, Celik I, Uluisik D. Immunohistochemical distribution of heat shock protein 70 and proliferating cell nuclear antigen in mouse placenta at different gestational stages. Microsc Res Tech. 2016;79:251–7. doi: 10.1002/jemt.22624. [DOI] [PubMed] [Google Scholar]
- 60.Oner H, Oner J, Demir R. Distributions of PCNA and Cas-3 in rat uterus during early pregnancy. Folia Histochem Cytobiol. 2010;48:71–7. doi: 10.2478/v10042-008-0088-2. [DOI] [PubMed] [Google Scholar]
- 61.Danihel L, Gomolcak P, Korbel M, et al. Expression of proliferation and apoptotic markers in human placenta during pregnancy. Acta Histochem. 2002;104:335–8. doi: 10.1078/0065-1281-00683. [DOI] [PubMed] [Google Scholar]
- 62.Acar N, Korgun ET, Cayli S, Sahin Z, Demir R, Ustunel I. Is there a relationship between PCNA expression and diabetic placental development during pregnancy? Acta Histochem. 2008;110:408–17. doi: 10.1016/j.acthis.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 63.Ozaydın T, Sur E, Oznurlu Y, Celık I, Uluısık D. Immunohistochemical distribution of heat shock protein 70 and proliferating cell nuclear antigen in mouse placenta at different gestational stages. Microsc Res Tech. 2016;79:251–7. doi: 10.1002/jemt.22624. [DOI] [PubMed] [Google Scholar]