

Abstract
From the dawn of civilization, humanity has dreamed of immortality. So why didn’t the discovery of the anti-aging properties of mTOR inhibitors change the world forever? I will discuss several reasons, including fear of the actual and fictional side effects of rapamycin, everolimus and other clinically-approved drugs, arguing that no real side effects preclude their use as anti-aging drugs today. Furthermore, the alternative to the reversible (and avoidable) side effects of rapamycin/everolimus are the irreversible (and inevitable) effects of aging: cancer, stroke, infarction, blindness and premature death. I will also discuss why it is more dangerous not to use anti-aging drugs than to use them and how rapamycin-based drug combinations have already been implemented for potential life extension in humans. If you read this article from the very beginning to its end, you may realize that the time is now.
Keywords: rapamycin, rapalogs, metformin, aging, anti-aging, fasting, lifespan, health span
“If you wait until you are ready, it is almost certainly too late.” Seth Godin
In one short-lived mutant strain of mice, the mTOR inhibitor rapamycin (known in the clinic as Sirolimus) extends maximum life span nearly three-fold [1]. Albeit less spectacularly, rapamycin also prolongs life in normal mice as well as in yeast, worms and flies, and it prevents age-related conditions in rodents, dogs, nonhuman primates and humans. Rapamycin and its analog, everolimus, are FDA approved for human use and have been used safely for decades. In 2006, it was suggested that rapamycin could be used immediately to slow down aging and all age-related diseases in humans [2], becoming an “anti-aging drug today” [3].
But rapamycin was unlucky
Rapamycin known in the clinic as Rapamune or Sirolimus, was unlucky from the start, however. Twenty years ago, it was labeled an immunosuppressant and used to treat renal transplant patients. If rapamycin had been labeled an immunomodulator and anti-inflammatory drug instead, it would sound much more appealing now. At anti-aging doses, rapamycin “eliminates hyperimmunity rather than suppresses immunity” or, more figuratively, it “rejuvenates immunity” [2]. This enables rapamycin and everolimus, a rapamycin analog, to act as immunostimulators [4–6], improving immunity in cancer patients [7] and the elderly [8,9]. For example, rapamycin reduces the risk of CMV infection in organ transplant patients [10–12], improves antipathogen and anticancer immunity in mice [13–15], prolongs lifespan in infection-prone mice [16] and protects aged mice against pneumonia [17]. Rapamycin also inhibits viral replication [18,19]. As a noteworthy example, rapamycin inhibits replication of the 1918 flu virus (the deadliest flu virus in history) by 100-fold [19], and also protects against lethal infection with influenza virus when administered during vaccination [13]. Still, as Dr. Allan Green advises, patients taking rapamycin should be carefully monitored for skin and subcutaneous bacterial infections, which should be treated with antibiotics https://rapamycintherapy.com.
Twenty years ago, it was thought that rapamycin might increase the risk of cancer (see a forthcoming review “Understanding the side effects of rapamycin”). Despite that concern, it was revealed that rapamycin actually prevents lymphoma and some types of cancer in transplant patients [20–27]. Currently, in fact, rapamycin analogs, everolimus and temsirolimus, are widely used in cancer therapy. Furthermore, rapamycin is the most effective known cancer-preventive agent in mice [25,28–32] extending the lifespan of cancer-prone mice [33–36]. It has even been suggested that rapamycin extends lifespan by preventing cancer [37].
Nevertheless, social media often warn that although rapamycin prevents cancer, its use to prevent cancer may come at the cost of getting cancer. This self-contradiction miscites a twenty-year-old warning by the FDA for all drugs marketed as immunosuppressants (including rapamycin and everolimus): “Increased susceptibility to infection and the possible development of malignancies such as lymphoma and skin cancer may result from immunosuppression.” This statement does not say that rapamycin or everolimus cause malignancies. (Just read it again). Although rapamycin and its analogs are now approved by the FDA for treatment of cancer and lymphomas, the rumors that these drugs may cause cancer persist. To my knowledge, no study has shown that mTOR inhibitors cause cancer.
At this point, most scientists agree that rapamycin is not counterindicated because of concerns about immunosuppressive effects. But a new objection against rapamycin has emerged, namely that rapamycin may cause diabetes. As discussed in detail [38], the new wave of “fear of rapamycin” is groundless. So, what are the metabolic effects of rapamycin?
Metabolic effects or rapamycin and starvation
When it is over-activated by nutrients and insulin, mTOR acts via S6K to inhibit insulin signaling, thereby causing insulin resistance [39–44]. Acute treatment with rapamycin abrogates insulin resistance in cells and animals including humans [45–51]. One study showed that chronic treatment with rapamycin may also prevent insulin resistance [52]. However, in some (but not all) rodent models, chronic treatment with rapamycin can also cause glucose intolerance and even insulin resistance [53–56]. This was interpreted as a deleterious side effect or even type 2 diabetes (T2D). Actually, however, these metabolic changes are features of benevolent starvation pseudo-diabetes (SPD), which was described 170 years ago in fasted animals and later in humans [57,58]. During prolonged fasting, utilization of glucose by non-brain tissues must be suppressed to ensure an adequate supply to the brain. When a fasted animal or human is then given a meal, glucose appears in the urine (glycosuria), which is a canonical symptom of diabetes. But this is because prolonged fasting (starvation) leads to decreased insulin secretion and to insulin resistance, and subsequent re-feeding then causes transient hyperglycemia and glycosuria. This SPD can be caused not only by prolonged fasting, but also by severe restriction of calorie and carbohydrate intake [38]. For example, severe calorie restriction can cause diabetes-like glucose intolerance [59]. Despite that, very low calorie diets are the most effective treatments for type 2 diabetes [60–62]. Some researchers re-discovered SPD in obese patients on strenuous weight loss program and erroneously warned that low calorie diets cause type 2 diabetes [63].
The obligatory symptom of starvation is ketosis, as ketones substitute for glucose as the main fuel for the brain. The ketogenic diet, a promising treatment for diabetes and obesity in humans, can cause symptoms of SPD in rodents (see for references [64]). Once again, some researchers warned that the ketogenic diet can favor type 2 diabetes [65]. As discussed, such warnings may not be justified [64,66–68].
Rapamycin can be viewed as a partial starvation-mimetic [69–71]. It is therefore predictable that, under some conditions, prolonged treatment with rapamycin may lead to the emergence of SPD [72]. This has been confirmed in rapamycin-fed mice, which developed insulin resistance, glucose intolerance and mild hyperglycemia [54]. Nevertheless, rapamycin-fed mice lived longer and thus were healthier than mice fed a standard diet [54]. It is not completely clear why SPD was observed in only some studies and was not observed in other studies (see for references [38,73]).
Importantly, SPD is reversible and does not lead to complications. Furthermore, rapamycin reduces the incidence of diabetic complications such as diabetic nephropathy in rodents [42,74–85]. In healthy elderly humans, chronic treatment with rapamycin or everolimus did not cause hyperglycemia [8,9,86]. On the contrary, the risk of hyperglycemia was lower in the mTOR inhibitor treatment group than the placebo group [9].
In some cancer patients, high doses of rapamycin or everolimus can cause hyperglycemia, which is usually mild (grade 1-2) and reversible, and does not lead to treatment interruption [87–89]. Hyperglycemia is a common side effect of many oncotargeted drugs and is not a manifestation of diabetes. Everolumus, for example, can cause hyperglycemia by decreasing insulin production [89].
To summarize, chronic treatment with high doses of rapamycin may cause symptoms of reversible SPD. Diet-induced SPD, at least, is beneficial and therapeutic. Rapamycin-induced SPD is a relatively rare side effect and probably can be avoided by administering the drug intermittently or at lower doses, and if SPD does occur, it can be reversed by discontinuation of the drug.
In some studies in transplant patients, rapamycin (sirolimus) and everolimus did not increase the risk of diabetes [90–96]. In one study, no patient, out of 21 patients treated with rapamycin, developed diabetes, while the incidence of diabetes was 7% in patients treated with either cyclosporine or tacrolimus [96]. Most importantly, cyclosporine- or tacrolimus-induced diabetes resolved in 80% of patients after conversion from tacrolimus/cyclosporine to rapamycin (sirolimus) [96].
On the other hand, a large retrospective study reported an association between Medicare billing for diabetes treatment and rapamycin use, implying that rapamycin may increase the risk of diabetes [97]. However, this association was explained by the interaction between rapamycin and calcineurin inhibitors, which increase each other’s levels [96,98,99]. Consequently, it remains unclear whether rapamycin per se increases the risk of diabetes in transplant patients [96]. Moreover, this is further complicated by the fact that most transplant patients develop type 2 diabetes spontaneously without rapamycin treatment [100].
Rapamycin is not much more dangerous than ordinary drugs
If used properly, rapamycin is not much more dangerous than ordinary aspirin. Aspirin, one of the most widely used nonprescription medications, may cause numerous side effects, including life threatening gastric bleeding. The manufacturer lists as possible side effects: ringing in ears, confusion, hallucinations, seizure, severe nausea, vomiting, bloody stools, coughing up blood, fever and swelling. Still, millions of people take aspirin daily to prevent cardiovascular disease and cancer. It was calculated that the benefits of aspirin are greater than their risks [101,102]. I believe the benefits of the anti-aging effects of rapamycin/everolimus may even be greater (Figure 1).
Figure 1.
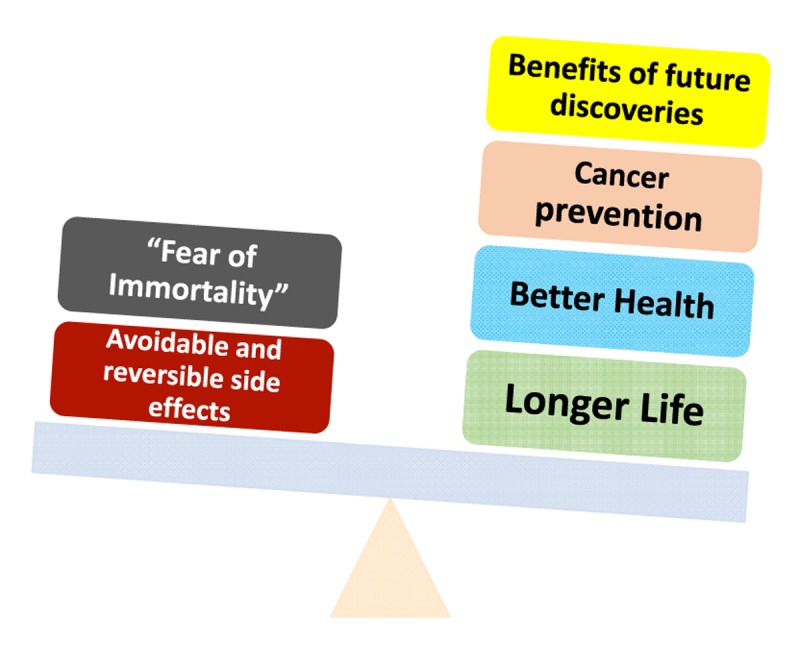
Potential risk vs benefits of rapamycin-based anti-aging therapy. Pros and Cons: Potential benefits of rapamycin may outweigh its risks.
In the case of rapamycin and everolimus, the most worrying side effects have not been confirmed. At low doses [8,9,86], or when administered as a single high dose [103], no side effects have been detected so far in the elderly. At high doses, rapamycin and everolimus slow cell proliferation, which decreases blood cell counts. As a result, mild and reversible thrombocytopenia (low platelet count), anemia and leukopenia are their most common side effects. But a mild reduction of platelets may be beneficial. In fact, one of the intended effects of aspirin is a decrease in platelet function.
There is one crucial reason why the side effects of rapamycin are exaggerated. The frequency of rapamycin side effects has often been estimated in studies lacking a placebo group. In cancer and transplant patients, numerous effects ascribed to rapamycin, such as fatigue (asthenia), for example, are often caused by the disease itself. In a placebo study of healthy volunteers, the placebo group reported more side effects such as fatigue than did the rapamycin group [104]. In recent placebo-controlled studies in healthy elderly people, no side effects were noticed as compared to placebo [9,86].
While aspirin may cause gastric ulceration and bleeding, rapamycin may cause stomatitis and mycositis (ulceration of the mucous membranes of the mouth and the digestive tract) when used at high doses or chronically. A rare side effect of rapamycin is noninfectious interstitial pneumonitis [105]. And by inhibiting neutrophil function, rapamycin may increase the severity of bacterial infections [106]. These side effects require rapamycin’s discontinuation. For antiaging purposes, however, rapamycin may be used either intermittently (e.g., once a week) or at low daily doses and can be discontinued if any unpleasant effects occur.
From a single dose to intermittent schedules
Although nearly all drugs, including nonprescription drugs such as aspirin, can be fatal at sufficiently high doses, there are no known fatal cases of acute rapamycin (sirolimus) overdose [103]. For example, in a failed suicide attempt, an 18-year-old woman ingested 103 rapamycin tablets (103 mg), and the only detected effect was an elevation in total blood cholesterol [103]. In rats, rapamycin’s LD50, a measure of drug lethality, could not be determined because it is higher than 2500 mg/kg. While a single dose of rapamycin is safe, it is sufficient to extend life and decrease obesity in several rodent models [1,107]. Furthermore, transient treatment with rapamycin can be long lasting, extending the lifespan and preventing obesity long after drug discontinuation [107–112]. The magnitude of life extension by rapamycin depends mostly on reaching a high peak blood level [113]. A similar conclusion was reached by a study of rapamycin use in obesity [112]. It was suggested in 2008 that a pulse (intermittent) schedule of rapamycin administration would improve regeneration of stem cells [114] while avoiding mTORC2 inhibition [54,115].
Therefore, to avoid side effects and maximize anti-aging effects [110], a feasible approach would be to prolong intervals between rapamycin administrations while keeping the total dose constant. For example, instead of daily administration, a weekly administration of a higher dose can be suggested to achieve a high peak blood level, followed by drug-free period to avoid undesirable effects. Still, everyday treatment of the elderly (1 mg/day for several weeks) was not associated with side effects and has been shown to be safe [86]. Similar results were achieved with low doses of other mTOR inhibitors [9]. Another option is an alternating schedule; for example, a 3- month course of weekly rapamycin alternating with a rapamycin-free month. Finally, anti-aging schedules can be very flexible to fit an individual patient. The optimal anti-aging dose is a personalized maximum dose that does not cause side effects in a particular patient (Figure 2).
Figure 2.

Optimal dose of rapamycin for maximal net benefits. Life extension by rapamycin is dose-dependent in rodents. The higher the dose, the higher the anti-aging benefits, including cancer prevention and life extension. In humans, side effects are dose-dependent and net benefits could potentially decrease at very high doses. This point of the highest net benefit is the optimal dose. The optimal dose varies in different individuals due to the variability of potential side effects. Thus, the optimal dose in a particular individual is determined by the emergence of side effects. The treatment can be viewed as life-long phase I/II clinical trial.
In conclusion, the side effects of rapamycin are well-known and reversible. When used on an anti-aging schedule, side effects may be absent but, if not, they may be mitigated by combining rapamycin with other anti-aging drugs (metformin, statins) or by temporarily discontinuing it.
Noteworthy, the alternative to the reversible (and avoidable) side effects of rapamycin/everolimus are the irreversible (and inevitable) effects of aging. And by living longer, our generation will benefit from future anti-aging discoveries (Figure 1).
But the fear of nonexistent side effects is not the only reason the use of mTOR inhibitors for life extension has been questioned. The second reason is that there is rightful skepticism about any claims made about anti-aging drugs because thousands of anti-aging remedies have already failed. What then makes rapamycin different?
The history of mankind: empty promises of immortality
On the one hand, from the dawn of civilization humans have dreamed of immortality. On the other hand, from the dawn of civilization a myriad of anti-aging remedies turned out to be empty promises. Even worse, they often shorten lifespan. Two notable examples are antioxidants and human growth hormone. The idea that free radicals, or reactive oxygen species (ROS), cause aging was based on a “wild guess,” as Harman, a father of the ROS theory, acknowledged when he titled his paper, “I thought, thought, thought for four months in vain and suddenly the idea came” [116]. The idea is simple and intuitive, and it was widely accepted based on circumstantial evidence. In fact, ROS are inevitable products of metabolism, and they do damage biomolecules. Moreover, excessive ROS can shorten lifespan. Similarly, the atomic bomb can shorten life span. Yet this does not mean that either atomic bombs or oxidants are the cause of normal aging as we know it.
Numerous experiments support the ROS theory. However, key experiments ruled the ROS theory out (see for references [2,117–122]. To make a long story short, antioxidants could in theory prolong lifespan if mTOR-driven (quasi-programmed) aging were suppressed and we lived long enough to die from ROS-induced post-aging syndrome (I will discuss the nuances in the forthcoming article “ROS and aging revisited”). Indeed, ROS will kill any organism eventually. However, organisms normally die from mTOR-driven, age-related diseases (aging as we know it) before ROS can kill them (see for discussion [2]). As an analogy, consider most of the passengers on the Titanic. Would antioxidant treatment have been useful to them for life extension? The best way to extend life for members of that group would have been to carry more life boats. Only after their safe rescue could one expect antioxidants to potentially increase their life further. Similarly, only after rescue from the quasi-program of aging may antioxidants potentially have an impact.
Not surprisingly, antioxidants did not extend lifespan in any clinical trials and were detrimental in some [122–133]. As Ristow put it, they were “worse than useless” [119]. For example, in two very large randomized controlled trials, antioxidants increased the incidence of cancer, especially of lung cancer in smokers [131–133]. Antioxidants also increased all-cause mortality. The results were so disturbing that two trials were stopped earlier than planned [131–133]. Also disturbing is the finding that antioxidants accelerate cancer progression and promote metastasis [134–136]. But despite their uselessness, antioxidants continue to be a multibillion-dollar business. They are widely sold as natural products in the forms of nutritional supplements and in foods “rich in antioxidants.”
Another example is human growth hormone (HGH), which is widely used for rejuvenation and longevity. Yet, it actually accelerates aging and shortens lifespan [137,138]. Growth hormone is a pro-aging hormone because it indirectly activates mTOR [139]. Notably, the hype around growth hormone is based on a single publication [140], which misinterpreted its acute effects [141].
Given that all previous anti-aging remedies have failed to meet expectations, it is not surprising that the discovery of the anti-aging effects of rapamycin are being met with skepticism too. But unlike HGH, the effects of rapamycin are not based on one single paper as were HGH, nor is it based on a wild guess as were ROS.
Rapamycin is a proven anti-aging drug
The evidence that rapamycin can function as an anti-aging drug is the product of thousands of scientists working independently all over the world, studying mTOR and its inhibitors for a variety of different reasons in diverse organisms, ranging from yeast to humans. Studies in model organisms, such as yeast, worms and flies, have revealed components of the TOR signaling pathway [142–145]. It was predicted in 2003 [146] that conversion from quiescence to senescence (geroconversion) is driven by growth-promoting mediators, such as mTOR, when the cell cycle is blocked [147]. Figuratively, geroconversion is “twisted” growth that occurs when actual growth is completed [2], [147]. In cell culture, mTOR is maximally activated and geroconversion lasts 3-6 days, whereas in the human body it may take decades. mTOR drives geroconversion, rendering cells hypertrophic and hyperfunctional (e.g. senescence-associated secretory phenotype), which eventually leads to the development of age-related pathologies [2]. Working independently, clinical researchers have studied rapamycin for the prevention and treatment of nearly every age-related disease, including cancer, obesity, atherosclerosis and neurodegeneration. If a drug is indicated for all age-related diseases, it must be an anti-aging drug in that it targets a common driver of age-related diseases – that is, aging (see for references [2]). This is because aging is the sum of all age-related diseases, which limit lifespan [148–150]. Does rapamycin suppress aging and extend lifespan by preventing diseases, or does it prevent diseases by slowing aging? Actually, both reflect the same process.
By 2006, an extensive body of work from several independent fields all pointed to rapamycin as an anti-aging drug [2]. According to hyperfunction theory, aging is an unintended (not programmed but quasi-programmed) continuation of the developmental growth program, driven in part by mTOR [2,120,121,151,152]. Testable predictions have been formulated [2,153] and confirmed in numerous independent studies (see for references: [150,154]).
In two dozen studies using different strains of mice, rapamycin extended life span. Starting from a thorough study by Harrison et al. [155] and followed by nearly simultaneous studies by others [33,108], the anti-aging effects of rapamycin have been confirmed many times (see for references: [113,150,156,157]). Importantly, rapamycin and everolimus are indicated in most, if not all, age-related diseases, from cancer to neurodegeneration [2,158].
Conventional drugs as anti-aging agents
Several conventional drugs used to treat age-related diseases (e.g., hypertension, ischemic heart disease, diabetes, cancer, prostate enlargement) can be viewed as somewhat anti-aging drugs [150,154]. First, these drugs extend lifespan in the same model organisms (see for references: [159]). For example, metformin extends lifespan not only in mice, but also in the worms, which do not suffer from human diseases [160,161]. ACE inhibitors prolong life not only in hypertensive rats, but also in healthy normotensive rats [162]. If these drugs were not ordinary drugs for human diseases, then gerontologists would call them anti-aging agents.
Second, these drugs prevent or treat more than one disease. For example, metformin is indicated to treat type 2 diabetes as well as pre-diabetes, obesity, metabolic syndrome, cancer, and polycystic ovary syndrome [163–168]. Aspirin not only reduces inflammation (a hallmark of aging), it also reduces the risk of cardiovascular disease, thrombosis and cancer. Low-dose aspirin prevents one-third of colorectal, gastric, and esophageal cancers [169]. PDE5 inhibitors such as Sildenafil and Tadalafil, which are widely used for erectile dysfunction, are also effective against benign prostatic hyperplasia (BPH) and pulmonary arterial hypertension in humans and suppress inflammation-driven colorectal cancer in mice [170]. Aging is the sum of all these age-related diseases. Given that humans and animals die from age-related diseases, life can be extended by treating multiple pre-diseases and diseases. Rapamycin and these drugs may complement each other in an anti-aging formulation by further extending life and/or by mitigating each others possible side effects [159]. For example, metformin may counteract rapamycin-induced hyperglycemia [171].
Not taking rapamycin may be as dangerous as smoking
Strangely, the fear of tobacco smoking is less intense than the fear of rapamycin. But whereas smoking shortens both the healthspan and lifespan, rapamycin extends them. Smoking increases the incidence of cancer and other age-related diseases. Rapamycin prevents cancer in mice and humans. Heavy smoking shortens life expectancy by 6-10 years. In other words, simply not smoking prolongs life by 6-10 years. In middle-aged mice, just 3 months of high-dose rapamycin treatment was sufficient to increase life expectancy up to 60% [109]. When taken late in life, rapamycin increases lifespan by 9-14% [155], despite the dosage being suboptimal [111]. This possibly equates to more than 7 years of human life. By comparison, smokers who quit late in life (at age 65 years), gain between 1.4 -3.7 years [172]. Considered in those terms, one could say that in the elderly, not taking rapamycin may be even more “dangerous” than smoking. Finally, rapamycin may be especially beneficial to smokers and former smokers. While the carcinogens from tobacco cause lung cancer in mice, rapamycin decreases tobacco-induced lung cancer multiplicity by 90% [28].
Diet and rapamycin
Calorie restriction (CR) and intermittent fasting (IF) extend both the lifespan and healthspan in diverse species. However, CR is of little benefit when started in old age [73,173–178]. Fasting inhibits the mTOR pathway in young but not old mice [179,180]. By contrast, rapamycin strongly inhibits mTORC1 at any age. It extends lifespan, whether started late or early in life [108,155,181], even if used transiently [109]. So, whereas CR is more beneficial early in life, rapamycin may be indicated later in life. In addition, the beneficial effects of rapamycin and CR may be additive, given that they are exerted through overlapping but distinct mechanisms [182–186]. Intermittent rapamycin and CR (24-48 hours after) can be combined, to avoid potential hyperglycemia. Physical exercise may be most beneficial starting immediately after rapamycin use, to take advantage of rapamycin-induced lipolysis as a fuel for the muscles. By itself, chronic rapamycin treatment does not compromise muscle endurance [187] and even prevents muscle loss [188–190].
Do we need new or safer rapalogs to start aging prevention?
Despite the metabolic side effects seen in some mouse models, mice treated with rapamycin live longer and are healthier. Humans also may want to live longer and healthier lives, regardless of whether one calls the means unsafe. Some basic researchers believe that rapamycin cannot be routinely used to treat aging in humans because of its metabolic effects and call for the development of safer analogs. First, rapamycin and everolimus are FDA-approved drugs, safe for human use. Since 1999, rapamycin has been used by millions of patients with no unexpected problems. One may suggest that rapamycin/everolimus are safe enough for very sick patients, not for healthy people.
First, healthy elderly people chronically treated with rapamycin or other mTOR inhibitors showed no ill effects (e.g. hyperglycemia) [8,9,86]. Logically, more threatening adverse effects could be expected in cancer and transplant patients, who are often heavily pre-treated and terminally ill than in healthy people. Second, there are no truly healthy people among the elderly; otherwise, they would be “immortal”, given that all humans die from age-related diseases, not from healthy aging. And the sooner they would be treated with anti-aging drugs, the longer they would remain relatively healthy.
That said, it is, of course, important to develop new rapalogs, but not because current rapalogs are unsafe. It is important because such research will help us to learn more about mTOR and aging and may lead to the discovery of agents capable inhibiting the rapamycin-insensitive functions of mTORC1. These future drugs could potentially complement current rapalogs to further extend lifespan. Non-rapalog rapamycin analogs will also be developed [191]. The limitation of current rapalogs is not that they are unsafe but that their ability to extend life is limited. The goal should be to develop new drugs that extend life span further.
Rapamycin is a natural anti-fungal antibiotic produced by soil bacteria of Eastern Island. The patent on rapamycin has expired, and pharmacological companies have developed other rapalogs such as everolimus. (I use the term rapalogs to encompass both rapamycin, everolimus and any other analogs). At equipotent doses, rapamycin and everolimus exert almost identical therapeutic and adverse effects; although, everolimus is weaker and has a shorter half-life in the organism compared with rapamycin.
All current rapalogs exhibit the same side effects as rapamycin and everolimus. Their real side effects are mTORC1-dependent. Inhibition of mTORC1 decreases cell proliferation and function, which is manifested as lower blood cell counts and insulin levels, especially when rapalogs are chronically administered at high doses. We could develop weaker rapalogs, which would have no side effects if used at the same dose as rapamycin. But then why not just use a lower dose of rapamycin? (I will discuss elsewhere how safer rapalogs are probably weaker rapalogs.) Given to mice at the same doses as rapamycin, weaker analogs would have neither side effects and no therapeutic effects. Consequently, their metabolic effects would be diminished and so would their therapeutic effects. However, the same negative result can be achieved simply by decreasing the dose of rapamycin. While waiting for silver bullets, we need to use the currently available rapalogs, such as rapamycin and everolimus, to live longer. When “safer” rapalogs are clinically available, we may use them too.
The time is now unless it’s too late
The overwhelming evidence suggests that rapamycin is a universal anti-aging drug – that is, it extends lifespan in all tested models from yeast to mammals, suppresses cell senescence and delays the onset of age-related diseases, which are manifestations of aging [discussed by me in [148,149,158,192]. Although rapamycin may reverse some manifestations of aging [181,193], it is more effective at slowing down aging than reversing it. Therefore, rapamycin will be most effective when administered at the pre-disease, or even pre-pre-disease stages of age-related diseases [150]. For example, Carosi et al. suggested that mTOR inhibitors could be useful in Alzheimer disease, but only in the earliest stages [194,195]. In addition, rapamycin and everolimus are more effective for preventing cancer than treating it. They may also be useful for treating osteoporosis, though not a broken hip after an osteoporotic fracture. Rapalogs may slow atherosclerosis, thereby preventing myocardial infarction, but they are unlikely to help reverse an infarction. In other words, anti-aging drugs extend the healthspan (Figure 3) and are most effective before overt diseases cause organ damage and loss of function.
Figure 3.

Effects of standard and anti-aging medicine on health- and life-span. (A) The relationship between health- and life-span. Aging is a sum of all age-related diseases, pre-diseases and pre-pre-diseases. Before overt age-related diseases become apparent, there is a seemingly healthy period of aging (so-called healthy aging). Starting from adulthood, pre-pre-diseases progress towards pre-diseases and then towards overt diseases. Unless treated with modern standard medical practice, the diseased stage is relatively brief. From (A) to (B) Standard medical treatment is usually started when overt diseases are diagnosed. Standard medicine extends life span mostly by preventing death from diseases, thus extending “unhealthy” phase of life, especially terminal stages of diseases, characterized by organ damage, failure and loss of functions. Standard medicine extends lifespan. From (B) to (C) Anti-aging medicine is most effective at the stage of pre-diseases and initial stages of diseases, characterized by increased functions before complications and organ damage occur. In terminal stages of deadly diseases, anti-aging therapy may not be useful. Thus, anti-aging medicine increases both health span and life span. Anti-aging medicine and standard medicine are additive when aging becomes unhealthy. The schema is simplified because, in reality, age-related diseases start at different ages (presbyopia vs sarcopenia), progress at different paces (atherosclerosis vs cancer), and most are not lethal, and some are well treated (cataract). Therefore, healthspan is an abstraction.
So, is it too late to take rapamycin once aging reaches an unhealthy stage? Actually, it is not too late. Even if one or a few age-related diseases renders aging unhealthy, other potential diseases are still at pre-disease stages, and anti-aging drugs may delay their development. And they may slow down further progression of existing overt diseases.
In addition to rapamycin/everolimus, the anti-aging formula metformin, aspirin, ACE inhibitors, angiotensin receptor blockers and PDE5 inhibitors, each of which can prevent or treat more than one age-related disease [159]. Note that I mention only clinically-approved drugs because they can be used now. Later, perhaps, we may be able to consider further life extension through the use of low doses of pan-mTOR [196,197], mdm-2 [198,199] and MEK inhibitors [200,201], lithium [201,202], as well as next-generation rapalogs.
There is currently no consensus around the short-term markers of anti-aging effects. Therefore, rapamycin trials should be focused on its potential side effects rather than anti-aging effects. We must be sure that the therapy is safe. In the future, the treatment should be conducted as a life-long phase I/II trial, with dose escalation of rapamycin/everolimus until the side effects are reached in an individual patient. The tailored optimal dose (see Figure 2) should be determined individually for each patient and may vary widely. Doses and frequencies should be limited by the side effects: stomatitis/mucositis, anemia, thrombopenia, leukopenia, edema, and pneumonitis. To be safe, even mild hyperglycemia should be avoided or mitigated with metformin. Treatment is intended to be life-long, unless discontinued due to side effects.
Self-medication (even by physicians themselves) should be avoided and strongly discouraged. Instead, we need anti-aging clinics that implement the entire anti-aging recipe, including a complementary low carbohydrate diet and life style changes. Blood levels of rapamycin should be measured, as the rapamycin concentration in blood varies greatly among individuals taking the same dose. Doses of rapamycin should be tailored: personalized dosing and schedules. There is no shortage of potential patients who unfortunately already employ self-medication with rapamycin, but there is a shortage of physicians to treat them. Fortunately, a prototype clinic already functions in the USA, demonstrating that it is feasible from a regulatory standpoint (see Alan Green’s practice, Little Neck, NY). We cannot wait for results from others if we want to live longer and healthier ourselves. The time is now.
Disclaimer
This article is addressed to clinical scientists and physicians. It is intended for informational and educational purposes only. Medical doctors interested in this topic may e-mail the author at Blagosklonny@rapalogs.com
Footnotes
CONFLICTS OF INTEREST: The author declares no conflicts of interest.
REFERENCES
- 1.Johnson SC, Yanos ME, Kayser EB, Quintana A, Sangesland M, Castanza A, Uhde L, Hui J, Wall VZ, Gagnidze A, Oh K, Wasko BM, Ramos FJ, et al. mTOR inhibition alleviates mitochondrial disease in a mouse model of Leigh syndrome. Science. 2013; 342:1524–28. 10.1126/science.1244360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Blagosklonny MV. Aging and immortality: quasi-programmed senescence and its pharmacologic inhibition. Cell Cycle. 2006; 5:2087–102. 10.4161/cc.5.18.3288 [DOI] [PubMed] [Google Scholar]
- 3.Blagosklonny MV. An anti-aging drug today: from senescence-promoting genes to anti-aging pill. Drug Discov Today. 2007; 12:218–24. 10.1016/j.drudis.2007.01.004 [DOI] [PubMed] [Google Scholar]
- 4.Dao V, Liu Y, Pandeswara S, Svatek RS, Gelfond JA, Liu A, Hurez V, Curiel TJ. Immune-Stimulatory Effects of Rapamycin Are Mediated by Stimulation of Antitumor γδ T Cells. Cancer Res. 2016; 76:5970–82. 10.1158/0008-5472.CAN-16-0091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jung JW, Veitch M, Bridge JA, Overgaard NH, Cruz JL, Linedale R, Franklin ME, Saunders NA, Simpson F, Frazer IH, Steptoe RJ, Wells JW. Clinically-Relevant Rapamycin Treatment Regimens Enhance CD8+ Effector Memory T Cell Function In The Skin and Allow their Infiltration into Cutaneous Squamous Cell Carcinoma. OncoImmunology. 2018; 7:e1479627. 10.1080/2162402X.2018.1479627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bravo-San Pedro JM, Senovilla L. Immunostimulatory activity of lifespan-extending agents. Aging (Albany NY). 2013; 5:793–801. 10.18632/aging.100619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Svatek RS, Ji N, de Leon E, Mukherjee NZ, Kabra A, Hurez V, Nicolas M, Michalek JE, Javors M, Wheeler K, Sharp ZD, Livi CB, Shu ZJ, et al. Rapamycin Prevents Surgery-Induced Immune Dysfunction in Patients with Bladder Cancer. Cancer Immunol Res. 2019; 7:466–75. 10.1158/2326-6066.CIR-18-0336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mannick JB, Del Giudice G, Lattanzi M, Valiante NM, Praestgaard J, Huang B, Lonetto MA, Maecker HT, Kovarik J, Carson S, Glass DJ, Klickstein LB. mTOR inhibition improves immune function in the elderly. Sci Transl Med. 2014; 6:268ra179. 10.1126/scitranslmed.3009892 [DOI] [PubMed] [Google Scholar]
- 9.Mannick JB, Morris M, Hockey HP, Roma G, Beibel M, Kulmatycki K, Watkins M, Shavlakadze T, Zhou W, Quinn D, Glass DJ, Klickstein LB. TORC1 inhibition enhances immune function and reduces infections in the elderly. Sci Transl Med. 2018; 10:eaaq1564. 10.1126/scitranslmed.aaq1564 [DOI] [PubMed] [Google Scholar]
- 10.Demopoulos L, Polinsky M, Steele G, Mines D, Blum M, Caulfield M, Adamkovic A, Liu Q, Harler MB, Hahn C, Singh A. Reduced risk of cytomegalovirus infection in solid organ transplant recipients treated with sirolimus: a pooled analysis of clinical trials. Transplant Proc. 2008; 40:1407–10. 10.1016/j.transproceed.2008.03.084 [DOI] [PubMed] [Google Scholar]
- 11.Hill JA, Hummel M, Starling RC, Kobashigawa JA, Perrone SV, Arizón JM, Simonsen S, Abeywickrama KH, Bara C. A lower incidence of cytomegalovirus infection in de novo heart transplant recipients randomized to everolimus. Transplantation. 2007; 84:1436–42. 10.1097/01.tp.0000290686.68910.bd [DOI] [PubMed] [Google Scholar]
- 12.Pascual J, Royuela A, Fernández AM, Herrero I, Delgado JF, Solé A, Guirado L, Serrano T, de la Torre-Cisneros J, Moreno A, Cordero E, Gallego R, Lumbreras C, Aguado JM, and Spanish Society of Transplantation Virological and Immune Response Investigation Study Group. Role of mTOR inhibitors for the control of viral infection in solid organ transplant recipients. Transpl Infect Dis. 2016; 18:819–31. 10.1111/tid.12601 [DOI] [PubMed] [Google Scholar]
- 13.Keating R, Hertz T, Wehenkel M, Harris TL, Edwards BA, McClaren JL, Brown SA, Surman S, Wilson ZS, Bradley P, Hurwitz J, Chi H, Doherty PC, et al. The kinase mTOR modulates the antibody response to provide cross-protective immunity to lethal infection with influenza virus. Nat Immunol. 2013; 14:1266–76. 10.1038/ni.2741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Diken M, Kreiter S, Vascotto F, Selmi A, Attig S, Diekmann J, Huber C, Türeci Ö, Sahin U. mTOR inhibition improves antitumor effects of vaccination with antigen-encoding RNA. Cancer Immunol Res. 2013; 1:386–92. 10.1158/2326-6066.CIR-13-0046 [DOI] [PubMed] [Google Scholar]
- 15.Ferrer IR, Wagener ME, Robertson JM, Turner AP, Araki K, Ahmed R, Kirk AD, Larsen CP, Ford ML. Cutting edge: rapamycin augments pathogen-specific but not graft-reactive CD8+ T cell responses. J Immunol. 2010; 185:2004–08. 10.4049/jimmunol.1001176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hurez V, Dao V, Liu A, Pandeswara S, Gelfond J, Sun L, Bergman M, Orihuela CJ, Galvan V, Padrón Á, Drerup J, Liu Y, Hasty P, et al. Chronic mTOR inhibition in mice with rapamycin alters T, B, myeloid, and innate lymphoid cells and gut flora and prolongs life of immune-deficient mice. Aging Cell. 2015; 14:945–56. 10.1111/acel.12380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hinojosa CA, Mgbemena V, Van Roekel S, Austad SN, Miller RA, Bose S, Orihuela CJ. Enteric-delivered rapamycin enhances resistance of aged mice to pneumococcal pneumonia through reduced cellular senescence. Exp Gerontol. 2012; 47:958–65. 10.1016/j.exger.2012.08.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuss-Duerkop SK, Wang J, Mena I, White K, Metreveli G, Sakthivel R, Mata MA, Muñoz-Moreno R, Chen X, Krammer F, Diamond MS, Chen ZJ, García-Sastre A, Fontoura BM. Influenza virus differentially activates mTORC1 and mTORC2 signaling to maximize late stage replication. PLoS Pathog. 2017; 13:e1006635. 10.1371/journal.ppat.1006635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ranadheera C, Coombs KM, Kobasa D. Comprehending a Killer: The Akt/mTOR Signaling Pathways Are Temporally High-Jacked by the Highly Pathogenic 1918 Influenza Virus. EBioMedicine. 2018; 32:142–63. 10.1016/j.ebiom.2018.05.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mathew T, Kreis H, Friend P. Two-year incidence of malignancy in sirolimus-treated renal transplant recipients: results from five multicenter studies. Clin Transplant. 2004; 18:446–49. 10.1111/j.1399-0012.2004.00188.x [DOI] [PubMed] [Google Scholar]
- 21.Kauffman HM, Cherikh WS, Cheng Y, Hanto DW, Kahan BD. Maintenance immunosuppression with target-of-rapamycin inhibitors is associated with a reduced incidence of de novo malignancies. Transplantation. 2005; 80:883–89. 10.1097/01.TP.0000184006.43152.8D [DOI] [PubMed] [Google Scholar]
- 22.Campistol JM, Eris J, Oberbauer R, Friend P, Hutchison B, Morales JM, Claesson K, Stallone G, Russ G, Rostaing L, Kreis H, Burke JT, Brault Y, et al. Sirolimus therapy after early cyclosporine withdrawal reduces the risk for cancer in adult renal transplantation. J Am Soc Nephrol. 2006; 17:581–89. 10.1681/ASN.2005090993 [DOI] [PubMed] [Google Scholar]
- 23.Euvrard S, Morelon E, Rostaing L, Goffin E, Brocard A, Tromme I, Broeders N, del Marmol V, Chatelet V, Dompmartin A, Kessler M, Serra AL, Hofbauer GF, et al. , and TUMORAPA Study Group. Sirolimus and secondary skin-cancer prevention in kidney transplantation. N Engl J Med. 2012; 367:329–39. 10.1056/NEJMoa1204166 [DOI] [PubMed] [Google Scholar]
- 24.Dantal J, Morelon E, Rostaing L, Goffin E, Brocard A, Tromme I, Broeders N, Del Marmol V, Chatelet V, Dompmartin A, Kessler M, Serra A, Hofbauer GF, et al. , and TUMORAPA Study Group. Sirolimus for Secondary Prevention of Skin Cancer in Kidney Transplant Recipients: 5-Year Results. J Clin Oncol. 2018; 36:2612–20. 10.1200/JCO.2017.76.6691 [DOI] [PubMed] [Google Scholar]
- 25.Patlolla JM, Kopelovich L, Qian L, Zhang Y, Kumar G, Madka V, Mohammed A, Biddick L, Sadeghi M, Lightfoot S, Rao CV. Early and delayed intervention with rapamycin prevents NNK-induced lung adenocarcinoma in A/J mice. Oncol Rep. 2015; 34:2925–34. 10.3892/or.2015.4277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stallone G, Infante B, Prisciandaro C, Grandaliano G. mTOR and Aging: An Old Fashioned Dress. Int J Mol Sci. 2019; 20:E2774. 10.3390/ijms20112774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Blagosklonny MV. Prevention of cancer by inhibiting aging. Cancer Biol Ther. 2008; 7:1520–24. 10.4161/cbt.7.10.6663 [DOI] [PubMed] [Google Scholar]
- 28.Granville CA, Warfel N, Tsurutani J, Hollander MC, Robertson M, Fox SD, Veenstra TD, Issaq HJ, Linnoila RI, Dennis PA. Identification of a highly effective rapamycin schedule that markedly reduces the size, multiplicity, and phenotypic progression of tobacco carcinogen-induced murine lung tumors. Clin Cancer Res. 2007; 13:2281–89. 10.1158/1078-0432.CCR-06-2570 [DOI] [PubMed] [Google Scholar]
- 29.Kopelovich L, Fay JR, Sigman CC, Crowell JA. The mammalian target of rapamycin pathway as a potential target for cancer chemoprevention. Cancer Epidemiol Biomarkers Prev. 2007; 16:1330–40. 10.1158/1055-9965.EPI-07-0045 [DOI] [PubMed] [Google Scholar]
- 30.Mercier I, Camacho J, Titchen K, Gonzales DM, Quann K, Bryant KG, Molchansky A, Milliman JN, Whitaker-Menezes D, Sotgia F, Jasmin JF, Schwarting R, Pestell RG, et al. Caveolin-1 and accelerated host aging in the breast tumor microenvironment: chemoprevention with rapamycin, an mTOR inhibitor and anti-aging drug. Am J Pathol. 2012; 181:278–93. 10.1016/j.ajpath.2012.03.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kawabata S, Mercado-Matos JR, Hollander MC, Donahue D, Wilson W 3rd, Regales L, Butaney M, Pao W, Wong KK, Jänne PA, Dennis PA. Rapamycin prevents the development and progression of mutant epidermal growth factor receptor lung tumors with the acquired resistance mutation T790M. Cell Rep. 2014; 7:1824–32. 10.1016/j.celrep.2014.05.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Blagosklonny MV. Immunosuppressants in cancer prevention and therapy. OncoImmunology. 2013; 2:e26961. 10.4161/onci.26961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Antoch MP, Blagosklonny MV. Rapamycin extends maximal lifespan in cancer-prone mice. Am J Pathol. 2010; 176:2092–97. 10.2353/ajpath.2010.091050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Komarova EA, Antoch MP, Novototskaya LR, Chernova OB, Paszkiewicz G, Leontieva OV, Blagosklonny MV, Gudkov AV. Rapamycin extends lifespan and delays tumorigenesis in heterozygous p53+/- mice. Aging (Albany NY). 2012; 4:709–14. 10.18632/aging.100498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hasty P, Livi CB, Dodds SG, Jones D, Strong R, Javors M, Fischer KE, Sloane L, Murthy K, Hubbard G, Sun L, Hurez V, Curiel TJ, Sharp ZD. eRapa restores a normal life span in a FAP mouse model. Cancer Prev Res (Phila). 2014; 7:169–78. 10.1158/1940-6207.CAPR-13-0299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Livi CB, Hardman RL, Christy BA, Dodds SG, Jones D, Williams C, Strong R, Bokov A, Javors MA, Ikeno Y, Hubbard G, Hasty P, Sharp ZD. Rapamycin extends life span of Rb1+/- mice by inhibiting neuroendocrine tumors. Aging (Albany NY). 2013; 5:100–10. 10.18632/aging.100533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ehninger D, Neff F, Xie K. Longevity, aging and rapamycin. Cell Mol Life Sci. 2014; 71:4325–46. 10.1007/s00018-014-1677-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Blagosklonny MV. Fasting and rapamycin: diabetes versus benevolent glucose intolerance. Cell Death Dis. 2019; 10:607. 10.1038/s41419-019-1822-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Manning BD. Balancing Akt with S6K: implications for both metabolic diseases and tumorigenesis. J Cell Biol. 2004; 167:399–403. 10.1083/jcb.200408161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Um SH, D’Alessio D, Thomas G. Nutrient overload, insulin resistance, and ribosomal protein S6 kinase 1, S6K1. Cell Metab. 2006; 3:393–402. 10.1016/j.cmet.2006.05.003 [DOI] [PubMed] [Google Scholar]
- 41.Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell. 2006; 124:471–84. 10.1016/j.cell.2006.01.016 [DOI] [PubMed] [Google Scholar]
- 42.Inoki K, Mori H, Wang J, Suzuki T, Hong S, Yoshida S, Blattner SM, Ikenoue T, Rüegg MA, Hall MN, Kwiatkowski DJ, Rastaldi MP, Huber TB, et al. mTORC1 activation in podocytes is a critical step in the development of diabetic nephropathy in mice. J Clin Invest. 2011; 121:2181–96. 10.1172/JCI44771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zoncu R, Efeyan A, Sabatini DM. mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol. 2011; 12:21–35. 10.1038/nrm3025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cornu M, Albert V, Hall MN. mTOR in aging, metabolism, and cancer. Curr Opin Genet Dev. 2013; 23:53–62. 10.1016/j.gde.2012.12.005 [DOI] [PubMed] [Google Scholar]
- 45.Krebs M, Brunmair B, Brehm A, Artwohl M, Szendroedi J, Nowotny P, Roth E, Fürnsinn C, Promintzer M, Anderwald C, Bischof M, Roden M. The Mammalian target of rapamycin pathway regulates nutrient-sensitive glucose uptake in man. Diabetes. 2007; 56:1600–07. 10.2337/db06-1016 [DOI] [PubMed] [Google Scholar]
- 46.Tremblay F, Krebs M, Dombrowski L, Brehm A, Bernroider E, Roth E, Nowotny P, Waldhäusl W, Marette A, Roden M. Overactivation of S6 kinase 1 as a cause of human insulin resistance during increased amino acid availability. Diabetes. 2005; 54:2674–84. 10.2337/diabetes.54.9.2674 [DOI] [PubMed] [Google Scholar]
- 47.Zhou W, Ye S. Rapamycin improves insulin resistance and hepatic steatosis in type 2 diabetes rats through activation of autophagy. Cell Biol Int. 2018; 42:1282–91. 10.1002/cbin.11015 [DOI] [PubMed] [Google Scholar]
- 48.Ueno M, Carvalheira JB, Tambascia RC, Bezerra RM, Amaral ME, Carneiro EM, Folli F, Franchini KG, Saad MJ. Regulation of insulin signalling by hyperinsulinaemia: role of IRS-1/2 serine phosphorylation and the mTOR/p70 S6K pathway. Diabetologia. 2005; 48:506–18. 10.1007/s00125-004-1662-6 [DOI] [PubMed] [Google Scholar]
- 49.Reifsnyder PC, Flurkey K, Te A, Harrison DE. Rapamycin treatment benefits glucose metabolism in mouse models of type 2 diabetes. Aging (Albany NY). 2016; 8:3120–30. 10.18632/aging.101117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.He S, Zhang Y, Wang D, Tao K, Zhang S, Wei L, Chen Q. Rapamycin/GABA combination treatment ameliorates diabetes in NOD mice. Mol Immunol. 2016; 73:130–37. 10.1016/j.molimm.2016.01.008 [DOI] [PubMed] [Google Scholar]
- 51.Leontieva OV, Demidenko ZN, Blagosklonny MV. Rapamycin reverses insulin resistance (IR) in high-glucose medium without causing IR in normoglycemic medium. Cell Death Dis. 2014; 5:e1214. 10.1038/cddis.2014.178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Makki K, Taront S, Molendi-Coste O, Bouchaert E, Neve B, Eury E, Lobbens S, Labalette M, Duez H, Staels B, Dombrowicz D, Froguel P, Wolowczuk I. Beneficial metabolic effects of rapamycin are associated with enhanced regulatory cells in diet-induced obese mice. PLoS One. 2014; 9:e92684. 10.1371/journal.pone.0092684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yang SB, Lee HY, Young DM, Tien AC, Rowson-Baldwin A, Shu YY, Jan YN, Jan LY. Rapamycin induces glucose intolerance in mice by reducing islet mass, insulin content, and insulin sensitivity. J Mol Med (Berl). 2012; 90:575–85. 10.1007/s00109-011-0834-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lamming DW, Ye L, Katajisto P, Goncalves MD, Saitoh M, Stevens DM, Davis JG, Salmon AB, Richardson A, Ahima RS, Guertin DA, Sabatini DM, Baur JA. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science. 2012; 335:1638–43. 10.1126/science.1215135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Houde VP, Brûlé S, Festuccia WT, Blanchard PG, Bellmann K, Deshaies Y, Marette A. Chronic rapamycin treatment causes glucose intolerance and hyperlipidemia by upregulating hepatic gluconeogenesis and impairing lipid deposition in adipose tissue. Diabetes. 2010; 59:1338–48. 10.2337/db09-1324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Deblon N, Bourgoin L, Veyrat-Durebex C, Peyrou M, Vinciguerra M, Caillon A, Maeder C, Fournier M, Montet X, Rohner-Jeanrenaud F, Foti M. Chronic mTOR inhibition by rapamycin induces muscle insulin resistance despite weight loss in rats. Br J Pharmacol. 2012; 165:2325–40. 10.1111/j.1476-5381.2011.01716.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lundbaek K. Metabolic abnormalities in starvation diabetes. Yale J Biol Med. 1948; 20:533–44. [PMC free article] [PubMed] [Google Scholar]
- 58.Peters JP. Starvation Diabetes, The Reason for the Use of Glucose in the Treatment of Diabetic Acidosis. Yale J Biol Med. 1945; 17:705–26. [PMC free article] [PubMed] [Google Scholar]
- 59.Fontana L, Klein S, Holloszy JO. Effects of long-term calorie restriction and endurance exercise on glucose tolerance, insulin action, and adipokine production. Age (Dordr). 2010; 32:97–108. 10.1007/s11357-009-9118-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Steven S, Lim EL, Taylor R. Population response to information on reversibility of Type 2 diabetes. Diabet Med. 2013; 30:e135–38. 10.1111/dme.12116 [DOI] [PubMed] [Google Scholar]
- 61.Steven S, Taylor R. Restoring normoglycaemia by use of a very low calorie diet in long- and short-duration Type 2 diabetes. Diabet Med. 2015; 32:1149–55. 10.1111/dme.12722 [DOI] [PubMed] [Google Scholar]
- 62.Taylor R, Al-Mrabeh A, Zhyzhneuskaya S, Peters C, Barnes AC, Aribisala BS, Hollingsworth KG, Mathers JC, Sattar N, Lean ME. Remission of Human Type 2 Diabetes Requires Decrease in Liver and Pancreas Fat Content but Is Dependent upon Capacity for β Cell Recovery. Cell Metab. 2018; 28:547–556.e3. 10.1016/j.cmet.2018.07.003 [DOI] [PubMed] [Google Scholar]
- 63.Koffler M, Kisch ES. Starvation diet and very-low-calorie diets may induce insulin resistance and overt diabetes mellitus. J Diabetes Complications. 1996; 10:109–12. 10.1016/1056-8727(94)00077-8 [DOI] [PubMed] [Google Scholar]
- 64.Blagosklonny MV. The mystery of the ketogenic diet: benevolent pseudo-diabetes. Cell Cycle. 2019; 18:2157–63. 10.1080/15384101.2019.1644765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lamont BJ, Waters MF, Andrikopoulos S. A low-carbohydrate high-fat diet increases weight gain and does not improve glucose tolerance, insulin secretion or β-cell mass in NZO mice. Nutr Diabetes. 2016; 6:e194. 10.1038/nutd.2016.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schofield G, Henderson G, Crofts C, Thornley S. What are we to think when results from mouse research contradict those from human experiments and clinical practice? Nutr Diabetes. 2016; 6:e224. 10.1038/nutd.2016.31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Caminhotto RO, Lima FB. Low carbohydrate high fat diets: when models do not match reality. Arch Endocrinol Metab. 2016; 60:405–06. 10.1590/2359-3997000000177 [DOI] [PubMed] [Google Scholar]
- 68.Menendez JA, Cuyàs E, Folguera-Blasco N, Verdura S, Martin-Castillo B, Joven J, Alarcón T. In silico clinical trials for anti-aging therapies. Aging (Albany NY). 2019; 11:6591–601. 10.18632/aging.102180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Longo VD, Fontana L. Intermittent supplementation with rapamycin as a dietary restriction mimetic. Aging (Albany NY). 2011; 3:1039–40. 10.18632/aging.100401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Mercken EM, Carboneau BA, Krzysik-Walker SM, de Cabo R. Of mice and men: the benefits of caloric restriction, exercise, and mimetics. Ageing Res Rev. 2012; 11:390–98. 10.1016/j.arr.2011.11.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Roth GS, Ingram DK. Manipulation of health span and function by dietary caloric restriction mimetics. Ann N Y Acad Sci. 2016; 1363:5–10. 10.1111/nyas.12834 [DOI] [PubMed] [Google Scholar]
- 72.Blagosklonny MV. Rapamycin-induced glucose intolerance: hunger or starvation diabetes. Cell Cycle. 2011; 10:4217–24. 10.4161/cc.10.24.18595 [DOI] [PubMed] [Google Scholar]
- 73.Carter CS, Khamiss D, Matheny M, Toklu HZ, Kirichenko N, Strehler KY, Tümer N, Scarpace PJ, Morgan D. Rapamycin Versus Intermittent Feeding: Dissociable Effects on Physiological and Behavioral Outcomes When Initiated Early and Late in Life. J Gerontol A Biol Sci Med Sci. 2016; 71:866–75. 10.1093/gerona/glu238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gödel M, Hartleben B, Herbach N, Liu S, Zschiedrich S, Lu S, Debreczeni-Mór A, Lindenmeyer MT, Rastaldi MP, Hartleben G, Wiech T, Fornoni A, Nelson RG, et al. Role of mTOR in podocyte function and diabetic nephropathy in humans and mice. J Clin Invest. 2011; 121:2197–209. 10.1172/JCI44774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Nagai K, Matsubara T, Mima A, Sumi E, Kanamori H, Iehara N, Fukatsu A, Yanagita M, Nakano T, Ishimoto Y, Kita T, Doi T, Arai H. Gas6 induces Akt/mTOR-mediated mesangial hypertrophy in diabetic nephropathy. Kidney Int. 2005; 68:552–61. 10.1111/j.1523-1755.2005.00433.x [DOI] [PubMed] [Google Scholar]
- 76.Sataranatarajan K, Mariappan MM, Lee MJ, Feliers D, Choudhury GG, Barnes JL, Kasinath BS. Regulation of elongation phase of mRNA translation in diabetic nephropathy: amelioration by rapamycin. Am J Pathol. 2007; 171:1733–42. 10.2353/ajpath.2007.070412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wittmann S, Daniel C, Stief A, Vogelbacher R, Amann K, Hugo C. Long-term treatment of sirolimus but not cyclosporine ameliorates diabetic nephropathy in the rat. Transplantation. 2009; 87:1290–99. 10.1097/TP.0b013e3181a192bd [DOI] [PubMed] [Google Scholar]
- 78.Lloberas N, Cruzado JM, Franquesa M, Herrero-Fresneda I, Torras J, Alperovich G, Rama I, Vidal A, Grinyó JM. Mammalian target of rapamycin pathway blockade slows progression of diabetic kidney disease in rats. J Am Soc Nephrol. 2006; 17:1395–404. 10.1681/ASN.2005050549 [DOI] [PubMed] [Google Scholar]
- 79.Yang Y, Wang J, Qin L, Shou Z, Zhao J, Wang H, Chen Y, Chen J. Rapamycin prevents early steps of the development of diabetic nephropathy in rats. Am J Nephrol. 2007; 27:495–502. 10.1159/000106782 [DOI] [PubMed] [Google Scholar]
- 80.Sakaguchi M, Isono M, Isshiki K, Sugimoto T, Koya D, Kashiwagi A. Inhibition of mTOR signaling with rapamycin attenuates renal hypertrophy in the early diabetic mice. Biochem Biophys Res Commun. 2006; 340:296–301. 10.1016/j.bbrc.2005.12.012 [DOI] [PubMed] [Google Scholar]
- 81.Mori H, Inoki K, Masutani K, Wakabayashi Y, Komai K, Nakagawa R, Guan KL, Yoshimura A. The mTOR pathway is highly activated in diabetic nephropathy and rapamycin has a strong therapeutic potential. Biochem Biophys Res Commun. 2009; 384:471–75. 10.1016/j.bbrc.2009.04.136 [DOI] [PubMed] [Google Scholar]
- 82.Flaquer M, Lloberas N, Franquesa M, Torras J, Vidal A, Rosa JL, Herrero-Fresneda I, Grinyó JM, Cruzado JM. The combination of sirolimus and rosiglitazone produces a renoprotective effect on diabetic kidney disease in rats. Life Sci. 2010; 87:147–53. 10.1016/j.lfs.2010.06.004 [DOI] [PubMed] [Google Scholar]
- 83.Reifsnyder PC, Doty R, Harrison DE. Rapamycin ameliorates nephropathy despite elevating hyperglycemia in a polygenic mouse model of type 2 diabetes, NONcNZO10/LtJ. PLoS One. 2014; 9:e114324. 10.1371/journal.pone.0114324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lu MK, Gong XG, Guan KL. mTOR in podocyte function: is rapamycin good for diabetic nephropathy? Cell Cycle. 2011; 10:3415–16. 10.4161/cc.10.20.17686 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wahl PR, Serra AL, Le Hir M, Molle KD, Hall MN, Wüthrich RP. Inhibition of mTOR with sirolimus slows disease progression in Han:SPRD rats with autosomal dominant polycystic kidney disease (ADPKD). Nephrol Dial Transplant. 2006; 21:598–604. 10.1093/ndt/gfi181 [DOI] [PubMed] [Google Scholar]
- 86.Kraig E, Linehan LA, Liang H, Romo TQ, Liu Q, Wu Y, Benavides AD, Curiel TJ, Javors MA, Musi N, Chiodo L, Koek W, Gelfond JA, Kellogg DL Jr. A randomized control trial to establish the feasibility and safety of rapamycin treatment in an older human cohort: Immunological, physical performance, and cognitive effects. Exp Gerontol. 2018; 105:53–69. 10.1016/j.exger.2017.12.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Xu KY, Shameem R, Wu S. Risk of hyperglycemia attributable to everolimus in cancer patients: A meta-analysis. Acta Oncol. 2016; 55:1196–203. 10.3109/0284186X.2016.1168939 [DOI] [PubMed] [Google Scholar]
- 88.Bono P, Oudard S, Bodrogi I, Hutson TE, Escudier B, Machiels JP, Thompson JA, Figlin RA, Ravaud A, Basaran M, Porta C, Bracarda S, Brechenmacher T, et al. Outcomes in Patients With Metastatic Renal Cell Carcinoma Who Develop Everolimus-Related Hyperglycemia and Hypercholesterolemia: Combined Subgroup Analyses of the RECORD-1 and REACT Trials. Clin Genitourin Cancer. 2016; 14:406–14. 10.1016/j.clgc.2016.04.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tanimura J, Nakagawa H, Tanaka T, Kikuchi A, Osada S, Tanaka Y, Tokuyama K, Takamura T. The clinical course and potential underlying mechanisms of everolimus-induced hyperglycemia. Endocr J. 2019; 66:615–20. 10.1507/endocrj.EJ18-0542 [DOI] [PubMed] [Google Scholar]
- 90.Franco A, Más-Serrano P, Perez Contreras J, Jiménez L, Rodriguez D, Olivares J. Mammalian Target of Rapamycin Inhibitor Monotherapy: Efficacy in Renal Transplantation. Transplant Proc. 2015; 47:2364–67. 10.1016/j.transproceed.2015.09.006 [DOI] [PubMed] [Google Scholar]
- 91.Veroux M, Corona D, Giuffrida G, Gagliano M, Sorbello M, Virgilio C, Tallarita T, Zerbo D, Giaquinta A, Fiamingo P, Macarone M, Li Volti G, Caglia P, Veroux P. New-onset diabetes mellitus after kidney transplantation: the role of immunosuppression. Transplant Proc. 2008; 40:1885–87. 10.1016/j.transproceed.2008.06.005 [DOI] [PubMed] [Google Scholar]
- 92.Veroux M, Corona D, Giuffrida G, Gagliano M, Vizcarra D, Tallarita T, Zerbo D, Giaquinta A, Sorbello M, Macarone M, Veroux P. Sirolimus-based immunosuppression in kidney transplantation for type 2 diabetic nephropathy. Urol Int. 2010; 84:301–04. 10.1159/000288232 [DOI] [PubMed] [Google Scholar]
- 93.Kälble F, Seckinger J, Schaier M, Morath C, Schwenger V, Zeier M, Sommerer C. Switch to an everolimus-facilitated cyclosporine A sparing immunosuppression improves glycemic control in selected kidney transplant recipients. Clin Transplant. 2017; 31:e13024. 10.1111/ctr.13024 [DOI] [PubMed] [Google Scholar]
- 94.Krentz AJ, Wheeler DC. New-onset diabetes after transplantation: a threat to graft and patient survival. Lancet. 2005; 365:640–42. 10.1016/S0140-6736(05)70914-4 [DOI] [PubMed] [Google Scholar]
- 95.Nanmoku K, Shinzato T, Kubo T, Shimizu T, Kimura T, Yagisawa T. Steroid Withdrawal Using Everolimus in ABO-Incompatible Kidney Transplant Recipients With Post-Transplant Diabetes Mellitus. Transplant Proc. 2018; 50:1050–55. 10.1016/j.transproceed.2018.01.028 [DOI] [PubMed] [Google Scholar]
- 96.Veroux M, Tallarita T, Corona D, Sinagra N, Giaquinta A, Zerbo D, Guerrieri C, D’Assoro A, Cimino S, Veroux P. Conversion to sirolimus therapy in kidney transplant recipients with new onset diabetes mellitus after transplantation. Clin Dev Immunol. 2013; 2013:496974. 10.1155/2013/496974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Johnston O, Rose CL, Webster AC, Gill JS. Sirolimus is associated with new-onset diabetes in kidney transplant recipients. J Am Soc Nephrol. 2008; 19:1411–18. 10.1681/ASN.2007111202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Pavlakis M, Goldfarb-Rumyantzev AS. Diabetes after transplantation and sirolimus: what’s the connection? J Am Soc Nephrol. 2008; 19:1255–56. 10.1681/ASN.2008050474 [DOI] [PubMed] [Google Scholar]
- 99.Cohen EE, Wu K, Hartford C, Kocherginsky M, Eaton KN, Zha Y, Nallari A, Maitland ML, Fox-Kay K, Moshier K, House L, Ramirez J, Undevia SD, et al. Phase I studies of sirolimus alone or in combination with pharmacokinetic modulators in advanced cancer patients. Clin Cancer Res. 2012; 18:4785–93. 10.1158/1078-0432.CCR-12-0110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Crutchlow MF, Bloom RD. Transplant-associated hyperglycemia: a new look at an old problem. Clin J Am Soc Nephrol. 2007; 2:343–55. 10.2215/CJN.03671106 [DOI] [PubMed] [Google Scholar]
- 101.Cuzick J, Thorat MA, Bosetti C, Brown PH, Burn J, Cook NR, Ford LG, Jacobs EJ, Jankowski JA, La Vecchia C, Law M, Meyskens F, Rothwell PM, et al. Estimates of benefits and harms of prophylactic use of aspirin in the general population. Ann Oncol. 2015; 26:47–57. 10.1093/annonc/mdu225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Thorat MA, Cuzick J. Prophylactic use of aspirin: systematic review of harms and approaches to mitigation in the general population. Eur J Epidemiol. 2015; 30:5–18. 10.1007/s10654-014-9971-7 [DOI] [PubMed] [Google Scholar]
- 103.Ceschi A, Heistermann E, Gros S, Reichert C, Kupferschmidt H, Banner NR, Krähenbühl S, Taegtmeyer AB. Acute sirolimus overdose: a multicenter case series. PLoS One. 2015; 10:e0128033. 10.1371/journal.pone.0128033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Brattström C, Säwe J, Jansson B, Lönnebo A, Nordin J, Zimmerman JJ, Burke JT, Groth CG. Pharmacokinetics and safety of single oral doses of sirolimus (rapamycin) in healthy male volunteers. Ther Drug Monit. 2000; 22:537–44. 10.1097/00007691-200010000-00006 [DOI] [PubMed] [Google Scholar]
- 105.Weiner SM, Sellin L, Vonend O, Schenker P, Buchner NJ, Flecken M, Viebahn R, Rump LC. Pneumonitis associated with sirolimus: clinical characteristics, risk factors and outcome--a single-centre experience and review of the literature. Nephrol Dial Transplant. 2007; 22:3631–37. 10.1093/ndt/gfm420 [DOI] [PubMed] [Google Scholar]
- 106.Dunn JL, Kartchner LB, Gast K, Sessions M, Hunter RA, Thurlow L, Richardson A, Schoenfisch M, Cairns BA, Maile R. Mammalian target of rapamycin regulates a hyperresponsive state in pulmonary neutrophils late after burn injury. J Leukoc Biol. 2018; 103:909–18. 10.1002/JLB.3AB0616-251RRR [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Hebert M, Licursi M, Jensen B, Baker A, Milway S, Malsbury C, Grant VL, Adamec R, Hirasawa M, Blundell J. Single rapamycin administration induces prolonged downward shift in defended body weight in rats. PLoS One. 2014; 9:e93691. 10.1371/journal.pone.0093691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Chen C, Liu Y, Liu Y, Zheng P. mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci Signal. 2009; 2:ra75. 10.1126/scisignal.2000559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Bitto A, Ito TK, Pineda VV, LeTexier NJ, Huang HZ, Sutlief E, Tung H, Vizzini N, Chen B, Smith K, Meza D, Yajima M, Beyer RP, et al. Transient rapamycin treatment can increase lifespan and healthspan in middle-aged mice. eLife. 2016; 5:5. 10.7554/eLife.16351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Johnson SC, Kaeberlein M. Rapamycin in aging and disease: maximizing efficacy while minimizing side effects. Oncotarget. 2016; 7:44876–78. 10.18632/oncotarget.10381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Johnson SC, Yanos ME, Bitto A, Castanza A, Gagnidze A, Gonzalez B, Gupta K, Hui J, Jarvie C, Johnson BM, Letexier N, McCanta L, Sangesland M, et al. Dose-dependent effects of mTOR inhibition on weight and mitochondrial disease in mice. Front Genet. 2015; 6:247. 10.3389/fgene.2015.00247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Leontieva OV, Paszkiewicz GM, Blagosklonny MV. Comparison of rapamycin schedules in mice on high-fat diet. Cell Cycle. 2014; 13:3350–56. 10.4161/15384101.2014.970491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Kaeberlein M. Rapamycin and ageing: when, for how long, and how much? J Genet Genomics. 2014; 41:459–63. 10.1016/j.jgg.2014.06.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Blagosklonny MV. Aging, stem cells, and mammalian target of rapamycin: a prospect of pharmacologic rejuvenation of aging stem cells. Rejuvenation Res. 2008; 11:801–08. 10.1089/rej.2008.0722 [DOI] [PubMed] [Google Scholar]
- 115.Arriola Apelo SI, Pumper CP, Baar EL, Cummings NE, Lamming DW. Intermittent Administration of Rapamycin Extends the Life Span of Female C57BL/6J Mice. J Gerontol A Biol Sci Med Sci. 2016; 71:876–81. 10.1093/gerona/glw064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Harman D, Harman H. “I thought, thought, thought for four months in vain and suddenly the idea came”--an interview with Denham and Helen Harman. Interview by K. Kitani and G.O. Ivy. Biogerontology. 2003; 4:401–12. 10.1023/B:BGEN.0000006561.15498.68 [DOI] [PubMed] [Google Scholar]
- 117.Blagosklonny MV. Aging: ROS or TOR. Cell Cycle. 2008; 7:3344–54. 10.4161/cc.7.21.6965 [DOI] [PubMed] [Google Scholar]
- 118.Lapointe J, Hekimi S. When a theory of aging ages badly. Cell Mol Life Sci. 2010; 67:1–8. 10.1007/s00018-009-0138-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Ristow M, Schmeisser K. Mitohormesis: Promoting Health and Lifespan by Increased Levels of Reactive Oxygen Species (ROS). Dose Response. 2014; 12:288–341. 10.2203/dose-response.13-035.Ristow [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Gems D, Partridge L. Genetics of longevity in model organisms: debates and paradigm shifts. Annu Rev Physiol. 2013; 75:621–44. 10.1146/annurev-physiol-030212-183712 [DOI] [PubMed] [Google Scholar]
- 121.Gems D, de la Guardia Y. Alternative Perspectives on Aging in Caenorhabditis elegans: Reactive Oxygen Species or Hyperfunction? Antioxid Redox Signal. 2013; 19:321–29. 10.1089/ars.2012.4840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Howes RM. The free radical fantasy: a panoply of paradoxes. Ann N Y Acad Sci. 2006; 1067:22–26. 10.1196/annals.1354.004 [DOI] [PubMed] [Google Scholar]
- 123.Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG, Gluud C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: systematic review and meta-analysis. JAMA. 2007; 297:842–57. 10.1001/jama.297.8.842 [DOI] [PubMed] [Google Scholar]
- 124.Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG, Gluud C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst Rev. 2012; 3:CD007176. 10.1002/14651858.CD007176.pub2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Bjelakovic G, Nikolova D, Gluud C. Meta-regression analyses, meta-analyses, and trial sequential analyses of the effects of supplementation with beta-carotene, vitamin A, and vitamin E singly or in different combinations on all-cause mortality: do we have evidence for lack of harm? PLoS One. 2013; 8:e74558. 10.1371/journal.pone.0074558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Bjelakovic G, Nikolova D, Gluud C. Antioxidant supplements and mortality. Curr Opin Clin Nutr Metab Care. 2014; 17:40–44. 10.1097/MCO.0000000000000009 [DOI] [PubMed] [Google Scholar]
- 127.Chong EW, Wong TY, Kreis AJ, Simpson JA, Guymer RH. Dietary antioxidants and primary prevention of age related macular degeneration: systematic review and meta-analysis. BMJ. 2007; 335:755. 10.1136/bmj.39350.500428.47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Myung SK, Ju W, Cho B, Oh SW, Park SM, Koo BK, Park BJ, and Korean Meta-Analysis Study Group. Efficacy of vitamin and antioxidant supplements in prevention of cardiovascular disease: systematic review and meta-analysis of randomised controlled trials. BMJ. 2013; 346:f10. 10.1136/bmj.f10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Ristow M, Zarse K, Oberbach A, Klöting N, Birringer M, Kiehntopf M, Stumvoll M, Kahn CR, Blüher M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc Natl Acad Sci USA. 2009; 106:8665–70. 10.1073/pnas.0903485106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ristow M, Schmeisser S. Extending life span by increasing oxidative stress. Free Radic Biol Med. 2011; 51:327–36. 10.1016/j.freeradbiomed.2011.05.010 [DOI] [PubMed] [Google Scholar]
- 131.Omenn GS, Goodman GE, Thornquist MD, Balmes J, Cullen MR, Glass A, Keogh JP, Meyskens FL Jr, Valanis B, Williams JH Jr, Barnhart S, Hammar S. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N Engl J Med. 1996; 334:1150–55. 10.1056/NEJM199605023341802 [DOI] [PubMed] [Google Scholar]
- 132.Goodman GE, Thornquist MD, Balmes J, Cullen MR, Meyskens FL Jr, Omenn GS, Valanis B, Williams JH Jr. The Beta-Carotene and Retinol Efficacy Trial: incidence of lung cancer and cardiovascular disease mortality during 6-year follow-up after stopping beta-carotene and retinol supplements. J Natl Cancer Inst. 2004; 96:1743–50. 10.1093/jnci/djh320 [DOI] [PubMed] [Google Scholar]
- 133.Wright ME, Virtamo J, Hartman AM, Pietinen P, Edwards BK, Taylor PR, Huttunen JK, Albanes D. Effects of alpha-tocopherol and beta-carotene supplementation on upper aerodigestive tract cancers in a large, randomized controlled trial. Cancer. 2007; 109:891–98. 10.1002/cncr.22482 [DOI] [PubMed] [Google Scholar]
- 134.Piskounova E, Agathocleous M, Murphy MM, Hu Z, Huddlestun SE, Zhao Z, Leitch AM, Johnson TM, DeBerardinis RJ, Morrison SJ. Oxidative stress inhibits distant metastasis by human melanoma cells. Nature. 2015; 527:186–91. 10.1038/nature15726 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Sayin VI, Ibrahim MX, Larsson E, Nilsson JA, Lindahl P, Bergo MO. Antioxidants accelerate lung cancer progression in mice. Sci Transl Med. 2014; 6:221ra15. 10.1126/scitranslmed.3007653 [DOI] [PubMed] [Google Scholar]
- 136.Le Gal K, Ibrahim MX, Wiel C, Sayin VI, Akula MK, Karlsson C, Dalin MG, Akyürek LM, Lindahl P, Nilsson J, Bergo MO. Antioxidants can increase melanoma metastasis in mice. Sci Transl Med. 2015; 7:308re8. 10.1126/scitranslmed.aad3740 [DOI] [PubMed] [Google Scholar]
- 137.Aguiar-Oliveira MH, Bartke A. Growth hormone deficiency: health and longevity. Endocr Rev. 2019; 40:575–601. 10.1210/er.2018-00216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Panici JA, Harper JM, Miller RA, Bartke A, Spong A, Masternak MM. Early life growth hormone treatment shortens longevity and decreases cellular stress resistance in long-lived mutant mice. FASEB J. 2010; 24:5073–79. 10.1096/fj.10-163253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Bartke A, Quainoo N. Impact of Growth Hormone-Related Mutations on Mammalian Aging. Front Genet. 2018; 9:586. 10.3389/fgene.2018.00586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Rudman D, Feller AG, Nagraj HS, Gergans GA, Lalitha PY, Goldberg AF, Schlenker RA, Cohn L, Rudman IW, Mattson DE. Effects of human growth hormone in men over 60 years old. N Engl J Med. 1990; 323:1–6. 10.1056/NEJM199007053230101 [DOI] [PubMed] [Google Scholar]
- 141.Vance ML. Can growth hormone prevent aging? N Engl J Med. 2003; 348:779–80. 10.1056/NEJMp020186 [DOI] [PubMed] [Google Scholar]
- 142.Kaeberlein M, Powers RW 3rd, Steffen KK, Westman EA, Hu D, Dang N, Kerr EO, Kirkland KT, Fields S, Kennedy BK. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science. 2005; 310:1193–96. 10.1126/science.1115535 [DOI] [PubMed] [Google Scholar]
- 143.Vellai T, Takacs-Vellai K, Zhang Y, Kovacs AL, Orosz L, Müller F. Genetics: influence of TOR kinase on lifespan in C. elegans. Nature. 2003; 426:620. 10.1038/426620a [DOI] [PubMed] [Google Scholar]
- 144.Kapahi P, Zid BM, Harper T, Koslover D, Sapin V, Benzer S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr Biol. 2004; 14:885–90. 10.1016/j.cub.2004.03.059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Stanfel MN, Shamieh LS, Kaeberlein M, Kennedy BK. The TOR pathway comes of age. Biochim Biophys Acta. 2009; 1790:1067–74. 10.1016/j.bbagen.2009.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Blagosklonny MV. Cell senescence and hypermitogenic arrest. EMBO Rep. 2003; 4:358–62. 10.1038/sj.embor.embor806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Demidenko ZN, Blagosklonny MV. Growth stimulation leads to cellular senescence when the cell cycle is blocked. Cell Cycle. 2008; 7:3355–61. 10.4161/cc.7.21.6919 [DOI] [PubMed] [Google Scholar]
- 148.Blagosklonny MV. Validation of anti-aging drugs by treating age-related diseases. Aging (Albany NY). 2009; 1:281–88. 10.18632/aging.100034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Blagosklonny MV. Answering the ultimate question “what is the proximal cause of aging?”. Aging (Albany NY). 2012; 4:861–77. 10.18632/aging.100525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Blagosklonny MV. Disease or not, aging is easily treatable. Aging (Albany NY). 2018; 10:3067–78. 10.18632/aging.101647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Blagosklonny MV. Aging is not programmed: genetic pseudo-program is a shadow of developmental growth. Cell Cycle. 2013; 12:3736–42. 10.4161/cc.27188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Wang H, Zhang Z, Gems D. Monsters in the uterus: teratoma-like tumors in senescent C. elegans result from a parthenogenetic quasi-program. Aging (Albany NY). 2018; 10:1188–89. 10.18632/aging.101486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Blagosklonny MV. Rapamycin and quasi-programmed aging: four years later. Cell Cycle. 2010; 9:1859–62. 10.4161/cc.9.10.11872 [DOI] [PubMed] [Google Scholar]
- 154.Blagosklonny MV. From rapalogs to anti-aging formula. Oncotarget. 2017; 8:35492–507. 10.18632/oncotarget.18033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460:392–95. 10.1038/nature08221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Dodds SG, Livi CB, Parihar M, Hsu HK, Benavides AD, Morris J, Javors M, Strong R, Christy B, Hasty P, Sharp ZD. Adaptations to chronic rapamycin in mice. Pathobiol Aging Age Relat Dis. 2016; 6:31688. 10.3402/pba.v6.31688 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Swindell WR. Rapamycin in mice. Aging (Albany NY). 2017; 9:1941–42. 10.18632/aging.101289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Blagosklonny MV. Prospective treatment of age-related diseases by slowing down aging. Am J Pathol. 2012; 181:1142–46. 10.1016/j.ajpath.2012.06.024 [DOI] [PubMed] [Google Scholar]
- 159.Blagosklonny MV. Koschei the immortal and anti-aging drugs. Cell Death Dis. 2014; 5:e1552. 10.1038/cddis.2014.520 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Cabreiro F, Au C, Leung KY, Vergara-Irigaray N, Cochemé HM, Noori T, Weinkove D, Schuster E, Greene ND, Gems D. Metformin retards aging in C. elegans by altering microbial folate and methionine metabolism. Cell. 2013; 153:228–39. 10.1016/j.cell.2013.02.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Chen J, Ou Y, Li Y, Hu S, Shao LW, Liu Y. Metformin extends C. elegans lifespan through lysosomal pathway. eLife. 2017; 6:6. 10.7554/eLife.31268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Santos EL, de Picoli Souza K, da Silva ED, Batista EC, Martins PJ, D’Almeida V, Pesquero JB. Long term treatment with ACE inhibitor enalapril decreases body weight gain and increases life span in rats. Biochem Pharmacol. 2009; 78:951–58. 10.1016/j.bcp.2009.06.018 [DOI] [PubMed] [Google Scholar]
- 163.Anisimov VN. Metformin: do we finally have an anti-aging drug? Cell Cycle. 2013; 12:3483–89. 10.4161/cc.26928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Anisimov VN. Metformin for cancer and aging prevention: is it a time to make the long story short? Oncotarget. 2015; 6:39398–407. 10.18632/oncotarget.6347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Haukka J, Niskanen L, Auvinen A. Risk of Cause-Specific Death in Individuals with Cancer-Modifying Role Diabetes, Statins and Metformin. Int J Cancer. 2017; 141:2437–49. 10.1002/ijc.31016 [DOI] [PubMed] [Google Scholar]
- 166.Heckman-Stoddard BM, DeCensi A, Sahasrabuddhe VV, Ford LG. Repurposing metformin for the prevention of cancer and cancer recurrence. Diabetologia. 2017; 60:1639–47. 10.1007/s00125-017-4372-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Pietrocola F, Kroemer G. Metformin: a metabolic modulator. Oncotarget. 2017; 8:9017–20. 10.18632/oncotarget.14794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Romero R, Erez O, Hüttemann M, Maymon E, Panaitescu B, Conde-Agudelo A, Pacora P, Yoon BH, Grossman LI. Metformin, the aspirin of the 21st century: its role in gestational diabetes mellitus, prevention of preeclampsia and cancer, and the promotion of longevity. Am J Obstet Gynecol. 2017; 217:282–302. 10.1016/j.ajog.2017.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Cuzick J. Progress in preventive therapy for cancer: a reminiscence and personal viewpoint. Br J Cancer. 2018; 118:1155–61. 10.1038/s41416-018-0039-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Islam BN, Sharman SK, Hou Y, Bridges AE, Singh N, Kim S, Kolhe R, Trillo-Tinoco J, Rodriguez PC, Berger FG, Sridhar S, Browning DD. Sildenafil Suppresses Inflammation-Driven Colorectal Cancer in Mice. Cancer Prev Res (Phila). 2017; 10:377–88. 10.1158/1940-6207.CAPR-17-0015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Weiss R, Fernandez E, Liu Y, Strong R, Salmon AB. Metformin reduces glucose intolerance caused by rapamycin treatment in genetically heterogeneous female mice. Aging (Albany NY). 2018; 10:386–401. 10.18632/aging.101401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Taylor DH Jr, Hasselblad V, Henley SJ, Thun MJ, Sloan FA. Benefits of smoking cessation for longevity. Am J Public Health. 2002; 92:990–96. 10.2105/AJPH.92.6.990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Lipman RD, Smith DE, Bronson RT, Blumberg J. Is late-life caloric restriction beneficial? Aging (Milano). 1995; 7:136–39. [DOI] [PubMed] [Google Scholar]
- 174.Dirks AJ, Leeuwenburgh C. Caloric restriction in humans: potential pitfalls and health concerns. Mech Ageing Dev. 2006; 127:1–7. 10.1016/j.mad.2005.09.001 [DOI] [PubMed] [Google Scholar]
- 175.Goto S. Health span extension by later-life caloric or dietary restriction: a view based on rodent studies. Biogerontology. 2006; 7:135–38. 10.1007/s10522-006-9011-4 [DOI] [PubMed] [Google Scholar]
- 176.Newman AB, Yanez D, Harris T, Duxbury A, Enright PL, Fried LP, and Cardiovascular Study Research Group. Weight change in old age and its association with mortality. J Am Geriatr Soc. 2001; 49:1309–18. 10.1046/j.1532-5415.2001.49258.x [DOI] [PubMed] [Google Scholar]
- 177.Lee CG, Boyko EJ, Nielson CM, Stefanick ML, Bauer DC, Hoffman AR, Dam TT, Lapidus JA, Cawthon PM, Ensrud KE, Orwoll ES, and Osteoporotic Fractures in Men Study Group. Mortality risk in older men associated with changes in weight, lean mass, and fat mass. J Am Geriatr Soc. 2011; 59:233–40. 10.1111/j.1532-5415.2010.03245.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Wannamethee SG, Shaper AG, Lennon L. Reasons for intentional weight loss, unintentional weight loss, and mortality in older men. Arch Intern Med. 2005; 165:1035–40. 10.1001/archinte.165.9.1035 [DOI] [PubMed] [Google Scholar]
- 179.Sengupta S, Peterson TR, Laplante M, Oh S, Sabatini DM. mTORC1 controls fasting-induced ketogenesis and its modulation by ageing. Nature. 2010; 468:1100–04. 10.1038/nature09584 [DOI] [PubMed] [Google Scholar]
- 180.White Z, White RB, McMahon C, Grounds MD, Shavlakadze T. High mTORC1 signaling is maintained, while protein degradation pathways are perturbed in old murine skeletal muscles in the fasted state. Int J Biochem Cell Biol. 2016; 78:10–21. 10.1016/j.biocel.2016.06.012 [DOI] [PubMed] [Google Scholar]
- 181.Flynn JM, O’Leary MN, Zambataro CA, Academia EC, Presley MP, Garrett BJ, Zykovich A, Mooney SD, Strong R, Rosen CJ, Kapahi P, Nelson MD, Kennedy BK, Melov S. Late-life rapamycin treatment reverses age-related heart dysfunction. Aging Cell. 2013; 12:851–62. 10.1111/acel.12109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Unnikrishnan A, Kurup K, Salmon AB, Richardson A. Is Rapamycin a Dietary Restriction Mimetic? J Gerontol A Biol Sci Med Sci. 2019glz060. 10.1093/gerona/glz060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Fok WC, Zhang Y, Salmon AB, Bhattacharya A, Gunda R, Jones D, Ward W, Fisher K, Richardson A, Pérez VI. Short-term treatment with rapamycin and dietary restriction have overlapping and distinctive effects in young mice. J Gerontol A Biol Sci Med Sci. 2013; 68:108–16. 10.1093/gerona/gls127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Fok WC, Bokov A, Gelfond J, Yu Z, Zhang Y, Doderer M, Chen Y, Javors M, Wood WH 3rd, Zhang Y, Becker KG, Richardson A, Pérez VI. Combined treatment of rapamycin and dietary restriction has a larger effect on the transcriptome and metabolome of liver. Aging Cell. 2014; 13:311–19. 10.1111/acel.12175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Yu Z, Wang R, Fok WC, Coles A, Salmon AB, Pérez VI. Rapamycin and dietary restriction induce metabolically distinctive changes in mouse liver. J Gerontol A Biol Sci Med Sci. 2015; 70:410–20. 10.1093/gerona/glu053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Karunadharma PP, Basisty N, Dai DF, Chiao YA, Quarles EK, Hsieh EJ, Crispin D, Bielas JH, Ericson NG, Beyer RP, MacKay VL, MacCoss MJ, Rabinovitch PS. Subacute calorie restriction and rapamycin discordantly alter mouse liver proteome homeostasis and reverse aging effects. Aging Cell. 2015; 14:547–57. 10.1111/acel.12317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Ye L, Widlund AL, Sims CA, Lamming DW, Guan Y, Davis JG, Sabatini DM, Harrison DE, Vang O, Baur JA. Rapamycin doses sufficient to extend lifespan do not compromise muscle mitochondrial content or endurance. Aging (Albany NY). 2013; 5:539–50. 10.18632/aging.100576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Tang H, Shrager JB, Goldman D. Rapamycin protects aging muscle. Aging (Albany NY). 2019; 11:5868–70. 10.18632/aging.102176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Joseph GA, Wang SX, Jacobs CE, Zhou W, Kimble GC, Tse HW, Eash JK, Shavlakadze T, Glass DJ. Partial inhibition of mTORC1 in aged rats counteracts the decline in muscle mass and reverses molecular signaling associated with sarcopenia. Mol Cell Biol. 2019; 39:e00141-19. 10.1128/MCB.00141-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Tang H, Inoki K, Brooks SV, Okazawa H, Lee M, Wang J, Kim M, Kennedy CL, Macpherson PC, Ji X, Van Roekel S, Fraga DA, Wang K, et al. mTORC1 underlies age-related muscle fiber damage and loss by inducing oxidative stress and catabolism. Aging Cell. 2019; 18:e12943. 10.1111/acel.12943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Aliper A, Jellen L, Cortese F, Artemov A, Karpinsky-Semper D, Moskalev A, Swick AG, Zhavoronkov A. Towards natural mimetics of metformin and rapamycin. Aging (Albany NY). 2017; 9:2245–68. 10.18632/aging.101319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Blagosklonny MV. How to save Medicare: the anti-aging remedy. Aging (Albany NY). 2012; 4:547–52. 10.18632/aging.100479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.An JY, Quarles EK, Mekvanich S, Kang A, Liu A, Santos D, Miller RA, Rabinovitch PS, Cox TC, Kaeberlein M. Rapamycin treatment attenuates age-associated periodontitis in mice. Geroscience. 2017; 39:457–63. 10.1007/s11357-017-9994-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Carosi JM, Sargeant TJ. Rapamycin and Alzheimer disease: a double-edged sword? Autophagy. 2019; 15:1460–62. 10.1080/15548627.2019.1615823 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Whyte LS, Lau AA, Hemsley KM, Hopwood JJ, Sargeant TJ. Endo-lysosomal and autophagic dysfunction: a driving factor in Alzheimer’s disease? J Neurochem. 2017; 140:703–17. 10.1111/jnc.13935 [DOI] [PubMed] [Google Scholar]
- 196.Leontieva OV, Blagosklonny MV. Gerosuppression by pan-mTOR inhibitors. Aging (Albany NY). 2016; 8:3535–51. 10.18632/aging.101155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Walters HE, Deneka-Hannemann S, Cox LS. Reversal of phenotypes of cellular senescence by pan-mTOR inhibition. Aging (Albany NY). 2016; 8:231–44. 10.18632/aging.100872 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Demidenko ZN, Korotchkina LG, Gudkov AV, Blagosklonny MV. Paradoxical suppression of cellular senescence by p53. Proc Natl Acad Sci USA. 2010; 107:9660–64. 10.1073/pnas.1002298107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Christy B, Demaria M, Campisi J, Huang J, Jones D, Dodds SG, Williams C, Hubbard G, Livi CB, Gao X, Weintraub S, Curiel T, Sharp ZD, Hasty P. p53 and rapamycin are additive. Oncotarget. 2015; 6:15802–13. 10.18632/oncotarget.4602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Demidenko ZN, Shtutman M, Blagosklonny MV. Pharmacologic inhibition of MEK and PI-3K converges on the mTOR/S6 pathway to decelerate cellular senescence. Cell Cycle. 2009; 8:1896–900. 10.4161/cc.8.12.8809 [DOI] [PubMed] [Google Scholar]
- 201.Castillo-Quan JI, Tain LS, Kinghorn KJ, Li L, Grönke S, Hinze Y, Blackwell TK, Bjedov I, Partridge L. A triple drug combination targeting components of the nutrient-sensing network maximizes longevity. Proc Natl Acad Sci USA. 2019; [Epub ahead of print]. 10.1073/pnas.1913212116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Castillo-Quan JI, Li L, Kinghorn KJ, Ivanov DK, Tain LS, Slack C, Kerr F, Nespital T, Thornton J, Hardy J, Bjedov I, Partridge L. Lithium Promotes Longevity through GSK3/NRF2-Dependent Hormesis. Cell Rep. 2016; 15:638–50. 10.1016/j.celrep.2016.03.041 [DOI] [PMC free article] [PubMed] [Google Scholar]