

SUMMARY
Termination and ribosome recycling are essential processes in translation. In eukaryotes, a stop codon in the ribosomal A site is decoded by a ternary complex consisting of release factors eRF1 and guanosine triphosphate (GTP)-bound eRF3. After GTP hydrolysis, eRF3 dissociates, and ABCE1 can bind to eRF1-loaded ribosomes to stimulate peptide release and ribosomal subunit dissociation. Here, we present cryoelectron microscopic (cryo-EM) structures of a pretermination complex containing eRF1-eRF3 and a termination/prerecycling complex containing eRF1-ABCE1. eRF1 undergoes drastic conformational changes: its central domain harboring the catalytically important GGQ loop is either packed against eRF3 or swung toward the peptidyl transferase center when bound to ABCE1. Additionally, in complex with eRF3, the N-terminal domain of eRF1 positions the conserved NIKS motif proximal to the stop codon, supporting its suggested role in decoding, yet it appears to be delocalized in the presence of ABCE1. These results suggest that stop codon decoding and peptide release can be uncoupled during termination.
INTRODUCTION
Translation termination and ribosome recycling are essential processes in ribosome-driven protein synthesis triggered by the appearance of a stop codon in the A site of the ribosome during elongation. In the first stage of this cycle, the release factor (RF) eRF1 is delivered to the ribosome by the guanosine triphosphatase (GTPase) eRF3, which departs following guanosine triphosphate (GTP) hydrolysis. Next, ABCE1 binds to the factor-binding site of ribosomes loaded with eRF1 and facilitates peptide release and then subunit dissociation. These events are tightly coordinated through their common utilization of eRF1 (Pisarev et al., 2010; Shoemaker et al., 2010; Shoemaker and Green, 2011).
To date, several X-ray and cryoelectron microscopic (cryo-EM) structures exist for individual eRFs as well as unbound and ribosome-bound eRF1-eRF3 complexes (Cheng et al., 2009; des Georges et al., 2014; Kong et al., 2004; Song et al., 2000; Taylor et al., 2012). Recent cryo-EM structures of a rabbit pretermination complex show eRF1 trapped in the process of delivery to the ribosome by eRF3 bound to the nonhydrolyzable GTP analog guanylyl imidodiphosphate (GDPNP). As a result, the catalytically essential GGQ motif of eRF1 is positioned approximately 90 Å apart from the peptidyl transferase center (PTC) where peptide release is ultimately catalyzed. Therefore, it remains an open question what the active conformation of eRF1 on the terminating ribosome might be. Moreover, whereas it has previously been shown that ABCE1 can stimulate eRF1-dependent peptide release before dissociating ribosomes into subunits, thereby coupling translation termination with ribosome recycling (Shoemaker and Green, 2011), we have little structural understanding of these processes.
Important clues regarding the possible behavior of eRF1 and eRF3 come from the closely related mRNA surveillance (or ribosome rescue) factors Pelota (Dom34p in yeast) and Hbs1. These factors are paralogs of eRF1 and eRF3, recognize stalled ribosomes, and initiate subsequent ribosome rescue/recycling together with ABCE1 that ends in degradation of aberrant mRNA and proteins (Barthelme et al., 2011; Doma and Parker, 2006; Pisareva et al., 2011; Shoemaker and Green, 2011). Cryo-EM structures of stalled ribosomes in complex with Pelota and either Hbs1 or ABCE1 showed that the central domain of Pelota undergoes a dramatic conformational change in these different complexes. In the prerescue state (in the presence of Hbs1:GDPNP), Pelota is packed against Hbs1 and not fully engaged in the A site, whereas in the recycling complex bound to ABCE1:adenylyl imidodiphosphate (ADPNP), Pelota stretches out within the A site reaching toward the P site-tRNA (Becker et al., 2011, 2012; Franckenberg et al., 2012). Based on the homology between these rescue factors and eukaryotic-RFs, similar behavior of eRF1 may explain how ABCE1 exerts its influence on peptide release. However, direct structural evidence for this model is not available so far.
RESULTS AND DISCUSSION
Generation and Cryo-EM of Pretermination and Termination/Prerecycling Complexes
Stable ribosomal complexes bound to eRF1 and eRF3 or ABCE1 were generated by employing a stalling polypeptide sequence from the human cytomegalovirus (CMV) gp48 uORF. This peptide sequence stalls translation by inhibiting eRF1-mediated peptide release with a UAA stop codonprogrammed ribosomal A site (Bhushan et al., 2010; Janzen et al., 2002). The detailed molecular changes responsible for prohibiting peptide release by eRF1 and also puromycin activity in this seemingly normal ribosomal termination complex are not known (Figure S1A).
We used a wheat germ in vitro translation system to generate CMV-stalled ribosome-nascent chain complexes (RNCs) (Bhushan et al., 2010) and then added either purified recombinant Saccharomyces cerevisiae eRF1-eRF3:GDPNP (Sup45p-Sup35p) ternary complex or eRF1 and ABCE1:ADPNP. eRF3 lacks the prion-forming domain (N-terminal 97 amino acids) that has been shown to be nonessential for termination activity in yeast (Alkalaeva et al., 2006; Frolova et al., 1996). To test the functional activity of these heterologous complexes, we performed release assays where we followed peptide release by immunodetection of the HA-tagged peptidyl tRNA and free peptide. In this case, the CMV-stalled RNCs were directly compared with RNCs prepared on a truncated mRNA. Although peptide was quantitatively released from the peptidyl tRNA by puromycin on the truncated mRNA RNCs, the CMV-stalled RNC peptides were substantially less reactive with puromycin (Figure S1A). These data confirmed the known downregulation of the PTC by the CMV-stalling peptide. Similarly, as expected, neither eRF1 alone nor in conjunction with eRF3 displayed detectable release activity with the CMV RNCs. Interestingly, eRF1 and ABCE1 together resulted in a detectable increase in the relative amounts of free peptide, consistent with earlier studies showing a stimulation of peptide release by ABCE1 (Shoemaker and Green, 2011). This limited peptide-release activity provides support for the functional relevance of the heterologous ribosome complexes analyzed in this manuscript.
We performed cryo-EM and single-particle analysis including in silico sorting procedures to obtain structures of CMV RNC-eRF1-eRF3:GDPNP and CMV RNC-eRF1-ABCE1:ADPNP at a resolution of 9.15 and 8.75 Å at a Fourier shell correlation (FSC) cutoff of 0.5, respectively (8.9 and 8.6 Å at a FSC cutoff at 0.143 after processing according to the so-called gold standard approach) (Figures 1A, S1B, and S1C). For molecular interpretation, we used an updated model of the Triticum aestivum ribosome (Gogala et al., 2014) and placed homology models of eRF1, eRF3, and ABCE1 in the assigned densities, where most secondary structure was resolved. This was validated by calculating the cross-resolution between the models and the maps (Figures 1B and S2).
Figure 1. Cryo-EM Structures of Pretermination and Termination/Prerecycling Complexes.
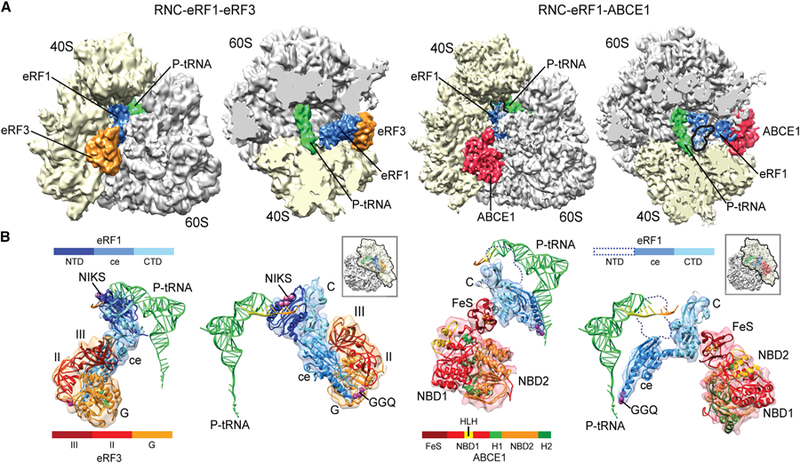
(A) Side and top views of the 80S ribosome pretermination complex with eRF1 and eRF3 (left) and termination/prerecycling complex with eRF1-ABCE1 (right). Density attributed to eRF1 occupies the A site. In the termination/prerecycling complex, the position of the flexible NTD of eRF1 is outlined with a black line.
(B) Molecular models for peptidyl tRNA, eRF1, eRF3, and ABCE1 on the ribosome. The NIKS motif (pink spheres) of eRF1 is positioned in close proximity to the stop codon (orange). The central domain of eRF1 containing the GGQ loop (magenta spheres) is packed against eRF3. In complex with ABCE1, the central domain of eRF1 is swung toward the PTC.
The pretermination complex showed extra densities for eRF1-eRF3 and P site-tRNA in positions consistent with previous observations in the rabbit eRF1:eRF3 pretermination complex (des Georges et al., 2014; Taylor et al., 2012) and in the yeast RNC-Pelota-Hbs1-complex (Becker et al., 2011). eRF1 is located in the A site, and its N-terminal domain (NTD) reaches into the decoding center of the small ribosomal subunit (SSU). The C-terminal domain (CTD) and central domain of eRF1 are packed against eRF3, which binds the ribosome like a classical EF-Tu-like translational GTPase. No density could be identified for the NTD of eRF3 (residues 97–255), suggesting a relatively flexible nature for this domain in our complex.
The termination/prerecycling complex showed dramatic conformational changes wherein eRF1 stretches between the P site-tRNA and ABCE1 that is located in the same position as seen previously in the Pelota ribosome complex (Becker et al., 2012). The CTD of eRF1 contacts the iron-sulfur (FeS) domain of ABCE1, whereas the central domain bearing the GGQ motif is stretched out toward the PTC of the large ribosomal subunit (LSU) where it contacts the P site-tRNA at the CCA end. Surprisingly, density for the NTD of eRF1 appeared to be fragmented and can only be visualized when the map is low-pass filtered at around 20 Å. This behavior is indicative of increased flexibility or disorder in this region that we confirmed in analysis of difference maps (Figure S3).
The Pretermination Complex
As mentioned briefly above, in the CMV RNC-eRF1-eRF3 pretermination complex, the ribosome adopts a similar overall conformation as observed for a stalled ribosome with Pelota-Hbs1 harboring a P site-tRNA (Becker et al., 2011) and the mammalian pretermination complex containing eRF1 and eRF3 (des Georges et al., 2014).
Consistent with the rabbit pretermination complex, the main contacts between eRF1 and the ribosome are found between the SSU and the NTD of eRF1 (Figure 2; Table S1). The conserved (TAS)NIKS loop is proximal to the stop codon poised in the A site, consistent with its critical role in stop codon recognition (Figure 2C). The NIKS loop is located in a similar position relative to the stop codon as for the equivalent loop (PVT/SPF) in bacterial RF1/RF2 that is involved in decoding (Korostelev et al., 2008; Laurberg et al., 2008; Weixlbaumer et al., 2008).
Figure 2. eRF1-Ribosome Interactions and Positioning of the NTD of eRF1 in the Pretermination Complex.
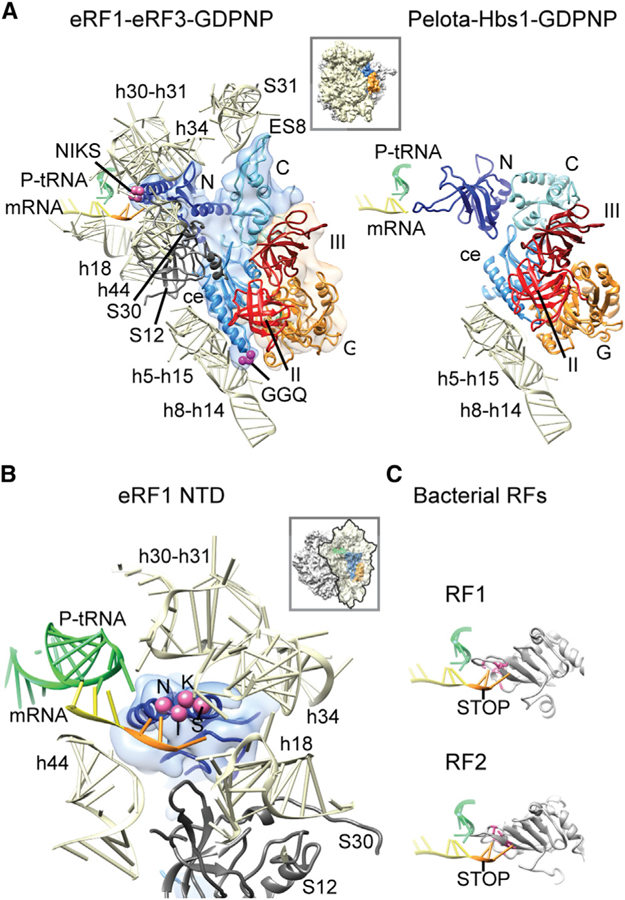
(A) eRF1 forms multiple contacts with the ribosome (left) that are mostly identical to those of Pelota in complex with Hbs1 (right) (Becker et al., 2011), apart from a contact at h8-h14 of the 18S rRNA. The minidomain of the CTD of eRF1 contacts ES8 and S31 near the beak of the SSU.
(B) The NTD reaches deep into the decoding center and establishes multiple contacts with 18S rRNA and S12 (left). The NIKS motif is close to the stop codon in the A site (orange).
(C) For decoding of the stop codon, bacterial RF1 and RF2 (Korostelev et al., 2008; Laurberg et al., 2008) rely on domain II that is unrelated to eRF1 NTD. Interacting amino acids are marked in pink.
Density for the NTD of eRF1 was not defined clearly enough as to allow for unambiguous positioning of the mRNA and individual residues of the (TAS)NIKS motif. Conformational changes of the NTD upon ribosome binding and during the event of decoding have indeed been postulated on the basis of toeprinting and chemical-crosslinking assays (Alkalaeva et al., 2006; Kryuchkova et al., 2013). In a previously proposed two-step model, recognition of the first two nucleotides in the codon is followed by a conformational change of the NTD of eRF1 that allows for decoding of the second and third nucleotides (Kryuchkova et al., 2013). The existence of distinct somewhat different conformations of eRF1 in this region might explain the limited resolution of the NTD during decoding in our structure.
The contacts between the CTD of eRF1 and domain III of eRF3 are formed by similar structural elements as previously reported in the crystal structure of human eRF1-eRF3 complex (Cheng et al., 2009). Here, however, helices α8 and α11 even more closely contact domain III of eRF3 (Figure S4). The minidomain in the CTD of eRF1 (that is only present in the nuclear magnetic resonance structure in the CTD of eRF1; Mantsyzov et al., 2010) anchors eRF1 to the beak of the SSU via the rRNA expansion segment ES8 and ribosomal protein (r-protein) S31. The central domain of eRF1 is tightly packed against all three domains of eRF3 and forms a large interaction surface of 1,088 Å2. As such, both the switch I and switch II regions of the G domain of eRF3 are in contact with eRF1 (Figure S4; Table S1).
Notably, we also observe a few differences when comparing our structure to the RNC-Pelota-Hbs1 structure or the rabbit pre-termination complex. In our structure, the inward movement of the stalk base compared to the factor-free state (rRNA helices H43 and H44 and r-protein L11, according to the nomenclature introduced by Jenner et al., 2012) is less pronounced (Figure S4). Concomitantly, the central domain and the CTD (including the minidomain) of eRF1 as well as eRF3 are bound in a more outward position such that the central domain of eRF1 is positioned closer to the small subunit and even contacts rRNA helix h14 with helix a5 that directly connects to the GGQ loop (Figure 2A; Table S1). As a result, the functionally critical GGQ loop is sandwiched between the G domain of eRF3 and the SSU in a tightly locked conformation that is incompatible with peptide-release activity at this pretermination stage. In order to be active for release, a dramatic conformational change is needed to position the GGQ motif of the eRF1 central domain in the peptidyl-transferase center.
The Termination/Prerecycling Complex
The overall conformation of the ribosome in the CMV RNC-eRF1-ABCE1 complex is indeed similar to that observed in the RNC-Pelota-ABCE1 complex (Becker et al., 2012). In both cases, the stalk base is moved downward toward the sarcin-ricin loop (SRL; H95) (Figure S4B), and as in the pretermination complex, we observe P site-tRNA and a nascent peptide in the ribosomal exit tunnel, indicating that minimal if any peptide release has occurred in this particle population (Figure 3B).
Figure 3. eRF1 Interactions and Positioning of Its Central Domain in the Termination/Prerecycling Complex.
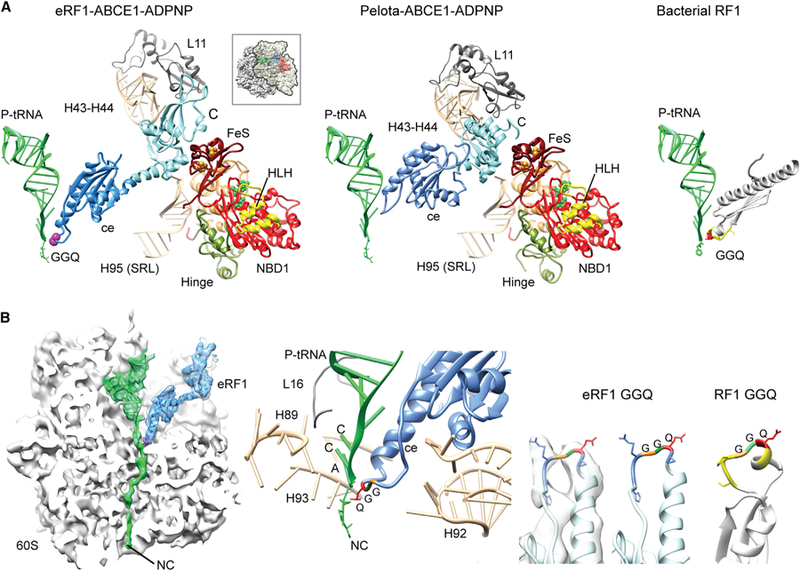
(A) The central domain of eRF1 undergoes a conformational change that positions the GGQ loop near the CCA end of the P site-tRNA (left). The CTD moves away from the SSU and forms contacts with the stalk base of the LSU and the SRL. These conformational changes are very similar to those of Pelota in complex with ABCE1 (middle). Unrelated domain III of bacterial RF1 possesses a different architecture but coordinates the highly conserved GGQ loop in an identical position (right).
(B) Cross-section and close-up view of the central domain of eRF1 with the GGQ loop close to the peptidyl tRNA (left and middle). Position and conformation of the GGQ loop are highly similar to that of bacterial RF1 (Laurberg et al., 2008).
The conformation of ABCE1 bound to the ribosome was also remarkably similar when compared to the Pelota-ABCE1-containing complex. ABCE1 binds in the translation factor-binding site and adopts an intermediate conformation of its nucleotide-binding domains (NBDs), somewhere between a fully open, ADP-bound structure, and the proposed closed ATP-bound form (Becker et al., 2012; Karcher et al., 2008). ABCE1 contacts the small subunit (h5-h15, h8-h14) mainly via its unique helix-loop-helix (HLH) and hinge motifs. Additional contacts are observed between NBD2 and L10.
Notably, eRF1 adopts a dramatically changed elongated conformation similar to ribosome-bound Pelota in the presence of ABCE1 (Figure 3). This elongated conformation is broadly similar to that of bacterial ribosome-bound RFs (Korostelev et al., 2008; Laurberg et al., 2008; Weixlbaumer et al., 2008), though in contrast to the bacterial structures, the NTD (the codon-interaction domain) of eRF1 appears to be delocalized.
The CTD of eRF1 contacts the FeS domain of ABCE1, the stalk base (H43-H44 and L11), and the SRL (H95) in the LSU. The central domain of eRF1 undergoes the most drastic conformational rearrangements in this structure, establishing multiple contacts to the rRNA (H71, H89, H91, H92, and H93) and stretching out toward the P site-tRNA. The conserved loop containing the GGQ motif is now located at the PTC of the LSU in close proximity to the CCA end of the peptidyl tRNA (Table S2). Modeling the GGQ region on the basis of previous crystal structures of bacterial RF1 and RF2 bound to the ribosome allowed for easily fitting of the density without further adjustments (Figure 3B). Although eRF1 is otherwise unrelated in sequence and structure (the class 1 RFs evolved independently in these two lineages), this structural finding suggests that the strictly conserved GGQ motif functions in the same way in these two systems.
Finally, we see stabilization of eRF1 in this active conformation by ABCE1 through contacting the CTD of eRF1. These structural observations rationalize how this ATPase can stimulate eRF1-dependent peptide-release activity (Shoemaker and Green, 2011). In order to fully appreciate the contribution that ABCE1 makes to positioning of eRF1 for catalysis, however, it will be useful to determine the structure of a ribosome complex loaded with eRF1 alone.
Conclusions
Our cryo-EM structures show that eukaryotic termination and ribosome recycling by eRF1, eRF3, and ABCE1 follow the same order of events and conformational transitions as observed previously for stalled ribosome rescue by Pelota, Hbs1, and ABCE1. In both pathways, the A site factor, eRF1 for termination and Pelota for ribosome rescue, is delivered by the EF-Tu-like GTPase eRF3 or Hbs1, respectively, which then dissociates from the ribosome after GTP hydrolysis (Figure 4). In their pre-GTP hydrolysis state, eRF1-eRF3 and Pelota-Hbs1 adopt similar conformations on the ribosome, though in the pre-termination (eRF1) complex, an additional prominent contact between the central domain of eRF1 and the SSU is established; as a consequence, this domain is more tightly locked between the SSU and eRF3. Notably, the central domain of eRF1 contacts both the switch I and switch II regions of eRF3 that control its GTP hydrolysis. Higher-resolution structures will be required to decipher how decoding of the stop codon coordinates these events on a molecular level.
Figure 4. Scheme of Eukaryotic Translation Termination and Ribosome Recycling.

For termination, the stop codon in the A site is recognized by the eRF1-eRF3-GTP ternary complex. eRF3 dissociates after GTP hydrolysis and allows the central domain of eRF1 to swing to the PTC. Proper positioning of the GGQ motif in the central domain of eRF1 may already allow peptide release, resulting in a termination complex with the deacyl-tRNA in the P state or P/E hybrid state. Alternatively, the active conformation of eRF1 in the pretermination complex is stabilized after binding of ABCE1. This stimulates peptide release while the NTD of eRF1 is delocalized, thus decoupling decoding from peptide release. Independent of the termination mechanism, ABCE1 together with eRF1 functions in concert to dissociate the ribosome into small and large subunits.
Next, we see that after eRF3 dissociation, eRF1 changes its conformation such that the central domain of eRF1 moves toward the PTC for catalysis of peptide release. The ribosome-eRF1 complex allows for binding of ABCE1 that appears to stabilize the fully extended active conformation of eRF1, thereby stimulating peptide release (Movie S1). Interestingly, the NTD of eRF1 appears to disengage the A site codon in this complex, indicating that codon engagement may not be required at this stage for peptide release. Yet, in contrast to the bacterial RFs that dissociate after termination (Freistroffer et al., 1997), eRF1 is still required for ABCE1-dependent ribosomal subunit splitting (Pisarev et al., 2010; Shoemaker and Green, 2011).
In a final stage, we know that ABCE1 functions in concert with bound eRF1 (on the posttermination complex) to promote subunit dissociation (Pisarev et al., 2010; Shoemaker and Green, 2011). Here, we see that ABCE1 adopts a remarkably similar conformation as observed in the prerecycling complex with Pelota (Becker et al., 2012). These data indicate that the mechanism of 80S splitting follows the same principle, independent of the nature of the ribosome to be recycled. Like Pelota in the context of ribosome rescue, eRF1 may act as a structural “bolt” that transmits conformational changes within ABCE1 upon ATP hydrolysis to the ribosome and induces splitting of the subunits. More structural and biochemical data will be needed to understand how this reaction is triggered and how ordered ATP binding and ATP hydrolysis in the two NBDs of ABCE1 contribute to this process.
EXPERIMENTAL PROCEDURES
Programmed CMV-stalled RNCs were prepared from a wheat germ in-vitro-translation extract as described by Bhushan et al. (2010). Recombinant yeast eRF1, eRF3, and ABCE1 were overexpressed in E. coli or S. cerevisiae and affinity purified. For release assays, RNCs were incubated together with the ligands, and tagged nascent peptidyl tRNA or free peptide was analyzed by western blotting.
Termination complexes were formed by in vitro reconstitution with recombinant-purified factors. The complexes were vitrified, and data were collected on a Titan Krios electron microscope (FEI). Single-particle analysis followed by 3D reconstruction was performed using the SPIDER software package (Frank et al., 1996). For molecular interpretation of the Triticum aestivum 80S ribosome, we used an updated model (Gogala et al., 2014). Models of eRF1, eRF3, and ABCE1 were based on existing crystal structures. See Supplemental Experimental Procedures for a detailed description of the Experimental Procedures.
Supplementary Material
ACKNOWLEDGMENTS
We thank Charlotte Ungewickell for assistance with cryo-EM and Heidemarie Sieber for assistance with biochemical experiments. This work was supported by grants from the German Research Council (GRK 1721, FOR 1805, and SFB646 to R.B., and SFB646 to T.B). R.B. acknowledges support by the Center for Integrated Protein Science and the European Research Council (Advanced Grant CRYOTRANSLATION).
Footnotes
ACCESSION NUMBERS
Cryo-EM maps for the pretermination complex (RNC-eRF1-eRF3) and the termination/prerecycling complex (RNC-eRF1-ABCE1) have been deposited in the EMDataBank under accession codes EMD-2597 and EMD-2598. The respective coordinates for EM-based models are deposited in the Protein Data Bank under ID codes 4rcn and 4rcm.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures, four figures, two tables, and one movie and can be found with this article online at http://dx.doi.org/10.1016/j.celrep.2014.04.058.
REFERENCES
- Alkalaeva EZ, Pisarev AV, Frolova LY, Kisselev LL, and Pestova TV. (2006). In vitro reconstitution of eukaryotic translation reveals cooperativity between release factors eRF1 and eRF3. Cell 125, 1125–1136. [DOI] [PubMed] [Google Scholar]
- Barthelme D, Dinkelaker S, Albers SV, Londei P, Ermler U, and Tampé R. (2011). Ribosome recycling depends on a mechanistic link between the FeS cluster domain and a conformational switch of the twin-ATPase ABCE1. Proc. Natl. Acad. Sci. USA 108, 3228–3233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berninghausen O, and Beckmann R. (2011). Structure of the no-go mRNA decay complex Dom34-Hbs1 bound to a stalled 80S ribosome. Nat. Struct. Mol. Biol 18, 715–720. [DOI] [PubMed] [Google Scholar]
- Becker T, Franckenberg S, Wickles S, Shoemaker CJ, Anger AM, Armache JP, Sieber H, Ungewickell C, Berninghausen O, Daberkow I, et al. (2012). Structural basis of highly conserved ribosome recycling in eukaryotes and archaea. Nature 482, 501–506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhushan S, Meyer H, Starosta AL, Becker T, Mielke T, Berninghausen O, Sattler M, Wilson DN, and Beckmann R. (2010). Structural basis for translational stalling by human cytomegalovirus and fungal arginine attenuator peptide. Mol. Cell 40, 138–146. [DOI] [PubMed] [Google Scholar]
- Cheng Z, Saito K, Pisarev AV, Wada M, Pisareva VP, Pestova TV, Gajda M, Round A, Kong C, Lim M, et al. (2009). Structural insights into eRF3 and stop codon recognition by eRF1. Genes Dev 23, 1106–1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- des Georges A, Hashem Y, Unbehaun A, Grassucci RA, Taylor D, Hellen CU, Pestova TV, and Frank J. (2014). Structure of the mammalian ribosomal pre-termination complex associated with eRF1.eRF3.GDPNP. Nucleic Acids Res 42, 3409–3418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doma MK, and Parker R. (2006). Endonucleolytic cleavage of eukaryotic mRNAs with stalls in translation elongation. Nature 440, 561–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franckenberg S, Becker T, and Beckmann R. (2012). Structural view on recycling of archaeal and eukaryotic ribosomes after canonical termination and ribosome rescue. Curr. Opin. Struct. Biol 22, 786–796. [DOI] [PubMed] [Google Scholar]
- Frank J, Radermacher M, Penczek P, Zhu J, Li Y, Ladjadj M, and Leith A. (1996). SPIDER and WEB: processing and visualization of images in 3D electron microscopy and related fields. J. Struct. Biol 116, 190–199. [DOI] [PubMed] [Google Scholar]
- Freistroffer DV, Pavlov MY, MacDougall J, Buckingham RH, and Ehrenberg M. (1997). Release factor RF3 in E. coli accelerates the dissociation of release factors RF1 and RF2 from the ribosome in a GTP-dependent manner. EMBO J 16, 4126–4133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frolova L, Le Goff X, Zhouravleva G, Davydova E, Philippe M, and Kisselev L. (1996). Eukaryotic polypeptide chain release factor eRF3 is an eRF1- and ribosome-dependent guanosine triphosphatase. RNA 2, 334–341. [PMC free article] [PubMed] [Google Scholar]
- Gogala M, Becker T, Beatrix B, Armache JP, Barrio-Garcia C, Berninghausen O, and Beckmann R. (2014). Structures of the Sec61 complex engaged in nascent peptide translocation or membrane insertion. Nature 506, 107–110. [DOI] [PubMed] [Google Scholar]
- Janzen DM, Frolova L, and Geballe AP. (2002). Inhibition of translation termination mediated by an interaction of eukaryotic release factor 1 with a nascent peptidyl-tRNA. Mol. Cell. Biol 22, 8562–8570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenner L, Melnikov S, Garreau de Loubresse N, Ben-Shem A, Iskakova M, Urzhumtsev A, Meskauskas A, Dinman J, Yusupova G, and Yusupov M. (2012). Crystal structure of the 80S yeast ribosome. Curr. Opin. Struct. Biol 22, 759–767. [DOI] [PubMed] [Google Scholar]
- Karcher A, Schele A, and Hopfner KP. (2008). X-ray structure of the complete ABC enzyme ABCE1 from Pyrococcus abyssi. J. Biol. Chem 283, 7962–7971. [DOI] [PubMed] [Google Scholar]
- Kong C, Ito K, Walsh MA, Wada M, Liu Y, Kumar S, Barford D, Nakamura Y, and Song H. (2004). Crystal structure and functional analysis of the eukaryotic class II release factor eRF3 from S. pombe. Mol. Cell 14, 233–245. [DOI] [PubMed] [Google Scholar]
- Korostelev A, Asahara H, Lancaster L, Laurberg M, Hirschi A, Zhu J, Trakhanov S, Scott WG, and Noller HF. (2008). Crystal structure of a translation termination complex formed with release factor RF2. Proc. Natl. Acad. Sci. USA 105, 19684–19689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kryuchkova P, Grishin A, Eliseev B, Karyagina A, Frolova L, and Alkalaeva E. (2013). Two-step model of stop codon recognition by eukaryotic release factor eRF1. Nucleic Acids Res 41, 4573–4586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laurberg M, Asahara H, Korostelev A, Zhu J, Trakhanov S, and Noller HF. (2008). Structural basis for translation termination on the 70S ribosome. Nature 454, 852–857. [DOI] [PubMed] [Google Scholar]
- Mantsyzov AB, Ivanova EV, Birdsall B, Alkalaeva EZ, Kryuchkova PN, Kelly G, Frolova LY, and Polshakov VI. (2010). NMR solution structure and function of the C-terminal domain of eukaryotic class 1 polypeptide chain release factor. FEBS J 277, 2611–2627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisarev AV, Skabkin MA, Pisareva VP, Skabkina OV, Rakotondrafara AM, Hentze MW, Hellen CU, and Pestova TV. (2010). The role of ABCE1 in eukaryotic posttermination ribosomal recycling. Mol. Cell 37, 196–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisareva VP, Skabkin MA, Hellen CU, Pestova TV, and Pisarev AV. (2011). Dissociation by Pelota, Hbs1 and ABCE1 of mammalian vacant 80S ribosomes and stalled elongation complexes. EMBO J. 30, 1804–1817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoemaker CJ, and Green R. (2011). Kinetic analysis reveals the ordered coupling of translation termination and ribosome recycling in yeast. Proc. Natl. Acad. Sci. USA 108, E1392–E1398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoemaker CJ, Eyler DE, and Green R. (2010). Dom34:Hbs1 promotes subunit dissociation and peptidyl-tRNA drop-off to initiate no-go decay. Science 330, 369–372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song H, Mugnier P, Das AK, Webb HM, Evans DR, Tuite MF, Hemmings BA, and Barford D. (2000). The crystal structure of human eukaryotic release factor eRF1—mechanism of stop codon recognition and peptidyl-tRNA hydrolysis. Cell 100, 311–321. [DOI] [PubMed] [Google Scholar]
- Taylor D, Unbehaun A, Li W, Das S, Lei J, Liao HY, Grassucci RA, Pestova TV, and Frank J. (2012). Cryo-EM structure of the mammalian eukaryotic release factor eRF1-eRF3-associated termination complex. Proc. Natl. Acad. Sci. USA 109, 18413–18418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weixlbaumer A, Jin H, Neubauer C, Voorhees RM, Petry S, Kelley AC, and Ramakrishnan V. (2008). Insights into translational termination from the structure of RF2 bound to the ribosome. Science 322, 953–956. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.