

Abstract
During patient colonization, Staphylococcus aureus is able to invade and proliferate within human cells to evade the immune system and last resort drugs such as vancomycin. Hijacking specific host molecular factors and/or pathways is necessary for pathogens to successfully establish an intracellular infection. In this study, we employed an unbiased shRNA screening coupled with ultra-fast sequencing to screen 16,000 human genes during S. aureus infection and we identified several host genes important for this intracellular pathogen. In addition, we interrogated our screening results to find novel host-targeted therapeutics against intracellular S. aureus. We found that silencing the human gene TRAM2 resulted in a significant reduction of intracellular bacterial load while host cell viability was restored, showing its importance during intracellular infection. Furthermore, TRAM2 is an interactive partner of the endoplasmic reticulum SERCA pumps and treatment with the SERCA-inhibitor Thapsigargin halted intracellular MRSA survival. Our results suggest that Thapsigargin could be repurposed to tackle S. aureus host cell infection in combination with conventional antibiotics.
Subject terms: Bacterial host response, Cellular microbiology
Introduction
Staphylococcus aureus is a facultative intracellular pathogen capable of surviving in a wide range of human cells1. It is becoming clear that the intracellular survival of S. aureus has an important role on nasal colonization2. In persistent carriers, this could lead to opportunistic infections, increasing the risk of mortality and medical costs3. Moreover, intracellular S. aureus may contribute to persistent rhinitis, recurrent tonsillitis and chronic osteomyelitis4–8. Importantly, many last resort antibiotics do not enter efficiently into the host cell to achieve intracellular killing9. Therefore, novel strategies to control intracellular S. aureus and patient decolonization are urgently needed2.
There are different strategies available to determine host factors hijacked by intracellular pathogens, combining genomics, computational biology, proteomics and transcriptomics10. These strategies could lead to the identification or development of new host-targeted treatments to combat intracellular infections. In particular, loss-of-function phenotypic analysis, such as RNA interference (RNAi), is a common procedure to identify host genes or proteins that are necessary for the pathogen internalization, growth or survival within mammalian cells11–13.
The assessment of gene function using RNAi approaches relies on degradation of specific messenger RNAs (mRNAs) to silence gene expression. In mammalian cells, RNAi is usually mediated by microRNAs (miRNAs) that are non-protein coding transcripts. shRNAs are miRNAs that are processed into shorter RNAs and contain a short-hairpin structure. These shRNAs are then further processed into short double-stranded pieces of RNA called short-interfering RNAs (siRNAs). The molecular mechanism of gene silencing comprises the binding of one strand of the siRNA duplex to a protein-coding mRNA transcript within the multi-protein RNA-Induced Silencing Complex (RISC). Consequently, this interaction triggers cleavage of the protein-coding mRNA by a nuclease in the RISC complex, thereby destroying the mRNA and silencing the expression of the gene14.
One of the main advantages of using lentiviral vectors encoding and expressing shRNAs, when compared to siRNAs, is their ability to stably integrate into the genome of mammalian cells, allowing the generation of established knockdown cell lines (e.g. after puromycin selection) and therefore longer experimental settings14,15. Furthermore, the availability of commercial shRNA libraries constitutes a powerful tool that allows large-scale loss-of-function genetic screenings in mammalian cells14,16.
We recently identified new host-directed therapeutics against intracellular S. aureus by means of metabolomics and high-throughput drug screening17,18. In this study, we employed an shRNA screening approach coupled to Illumina sequencing-based deconvolution to identify novel host-genes that could be involved in S. aureus cell infection. Our results indicate that TRAM2 is a promising host target for the development of new anti-infectives against intracellular MRSA.
Results
Identification of novel host-genes involved in S. aureus cell infection
We tested the effect of silencing 16,000 human genes during S. aureus cell infection by using the Mission® LentiPlex® Human Pooled shRNA Library (Fig. 1). Most of the genes were targeted by at least 5 different shRNA constructs to ensure reproducibility (Supplementary Fig. 1). We transduced HeLa cells with the shRNA library and performed puromycin selection to obtain cells stably expressing the library (shRNA-HeLa cells; Fig. 1). Subsequently, we infected shRNA-HeLa cells with the clinically relevant S. aureus NCTC 13626 strain to investigate the effect of silencing specific genes on intracellular MRSA infection; vancomycin was added after one hour of incubation to kill extracellular bacteria17, and uninfected shRNA-HeLa cells were used as negative control. After six hours of infection, we extracted total genomic DNA to determine by Illumina sequencing the total occurrence of each shRNA integrated on the shRNA-HeLa cells after infection (Fig. 1). By calculating the ratio between the occurrence of all shRNA constructs targeting the same gene found in S. aureus-infected cells (sample) versus the uninfected condition (control) (Supplementary Table 1), we identified 5,674 shRNAs that were under-represented after S. aureus infection, whereas 9,997 shRNAs were over-represented when compared to uninfected cells (Supplementary Fig. 2).
Figure 1.

Layout of shRNA screening approach. HeLa cells were first transduced with shRNA library until stable cells were produced (shRNA-HeLa). shRNA-HeLa cells were infected with MRSA and genomic DNA was extracted to determine by Illumina sequencing the occurrence of each shRNA after infection. Uninfected HeLa cells were used as negative control.
Host cellular death is the last step of a successful intracellular infection, thus over-represented shRNA vectors may select for host cells that halt intracellular MRSA by silencing host genes that support the cell invasion or intracellular replication of S. aureus. In other words, over-represented shRNA vectors indicate an increase in host cell viability (via their copy number in the population of infected cells). The targets of these over-represented shRNAs, i.e. transcripts encoding proteins used by the pathogen to enter or proliferate inside the host cells, represent the main focus of our downstream analysis.
As an initial step, shRNA abundance/host cell viability data were analysed based on their significance and magnitude of change, and 38 transcript targets were highlighted after applying a strict cut-off (Supplementary Fig. 3). However, the silencing effect of empirically-designed shRNA vectors could be limited by their variable efficiency and specificity19. Therefore, we further interrogated our screening results by considering the percentage of shRNAs that were under- or over-represented for each gene to estimate the consistency of the silencing effect. This is important because a consistent silencing effect produced by multiple shRNA constructs targeting the same gene is less likely to be due to an off-target effect15. We then established several cut-offs to identify the most interesting gene candidates in our model of infection, which included a mean ratio greater than 1 (Log Ratio >1) compared to uninfected cells, a standard error below 2 (SE <2) and a consistency over 60% (i.e. 60% of gene-specific shRNAs caused an effect in the same direction). In total, 2,888 (≈18% of all genes) met these criteria (Fig. 2A). The identified genes were functionally classified into 63 cellular pathways using DAVID Bioinformatics Resources 6.820 (Supplementary Fig. 4 and Supplementary Table 2). We identified overrepresented pathways that may be important during S. aureus cell infection, e.g. regulation of autophagy, regulation of actin cytoskeleton, MAPK signalling and Toll-like receptor signalling pathways.
Figure 2.
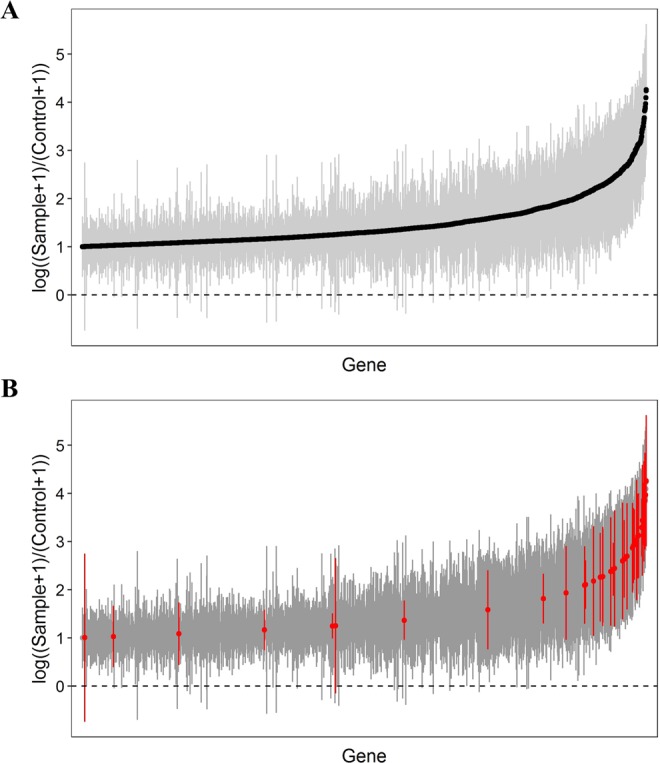
shRNA screening data after applying a strict cut-off. (A) shRNA screening data were filtered by establishing three criteria: mean ratio higher than 1 (Log Ratio >1) compared to the uninfected control, standard error lower than 2 (SE <2) and a consistency higher than 60%. (B) Out of 2,888 genes, twenty-nine host genes were selected for further validation. High effect in the shRNA screening after MRSA infection and/or a biological relevance for intracellular pathogenesis were used as criteria to choose the most interesting hits. Selected genes are highlighted in red.
Following the aforementioned principles (i.e. magnitude and significance of change, consistent silencing effect and biological relevance), we selected a total of 29 genes for further validation (Fig. 2B and Supplementary Table 3). By choosing these genes, we covered a diverse range of cellular pathways known to be exploited by MRSA during intracellular infection and - importantly - novel genes whose functions (and/or involvement in MRSA invasion) may not have been thoroughly described.
Among the shortlisted genes candidates, the Autophagy Related 10 (ATG10) gene is directly involved in cell autophagy21,22. Other genes are associated with programmed cell death, such as Apoptosis and Caspase Activation Inhibitor (AVEN) and CD5 molecule-like (CD5L) genes. RAB genes are linked to membrane trafficking23 and, indeed, several intracellular pathogens hijack host-RAB proteins during infection24. The components of the cell cytoskeleton also play an important role during intracellular infection, and we identified at least three proteins, CLTB and MYL2/9, which may be involved in cytoskeleton rearrangement25. We also observed proteins linked to the host cell metabolism, such as the branched-chain amino acids regulator (BCKDK) and the amino acids transporters SLC43A1 and SLC63A3, which could be important for intracellular MRSA survival17. Additionally, we also included the CD83-molecule (CD83) and Toll-Like Receptor 2 (TLR2) genes in our downstream analysis, which are involved in a wide range of host immune responses to fight against bacterial and viral infections26,27.
To validate our results, we used individual shRNA constructs with the strongest silencing effect for each gene to produce individual knockdowns in HeLa cells for further investigation. An established cell line expressing an shRNA that does not silence any human gene (Non-target HeLa cells) was used as a negative control in all downstream experiments.
We first assessed the host cell viability of the selected 29 knockdowns after MRSA infection (Fig. 3 and Supplementary Table 3). We performed infection assays with two different strains of MRSA – S. aureus NCTC 13626 (Healthcare-Acquired MRSA) and S. aureus USA300 LAC (Community-Acquired MRSA; Fig. 3) – in the presence of vancomycin and gentamycin respectively, since NCTC 13626 is resistant to gentamycin17. We selected CD83, FAM63B, MYL2 and TRAM2 knockdowns for further analysis since the silencing of these four host genes resulted in increased host cell viability regardless of the strain that was employed, and these genes were not studied before in relation to S. aureus cell infection.
Figure 3.

Validation of the screening results. Host cell viability after intracellular infection with S. aureus NCTC 13626 strain (A) and S. aureus USA300 LAC strain (B). After 6 hours of MRSA infection, cell viability was quantified by flow cytometry using a double staining of Annexin V-FITC and propidium iodide. Cell viability was normalized in relation to the uninfected control. Data are expressed by means ± standard error (SE) of two biological replicates performed in duplicates. One-way ANOVA and multiple comparison Tukey’s tests were performed to assess statistically significant differences between each knockdown and the non-target control. p-value ≤ 0.05 (*); ≤0.01 (**); ≤0.001 (***). In order to recognize the most interesting hits, we selected knockdowns whose cell viability was at least 1.2-fold higher than non-target cells after infection with both strains.
We then assessed the intracellular bacterial load of S. aureus USA300 in CD83, FAM63B, MYL2 and TRAM2 knockdowns in parallel to host cell viability (Fig. 4). We observed a clear inhibitory effect on the intracellular growth in every knockdown tested in comparison to the non-targeted control, with the most pronounced effects after silencing FAM63B, MYL2 or TRAM2 (Fig. 4A). We finally found drug inhibitors linked to two proteins that were consistently identified in all tests: MYL2 and TRAM2, which were chosen for further analysis.
Figure 4.

Bacterial and host cell viability after S. aureus USA300 infection in CD83, FAM63B, MYL2, and TRAM2 knockdowns. (A) Intracellular bacteria were quantified by colony forming units (CFU) after 6 hours of infection. (B) Cell viability was quantified in parallel after 6 hours of infection by flow cytometry, using a double staining of Annexin V-FITC and propidium iodide. Uninfected HeLa cells were employed as negative control and cell viability was normalized in relation to the control’s viability. Data are expressed by means ± standard error (SE) of three experiments performed in duplicates. One-way ANOVA and multiple comparison Tukey’s tests were performed to assess statistically significant differences between each knockdown and the non-target control. p-value ≤ 0.05 (*); ≤0.01 (**); ≤0.001 (***).
Thapsigargin impairs intracellular S. aureus infection
Blebbistatin inhibits the ATPase activity of myosin II and stops motility based on actomyosin association28. However, the treatment with this inhibitor had no effect on the host cell viability during infection (Supplementary Fig. 5), discarding MYL2 for further downstream experiments.
On the other hand, TRAM2 is part of the translocon, a complex of proteins involved in the transport of polypeptides across the endoplasmic reticulum (ER) membrane29. The C-terminal end of TRAM2 interacts with the Ca2+ pump of the endoplasmic reticulum SERCA2b, which is necessary for the correct protein folding of proteins such as collagen type I29. Based on this evidence, we hypothesized that by silencing TRAM2 in HeLa cells we may in turn block Ca2+ pumps of the ER (e.g. SERCA2b), and consequently alter the maturation of proteins that are essential for S. aureus during cell infection. In particular, we speculated with the possibility that TRAM2 depletion or SERCA inhibition may lead to an alteration in collagen type I folding and consequently its increased intracellular degradation, as previously reported29. This in turn could affect the adhesion of S. aureus to the host tissues through the collagen-binding adhesin Cna30, which may lead to a lower internalization of S. aureus into the host cell.
To test this hypothesis, we performed S. aureus infection assays in the presence of Thapsigargin, a selective SERCA inhibitor31, and we measured the host cell viability and intracellular MRSA survival. We employed a low concentration of this inhibitor (0.1 µM), because higher doses have been reported to be cytotoxic32,33. In the presence of Thapsigargin, host cell viability was enhanced at 6 hours post-infection both in human umbilical vein endothelial cells (HUVECs) and HeLa cells (Fig. 5). This is important, because host-cell interactions may profoundly differ depending on the cell line tested in the context of S. aureus cell infection1.
Figure 5.

Thapsigargin treatment during S. aureus infection increases host cell viability whereas the intracellular bacterial load is reduced. HeLa cells and HUVEC were infected with S. aureus USA300-GFP (MOI 100) for 2 and 6 hours in the presence of DMSO (D) or 0.1 µM of Thapsigargin (T). Host cell viability in HeLa (A) and HUVEC (B) was quantified after 6 hours of infection by flow cytometry. Cell viability was normalized by the percentage of uninfected and untreated cells (U). (C) Intracellular MRSA survival was quantified by measuring the percentage of host cells with intracellular S. aureus USA300-GFP after 2 and 6 hours of infection. Data are expressed as means ± standard errors (SE) of at least two different experiments performed in duplicates and Student’s t-tests were performed to validate statistical significance across conditions. p-value ≤ 0.05 (*); ≤0.001 (***).
In concordance to the observed effect on host cell viability, intracellular MRSA survival was reduced by two-fold even at early time points of infection when infected HeLa cells where treated with Thapsigargin (Fig. 5C). On the other hand, we did not observe any significant differences in S. aureus growth curves in vitro in the presence of Thapsigargin, ruling out a direct inhibitory effect of the drug on S. aureus (Supplementary Fig. 6).
Our findings suggested that Thapsigargin halts S. aureus internalization in HeLa cells. However, this is not mediated by an altered production of collagen type 1, since COL1A2 (Collagen α2 Type I) levels were unaffected during S. aureus infection in wild type cells treated with Thapsigargin or in the TRAM2 knockdown, where TRAM2 protein levels are reduced by 50% when compared to a non-targeted control (Supplementary Fig. 7). Moreover, TRAM2 silencing did not have an effect during early time points of infection (i.e. 2 hours post-infection), pointing towards a role on intracellular proliferation rather than cell entry (Supplementary Fig. 8). Interestingly, autophagy is significantly reduced in response to TRAM2 depletion or Thapsigargin treatment in S. aureus infected cells (Supplementary Fig. 7e), suggesting a role of TRAM2 on the activation of the autophagic machinery.
Characterization of the host gene TRAM2 on S. aureus cell infection
To shed light on the role of TRAM2 on S. aureus intracellular survival, we have first evaluated the effect of S. aureus infection on the protein levels of TRAM2 in wild type HeLa cells at different time points and multiplicity of infections (MOI; Supplementary Fig. 9). As a control of bacterial load, we detected S. aureus Protein A by Western-blot, whose levels directly correlate with increasing times of infection and MOI (Supplementary Fig. 9a). Protein levels of TRAM2 were increased after 2 hours of S. aureus infection to become up to 40% higher in infected HeLa cells (Supplementary Fig. 9b). However, at 6 hours post-infection, we noticed a change in the migration pattern of TRAM2 upon denaturing protein electrophoresis that apparently generates a band of lower molecular weight, indicating that TRAM2 may have been subjected to post-translational modification such as proteolysis (Supplementary Fig. 9a). This change in the migration pattern was proportional to the MOI employed for the infection, suggesting that S. aureus may be interfering with the post-translational modification of TRAM2 to promote its own survival or replication. This strategy is commonly employed by many bacterial pathogens to control host proteins during intracellular infection34.
Given that TRAM2 is a membrane protein, we hypothesized that it may be involved in the maturation of the vacuole containing S. aureus. Therefore, HeLa cells stably expressing TRAM2-mCherry were infected with S. aureus USA300-GFP35. In most of the cases, we observed large sacs of S. aureus cells surrounded by a scattered pattern of TRAM2-mCherry (Fig. 6A), which was not considered a conclusive pattern of colocalization. However, some individual bacterial cells were enclosed by a membrane containing TRAM2-mCherry (Fig. 6B). Nevertheless, these clear co-localization patterns of S. aureus-TRAM2 were restricted to approximately 9% of infected cells (Fig. 6C). Taken together the low number of co-localization events, we can only conclude that either TRAM2 co-localization is highly dynamic or this protein may have other roles during cell infection rather than on the maturation of the vacuole containing intracellular S. aureus.
Figure 6.

Co-localization of MRSA-GFP and TRAM2-mCherry by confocal microscopy. TRAM2-mCherry HeLa cells were infected with S. aureus USA300-GFP (MOI 100) and samples were collected after 6 hours of infection for confocal microscopy. DAPI staining was employed for nuclear staining. (A) Confocal picture showing a representative pattern of TRAM2-mCherry localization in S. aureus infected cells. (B) White arrows depict where co-localization between MRSA and TRAM2 was observed. (C) Percentage of infected cells showing co-localization of USA300-GFP and TRAM2-mCherry.
Discussion
A functional genomics approach identifies novel host-genes important for S. aureus during cell infection
The coupling of RNA interference (RNAi) with ultra-fast sequencing provides an exceptional opportunity to simultaneously screen the implication of thousands of host-genes in different areas of research. In particular, RNAi approaches have been employed before to screen novel host-genes implicated in intracellular bacterial36, parasitical37 and viral38 infections.
The main limitation of this high-throughput approach is the possibility of encountering phenotypes caused by off-target effects. To minimize this, transductions should be performed with a low multiplicity of infection14, and different shRNA constructs targeting the same gene should be included in the pooled libraries16. Accordingly, we performed lentiviral transductions using a MOI of 1, and most genes included in the commercial shRNA library were targeted by at least 5 shRNA constructs.
By employing ultra-fast sequencing, we were able to rapidly estimate the occurrence of shRNAs in S. aureus-infected cells in relation to the uninfected control. Over-represented shRNA constructs may be silencing human genes that are important for the intracellular bacterial survival and thus, any of these genes could have an essential role on S. aureus cell infection. We found approximately 10,000 human genes that were targeted by shRNAs over-represented in HeLa cells at 6 hours post-infection.
Further validation of our preliminary screening results was necessary and thus, the number of potential candidates was filtered. A combination of high consistency of the silencing effect, magnitude of the change and biological relevance was employed to select genes for further downstream analysis. As a result, 29 human genes were selected for further validation by producing individual knockdowns in HeLa cells and testing host cell viability after intracellular MRSA infection. Among these genes, we included a varied range of cellular processes, such as autophagy, apoptosis, membrane trafficking and cytoskeleton rearrangement, amino-acids metabolism and host-immune responses. Many of these host cell pathways have previously been described to play a role in intracellular infection and therefore, we evaluated how host cell viability was affected during S. aureus infection after individually silencing these host-genes.
Of particular significance is ATG10, which silencing significantly restored cell viability after infection with two different MRSA strains (Fig. 3). This is in agreement with findings from our and other groups, suggesting that intracellular S. aureus infection in non-phagocytic cells is halted with autophagy inhibitors such as dorsomorphin17, while 3-Methyladenine treatment protects mice from MRSA pneumonia39.
To further identify other host-targeted therapeutics against intracellular MRSA, we interrogated our shRNA screening results to identify four genes, i.e. CD83, FAM63B, MYL2 and TRAM2, whose silencing in HeLa cells resulted in a significant reduction of intracellular S. aureus survival while host cell viability was enhanced after cell infection.
The protein encoded by CD83 gene is a host membrane protein that is part of the immunoglobulin family and it is mainly expressed in mature dendritic cells (DCs). The main function of these antigen-presenting cells is the activation of host immune responses against invading pathogens40. Eukaryotic cells have evolved complex immune mechanisms to combat invading pathogens. Consequently, and in order to successfully establish infections, intracellular microorganisms have also developed sophisticated mechanisms to subvert and overcome host immune responses41. Specifically, it has been shown that interleukin 10 (IL-10) – a cytokine produced by macrophages, T-cells, B-cells, mast cells and keratinocytes, among others – can be an effective immunosuppressive factor and therefore, the exploitation of this cytokine is a common mechanism to evade host immune responses by several pathogens. Some viruses induce the production of host IL-10, while others produce their own IL-10-homologs. Bacterial intracellular pathogens, such as Mycobacterium tuberculosis and Listeria monocytogenes, are able to replicate within macrophages by inducing IL-1042. However, further investigation is needed to explain the molecular mechanism behind CD83-S. aureus interaction in cells that are not part of the immune system.
Although its function is unknown, FAM63B has been recently identified as an interaction partner of kinesin light chain-1 (KLC1), which is involved in the intracellular trafficking of vaccinia virus43. Kinesins, dyneins and myosins comprise three classes of molecular motors that are part of the host cell cytoskeleton and are involved in many biological processes related to cell movement. Intracellular bacterial pathogens and viruses commonly use this host machinery to reach their intracellular niche, as well as to control the membrane remodelling of their containing vacuoles44. In particular, Kinesin-1 has been implicated in the intracellular dissemination of adenovirus and Herpes virus43,45,46. Moreover, Salmonella effector protein SifA targets the host protein SKIP, which down-regulates the recruitment of Kinesin-1 to the Salmonella-containing vacuole and thus, controls its dynamics47. Therefore, FAM63B may play a role in the intracellular trafficking of S. aureus by targeting kinesin proteins.
Myosins comprise a superfamily of motor proteins found in actin filaments and have an essential role in the organization of actin cytoskeleton. Polymerization and depolymerization of actin filaments promote changes in cells shape and, along with myosin proteins, control the intracellular organization25. The host cell cytoskeleton is commonly hijacked by intracellular pathogens to support their own intracellular invasion, survival and replication and actin has been specifically described as a common target of many bacterial pathogens48. As above mentioned, bacterial pathogens exploit host-actin for different purposes such as intracellular invasion, actin-based motility through cell cytoplasm and pathogen dissemination49. Likewise, the use of microtubules, motor proteins and intermediate filaments in intracellular invasion and dissemination has been highlighted49. For instance, the actin-base motor protein myosin 2 has been associated with the integrity and location of Salmonella-containing vacuoles50. In addition, myosin 2 has been associated with the promotion of vesicle fission from the Golgi complex51. Furthermore, there is a proportion of cytosolic Shigella that become covered with septin filaments in a myosin 2-dependent manner52. Moreover, the release of Chlamydia-intrusions into the extracellular space is mediated by actin, N-WASP, myosin 2 and GTPase Rho proteins53. Collectively, these studies suggest that the intracellular fate of many intracellular pathogens is determined by an organized collaboration between actin, microtubules, intermediate filaments and motor proteins. Therefore, the gene coding for the myosin light chains 2 (MYL2) could be involved in the intracellular dissemination of S. aureus by controlling host actin cytoskeleton. However, a treatment with the myosin II inhibitor Blebbistatin did not restore the host cell viability during S. aureus infection (Supplementary Fig. 5), suggesting a more complex scenario.
TRAM2 plays an important role during intracellular MRSA survival
TRAM2 (Translocating chain-associated membrane protein 2) was initially identified as being responsible for the translocation of proteins across the endoplasmic reticulum membrane54. Furthermore, TRAM2 interacts with the endoplasmic reticulum Ca2+ ATPase transporter SERCA2b, being involved in collagen type I protein folding29.
Different intracellular pathogens rely on the endoplasmic reticulum and its components to ensure intracellular survival and proliferation55. For instance, Legionella pneumophila, an intravacuolar pathogen, hijacks host membrane components to contribute to the formation of Legionella-containing vacuoles that are mainly derived from endoplasmic reticulum material56,57. Similarly, Brucella abortus58,59 and Chamydia trachomatis60,61 also exploit host-endoplasmic reticulum to promote their intracellular survival and replication within mammalian cells. In addition, SERCA2b has been closely associated with Chlamydial infections in HeLa cells62. In concordance with these observations, co-localization assays showed that several chlamydial antigens, such as major outer membrane protein (MOMP), inclusion membrane protein and lipopolysaccharides (LPS), were specifically associated with the ER and ER-markers63.
Because of the interaction of TRAM2 and SERCA transporters, we investigated the intracellular S. aureus survival under treatment with Thapsigargin – a promising anticancer drug that is a specific host-SERCA inhibitor31,64. High doses of Thapsigargin lead to host cell death due to the ER stress caused by this inhibitor33. In contrast, treatment of infected HeLa cells with low doses of Thapsigargin resulted in a significantly attenuated bacterial load at both time points of infection (at 2 and 6 hours post infection, Fig. 5), whereas this reduction was only observed after 6 hours of infection in TRAM2 knockdown cells (Supplementary Fig. 8). Therefore, Thapsigargin may have an effect on cell entry, whereas TRAM2 depletion is not altering cell invasion but it can halt S. aureus intracellular replication. Nonetheless, our findings suggest that the anti-cancer drug Thapsigargin may be repurposed to block S. aureus cell infection when combined with conventional antibiotics. On the other hand, neither TRAM2 depletion nor Thapsigargin treatment had any effects on type I collagen production during host cell infection, pointing towards an alternative role of TRAM2 on S. aureus intracellular survival in this context. In concordance with this hypothesis, we have not observed any significant changes in the occurrence of shRNAs in S. aureus-infected cells that are silencing essential components of the translocon such as SEC61A1 or SEC61G (Supplementary Table 1), suggesting that the role of TRAM2 on the intracellular survival of S. aureus is not related to protein translocation across the endoplasmic reticulum membrane. Interestingly, we have observed autophagy inhibition during S. aureus infection in the context of TRAM2 depletion or Thapsigargin treatment, but this effect may be indirect due to a lower bacterial load. Undoubtedly, more research is needed to fully understand the role of TRAM2 during S. aureus infection.
Conclusions
By using a functional genomics approach, we screened the implication of 16,000 host-genes on intracellular S. aureus infection to identify many potential host targets that could be hijacked by S. aureus during intracellular infection. Here, we studied the human gene TRAM2, which plays an important role during S. aureus infection. The data presented in this study serve as proof-of-principle of a functional genomics approach to unravel novel molecular mechanisms hijacked by S. aureus during intracellular proliferation in human cells.
Materials and Methods
Bacterial strains, cell lines and culture conditions
S. aureus NCTC 1362665, USA300 LAC66 and USA300-GFP35 strains were cultured and pre-inocula for infection assays were prepared as previously described17. Briefly, bacterial culture was grown from an overnight culture until an optical density (OD600 nm) of 1 was reached, which corresponds to the mid-exponential phase (Supplementary Fig. 10). Cultures were centrifuged, and pellets were washed before being resuspended in PBS supplemented with 20% glycerol to aliquot and store at −80 °C until needed.
HeLa cells (ECACC 93021013) and their derivatives were incubated at 37 °C and 5% of CO2 in Dulbecco’s Modified Eagle’s medium (DMEM, Gibco) containing pyruvate, glucose and glutamine and supplemented with 10% heat-inactivated foetal bovine serum (FBS, Gibco) and 5% of penicillin and streptomycin solution (Gibco), unless otherwise specified. Human umbilical vein endothelial cells (HUVECs; Sigma) were grown in endothelial cell growth medium (Sigma).
shRNA screening
HeLa cells were transduced with the Mission® LentiPlex Human shRNA Library (Sigma-Aldrich). This library is comprised by 10 sub-pools of lentiviral particles carrying around 75,000 shRNA constructs that target 16,000 human genes. HeLa cells were seeded in ten different 100 mm plates (Sarstedt) in complete DMEM medium at a cell density of 2 × 106 cells per dish and placed in the incubator overnight (37C, 5% CO2). The following day, the ten-different lentiviral subpools were thawed on ice and added to the HeLa cells at a MOI of 1 in the presence of hexadimethrine bromide (8 µg/ml; Sigma-Aldrich). After an overnight incubation, the virus-containing media were replaced with fresh DMEM without hexadimethrine bromide. The next day, HeLa cells were exposed to puromycin selection (1 µg/ml; Sigma-Aldrich) and medium was replaced with fresh puromycin-containing media every two days until only positive transduced-cells remained.
HeLa cell lines stably expressing the shRNA library were infected with S. aureus NCTC 13626 at a MOI of 100. After 6 hours of infection, total DNA was extracted from uninfected and S. aureus-infected cells by using GenElute Mammalian Genomic DNA Miniprep Kit (Sigma-Aldrich), following manufacturer’s instructions. Genomic DNA samples were submitted to the Sigma Deconvolution platform, and the abundance of each shRNA clone in samples was tested by amplifying and next-generation sequencing (at × 1,000) shRNA regions and barcoding samples; short reads were aligned to the reference. Data were obtained as the number of shRNA sequences per clone per sample, as previously described67. Screening results are shown in Supplementary Table 1.
Production of individual knockdowns in HeLa cells
To produce individual knockdowns, HeLa cells were transduced with 29 individual shRNA lentiviral constructs (Supplementary Table 4, Sigma-Aldrich). HeLa cells were seeded in 24 wells-plates at a cell density of 8 × 104 cells per well and incubated overnight (37 °C, 5% CO2). Transduction of mammalian cells was performed as described above, employing a MOI of 1.
HeLa cells expressing TRAM2-mCherry were created by transduction with P12-MMP-TRAM2-mCherry, as previously described68. To create this vector, mCherry-LC3 was replaced with TRAM2-mCherry in P12-MMP-mCherry-LC368. TRAM2 was amplified from the cDNA Clone IRATp970H1249D (SourceBioscience) with primers TAGCTAAAGCTTGCCACCATGGCTTTCCGCAGGAGG and GCCCTTGCTCACCATGGGAGACTTGAGTTT, whereas mCherry was amplified from P12-MMP-mCherry-LC3 with AAACTCAAGTCTCCCATGGTGAGCAAGGGC and TAGCTAGCGGCCGCTTTACTTGTACAGCTCGTCCAT, and subjected to fusion PCR. The DNA amplicon was verified by sequencing, digested with HindIII/NotI, and cloned into P12-MMP to create P12-MMP-TRAM2-mCherry.
Intracellular infection assays
Intracellular infection assays were performed as described before17. For host cell viability assays, cells were double stained with annexin V-FITC and propidium iodide according to manufacturer’s recommendations (Becton Dickinson, BD), and host cell viability was measured by flow cytometry (BD AccuriTM C6 Plus) as previously described17. To estimate the intracellular bacterial load at specific time points after infection, cells were lysed using 0.1% Triton X-100 diluted in PBS (5 min, RT) and serial dilutions were plated in Nutrient Agar plates for colony forming unit (CFU) counting. For assays involving Thapsigargin treatment, a flow cytometry-based protocol developed for drug screening was applied as previously described18.
Western-blot
Western-blots were performed as described before17. Primary antibodies were purchased from Santa Cruz Biotechnologies (anti-GAPDH [reference no. sc-47724]; anti-COL1A2 [sc-166865]), Fisher (anti-LC3 [13278218]), and Novus Biologicals (anti-TRAM2 [NBP1-83052]). Secondary antibodies were purchased from Li-Cor (IRDye 680LT goat anti-mouse [926–68070] and IRDye 800LT goat anti-rabbit [926–32211]). Full-length blots are shown in Supplementary Fig. 11.
Confocal microscopy
HeLa cells expressing TRAM2-mCherry were seeded in coverslips at a cell density of 7 × 104 cells per well in 24 well plates and S. aureus USA300-GFP35 was used to infect the cells at a MOI of 100 as previously described18. After 6 hours of infection, cells were fixed with 0.5 ml of freshly prepared 4% of paraformaldehyde (PFA; Fisher Scientific) for 15 minutes. Coverslips were then washed twice with PBS and mounted on microscope slides (Thermo Fisher Scientific) using ProLong Gold Antifade mountant with DAPI (Thermo Fisher Scientific) for nuclear staining. Slides were observed on a confocal microscope ZEISS LSM 800 with Airyscan by using Zen Blue Software (Zeiss).
Statistical analyses
Statistical tests and graphs plotting were conducted using GraphPad Prism software and significant differences across treatments were assessed by running Student’s t-test and post hoc Tukey’s multiple comparison tests when required.
Supplementary information
Acknowledgements
We thank Dr. Cokro Leksmono and Ms Martha Villegas-Montes for technical assistance. This work was supported by a Roehampton Vice Chancellor’s Scholarship to NBS, a Royal Society Research Grant to M.L. (Ref. RG130658), a New Lecturer Research Grant of the Society for Applied Microbiology to V.B., and intramural funding from the University of Roehampton to M.L. and V.B. P.C.L. was funded by a BBSRC-funded South West Biosciences Doctoral Training Partnership (BB/M009122/1).
Author contributions
M.L. and V.B. conceived the experiments; M.L., V.B., L.M. and Y.C. were involved in the design of the experiments; N.B.S. performed the experiments and wrote the first draft of the manuscript; N.B.S. and P.C.L. analysed the data and performed the statistical analysis. All authors contributed to manuscript revision, read and approved the submitted version.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Volker Behrends, Email: Volker.Behrends@roehampton.ac.uk.
Michal Letek, Email: Michal.Letek@roehampton.ac.uk.
Supplementary information
is available for this paper at 10.1038/s41598-019-51894-3.
References
- 1.Fraunholz M, Sinha B. Intracellular Staphylococcus aureus: live-in and let die. Front. Cell. Infect. Microbiol. 2012;2:1–10. doi: 10.3389/fcimb.2012.00043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rigaill, J. et al. Intracellular activity of antimicrobial compounds used for Staphylococcus aureus nasal decolonization. J. Antimicrob. Chemother. 1–5, 10.1093/jac/dky318 (2018). [DOI] [PubMed]
- 3.Sakr A, Brégeon F, Mege J-L, Rolain J-M, Blin O. Staphylococcus aureus nasal colonization: an update on mechanisms, epidemiology, risk factors and subsequent infections. Front. Microbiol. 2018;9:1–15. doi: 10.3389/fmicb.2018.02419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sinha B, Herrmann M. Mechanism and consequences of invasion of endothelial cells by Staphylococcus aureus. Thromb Haemost. 2005;94:266–277. doi: 10.1160/TH05-04-0235. [DOI] [PubMed] [Google Scholar]
- 5.Rollin G, et al. Intracellular survival of Staphylococcus aureus in endothelial cells: a matter of growth or persistence. Front. Microbiol. 2017;8:1354. doi: 10.3389/fmicb.2017.01354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ou J, et al. The persistence of intracellular Staphylococcus aureus in the sinuses: a longitudinal study. Rhinology. 2017;55:305–311. doi: 10.4193/Rhin16.218. [DOI] [PubMed] [Google Scholar]
- 7.Yang D, et al. Novel insights into Staphylococcus aureus deep bone infections: The involvement of osteocytes. MBio. 2018;9:1–10. doi: 10.3391/mbi.2018.9.1.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zautner, A. E. et al. Intracellular persisting Staphylococcus aureus is the major pathogen in recurrent tonsillitis. PLoS One5 (2010). [DOI] [PMC free article] [PubMed]
- 9.Lehar SM, et al. Novel antibody–antibiotic conjugate eliminates intracellular S. aureus. Nature. 2015;527:323–328. doi: 10.1038/nature16057. [DOI] [PubMed] [Google Scholar]
- 10.Schwegmann A, Brombacher F. Host-directed drug targeting of factors hijacked by pathogens. Sci. Signal. 2008;1:1–8. doi: 10.1126/scisignal.129re8. [DOI] [PubMed] [Google Scholar]
- 11.Agaisse H, et al. Genome-wide RNAi screen for host factors required for intracellular bacterial infection. Science (80-.). 2005;309:1248–1252. doi: 10.1126/science.1116008. [DOI] [PubMed] [Google Scholar]
- 12.Ng TI, et al. Identification of host genes involved in Hepatitis C virus replication by Small Interfering RNA Technology. Hepatology. 2007;45:1413–1421. doi: 10.1002/hep.21608. [DOI] [PubMed] [Google Scholar]
- 13.Brass AL, et al. Identification of host proteins required for HIV infection through a functional genomic screen. Science (80-.). 2008;319:921–927. doi: 10.1126/science.1152725. [DOI] [PubMed] [Google Scholar]
- 14.Sims D, et al. High-throughput RNA interference screening using pooled shRNA libraries and next generation sequencing. Genome Biol. 2011;12:R104. doi: 10.1186/gb-2011-12-10-r104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Coussens, M. J., Corman, C., Fischer, A. L., Sago, J. & Swarthout, J. MISSION LentiPlex pooled shRNA library screening in mammalian cells. J. Vis. Exp. 3–9, 10.3791/3305 (2011). [DOI] [PMC free article] [PubMed]
- 16.Root DE, Hacohen N, Hahn WC, Lander ES, Sabatini DM. Genome-scale loss-of-function screening with a lentiviral RNAi library. Nat. Methods. 2006;3:715–719. doi: 10.1038/nmeth924. [DOI] [PubMed] [Google Scholar]
- 17.Bravo-Santano N, et al. Intracellular Staphylococcus aureus modulates host central carbon metabolism to activate autophagy. mSphere. 2018;3:e00374–18. doi: 10.1128/mSphere.00374-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bravo-Santano N, et al. Host-directed kinase inhibitors act as novel therapies against intracellular Staphylococcus aureus. Sci. Rep. 2019;9:1–12. doi: 10.1038/s41598-019-41260-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Khandelwal N, et al. A high-throughput RNAi screen for detection of immune-checkpoint molecules that mediate tumor resistance to cytotoxic T lymphocytes. EMBO Mol. Med. 2015;7:450–463. doi: 10.15252/emmm.201404414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009;4:44–57. doi: 10.1038/nprot.2008.211. [DOI] [PubMed] [Google Scholar]
- 21.Jung CH, Ro SH, Cao J, Otto NM, Kim DH. MTOR regulation of autophagy. FEBS Lett. 2010;584:1287–1295. doi: 10.1016/j.febslet.2010.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bernard A, Jin M, Xu Z, Klionsky DJ. A large-scale analysis of autophagy-related gene expression identifies new regulators of autophagy. Autophagy. 2015;8627:2114–2122. doi: 10.1080/15548627.2015.1099796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hutagalung AH, Novick PJ. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011;91:119–149. doi: 10.1152/physrev.00059.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Spanò S, Galán JE. Taking control: Hijacking of Rab GTPases by intracellular bacterial pathogens. Small GTPases. 2017;9:1–10. doi: 10.1080/21541248.2017.1336192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fletcher DA, Mullins RD. Cell mechanics and the cytoskeleton. Nature. 2010;463:485–492. doi: 10.1038/nature08908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Christiaansen A, Varga SM, Spencer JV. Viral manipulation of the host immune response. Curr. Opin. Immunol. 2015;36:54–60. doi: 10.1016/j.coi.2015.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mukherjee S, Karmakar S, Babu SPS. TLR2 and TLR4 mediated host immune responses in major infectious diseases: A review. Brazilian J. Infect. Dis. 2016;20:193–204. doi: 10.1016/j.bjid.2015.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kovács M, Tóth J, Hetényi C, Málnási-Csizmadia A, Sellers JR. Mechanism of Blebbistatin inhibition of Myosin II. J. Biol. Chem. 2004;279:35557–35563. doi: 10.1074/jbc.M405319200. [DOI] [PubMed] [Google Scholar]
- 29.Stefanovic B, Stefanovic L, Schnabl B, Bataller R, Brenner DA. TRAM2 protein interacts with endoplasmic reticulum Ca2+ pump Serca2b and is necessary for collagen type I synthesis. Mol. Cell. Biol. 2004;24:1758–1768. doi: 10.1128/MCB.24.4.1758-1768.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Herman-Bausier P, et al. Mechanical strength and inhibition of the Staphylococcus aureus Collagen-Binding Protein Cna. MBio. 2016;7:1–11. doi: 10.1128/mBio.01529-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lytton J, Westlin M, Hanley MR. Thapsigargin inhibits the sarcoplasmic or endoplasmic reticulum Ca-ATPase family of calcium pumps. J. Biol. Chem. 1991;266:17067–17071. [PubMed] [Google Scholar]
- 32.Stetler, C., Murali, R., Chen, E. & Miller, G. E. Endoplasmic-reticulum calcium depletion and disease. Cold Spring Harb. Perspect. Biol. 3 (2011). [DOI] [PMC free article] [PubMed]
- 33.Sehgal, P. et al. Inhibition of the sarco/endoplasmic reticulum (ER) Ca2+-ATPase by thapsigargin analogs induces cell death via ER Ca2+ depletion and the unfolded protein response. J. Biol. Chem. jbc.M117.796920, 10.1074/jbc.M117.796920 (2017). [DOI] [PMC free article] [PubMed]
- 34.Ribet D, Cossart P. Post-translational modifications in host cells during bacterial infection. FEBS Lett. 2010;584:2748–2758. doi: 10.1016/j.febslet.2010.05.012. [DOI] [PubMed] [Google Scholar]
- 35.Reichmann NT, et al. Differential localization of LTA synthesis proteins and their interaction with the cell division machinery in Staphylococcus aureus. Mol. Microbiol. 2014;92:273–286. doi: 10.1111/mmi.12551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jayaswal S, et al. Identification of host-dependent survival factors for intracellular Mycobacterium tuberculosis through an siRNA Screen. PLoS Pathog. 2010;6:e1000839. doi: 10.1371/journal.ppat.1000839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Moser LA, Pollard AM, Knoll LJ. A Genome-Wide siRNA screen to identify host factors necessary for growth of the parasite Toxoplasma gondii. PLoS One. 2013;8:1–9. doi: 10.1371/journal.pone.0068129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hirsch A. The use of RNAi-based screens to identify host proteins involved in viral replication. Future Microbiol. 2010;5:303–311. doi: 10.2217/fmb.09.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zhu Y, Li H, Ding S, Wang Y. Autophagy inhibition promotes phagocytosis of macrophage and protects mice from methicillin-resistant Staphylococcus aureus pneumonia. J. Cell. Biochem. 2018;119:4808–4814. doi: 10.1002/jcb.26677. [DOI] [PubMed] [Google Scholar]
- 40.Schmidt SV, Nino-Castro AC, Schultze JL. Regulatory dendritic cells: There is more than just immune activation. Front. Immunol. 2012;3:1–17. doi: 10.3389/fimmu.2012.00274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Reddick LE, Alto NM. Bacteria fighting back: How pathogens target and subvert the host innate immune system. Mol. Cell. 2014;54:321–328. doi: 10.1016/j.molcel.2014.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Redpath S, Ghazal P, Gascoigne NRJ. Hijacking and exploitation of IL-10 by intracellular pathogens. Trends Microbiol. 2001;9:86–92. doi: 10.1016/S0966-842X(00)01919-3. [DOI] [PubMed] [Google Scholar]
- 43.Dodding MP, Mitter R, Humphries AC, Way M. A kinesin-1 binding motif in vaccinia virus that is widespread throughout the human genome. EMBO J. 2011;30:4523–4538. doi: 10.1038/emboj.2011.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Henry T, Gorvel JP, Méresse S. Molecular motors hijacking by intracellular pathogens. Cell. Microbiol. 2006;8:23–32. doi: 10.1111/j.1462-5822.2005.00649.x. [DOI] [PubMed] [Google Scholar]
- 45.Greber UF, Way M. A superhighway to virus infection. Cell. 2006;124:741–754. doi: 10.1016/j.cell.2006.02.018. [DOI] [PubMed] [Google Scholar]
- 46.Zhou J, Scherer J, Yi J, Vallee RB. Role of kinesins in directed adenovirus transport and cytoplasmic exploration. PLoS Pathog. 2018;14:1–22. doi: 10.1371/journal.ppat.1007055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Boucrot E, Henry T, Borg J-P, Gorvel J-P, Méresse S. The intracellular fate of Salmonella depends on the recruitment of kinesin. Sci. (New York, NY) 2005;308:1174–1178. doi: 10.1126/science.1110225. [DOI] [PubMed] [Google Scholar]
- 48.Choe JE, Welch MD. Actin-based motility of bacterial pathogens: mechanistic diversity and its impact on virulence. Pathog. Dis. 2016;74:ftw099. doi: 10.1093/femspd/ftw099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Haglund CM, Welch MD. Pathogens and polymers: Microbe-host interactions illuminate the cytoskeleton. J. Cell Biol. 2011;195:7–17. doi: 10.1083/jcb.201103148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wasylnka JA, et al. Role for myosin II in regulating positioning of Salmonella-containing vacuoles and intracellular replication. Infect. Immun. 2008;76:2722–2735. doi: 10.1128/IAI.00152-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Miserey-Lenkei S, et al. Rab and actomyosin-dependent fission of transport vesicles at the golgi complex. Nat. Cell Biol. 2010;12:645–654. doi: 10.1038/ncb2067. [DOI] [PubMed] [Google Scholar]
- 52.Mostowy S, et al. Entrapment of intracytosolic bacteria by septin cage-like structures. Cell Host Microbe. 2010;8:433–444. doi: 10.1016/j.chom.2010.10.009. [DOI] [PubMed] [Google Scholar]
- 53.Hybiske K, Stephens RS. Mechanisms of host cell exit by the intracellular bacterium Chlamydia. Proc. Natl. Acad. Sci. 2007;104:11430–11435. doi: 10.1073/pnas.0703218104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Walter P. Travelling by TRAM. Nature. 1992;357:22–23. doi: 10.1038/357022a0. [DOI] [PubMed] [Google Scholar]
- 55.Celli J, Tsolis RM. Bacteria, the endoplasmic reticulum and the unfolded protein response: Friends or foes? Nat. Rev. Microbiol. 2015;13:71–82. doi: 10.1038/nrmicro3393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tilney LG, Harb OS, Connelly PS, Robinson CG, Roy CR. How the parasitic bacterium Legionella pneumophila modifies its phagosome and transforms it into rough ER: implications for conversion of plasma membrane to the ER membrane. J. Cell Sci. 2001;114:4637–4650. doi: 10.1242/jcs.114.24.4637. [DOI] [PubMed] [Google Scholar]
- 57.Isberg RR, O’Connor TJ, Heidtman M. The Legionella pneumophila replication vacuole: Making a cosy niche inside host cells. Nat. Rev. Microbiol. 2009;7:13–24. doi: 10.1038/nrmicro1967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Pizarro-cerda J, Moreno E, Sola-landa A, Lopez-gon I. Brucella abortus Transits through the autophagic pathway and replicates in the endoplasmic reticulum of nonprofessional phagocytes. Infect. Immun. 1998;66:5711–5724. doi: 10.1128/iai.66.12.5711-5724.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Celli J, et al. Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum. J. Exp. Med. 2003;198:545–556. doi: 10.1084/jem.20030088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Derré, I., Swiss, R. & Agaisse, H. The lipid transfer protein CERT interacts with the Chlamydia inclusion protein IncD and participates to ER-Chlamydia inclusion membrane contact sites. PLoS Pathog. 7 (2011). [DOI] [PMC free article] [PubMed]
- 61.Dumoux M, Clare DK, Saibil HR, Hayward RD. Chlamydiae assemble a pathogen synapse to hijack the host endoplasmic reticulum. Traffic. 2012;13:1612–1627. doi: 10.1111/tra.12002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Majeed M, Krause KH, Clark RA, Kihlström E, Stendahl O. Localization of intracellular Ca2+ stores in HeLa cells during infection with Chlamydia trachomatis. J. Cell Sci. 1999;112(Pt 1):35–44. doi: 10.1242/jcs.112.1.35. [DOI] [PubMed] [Google Scholar]
- 63.Giles DK, Wyrick PB. Trafficking of chlamydial antigens to the endoplasmic reticulum of infected epithelial cells. Microbes Infect. 2008;10:1494–1503. doi: 10.1016/j.micinf.2008.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mahalingam D, et al. Mipsagargin, a novel thapsigargin-based PSMA-Activated prodrug: Results of a first-in-man phase i clinical trial in patients with refractory, advanced or metastatic solid tumours. Br. J. Cancer. 2016;114:986–994. doi: 10.1038/bjc.2016.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Edgeworth JD, et al. An outbreak in an intensive care unit of a strain of Methicillin-Resistant Staphylococcus aureus Sequence Type 239 associated with an increased rate of vascular access device–related bacteremia. Clin. Infect. Dis. 2007;44:493–501. doi: 10.1086/511034. [DOI] [PubMed] [Google Scholar]
- 66.Edwards AM. Phenotype switching is a natural consequence of Staphylococcus aureus replication. J. Bacteriol. 2012;194:5404–5412. doi: 10.1128/JB.00948-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gawrzak S, et al. MSK1 regulates luminal cell differentiation and metastatic dormancy in ER+breast cancer. Nat. Cell Biol. 2018;20:211–221. doi: 10.1038/s41556-017-0021-z. [DOI] [PubMed] [Google Scholar]
- 68.Boada-Romero E, et al. TMEM59 defines a novel ATG16L1-binding motif that promotes local activation of LC3. EMBO J. 2013;32:566–582. doi: 10.1038/emboj.2013.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.