

Abstract
Key message
Subgroup IVc basic helix-loop-helix transcription factors OsbHLH058 and OsbHLH059 positively regulate major iron deficiency responses in rice in a similar but distinct manner, putatively under partial control by OsHRZs.
Abstract
Under low iron availability, plants transcriptionally induce the expression of genes involved in iron uptake and translocation. OsHRZ1 and OsHRZ2 ubiquitin ligases negatively regulate this iron deficiency response in rice. The basic helix-loop-helix (bHLH) transcription factor OsbHLH060 interacts with OsHRZ1, and positively regulates iron deficiency-inducible genes. However, the functions of three other subgroup IVc bHLH transcription factors in rice, OsbHLH057, OsbHLH058, and OsbHLH059, have not yet been characterized. In the present study, we investigated the functions of OsbHLH058 and OsbHLH059 related to iron deficiency response. OsbHLH058 expression was repressed under iron deficiency, whereas the expression of OsbHLH057 and OsbHLH060 was moderately induced. Yeast two-hybrid analysis indicated that OsbHLH058 interacts with OsHRZ1 and OsHRZ2 more strongly than OsbHLH060, whereas OsbHLH059 showed no interaction. An in vitro ubiquitination assay detected no OsbHLH058 and OsbHLH060 ubiquitination by OsHRZ1 and OsHRZ2. Transgenic rice lines overexpressing OsbHLH058 showed tolerance for iron deficiency and higher iron concentration in seeds. These lines also showed enhanced expression of many iron deficiency-inducible genes involved in iron uptake and translocation under iron-sufficient conditions. Conversely, OsbHLH058 knockdown lines showed susceptibility to iron deficiency and reduced expression of many iron deficiency-inducible genes. OsbHLH059 knockdown lines were also susceptible to iron deficiency, and formed characteristic brownish regions in iron-deficient new leaves. OsbHLH059 knockdown lines also showed reduced expression of many iron deficiency-inducible genes. These results indicate that OsbHLH058 and OsbHLH059 positively regulate major iron deficiency responses in a similar but distinct manner, and that this function may be partially controlled by OsHRZs.
Electronic supplementary material
The online version of this article (10.1007/s11103-019-00917-8) contains supplementary material, which is available to authorized users.
Keywords: bHLH transcription factor, HRZ ubiquitin ligase, Iron deficiency, Rice
Introduction
Iron (Fe) is an essential element for living organisms, functioning as an essential component of prosthetic groups containing heme and Fe-sulfur clusters, and also as free Fe ions that bind to proteins for specific functions. Both Fe uptake and internal translocation are crucial for Fe homeostasis in multicellular organisms, including animals and plants. Plants take up Fe from the rhizosphere, where Fe is abundant but only slightly soluble. Fe solubility and, thus, availability is especially low in aerobic and high-pH soils, including calcareous soils, which cover one-third of cultivated areas globally, severely limiting plant productivity and nutritional quality (Marschner 1995; Briat et al. 2015). Low Fe content in staple foods such as polished rice (Oryza sativa L.) is also a main cause of human anemia, which affects at least 2 billion people worldwide (Murgia et al. 2012; Briat et al. 2015). Therefore, understanding the mechanisms of plant Fe uptake and translocation is important for improving plant productivity in problem soils and Fe nutritional quality for the human diet.
Graminaceous plants such as rice and barley utilize a unique Fe uptake system characterized by the production of high-affinity ferric Fe [Fe(III)] chelators, which are designated mugineic acid family phytosiderophores (MAs); these solubilize Fe(III) in the rhizosphere, and the resulting Fe(III)-MA complexes are taken up by roots (Takagi 1976). In contrast, non-graminaceous plants rely on the reduction of rhizospheric Fe(III) and subsequent uptake of ferrous Fe ion (Fe2+) for Fe uptake (Römheld and Marschner 1986). Fe2+ uptake also occurs in graminaceous species such as rice (Ishimaru et al. 2006). Various types of chelators and transporters also play essential roles in Fe translocation within the plant body (Kobayashi and Nishizawa 2012; Kobayashi et al. 2019).
In response to low iron availability, plants induce the expression of a set of genes involved in Fe uptake and translocation. This response is mediated at the transcript level by cascades comprising transcription factors (TFs), and modulated by ubiquitin ligases (Kobayashi and Nishizawa 2012; Gao et al. 2019; Kobayashi 2019). This regulation has mainly been investigated in rice and Arabidopsis thaliana. Irrespective of clear divergences in Fe uptake systems between rice and Arabidopsis, many regulatory components of TFs and ubiquitin ligases are conserved regarding similarities in function and sequence (Kobayashi 2019). The conserved TFs belong to the Ib, IVb, and IVc subgroups of the basic helix-loop-helix (bHLH) family (Gao et al. 2019; Kobayashi 2019). The bHLH TFs are widely distributed among eukaryotic organisms, among which higher plants possess especially large numbers; for example, rice and Arabidopsis have 173 and 158 bHLH proteins, respectively (Pires and Dolan 2010). Plant bHLH proteins can be phylogenetically divided into 26 subfamilies (Pires and Dolan 2010). The formation of homodimers or heterodimers is a prerequisite for DNA binding and thus transcriptional regulation of bHLH TFs.
Subgroup Ib bHLH TFs, including rice OsIRO2 and Arabidopsis AtbHLH38, AtbHLH39, AtbHLH100, and AtbHLH101, induce the expression of large subsets of genes involved in Fe uptake and translocation (Ogo et al. 2007, 2011; Wang et al. 2007, 2013; Yuan et al. 2008; Sivitz et al. 2012). In contrast, subgroup IVb bHLH TFs, including rice OsIRO3 and Arabidopsis PYE and AtbHLH11, repress the expression of subsets of genes involved in Fe uptake and translocation (Long et al. 2010; Zheng et al. 2010; Tanabe et al. 2019). The expression of these subgroup Ib and IVb bHLH genes, except for AtbHLH11, is strongly induced under Fe deficiency at the transcript level (Ogo et al. 2006; Long et al. 2010; Zheng et al. 2010), indicating the presence of upstream TFs. Rice IDEF1, which belongs to ABI3/VP1 but is not a bHLH family TF, induces the expression of OsIRO2 and OsIRO3 (Kobayashi et al. 2007, 2009, 2014); however, Arabidopsis does not possess an apparent IDEF1 ortholog. Arabidopsis Ib bHLH TFs form heterodimers with FIT, a subgroup IIIa bHLH TF, to induce Fe uptake-related genes (Yuan et al. 2008; Wang et al. 2013), whereas rice lacks an apparent FIT ortholog, and OsIRO2 does not appear to require heterodimer counterparts for its function.
Recent studies have clarified that some subgroup IVc bHLH TFs, including rice OsbHLH060 (also designated OsPRI1) and Arabidopsis AtbHLH34, AtbHLH104, AtbHLH105 (also designated ILR3), and AtbHLH115, act upstream of subgroup Ib and IVb bHLH TFs by inducing their transcriptional expression (Selote et al. 2015; Zhang et al. 2015; Li et al. 2016; Liang et al. 2017; Zhang et al. 2017). The transcript levels of these subgroup IVc bHLHs are affected little by Fe nutritional status (Zhang et al. 2015; Liang et al. 2017; Zhang et al. 2017). Arabidopsis subgroup IVc bHLH TFs interact with each other and with PYE (Long et al. 2010; Selote et al. 2015; Zhang et al. 2015; Li et al. 2016; Liang et al. 2017), suggesting transcriptional regulation diversity through variation among heterodimers.
The expression of these Fe deficiency-inducible genes involved in Fe uptake and translocation is negatively regulated by HRZ/BTS ubiquitin ligases at the transcript level (Kobayashi et al. 2013; Selote et al. 2015; Hindt et al. 2017). HRZ/BTS proteins share conserved domain structures including hemerythrin, CHY, CTCHY and RING zinc (Zn)-fingers, and rubredoxin-type fold (Urzica et al. 2012; Kobayashi et al. 2013). The hemerythrin domain binds Fe and Zn, putatively participating in intracellular Fe sensing, analogous to mammal Fe sensor protein FBXL5 (Hindt and Guerinot 2012; Kobayashi et al. 2013; Selote et al. 2015; Rodríguez-Celma et al. 2019a). The RING Zn-finger usually binds Zn and serves as a component of E3 ubiquitin ligase, which ubiquitinates specific proteins for 26S proteasome-mediated degradation or modification (Hua and Vierstra 2011). This domain in Arabidopsis BTS is crucial for both its E3 ligase activity in vitro and its function in vivo (Selote et al. 2015; Matthiadis and Long 2016; Hindt et al. 2017). CHY and CTCHY Zn-fingers are also predicted to bind Zn and putatively mediate transcriptional, post-transcriptional, or post-translational gene regulation (Gamsjaeger et al. 2007; Sheng et al. 2008). The rubredoxin-type fold (also designated the Zn ribbon) is predicted to bind Fe or Zn, forming a Fe–sulfur cluster when bound to Fe (Sieker et al. 1986; Sheng et al. 2008).
Knockdown of one or both of the rice HRZ genes, OsHRZ1 and OsHRZ2, results in markedly enhanced transcript expression of most of the Fe deficiency-inducible genes involved in Fe uptake and translocation, tolerance to low Fe availability, hypersensitivity to extreme Fe excess conditions, and Fe accumulation in shoots and seeds (Kobayashi et al. 2013; Aung et al. 2018). Similar phenotypes are also observed in knockout mutants of OsHRZ1 (Zhang et al. 2017) and knockdown and point mutants of Arabidopsis BTS (Long et al. 2010; Selote et al. 2015; Hindt et al. 2017), a functional ortholog of rice OsHRZ1 and OsHRZ2. Transcript levels of OsHRZ1, OsHRZ2, and BTS are relatively high in shoots and root steles, and are induced under Fe deficiency (Long et al. 2010; Kobayashi et al. 2013). In addition, BTS protein accumulation is increased under lower Fe abundance in in vitro translation reactions (Selote et al. 2015). However, physiological analyses suggested that HRZ/BTS regulatory activity increase with Fe abundance (Kobayashi et al. 2013; Hindt et al. 2017; Aung et al. 2018). This Fe-dependent activity of HRZ/BTS may be important for inhibiting excessive Fe uptake and translocation, thus avoiding Fe toxicity. Arabidopsis possesses two more HRZ/BTS homologs, BTSL1 and BTSL2, which have been suggested to play similar but distinct roles, mainly functioning in the root epidermis and cortex (Hindt et al. 2017; Rodríguez-Celma et al. 2019b).
Recent studies have identified some members of subgroup IVc bHLHs as putative ubiquitination targets of HRZ/BTS through the investigation of protein binding, in vivo protein expression, ubiquitination, or degradation assays (Selote et al. 2015, 2018; Zhang et al. 2017). Rice OsbHLH060 interacts with OsHRZ1; the latter is thought to ubiquitinate the former for degradation (Zhang et al. 2017). Genetic studies have indicated that the Fe deficiency tolerance and enhanced expression of Fe-related genes observed in an OsHRZ1 knockout line is partly dependent on the presence of OsbHLH060 (Zhang et al. 2017). Similarly, Arabidopsis AtbHLH105 and AtbHLH115 interact with BTS, and the abundance of these IVc bHLH proteins is negatively regulated by BTS (Selote et al. 2015), suggesting ubiquitination-dependent degradation. AtbHLH104 also interacts with BTS; however, the protein level of AtbHLH104 is apparently not regulated by BTS (Selote et al. 2015). Two NAC-family TFs, VOZ1 and VOZ2, may also be degraded via BTS-dependent ubiquitination (Selote et al. 2018). Although VOZ1 and VOZ2 have no apparent function related to Fe deficiency tolerance, decreased abundance of the VOZ2 protein through BTS overexpression results in tolerance to drought stress in Arabidopsis (Selote et al. 2018). A similar approach showed that FIT, a master regulator in Arabidopsis Fe deficiency response (Colangelo and Guerinot 2004; Jakoby et al. 2004), is an ubiquitination target of BTSL1 and BTSL2 (Rodríguez-Celma et al. 2019b).
These findings regarding bHLH TFs and HRZ/BTS ubiquitin ligases suggest the presence of a general regulatory pathway of HRZ/BTS → IVc bHLHs → Ib/IVb bHLHs → Fe deficiency-inducible genes (Online Resource Fig. S1a; Gao et al. 2019; Kobayashi 2019). However, it remains an open question whether this pathway fully explains the wide-ranging inhibitory functions of HRZ/BTS, due to a lack of comprehensive studies on ubiquitination substrates of HRZ/BTS and regulatory targets of all IVc bHLH TFs. Although OsbHLH060 and all four IVc bHLH TFs in Arabidopsis induce both Ib and IVb bHLH genes (Zhang et al. 2015; Li et al. 2016; Liang et al. 2017; Zhang et al. 2017), it is unknown how the balance between gene induction by Ib bHLHs and repression by IVb bHLHs could coordinate to produce proper Fe deficiency responses. In addition, IVc bHLHs may directly regulate some Fe deficiency-inducible genes without mediating Ib and IVb bHLHs; however, this possibility has not yet been explored. Moreover, rice possesses three other subgroup IVc bHLH genes in addition to OsbHLH060, namely OsbHLH057, OsbHLH058, and OsbHLH059 (Zheng et al. 2010), but their functions have yet to be investigated. To date, there has been no report of possible ubiquitination targets of OsHRZ2, except for self-ubiquitination activity in OsHRZ2 as well as OsHRZ1 (Kobayashi et al. 2013). Therefore, the objective of the present study was to characterize OsbHLH058 and OsbHLH059 functions related to Fe deficiency responses involving HRZ ubiquitin ligases. We show that OsbHLH058 and OsbHLH060, but not OsbHLH059, interact with OsHRZ1 and OsHRZ2. Our results further indicate that OsbHLH058 and OsbHLH059 positively regulate major iron deficiency responses in rice (Fig. S1b).
Materials and methods
Plant materials and growth conditions
We used wild-type rice (O. sativa L.) cultivar Nipponbare, cultured for a previous study (Kobayashi et al. 2013) for OsbHLH gene expression analysis. Transgenic lines possessing OsbHLH genes were obtained as described below. T-DNA insertion lines b059-1 (1B-08437) and b059OX-1 (3A-09300) (background cultivar: Dongjin) were obtained from the Rice T-DNA Insertion Sequence Database (POSTECH; Pohang University of Science and Technology, Pohang, Korea; Jeong et al. 2002).
Seeds were germinated on Murashige and Skoog (MS) medium (Murashige and Skoog 1962) with or without hygromycin B (50 mg L−1) for transformants and non-transformants (NT), respectively. After 18–22 d of culture at 28 °C under a 14-h light/10-h dark cycle followed by 3 d of acclimation, plantlets were transferred to a hydroponic solution in a greenhouse under a 30 °C light/25 °C dark cycle with natural light conditions. The composition of the hydroponic solution was as follows: 0.70 mM K2SO4, 0.10 mM KCl, 0.10 mM KH2PO4, 2.0 mM Ca(NO3)2, 0.50 mM MgSO4, 10 μM H3BO3, 0.50 μM MnSO4, 0.50 μM ZnSO4, 0.20 μM CuSO4, 0.01 μM (NH4)6Mo7O24, and 100 μM Fe(III)–EDTA. After 5–6 days of preculture in hydroponic solution, Fe deficiency was initiated by omitting Fe(III)-EDTA from the solution. The nutrient solution was renewed after 4 days of Fe deficiency and sufficiency (control) treatments. After 7–9 days, whole roots and leaf blades were harvested from three plants for each replicate, and immediately frozen in liquid nitrogen for expression analysis, or cut into pieces and immediately dried for metal concentration analysis.
Pot cultivation was conducted using normal soil by transplanting each seedling into a 1-L pot filled with a 1:1 mixture of Bonsol (Sumitomo Chemical, Japan) and vermiculite (Protoleaf, Japan), supplied with 0.5 g each of EcoLongTotal 70 and LongTotal 140 controlled-release fertilizers (JCAM AGRI, Japan) until seed maturation.
Quantitative reverse-transcription polymerase chain reaction (qRT-PCR) analysis
RNA extraction, DNase treatment, reverse-transcription and qRT-PCR were performed as described in Kobayashi et al. (2016), using primers listed in Online Resource Table S1. Transcript abundance was expressed as a ratio relative to the levels in Fe-sufficient NT roots in each experiment.
Yeast two-hybrid assay
To construct OsHRZ1 and OsHRZ2 bait vectors, the RING Zn-finger domain was deleted by inverse PCR using primers listed in Table S1 and pENTR/D-TOPO-OsHRZ vectors (Kobayashi et al. 2013) as templates. For the HRZ1ΔHrRi, HRZ1ΔHrRiRub and HRZ1ΔHrRiZnF baits, OsHRZ12-delRING-F and OsHRZ1-delRING-R were used as the primers and the pENTR/D-TOPO-OsHRZ1 ΔH vector was used as a template. For the HRZ2ΔRi bait, OsHRZ12-delRING-F and OsHRZ2-delRING-R were used as the primers and the pENTR/D-TOPO-OsHRZ2 FL vector was used as a template. Amplified fragments were purified and self-ligated, and the sequences were verified. The resultant plasmids with expected RING Zn-finger deletions of amino acids 1112–1180 for OsHRZ1 and 687–755 for OsHRZ2 in frame were used for the HRZ1ΔHrRi and HRZ2ΔRi baits, respectively. Using the same method, plasmids with spontaneous OsHRZ1 deletions of amino acids 1109–1212 and 888–1180 in frame were also obtained; these were used for HRZ1ΔHrRiRub and HRZ1ΔHrRiZnF baits, respectively. The HRZ inserts of these plasmids were excised using EcoRI and SalI, and inserted into pGBKT7 (TaKaRa, Japan) at the same sites. To construct HRZ1Hr baits, the plasmid pENTR/D-TOPO-OsHRZ1 ΔRZ (Kobayashi et al. 2013) was digested with NcoI, and the excised partial insert corresponding to OsHRZ1 amino acids 7–456 was subcloned into pGBKT7 at the same site.
Full-length open reading frames of OsbHLH057, OsbHLH058, OsbHLH059, and OsbHLH060 were amplified by PCR using primers listed in Table S1 and a complementary DNA (cDNA) pool of Nipponbare rice cultivar roots and leaves (Kobayashi et al. 2013), and cloned into pENTR/D-TOPO (Invitrogen, USA); the sequences were then verified. Fragments were excised at the primer sites using restriction enzymes EcoRI and SalI for OsbHLH057, or EcoRI and BamHI for OsbHLH058, OsbHLH059, and OsbHLH060, and inserted into pGADT7AD (TaKaRa) at the same sites.
A yeast two-hybrid assay was performed using the Matchmaker Gold Yeast Two-Hybrid System (TaKaRa) following the manufacturer’s protocol (PT4084-1). Briefly, bait and prey vectors were separately transformed into yeast (Saccharomyces cerevisiae) strains Y2HGold and Y187, respectively, and grown on Synthetic Defined (SD) medium without tryptophan and leucine (SD-W and SD-L), respectively. Control bait plasmid pGBKT7-53, which expresses the GAL4 DNA-binding domain (BD) fused with murine p53, and prey plasmid pGADT7-T, which expresses the GAL4 activation domain (AD) fused with SV40 large T-antigen, supplied by the manufacturer, were also transformed. Bait–prey combinations were then produced by mating on SD-LW medium, and growth was tested by spotting 10 μL of each culture adjusted to OD600 = 1.0 onto SD selective media without leucine, tryptophan, histidine and adenine (–L–W–H–A), that without leucine, tryptophan and histidine (–L–W–H), and that without leucine and tryptophan supplemented with 5-bromo-4-chloro-3-indolyl-α-D-galactopyranoside (–L–W + X-α-Gal), followed by culture for 5 days at 30 °C. For further experiments (Fig. S3), bait–prey mated colonies were applied on SD selective medium –L–W–H–A + Aureobasidin A (AbA) or control medium SD–L–W, and grown for 5 days at 30 °C.
In vitro ubiquitination assay
To produce recombinant proteins, OsbHLH gene fragments were excised from the abovementioned pENTR/D-TOPO vectors using EcoRI and PstI for OsbHLH058 and OsbHLH060, or EcoRI and SalI for OsbHLH059, and inserted into pET42a(+) (Novagen, USA) at the same sites, constructing the OsbHLH in-frame downstream of glutathione-S-transferase (GST)-tag-His-tag-S-tag. These pET42a(+)-OsbHLH plasmids, as well as pET42a(+), which expresses GST-tag-His-tag-S-tag protein, as a negative control, were introduced into Escherichia coli strain BL21(DE3)pLysS. Recombinant proteins were obtained and purified using the GST fusion system (Novagen) according to the manufacturer’s instructions, except that E. coli was cultured at 16 °C. Purified proteins were concentrated using a VivaSpin 2-30 K column (GE Healthcare, USA).
Maltose binding protein (MBP)-fused OsHRZ proteins and MBP-LacZ control were prepared, and a ubiquitination assay conducted, as described in Kobayashi et al. (2013) using the Ubiquitinylation Kit (ENZO Life Sciences, USA), except that the potential GST-fused protein substrate (300 nM each) was added to the reaction. GST- and MBP-fused proteins were detected using an anti-GST primary antibody (MBL, Japan) and an anti-MBP primary antibody (Sigma, USA), respectively, at 1:1500 dilution in Can Get Signal solution 1 (Toyobo), followed by horseradish peroxidase-conjugated anti-mouse secondary antibody (GE Healthcare) at 1:10000 dilution in Can Get Signal solution 2 (Toyobo).
Production of transgenic rice
To produce rice overexpression lines, the LR Clonase reaction (Invitrogen) was conducted using pH7FWG2 (Karimi et al. 2002) as the destination vector and abovementioned OsbHLH058-pENTR/D-TOPO as the entry vector, constructing in-frame fusion of OsbHLH058 downstream of the cauliflower mosaic virus 35S promoter. To produce knockdown rice lines by RNA interference, a 295-bp fragment corresponding to the 3′-untranslated region (3′UTR) and flanking coding region of OsbHLH058, or a 292-bp fragment corresponding to the 3′UTR of OsbHLH059, was amplified by PCR using primers listed in Table S1 and a cDNA pool of Tsukinohikari rice cultivar roots and leaves (Kobayashi et al. 2013) and inserted into pENTR/D-TOPO; the sequence was then verified. Using the LR Clonase reaction, the fragment was transferred into the destination vector, pIG121-RNAi-DEST (Ogo et al. 2007), to construct the RNA interference vector. Transformation of rice cultivar Tsukinohikari was performed as previously described (Hiei et al. 1994; Kobayashi et al. 2001), and T1 seeds were used for further analysis.
Metal concentration analysis
Leaf segments and T1 brown seeds were dried for 2–3 days at 70 °C, and portions weighing 80–200 mg were wet-ashed with 1.5 mL 13.4 M HNO3 and 1.5 mL 8.8 M H2O2 for 20 min at 220 °C using a MarsXpress oven. Fe, Zn, Mn, and Cu concentrations were measured by inductively coupled plasma optical emission spectrometry (ICPS-8100; Shimadzu, Japan).
Results
Each of the four subgroup IVc bHLH genes show a distinct expression pattern
Amino acid sequences of the four rice subgroup IVc bHLH proteins, OsbHLH057, OsbHLH058, OsbHLH059, and OsbHLH060, are closely related (Fig. S2; Zheng et al. 2010). To estimate possible functions of each of these genes related to Fe deficiency responses in rice, we first examined the transcript levels of these genes in wild-type rice plants hydroponically cultured under Fe sufficiency and deficiency for 7 days by qRT-PCR (Fig. 1). In both roots and leaves, OsbHLH057 and OsbHLH060 expression was moderately induced under Fe deficiency (Fig. 1a, b). In contrast, OsbHLH058 expression was moderately repressed in Fe-deficient roots and strongly repressed in Fe-deficient leaves (Fig. 1a, b). OsbHLH059 expression was slightly repressed in Fe-deficient roots and leaves, although there were no significant differences in expression levels between Fe sufficiency and deficiency (Fig. 1a, b). OsbHLH057, OsbHLH059, and OsbHLH060 expression levels were higher in leaves than in roots; however, that of OsbHLH058 was similar between roots and leaves (Fig. 1a, b).
Fig. 1.

Expression levels of subgroup IVc bHLH genes. The transcript levels of OsbHLH057, OsbHLH058, OsbHLH059, and OsbHLH060 were quantified by reverse-transcription polymerase chain reaction (RT-PCR) in roots (a) and leaves (b) of wild-type rice cultured under Fe sufficiency (+Fe) or deficiency (−Fe) for 7 days. Transcript abundance was normalized against the rice α-2 tubulin transcript level and expressed as a ratio relative to levels in +Fe roots. Means ± standard deviation (SD; n = 3) are shown. Asterisks indicate significant differences from the +Fe value (two-sample Student’s t test; *P < 0.05; **P < 0.01)
We also examined the expression pattern of these four genes using our previous 44 K microarray results (Ogo et al. 2008, 2014; Kobayashi et al. 2009, 2013; Table S2) and a publically available database. Fe deficiency responses in whole roots and shoots determined by microarray (Ogo et al. 2008) were similar to our qRT-PCR results (Fig. 1; Table S2). Tissue-specific microarray analysis after microdissection (Ogo et al. 2014) indicated that OsbHLH058 expression was repressed in the root vascular bundle, cortex, and epidermis/exodermis under Fe deficiency, whereas OsbHLH059 expression was repressed in the root cortex and epidermis/exodermis under Fe deficiency (Table S2). In contrast, OsbHLH057 expression was induced in the root epidermis/exodermis under Fe deficiency (Table S2). Microarray results using HRZ, IDEF1, and IDEF2 transformants (Kobayashi et al. 2009, 2013; Ogo et al. 2008) revealed that OsbHLH057 expression was induced in both OsHRZ and IDEF2 knockdown lines, but repressed in both IDEF1 induction and IDEF1 knockdown lines, whereas OsbHLH058, OsbHLH059, and OsbHLH060 expression was little affected in these transformants (Table S2).
Inspection of the publicly available microarray database RiceXPro (Sato et al. 2011, 2013; http://ricexpro.dna.affrc.go.jp/index.html) revealed that all four subgroup IVc bHLH genes are widely expressed in various organs throughout the plant lifespan; however, some differences in expression patterns were observed. Notably, leaf expression of OsbHLH058 and OsbHLH060 showed diurnal rhythm with a steep peak at early daytime and late night-time, respectively, whereas diurnal alteration of OsbHLH057 and OsbHLH059 expression was less obvious. OsbHLH058, OsbHLH059, and OsbHLH060 expression was induced in roots by jasmonic acid treatment, but OsbHLH057 expression was not. Collectively, these results indicate that each of the subgroup IVc bHLH genes shows a distinct expression pattern at the transcript level. We mainly focused on OsbHLH058 and OsbHLH059 for further analysis, because expression pattern of these two genes in response to Fe deficiency was clearly distinct from that of previously characterized OsbHLH060 (Fig. 1; Table S2).
OsbHLH058 shows strong interaction with OsHRZs in yeast, but may not be ubiquitinated by OsHRZs
To explore possible relationships between subgroup IVc bHLH TFs and OsHRZs, we performed yeast two-hybrid analysis to examine interactions between these proteins. Because the RING Zn-finger domain of both OsHRZ1 and OsHRZ2 showed strong auto-activation activity when used as bait (data not shown), we used OsHRZ1 or OsHRZ2 lacking the RING Zn-finger domain, due to deletion, as a bait (Fig. 2a). For OsHRZ1, we used four partial sequences: an N-terminal fragment possessing the first two hemerythrin domains but no other domains (HRZ1Hr), a C-terminal fragment lacking the three hemerythrin domains and the RING Zn-finger domain (HRZ1ΔHrRi), a C-terminal fragment further lacking a major region of the rubredoxin-type fold and a minor part of the CTCHY Zn-finger domain due to deletion (HRZ1ΔHrRiRub), and a C-terminal fragment possessing the rubredoxin-type fold but lacking all three Zn-finger domains due to deletion (HRZ1ΔHrRiZnF). For OsHRZ2, we used a full-length sequence lacking only the RING Zn-finger domain due to deletion (HRZ2ΔRi). We also used p53 as a control bait. We used subgroup IVc bHLH TF or large T-antigen (T-antigen) as prey. As expected, T-antigen and p53 interacted with each other, as shown by superior growth in high-stringency and low-stringency selective media, –L-W–H–A and –L–W–H, respectively, as well as blue staining in –L–W + X-α-Gal medium (Fig. 2b). HRZ partial fragments did not show auto-activation activity when introduced with T-antigen control, except for mild auto-activation by HRZ1ΔHrRiZnF in –L–W–H and –L–W + X-α-Gal media (Fig. 2b).
Fig. 2.

Yeast two-hybrid analysis of OsHRZ and subgroup IVc bHLH proteins. a Domain structures of OsHRZ partial sequences used for GAL4 DNA-binding domain (BD)-fusion. b Growth and color of yeasts transformed with BD-fused bait and GAL4 activation domain (AD)-fused prey after 5 days of culture. Interaction was detected by enhanced growth in high-stringency (–L–W–H–A) and low-stringency (–L–W–H) selective medium, or by blue staining in –L–W + X-α-Gal medium. Basal growth was confirmed in –L–W + X-α-Gal medium. BD-p53 and AD-large T antigen (T-antigen) are controls provided by the manufacturer
In the high-stringency selective medium –L–W–H–A, OsbHLH058 clearly enhanced growth when introduced with HRZ1ΔHrRi or HRZ2ΔRi, but not with other baits analyzed (Fig. 2b). This result suggests that OsbHLH058 interacts with OsHRZ1 and OsHRZ2, in which the rubredoxin-type fold and the CHY or CTCHY Zn-finger or their N-terminal flanking region have essential roles, but hemerythrin and the RING Zn-finger domains are not required for interaction. In contrast, neither OsbHLH059 nor OsbHLH060 showed any interaction in –L–W–H–A medium (Fig. 2b). The same interaction pattern was also confirmed by blue staining in –L–W + X-α-Gal medium (Fig. 2b).
In the low-stringency selection medium –L–W–H, OsbHLH058 weakly enhanced growth with HRZ1ΔHrRiRub bait, in addition to strong growth activation with HRZ1ΔHrRi or HRZ2ΔRi (Fig. 2b). In this medium, mild growth activation was also observed by OsbHLH060 with HRZ1ΔHrRi or HRZ2ΔRi (Fig. 2b). This result suggests that OsbHLH060 interacts with OsHRZs in a manner similar to OsbHLH058 but with lower affinity. No interaction was observed for OsbHLH059, even in –L–W–H medium (Fig. 2b). We also performed a separate yeast two-hybrid analysis using OsbHLH057 or OsbHLH058 as prey. OsbHLH057 also showed no strong interaction with OsHRZs (Fig. S3). These results suggest that OsbHLH058 interacts with OsHRZ1 and OsHRZ2 more strongly than other subgroup IVc bHLH TFs.
We then tried to detect ubiquitination of subgroup IVc bHLH TFs by OsHRZs. We produced recombinant proteins of GST-fused OsbHLH058, OsbHLH059, and OsbHLH060, as well as GST-His-S as a negative control, and used these as potential substrates for our in vitro ubiquitination assay system (Kobayashi et al. 2013). We used MBP-fused full-length OsHRZ1 and OsHRZ2 as active E3 ubiquitin ligases, as well as MBP-LacZ and MBP-OsHRZ1ΔRZ, the latter of which lacks the RING and other Zn-finger domains and thus lacks E3 ubiquitin ligase activity (Kobayashi et al. 2013), as E3 negative controls. When we added either OsbHLH058 or OsbHLH060 to the reaction solution, high-molecular-weight smear bands appeared in all lanes, irrespective of presence or absence of E3 ligase activity, as determined by substrate detection using anti-GST antibody (Fig. 3). We did not observe more intense bands or smear staining of OsbHLH058 or OsbHLH060 with active OsHRZ1 or OsHRZ2. A similar result was obtained with OsbHLH059 with or without active OsHRZ1. E3 ligase activity in OsHRZ1 and OsHRZ2 was confirmed by appearance of much stronger high-molecular-weight smear bands than that in negative controls using anti-MBP antibody (Fig. S4; Kobayashi et al. 2013). These results indicate that OsbHLH058, OsbHLH059, and OsbHLH060 were not ubiquitinated by OsHRZ1 or OsHRZ2 in the experimental conditions of this study.
Fig. 3.
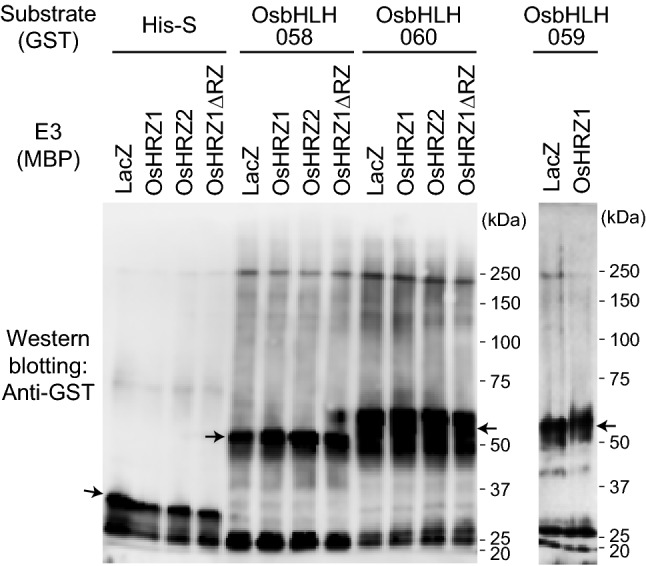
In vitro ubiquitination assay of OsbHLH058, OsbHLH060, and OsbHLH059 with OsHRZ ubiquitin ligases. Reaction was conducted with maltose binding protein (MBP) fusions of full-length OsHRZs as E3 ligases, and MBP-LacZ and MBP-OsHRZ1ΔRZ as negative controls, and GST fusions of OsbHLH058, OsbHLH060, and OsbHLH059 as potential substrates, or GST-tag-His-tag-S-tag (His-S) as a negative control. GST-fused proteins were detected by Western blotting using anti-GST antibody. The positions and sizes of the molecular mass markers are shown to the right of each blot. Monomeric free GST-fused proteins are indicated with arrows
OsbHLH058 overexpression rice shows tolerance to Fe deficiency and enhanced expression of Fe deficiency-inducible genes
Although we did not detect ubiquitination of OsbHLH058 by OsHRZs, their protein interaction and Fe deficiency-repressed OsbHLH058 expression suggest the possible involvement of OsbHLH058 in Fe responses and homeostasis. To address this possibility, we produced transgenic rice lines constitutively overexpressing OsbHLH058 under the control of the cauliflower mosaic virus 35S promoter (designated b058OX lines). We selected representative lines, b058OX-2 and -4, showing enhanced OsbHLH058 expression (Fig. S5a), for further analysis.
These lines and NT rice were cultured hydroponically and subjected to Fe deficiency treatment. The chlorophyll level in the newest leaf as determined by SPAD value, which is indicative of Fe nutrition, was higher in b058OX lines 2 and 4 than in NT especially during days 4–5 of Fe deficiency treatment (Fig. 4a). The newest leaves of b058OX line 4 still remained greener than those of NT after 7 days of Fe deficiency treatment (Fig. 4b, c). These results suggest that the b058OX lines are tolerant to low Fe availability. These lines and NT showed similar leaf colors under Fe sufficiency (Fig. 4d). The b058OX lines showed retarded growth from the beginning of seedling stages, which led to shorter shoot length until the end of hydroponic culture under either Fe condition (Fig. 4b, d). However, the growth rates were similar between the genotypes during the Fe treatments (data not shown).
Fig. 4.

Fe deficiency tolerance of OsbHLH058 overexpression plants. a Fe deficiency tolerance in hydroponic culture. Relative chlorophyll contents (SPAD values) of the newest leaves were measured after the onset of Fe deficiency treatment. b Representative plants on day 7 of Fe-deficient hydroponic culture. Three plants were bundled for culture of each line. c Representative newest leaves on day 7 of Fe-deficient hydroponic culture. White arrowheads in b and c indicate the same positions. d Representative plants on day 7 of Fe-sufficient hydroponic culture. Three plants were bundled for culture of each line. e, f Fe concentration in leaf blades after Fe-deficient (e) or -sufficient (f) hydroponic culture for 7 days. g Fe concentration in brown seeds after pot culture in Fe-sufficient soil. NT, non-transformant; b058OX-2 and 4, OsbHLH058 overexpression lines 2 and 4, respectively. Means ± SD are shown (a n = 12 for NT, n = 5 for b058OX-2, n = 4 for b058OX-4; e n = 4 for NT, n = 2 for b058OX-2 and 4; f n = 3 for NT, n = 2 for b058OX-4; n = 1 for b058OX-2; g n = 7 for NT, n = 3 for b058OX-2 and 4). Asterisks indicate significant differences compared to NT at each time point and condition (two-sample Student’s t-test; *P < 0.05; **P < 0.01)
We then measured Fe concentrations in the leaves of these plants. Both b058OX lines contained leaf Fe concentrations similar to NT levels after Fe-deficient hydroponic culture (Fig. 4e), but tended to contain higher leaf Fe concentration than NT after Fe-sufficient hydroponic culture (Fig. 4f). Both b058OX lines contained significantly higher Fe concentrations in brown seeds than NT after normal soil culture (Fig. 4g).
We then investigated the expression patterns of Fe-related genes in roots of these b058OX lines after hydroponic culture. We quantified the transcript levels of six representative Fe deficiency-inducible genes: OsNAS1, TOM1, and OsYSL15, which are responsible for deoxymugineic acid (DMA)-based Fe(III) uptake and translocation, OsIRT1 for Fe2+ uptake, OsYSL2 for Fe(II)-nicotianamine transport for internal Fe translocation, and OsIRO2 for positive regulation of genes involved in Fe(III) uptake and translocation (Kobayashi et al. 2014). Under Fe-sufficient conditions, the expression of all genes was markedly induced in the b058OX-2 and 4 lines compared with NT (Fig. 5a), suggesting positive regulation of these genes by OsbHLH058. Under Fe deficiency, however, these genes were not more highly expressed in b058OX-2 and 4 than in NT, except for still greater induction of OsYSL2 (Fig. 5b). OsNAS1, TOM1, and OsIRO2 expression was moderately lower in the b058OX-2 and 4 lines than in NT under Fe deficiency (Fig. 5b).
Fig. 5.

Expression levels of representative Fe-related genes in OsbHLH058 overexpression plants. Transcript levels in roots of OsbHLH058 overexpression (b058OX) lines 2 and 4 and non-transformants (NT) were quantified by RT-PCR, after 7 days of hydroponic culture under Fe sufficiency (a) or deficiency (b). Transcript abundance was normalized against the rice α-2 tubulin transcript level and expressed as a ratio relative to the levels in the +Fe NT roots. Means ± SD (n = 3) are shown. Asterisks indicate significant differences from the NT value at each condition (two-sample Student’s t-test; *P < 0.05; **P < 0.01)
Altered expression level of OsbHLH059 influences rice Fe deficiency responses
To explore further the possible role of OsbHLH058 and OsbHLH059 in Fe nutrition, we then searched for T-DNA insertion rice lines from the POSTECH database (Jeong et al. 2002). Although we could not find putative T-DNA insertion lines for OsbHLH058, we found two T-DNA insertion rice lines for OsbHLH059. The insertion line 1B-08437, which we designated b059-1, possessed a T-DNA insertion in the 5′-untranslated region, and showed decreased OsbHLH059 expression (Fig. S5b, c). The insertion line 3A-09300, which we designated b059OX-1, showed slightly enhanced OsbHLH059 expression, presumably due to the insertion of enhancer sequences in the OsbHLH059 promoter region adjacent to the OsbHLH059 gene (Fig. S5b, c). When cultured hydroponically under Fe deficiency, the b059-1 line showed a lower newest-leaf SPAD value than NT at days 2 and 3, whereas the b059OX-1 line showed a higher SPAD value than NT at days 1, 4, and 5 (Fig. 6a). The SPAD values of these lines settled nearer to the NT value at day 6 and thereafter (Fig. 6a). Notably, brownish or necrotic regions appeared in the newest leaves of the b059-1 line, but not in those of NT or the b059OX-1 line, from day 5 of the Fe deficiency treatment (Fig. 6b, c). This symptom was not observed in any older leaves under Fe deficiency, or any leaves under Fe sufficiency. These results suggest that OsbHLH059 expression levels also affect Fe nutrition in rice, in addition to OsbHLH058.
Fig. 6.

Fe deficiency tolerance of OsbHLH059 T-DNA insertion lines. a Fe deficiency tolerance in hydroponic culture. Relative chlorophyll contents (SPAD values) of the newest leaves were measured after the onset of Fe deficiency treatment. b Representative plants on day 9 of Fe-deficient hydroponic culture. Three plants were bundled for culture of each line. White arrowheads indicate representative positions of brown spots in the newest leaves. c Representative newest leaves on day 9 of Fe-deficient hydroponic culture. NT, non-transformant; b059-1, OsbHLH059 knockdown line by T-DNA insertion; b059OX-1, OsbHLH059 overexpression line by T-DNA insertion. Means ± SD (n = 12) are shown for a. Asterisks indicate significant differences compared to NT (two-sample Student’s t-test; *P < 0.05; **P < 0.01)
Repression of OsbHLH058 or OsbHLH059 results in susceptibility to Fe deficiency and reduced expression of Fe-related genes
To further investigate the roles of OsbHLH058 and OsbHLH059 in Fe deficiency responses, we produced transgenic rice lines with repressed expression of OsbHLH058 (designated b058i lines) or OsbHLH059 (b059i lines) using the RNA interference method. We selected representative lines with decreased expression of OsbHLH058 for b058i or OsbHLH059 for b059i (Fig. S5d). We performed three separate hydroponic experiments to impose Fe deficiency on these lines, as compared with NT (Figs. 7, S6). All selected b058i and b059i lines showed susceptibility to low Fe availability, as indicated by lower newest-leaf SPAD values than NT after the progression of Fe deficiency at around day 5 and thereafter (Figs. 7a, S6a). These lines had yellower leaves than NT at day 7 (Fig. 7b). Moreover, b059i lines, but not b058i lines or NT, exhibited severe browning or necrosis in partial regions of the newest and the second newest leaves at day 6 and thereafter of Fe deficiency treatment, but not in older leaves (Fig. 7b, c, S6b, c), as was observed in the b059-1 line (Fig. 6b, c). The b058i and b059i lines showed similar growth to NT, with no visible phenotypes, under Fe sufficiency (Fig. 7d).
Fig. 7.

Fe deficiency tolerance of OsbHLH058 and OsbHLH059 knockdown plants produced by RNA interference. a Fe deficiency tolerance in hydroponic culture. Relative chlorophyll contents (SPAD values) of the newest leaves were measured after the onset of Fe deficiency treatment. b Representative plants on day 7 of Fe-deficient hydroponic culture. Three plants were bundled for culture of each line. White arrowheads indicate representative positions of brown spots in the newest leaves. c Representative newest leaves on day 7 of Fe-deficient hydroponic culture. d Representative plants on day 7 of Fe-sufficient hydroponic culture. Three plants were bundled for culture of each line. e, f Fe concentration in leaf blades after Fe-deficient (e) or -sufficient (f) hydroponic culture for 7 d. g Fe concentration in brown seeds after pot culture in Fe-sufficient soil. NT, non-transformant; b058i, OsbHLH058 knockdown lines; b059i, OsbHLH059 knockdown lines produced by RNA interference. Means ± SD are shown (a n = 12 for NT and b059i-3, n = 6 for b058i-8, n = 9 for b058i-10; e n = 4 for NT and b059i-3, n = 2 for b058i-8, n = 5 for b058i-10; f n = 3 for NT and b058i-10, n = 4 for b059i-3; g: n = 7 or NT, n = 3 for b058i-1, 8 and 10, n = 4 for b058i-2, n = 2 for b059i-3 and 6). Asterisks indicate significant differences compared to NT at each time point and condition (two-sample Student’s t-test; *P < 0.05; **P < 0.01)
Leaf Fe concentrations among these lines after hydroponic culture were similar to NT levels under either Fe-deficient or -sufficient conditions (Fig. 7e, f). In brown seeds, b058i lines 1 and 10 contained lower concentrations of Fe, whereas b059i lines 3 and 6 showed higher Fe concentrations, compared with NT (Fig. 7g). We also tried to dissect Fe localization in the newest leaves of Fe-deficienct and -sufficient plants by Perls DAB staining (Roschzttardtz et al. 2009), but no clear differences between the genotypes were observed (data not shown).
We then investigated the expression levels of the Fe-related genes OsNAS1, TOM1, OsYSL15, OsIRT1, OsYSL2, and OsIRO2 in roots of the b058i and b059i lines and NT hydroponically grown under Fe sufficiency (Fig. 8). In b058i lines 1 and 2, OsNAS1, TOM1, and OsYSL2 expression was strongly repressed compared with NT. OsYSL15 and OsIRO2 expression also tended to be repressed in these lines, whereas that of OsIRT1 was not repressed but, rather, induced, in b058i line 2. In b059i lines 3 and 6, OsNAS1, TOM1, OsIRT1, and OsYSL2 expression was strongly repressed compared with NT. OsYSL15 expression was repressed only in line 6, whereas OsIRO2 expression levels were similar between the b059i lines and NT.
Fig. 8.

Expression levels of representative Fe-related genes in OsbHLH058 and OsbHLH059 knockdown plant roots. Transcript levels in Fe-sufficient roots of OsbHLH058 knockdown (b058i) lines 1 and 2 and NT after 8 days of hydroponic culture, and in roots of OsbHLH059 knockdown (b059i) lines 3 and 6 and NT lines after 9 days of hydroponic culture, were quantified by RT-PCR. Transcript abundance was normalized against the rice α-2 tubulin transcript level and expressed as a ratio relative to NT root levels in each experiment. Means ± SD (n = 3) are shown. Asterisks indicate significant differences from the NT value at each condition (two-sample Student’s t-test; *P < 0.05; **P < 0.01)
We also examined the expression of these genes in leaves of the identical plants (Fig. S7). TOM1 and OsYSL15 expression was not detected in Fe-sufficient leaves. The b058i lines showed repressed expression of OsYSL2 and OsIRO2, but not OsNAS1 and OsIRT1, compared with NT. The b059i line 6 showed repressed expression of OsYSL2 and tendency of reduced expression of OsNAS1, while the b059i line 3 did not show repressed expression but rather showed increased expression of OsIRT1, OsYSL2 and OsIRO2, compared with NT.
Collectively, these results indicate that both OsbHLH058 and OsbHLH059 positively regulate these Fe-related genes in a similar but not identical manner.
Discussion
OsbHLH058 positively regulates Fe deficiency responses presumably downstream of OsHRZs
In the present study, we investigated the roles of OsbHLH058 and OsbHLH059 in comparison with those of other members of subgroup IVc bHLH TFs in rice: OsbHLH057 and OsbHLH060. We observed clear differences in expression patterns among these four genes (Fig. 1, Table S2). OsbHLH058 expression was markedly repressed under Fe deficiency in roots and leaves. OsbHLH059 expression also tended to be repressed in some tissues, whereas OsbHLH057 expression was significantly induced in roots and leaves. We also detected slight OsbHLH060 induction in roots and leaves, whereas our previous microarray results (Ogo et al. 2008) indicated subtle but statistically insignificant induction of OsbHLH060 in roots and leaves (Table S2). Zhang et al. (2017) reported no induction of OsbHLH060 under Fe deficiency. Notwithstanding slight differences in these results, possibly due to growth conditions or cultivars, these results suggest that OsbHLH060 transcript levels are not much affected by Fe nutritional conditions. Each of the four IVc bHLH genes also showed distinct diurnal and jasmonate-responsive expression patterns based on microarray results, which may underlie functional diversification within this subgroup.
Our yeast two-hybrid assay showed clear interaction of OsbHLH058, but not OsbHLH059, with OsHRZ1 and OsHRZ2 (Fig. 2). Domain deletion analysis suggested that these interactions require the rubredoxin-type fold and the CHY or CTCHY Zn-finger or their N-terminal flanking region (Fig. 2). The functions of these domains have not yet been determined, but structural analysis of the human Pirh2 protein, which shares these domains and the RING Zn-finger domain with HRZs/BTS (Matthiadis and Long 2016), has indicated that these domains in Pirh2 bind Zn to form tertiary structure, which is presumably essential for interaction with other proteins (Sheng et al. 2008). Because HRZs are thought to bind both Fe and Zn via these domains (Kobayashi et al. 2013), intracellular metal status may influence the tertiary structure around these HRZ domains to alter their affinity to interacting partners, including OsbHLH058. Under our assay conditions, OsbHLH060 also showed interaction with OsHRZ1 and OsHRZ2, with similar domain requirements; however, this interaction was observed only in low-stringency medium, suggesting that the interaction with HRZs is weaker in OsbHLH060. Zhang et al. (2017) reported interaction between OsbHLH060 and a C-terminal portion of OsHRZ1 including the RING Zn-finger domain, in a high-stringency medium. Differences in the partial sequences of OsHRZ1 used in these two experiments may explain differences in interaction strength observed between OsbHLH060 and OsHRZ1. We could not include the RING Zn-finger domain in a HRZ bait due to strong auto-activation (data not shown), although Zhang et al. (2017) did not report such auto-activation.
The interaction of OsbHLH058 with HRZ ubiquitin ligases suggests possible ubiquitination of the former by the latter. Indeed, Zhang et al. (2017) reported that OsbHLH060 was ubiquitinated by OsHRZ1, based on in vitro ubiquitination and a degradation assay. Therefore, we used OsbHLH058 and OsbHLH060 as potential targets in our in vitro ubiquitination assay. However, we did not detect ubiquitination of these bHLH proteins by either OsHRZ1 or OsHRZ2 (Fig. 3). We also did not detect ubiquitination of OsbHLH059 by OsHRZ1 (Fig. 3). These bHLH proteins formed high-molecular-weight smear bands, even with inactive E3 controls (Fig. 3); this type of negative control experiment was not reported in Zhang et al. (2017). Similarly, no ubiquitination reaction of AtbHLH105, AtbHLH115, VOZ1, and VOZ2 was reported in Arabidopsis BTS in two recent studies (Selote et al. 2015, 2018). Collectively, no concrete evidence of HRZs/BTS ubiquitination targets has been reported to date, although subgroup IVc bHLH TFs are promising candidates in both rice and Arabidopsis. Very recently, FIT, a subgroup IIIa bHLH TF, was shown to be an ubiquitination target of Arabidopsis BTSL2 by in vitro ubiquitination assay, but this assay also lacked inactive E3 controls (Rodríguez-Celma et al. 2019b).
Although the physiological relevance of interaction between OsbHLH058 and HRZs is unclear, analysis of transgenic lines with overexpressed or repressed expression of OsbHLH058 clearly showed its involvement in Fe deficiency responses (Figs. 4, 5, 7, 8, S5, S6). OsbHLH058 overexpression lines showed enhanced tolerance to Fe deficiency, accumulation of higher Fe concentrations in seeds and Fe-sufficient leaves (Fig. 4), and induced expression of various Fe deficiency-inducible genes for Fe uptake and translocation in Fe-sufficient roots (Fig. 5a). These results suggest that OsbHLH058 positively regulates these genes to facilitate Fe uptake and translocation to shoots and seeds. This hypothesis is further supported by basically the opposite phenotype of the OsbHLH058 knockdown lines, including susceptibility to Fe deficiency, lower Fe concentrations in seeds (Figs. 7, S6), and repressed expression of various Fe deficiency-inducible genes examined in Fe-sufficient roots and leaves (Fig. 8, S7). Fe concentration in Fe-sufficient leaves and OsIRT1 expression in roots did not show a clear and opposite change from that of the overexpression lines, perhaps due to possible complementation of OsbHLH058 knockdown by other bHLH genes, or secondary effects of OsbHLH058 overexpression. Fe deficiency-inducible Fe-related genes in rice can be categorized into four groups according to their functions and expressional patterns in the regulatory network: (i) genes for Fe(III)-DMA uptake and translocation (including OsNAS1, TOM1, and OsYSL15); (ii) genes for Fe2+ uptake (including OsIRT1); (iii) genes for internal Fe translocation (including OsYSL2); (iv) genes regulating group (i)–(iii) genes [including OsIRO2, which positively regulates group (i) genes] (Kobayashi et al. 2014). The expression patterns of OsbHLH058 overexpression and knockdown lines (Figs. 5a, 8, S7) suggest that OsbHLH058 positively regulates all four groups, among which group (i) and (iii) genes may be more strongly regulated by OsbHLH058 in Fe-sufficient roots.
Interestingly, enhanced expression of these Fe-related genes in OsbHLH058 overexpression lines was evident only under Fe sufficiency, but was diminished or even reversed to lower expression levels than in NT under Fe deficiency, except for OsYSL2 (Fig. 5a vs. b). This pattern of expressional change is strikingly similar to that observed in HRZ knockdown lines, including extremely high OsYSL2 induction, even under Fe deficiency (Kobayashi et al. 2013). This similarity, along with the interaction of OsbHLH058 and HRZs, suggests that OsbHLH058 function may be regulated by, or linked to, HRZs. Enhanced expression of Fe-related genes in HRZ knockdown lines is still more prominent under Fe excess conditions (Aung et al. 2018), suggesting that HRZ function is regulated by Fe abundance, which may account for intracellular Fe sensing. OsbHLH058 may be an output of an HRZ-mediated Fe sensing mechanism, in which the function of HRZs may be partially mediated by inhibition of OsbHLH058. Although its molecular mechanism remains completely unknown, binding to HRZs may inhibit homo- or heterodimer formation of OsbHLH058 for its proper function.
The role of OsbHLH059 in positive regulation of Fe deficiency responses is similar to but distinct from that of OsbHLH058
Although OsbHLH059 did not show interaction with HRZs or detectable ubiquitination by OsHRZ1 (Figs. 2, 3), analysis of the transgenic lines showed that OsbHLH059 is also involved in Fe deficiency responses. The OsbHLH059 overexpression line with enhancer-tagged T-DNA showed slight tolerance to Fe deficiency (Fig. 6a), although its phenotype was not strong, possibly due to a very weak increase in OsbHLH058 expression (Fig. S5). Clearer results were obtained from OsbHLH059 knockdown lines (Figs. 6, 7, 8, S5, S6, S7). Similar to OsbHLH058, OsbHLH059 knockdown lines showed susceptibility to Fe deficiency and repressed expression of various Fe-related genes in roots (Figs. 6, 7, 8, S6). In addition, OsbHLH059 knockdown lines by either T-DNA insertion or RNA interference exhibited characteristic brownish or necrotic regions in new leaves after prolonged Fe deficiency treatment (Figs. 6, 7, S5), which were never observed in NT or OsbHLH058 knockdown lines. Because this phenotype was restricted to severely chlorotic regions suffering from Fe deficiency, this result may be due to local leaf damage from either extremely low Fe abundance or ectopic accumulation of other metals. Although we detected no decrease in Fe concentrations in whole leaves of the OsbHLH059 knockdown lines (Fig. 7e), some impairment of Fe distribution in restricted leaf areas may have occurred due to OsbHLH059 knockdown. We did not observe clear differences in Fe localization in the newest leaves by Perls DAB staining (data not shown), perhaps because of very low Fe concentration. A similar occurrence of brown spots in Fe-deficient leaves was reported for OsbHLH060 knockout lines pri1-1 and pri1-2 (Zhang et al. 2017); however, this phenomenon has not been investigated further.
Another distinguishable difference between OsbHLH059 and OsbHLH058 knockdown lines was observed in seed Fe concentration; OsbHLH059 knockdown lines showed higher Fe concentrations than NT, in contrast to OsbHLH058 knockdown lines (Fig. 7g). Expression patterns of Fe-related genes were also different between OsbHLH059 and OsbHLH058 knockdown lines (Figs. 8, S7). These results indicate that OsbHLH059 enhances the pathway of Fe deficiency responses for proper Fe uptake and distribution to rice tissues, in a manner similar to but distinct from that of OsbHLH058. Based on our gene expression results (Figs. 8, S7), OsbHLH059 may positively regulate a large subset of Fe-related genes in Fe-sufficient roots, including those for (i) genes for Fe(III)-DMA uptake and translocation, (ii) genes for Fe2+ uptake, and (iii) genes for internal Fe translocation, but not (iv) OsIRO2. In Fe-sufficient leaves, OsbHLH059 knockdown lines did not show repression of Fe-related genes except for OsYSL2 in b059i line 6 (Fig. S7). Instead, enhanced expression of OsIRT1, OsYSL2 or OsIRO2 was observed in leaves of one or both b059i lines analyzed (Fig. S7), possibly due to secondary effects of disturbed Fe availability, or possible negative regulation by OsbHLH059, especially for OsIRO2 which showed much enhanced expression in both b059i lines (Fig. S7).
In contrast to OsbHLH059, OsbHLH060 appears to positively regulate many Fe-related genes representative of all groups (i)–(iv) under both Fe sufficiency and deficiency, based on the expression patterns of OsbHLH060 knockdown lines in roots (Zhang et al. 2017). Together, these results suggest that three members of the subgroup IVc bHLH TFs, OsbHLH058, OsbHLH059, and OsbHLH060, regulate largely overlapping, but not identical, subsets of Fe deficiency-inducible genes (Fig. S1b).
In OsbHLH058 and OsbHLH059 knockdown roots, the expression levels of many target genes such as OsNAS1, TOM1, and OsYSL2 often decreased to less than 50% of each NT control (Fig. 8), irrespective of high sequence and functional similarity with subgroup IVc bHLH TFs. This result suggests that the expression of these target genes requires both OsbHLH058 and OsbHLH059 in a synergistic, rather than additive, fashion. Although we did not detect interaction between OsbHLH059 and HRZs (Fig. 2), a possible synergistic effect between OsbHLH059 and OsbHLH058 suggests indirect involvement of OsbHLH059 in the HRZ pathway.
Such transactivation function of OsbHLH058 and OsbHLH059 could be mediated by heterodimer formation or synergistic effects of plural cis-acting elements in target genes. Many bHLH TFs preferentially recognize E-box (CANNTG) or G-box (CACGTG) sequences; sequence recognition preferences can be predicted from amino acid sequences of specific residues in the basic region (Toledo-Ortiz et al. 2003). Based on this criterion, all subgroup IVc bHLH TFs in rice and Arabidopsis are predicted to bind preferentially to G-box (Toledo-Ortiz et al. 2003), suggesting significant sharing of cis-acting sequences among the subgroup, with heterodimer formation possibly playing important roles. To discover the precise functions of the apparently diversified bHLH TFs related to Fe homeostasis would require precise and comprehensive characterization of the target genes of each member, including the distribution of corresponding cis-acting elements in each gene promoter context, homo/heterodimer formations of each member, the functional significance of these homo/heterodimer formations, and clarification of the possible involvement of HRZs/BTS in the abundance and activity of each bHLH member. The clarification of these issues would be key to understanding overall regulation cascades in plant Fe deficiency responses.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
We thank Ms. Kazuko Arai and Ms. Mayumi Ura (Ishikawa Prefectural University) for assistance with experiments. This research was supported by the Japan Society for the Promotion of Sciences (JSPS) KAKENHI Grant Numbers 15H01187, 15H05617 and 18H02115 (to T.K.), JSPS KAKENHI Grant Number 16H04891 (to N.K.N.), and by the Advanced Low Carbon Technology Research and Development Program (ALCA) of the Japan Science and Technology Agency (to N.K.N.).
Author contributions
TK designed the research, carried out most of the experiments, and wrote the manuscript with discussion from all the co-authors. AO carried out some experiments of the OsbHLH059 T-DNA lines and production of the OsbHLH059 RNA interference lines. SK carried out some experiments of the OsbHLH059 RNA interference lines. GA and JSJ provided the OsbHLH059 T-DNA lines. NKN supervised the research.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Aung MS, Kobayashi T, Masuda H, Nishizawa NK. Rice HRZ ubiquitin ligases are crucial for response to excess iron. Physiol Plant. 2018;163:282–296. doi: 10.1111/ppl.12698. [DOI] [PubMed] [Google Scholar]
- Briat JF, Dubos C, Gaymard F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015;20:33–40. doi: 10.1016/j.tplants.2014.07.005. [DOI] [PubMed] [Google Scholar]
- Colangelo EP, Guerinot ML. The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell. 2004;16:3400–3412. doi: 10.1105/tpc.104.024315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamsjaeger R, Liew CK, Loughlin FE, Crossley M, Mackay JP. Sticky fingers: zinc-fingers as protein-recognition motifs. Trends Biochem Sci. 2007;32:63–70. doi: 10.1016/j.tibs.2006.12.007. [DOI] [PubMed] [Google Scholar]
- Gao F, Robe K, Gaymard F, Izquierdo E, Dubos C. The transcriptional control of iron homeostasis in plants: a tale of bHLH transcription factors? Front Plant Sci. 2019;10:6. doi: 10.3389/fpls.2019.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiei Y, Ohta S, Komari T, Kumashiro T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994;6:271–282. doi: 10.1046/j.1365-313x.1994.6020271.x. [DOI] [PubMed] [Google Scholar]
- Hindt MN, Guerinot ML. Getting a sense for signals: regulation of the plant iron deficiency response. Biochim Biophys Acta. 2012;1823:1521–1530. doi: 10.1016/j.bbamcr.2012.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hindt MN, Akmakjian GZ, Pivarski KL, Punshon T, Baxter I, Salt DE, Guerinot ML. BRUTUS and its paralogs, BTS LIKE1 and BTS LIKE2, encode important negative regulators of the iron deficiency response in Arabidopsis thaliana. Metallomics. 2017;9:876–890. doi: 10.1039/c7mt00152e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hua Z, Vierstra RD. The Cullin-RING ubiquitin-protein ligases. Annu Rev Plant Biol. 2011;62:299–334. doi: 10.1146/annurev-arplant-042809-112256. [DOI] [PubMed] [Google Scholar]
- Ishimaru Y, Suzuki M, Tsukamoto T, Suzuki K, Nakazono M, Kobayashi T, Wada Y, Watanabe S, Matsuhashi S, Takahashi M, Nakanishi H, Mori S, Nishizawa NK. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+ Plant J. 2006;45:335–346. doi: 10.1111/j.1365-313X.2005.02624.x. [DOI] [PubMed] [Google Scholar]
- Jakoby M, Wang HY, Reidt W, Weisshaar B, Bauer P. FRU (BHLH029) is required for induction of iron mobilization genes in Arabidopsis thaliana. FEBS Lett. 2004;577:528–534. doi: 10.1016/j.febslet.2004.10.062. [DOI] [PubMed] [Google Scholar]
- Jeong DH, An S, Kang HG, Moon S, Han JJ, Park S, Lee HS, An K, An G. T-DNA insertional mutagenesis for activation tagging in rice. Plant Physiol. 2002;130:1636–1644. doi: 10.1104/pp.014357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karimi M, Inze D, Depicker A. Gateway vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002;7:193–195. doi: 10.1016/s1360-1385(02)02251-3. [DOI] [PubMed] [Google Scholar]
- Kobayashi T. Understanding the complexity of iron sensing and signaling cascades in plants. Plant Cell Physiol. 2019;60:1440–1446. doi: 10.1093/pcp/pcz038. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Nishizawa NK. Iron uptake, translocation, and regulation in higher plants. Ann Rev Plant Biol. 2012;63:131–152. doi: 10.1146/annurev-arplant-042811-105522. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Nakanishi H, Takahashi M, Kawasaki S, Nishizawa NK, Mori S. In vivo evidence that Ids3 from Hordeum vulgare encodes a dioxygenase that converts 2′-deoxymugineic acid to mugineic acid in transgenic rice. Planta. 2001;212:864–871. doi: 10.1007/s004250000453. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Ogo Y, Itai RN, Nakanishi H, Takahashi M, Mori S, Nishizawa NK. The transcription factor IDEF1 regulates the response to and tolerance of iron deficiency in plants. Proc Natl Acad Sci USA. 2007;104:19150–19155. doi: 10.1073/pnas.0707010104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi T, Itai RN, Ogo Y, Kakei Y, Nakanishi H, Takahashi M, Nishizawa NK. The rice transcription factor IDEF1 is essential for the early response to iron deficiency, and induces vegetative expression of late embryogenesis abundant genes. Plant J. 2009;60:948–961. doi: 10.1111/j.1365-313X.2009.04015.x. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Nagasaka S, Senoura T, Itai RN, Nakanishi H, Nishizawa NK. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation. Nat Commun. 2013;4:2792. doi: 10.1038/ncomms3792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi T, Itai RN, Nishizawa NK. Iron deficiency responses in rice roots. Rice. 2014;7:27. doi: 10.1186/s12284-014-0027-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi T, Itai RN, Senoura T, Oikawa T, Ishimaru Y, Ueda M, Nakanishi H, Nishizawa NK. Jasmonate signaling is activated in the very early stages of iron deficiency responses in rice roots. Plant Mol Biol. 2016;91:533–547. doi: 10.1007/s11103-016-0486-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi T, Nozoye T, Nishizawa NK. Iron transport and its regulation in plants. Free Radic Biol Med. 2019;133:11–20. doi: 10.1016/j.freeradbiomed.2018.10.439. [DOI] [PubMed] [Google Scholar]
- Li X, Zhang H, Ai Q, Liang G, Yu D. Two bHLH transcription factors, bHLH34 and bHLH104, regulate iron homeostasis in Arabidopsis thaliana. Plant Physiol. 2016;170:2478–2493. doi: 10.1104/pp.15.01827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang G, Zhang H, Li X, Ai Q, Yu D. bHLH transcription factor bHLH115 regulates iron homeostasis in Arabidopsis thaliana. J Exp Bot. 2017;68:1743–1755. doi: 10.1093/jxb/erx043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Long TA, Tsukagoshi H, Busch W, Lahner B, Salt D, Benfey PN. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell. 2010;22:2219–2236. doi: 10.1105/tpc.110.074096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marschner H. Mineral nutrition of higher plants. 2. London: Academic Press; 1995. [Google Scholar]
- Matthiadis A, Long TA. Further insight into BRUTUS domain composition and functionality. Plant Signal Behav. 2016;11:e1204508. doi: 10.1080/15592324.2016.1204508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant. 1962;15:473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x. [DOI] [Google Scholar]
- Ogo Y, Itai RN, Nakanishi H, Inoue H, Kobayashi T, Suzuki M, Takahashi M, Mori S, Nishizawa NK. Isolation and characterization of IRO2, a novel iron-regulated bHLH transcription factor in graminaceous plants. J Exp Bot. 2006;57:2867–2878. doi: 10.1093/jxb/erl054. [DOI] [PubMed] [Google Scholar]
- Ogo Y, Itai RN, Nakanishi H, Kobayashi T, Takahashi M, Mori S, Nishizawa NK. The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe-deficient conditions. Plant J. 2007;51:366–377. doi: 10.1111/j.1365-313X.2007.03149.x. [DOI] [PubMed] [Google Scholar]
- Ogo Y, Kobayashi T, Itai RN, Nakanishi H, Kakei Y, Takahashi M, Toki S, Mori S, Nishizawa NK. A novel NAC transcription factor IDEF2 that recognizes the iron deficiency-responsive element 2 regulates the genes involved in iron homeostasis in plants. J Biol Chem. 2008;283:13407–13417. doi: 10.1074/jbc.M708732200. [DOI] [PubMed] [Google Scholar]
- Ogo Y, Itai RN, Kobayashi T, Aung MS, Nakanishi H, Nishizawa NK. OsIRO2 is responsible for iron utilization in rice and improves growth and yield in calcareous soil. Plant Mol Biol. 2011;75:593–605. doi: 10.1007/s11103-011-9752-6. [DOI] [PubMed] [Google Scholar]
- Ogo Y, Kakei Y, Itai RN, Kobayashi T, Nakanishi H, Takahashi H, Nakazono M, Nishizawa NK. Spatial transcriptomes of iron-deficient and cadmium-stressed rice. New Phytol. 2014;201:781–794. doi: 10.1111/nph.12577. [DOI] [PubMed] [Google Scholar]
- Pires N, Dolan L. Origin and diversification of basic-helix-loop-helix proteins in plants. Mol Biol Evol. 2010;27:862–874. doi: 10.1093/molbev/msp288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Celma J, Chou H, Kobayashi T, Long TA, Balk J. Hemerythrin E3 ubiquitin ligases as negative regulators of iron homeostasis in plants. Front Plant Sci. 2019;10:98. doi: 10.3389/fpls.2019.00098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Celma J, Connorton JM, Kruse I, Green RT, Franceschetti M, Chen YT, Cui Y, Ling HQ, Yeh KC, Balk J. Arabidopsis BRUTUS-LIKE E3 ligases negatively regulate iron uptake by targeting transcription factor FIT for recycling. Proc Natl Acad Sci USA. 2019 doi: 10.1073/pnas.1907971116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Römheld V, Marschner H. Evidence for a specific uptake system for iron phytosiderophore in roots of grasses. Plant Physiol. 1986;80:175–180. doi: 10.1104/pp.80.1.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roschzttardtz H, Conéjéro G, Curie C, Mari S. Identification of the endodermal vacuole as the iron storage compartment in the Arabidopsis embryo. Plant Physiol. 2009;151:1329–1338. doi: 10.1104/pp.109.144444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato Y, Antonio BA, Namiki N, Takehisa H, Minami H, Kamatsuki K, Sugimoto K, Shimizu Y, Hirochika H, Nagamura Y. RiceXPro: a platform for monitoring gene expression in japonica rice grown under natural field conditions. Nucleic Acids Res. 2011;39:D1141–1148. doi: 10.1093/nar/gkq1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato Y, Takehisa H, Kamatsuki K, Minami H, Namiki N, Ikawa H, Ohyanagi H, Sugimoto K, Antonio B, Nagamura Y. RiceXPro Version 3.0: expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 2013;41:D1206–D1213. doi: 10.1093/nar/gks1125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selote D, Samira R, Matthiadis A, Gillikin JW, Long TA. Iron-binding E3 ligase mediates iron response in plants by targeting bHLH transcription factors. Plant Physiol. 2015;167:273–286. doi: 10.1104/pp.114.250837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selote D, Matthiadis A, Gillikin JW, Sato MH, Long TA. The E3 ligase BRUTUS facilitates degradation of VOZ1/2 transcription factors. Plant Cell Environ. 2018;41:2463–2474. doi: 10.1111/pce.13363. [DOI] [PubMed] [Google Scholar]
- Sheng Y, Laister RC, Lemak A, Wu B, Tai E, Duan S, Lukin J, Sunnerhagen M, Srisailam S, Karra M, Benchimol S, Arrowsmith CH. Molecular basis of Pirh2-mediated p53 ubiquitylation. Nat Struct Mol Biol. 2008;15:1334–1342. doi: 10.1038/nsmb.1521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieker LC, Stenkamp RE, Jensen LH, Prickril B, LeGall J. Structure of rubredoxin from the bacterium Desulfovibrio desulfuricans. FEBS Lett. 1986;208:73–76. doi: 10.1016/0014-5793(86)81535-6. [DOI] [PubMed] [Google Scholar]
- Sivitz AB, Hermand V, Curie C, Vert G. Arabidopsis bHLH100 and bHLH101 control iron homeostasis via a FIT-independent pathway. PLoS ONE. 2012;7:e44843. doi: 10.1371/journal.pone.0044843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takagi S. Naturally occurring iron-chelating compounds in oat- and rice-root washing. I. Activity measurement and preliminary characterization. Soil Sci Plant Nutr. 1976;22:423–433. [Google Scholar]
- Tanabe N, Noshi M, Mori D, Nozawa K, Tamoi M, Shigeoka S. The basic helix-loop-helix transcription factor, bHLH11 functions in the iron-uptake system in Arabidopsis thaliana. J Plant Res. 2019;132:93–105. doi: 10.1007/s10265-018-1068-z. [DOI] [PubMed] [Google Scholar]
- Toledo-Ortiz G, Huq E, Quail PH. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell. 2003;15:1749–1770. doi: 10.1105/tpc.013839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urzica EI, Casero D, Yamasaki H, Hsieh SI, Adler LN, Karpowicz SJ, Blaby-Haas CE, Clarke SG, Loo JA, Pellegrini M, Merchant SS. Systems and trans-system level analysis identifies conserved iron deficiency responses in the plant lineage. Plant Cell. 2012;24:3921–3948. doi: 10.1105/tpc.112.102491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang HU, Klatte M, Jakoby M, Bäumlein H, Weisshaar B, Bauer P. Iron deficiency-mediated stress regulation of four subgroup Ib BHLH genes in Arabidopsis thaliana. Planta. 2007;226:897–908. doi: 10.1007/s00425-007-0535-x. [DOI] [PubMed] [Google Scholar]
- Wang N, Cui Y, Liu Y, Fan H, Du J, Huang Z, Yuan Y, Wu H, Ling HQ. Requirement and functional redundancy of Ib subgroup bHLH proteins for iron deficiency responses and uptake in Arabidopsis thaliana. Mol Plant. 2013;6:503–513. doi: 10.1093/mp/sss089. [DOI] [PubMed] [Google Scholar]
- Yuan Y, Wu H, Wang N, Li J, Zhao W, Du J, Wang D, Ling HQ. FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res. 2008;18:385–397. doi: 10.1038/cr.2008.26. [DOI] [PubMed] [Google Scholar]
- Zhang J, Liu B, Li M, Feng D, Jin H, Wang P, Liu J, Xiong F, Wang J, Wang HB. The bHLH transcription factor bHLH104 interacts with IAA-LEUCINE RESISTANT3 and modulates iron homeostasis in Arabidopsis. Plant Cell. 2015;27:787–805. doi: 10.1105/tpc.114.132704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H, Li Y, Yao X, Liang G, Yu D. Positive regulator of iron homeostasis1, OsPRI1, facilitates iron homeostasis. Plant Physiol. 2017;175:543–554. doi: 10.1104/pp.17.00794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng L, Ying Y, Wang L, Wang F, Whelan J, Shou H. Identification of a novel iron regulated basic helix-loop-helix protein involved in Fe homeostasis in Oryza sativa. BMC Plant Biol. 2010;10:166. doi: 10.1186/1471-2229-10-166. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.