

Summary
Epithelia of the oral cavity exhibit variations in morphologies and turnover rates. Are these differences related to environment or to region-specific stem cell populations? A lineage-tracing strategy allowed visualization of Wnt-responsive cells, and their progeny, in the hard and soft palates. In both anatomic locations, Wnt-responsive basal cells self-renewed and gave rise to supra-basal cells. Palatal injuries triggered an enlargement of this population, and their descendants were responsible for wound re-epithelialization. Compared with the hard palate, soft palate stem cells exhibited an earlier, more robust burst in proliferation, culminating in significantly faster repair. Thereafter, excess Wnt-responsive basal cells were removed, and stem cell numbers were restored back to homeostatic level. Thus, we uncovered a stem cell population in oral mucosa, and its relative abundance is correlate with the rate of oral wound healing. Besides the activation during injury, an endogenous mechanism exists to constrain the stem cell pool after repair.
Subject Areas: Biological Sciences, Cell Biology, Stem Cells Research
Graphical Abstract
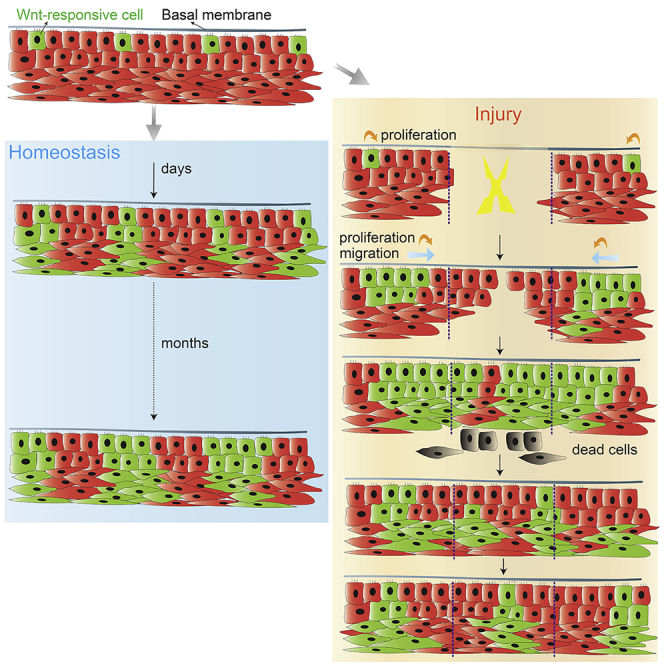
Highlights
-
•
During homeostasis, long-lived stem cells in the oral epithelium are Wnt responsive
-
•
After injury, the soft palate re-epithelizes faster than the hard palate
-
•
Faster healing is attributable to an abundance of Wnt-responsive cells
-
•
After healing, the Wnt-responsive stem cell pool returns to homeostatic levels
Biological Sciences; Cell Biology; Stem Cells Research
Introduction
The mucosal lining of the oral cavity serves as a barrier to protect the underlying tissues from mechanical damage and noxious stimuli. Like other covering and lining tissues of the body, the oral mucosa consists of a surface epithelial layer and a supporting fibrous connective tissue layer (Jones and Klein, 2013, Winning and Townsend, 2000). The connective tissue mainly provides mechanical support and nutrients for the epithelium, whereas the epithelium constantly turns over to protect the underlying connective tissue against mechanical and chemical insults (Squier and Kremer, 2001, Winning and Townsend, 2000).
In mammals, the oral mucosa can be divided into three categories based on function and histology: masticatory mucosa that includes the hard palate and gingiva; lining mucosa that covers the soft palate, the ventral surface of the tongue, as well as buccal, alveolar, and labial surfaces; and specialized mucosa on the dorsal surface of the tongue (Jones and Klein, 2013). Despite distinct functions and morphologies, all oral mucosa is contiguous. Whether they share the same stem cell population and similar remodeling mechanism is largely unknown.
The Wnt pathway is essential for stem cell maintenance and differentiation (Clevers et al., 2014, Kretzschmar and Clevers, 2017, Lien and Fuchs, 2014). The advent of lineage tracing techniques has made it possible to precisely tag populations that arise from Wnt-responsive cells and follow their fate(s) over time. Using Wnt reporters, Wnt responsiveness appears to be a general feature of stem cells in different tissues (Bowman et al., 2013, van Amerongen et al., 2012, Wang et al., 2015). In this study, we used two Wnt reporter strains to specifically visualize Wnt-responsive cells and examine their fates in the oral epithelia, here focusing on the hard and soft palates. Although these two tissues represent two different types of oral mucosa with distinct morphologies, we discovered that they shared a similar homeostatic mechanism. Upon injury, however, the hard and soft palates displayed distinctive, dynamic changes that led to significantly different healing rates. In addition to the critical role of Wnt-responsive cells in turnover and regeneration, we also demonstrated that in oral epithelia the number of Wnt-responsive basal cells is tightly controlled: instead of producing progeny to occupy all layers of the oral epithelia, a large portion of injury-induced progeny remain in the basal layer to cover the injury site; once the tissue has undergone repair, a subset of those labeled basal cells are removed via a non-apoptotic mechanism, which returns the tissue to its initial homeostatic state. These data have implications for improving the repair of slow-to-heal oral injuries.
Results
The Hard and Soft Palates Contain Wnt-Responsive Cells
The hard palate comprises the roof of the oral cavity (Figure 1A) and is characterized by a thick, keratinized epithelium with dense connective tissue underneath (Figure 1B), which makes it an ideal tissue to support mastication. Posteriorly, the hard palate is contiguous with the soft palate (Figure 1A), which has a thin epithelium and loose underlying connective tissue (Figure 1C). This flexible tissue is ideally suited for vocalization. Using an Axin2LacZ/+ Wnt reporter mouse strain, in which Xgal signal specifically reports Wnt activation in epithelia (Figure S1A), we found Xgal+ve, Wnt-responsive cells in both hard (Figure 1D) and soft palates (Figure 1E). A closer look at the epithelial layers revealed that majority of Xgal+ve Wnt-responsive cells were restricted to the basal layer (Figures 1D’ and 1E′). Basal cells were easy to be recognized by their position, size, and distinctive staining with nuclear fast red (Figure S1B). Quantification of basal cells co-stained with Xgal and nuclear fast red indicated that the soft palate harbored slightly more Wnt-responsive cells than the hard palate (Figure 1F).
Figure 1.

The Hard and Soft Palates Contain Wnt-Responsive Cells
(A–C) (A) Pentachrome staining of the maxilla (sagittal section) showing the different morphology of (B) the hard palate and (C) the soft palate. In pentachrome staining, the epithelial layer stains red and the lamia proper stains yellow to blue.
(D–E) In 3-month-old Axin2LacZ/+ mice, Xgal staining was performed to detect Wnt-responsive cells in (D) the hard palate and (E) the soft palate. The box area in D and E were enlarged in (D′) and (E′). Orange dotted lines indicate the cell shape.
(F) Quantification of Xgal+ve basal cells in both hard and soft palates (n = 3, 5 sections per mouse).
(G) Co-staining of GFP+ve cells (tamoxifen was injected 1 day before harvest) with β-catenin (red). The dotted line indicates the basement membrane. Abbreviation: lp, lamia proper; bl, basal layer; hp, hard palate; sp, soft palate; k, keratinized layer. Scale bars: black, 100 μm; orange, 10 μm.
Data represent mean ± SD. See also Figure S1.
Another genetic approach to labeling Wnt-responsive cells, as well as their progeny, involves the use of a Axin2CreErt2/+; R26mTmG/+ strain, in which tamoxifen converts Wnt-responsive cells to permanently labeled GFP-expressing cells (van Amerongen et al., 2012).
Based on prior experience (Yuan et al., 2018a, Yuan et al., 2018b), a single dose of tamoxifen (5 mg/25 g body weight) resulted in efficient labeling of the palatal epithelium (Figure S2).
To validate that GFP+ve cells indicated a Wnt-responsive status, tamoxifen was delivered to 3-months-old mice and 1 day later, oral epithelium was analyzed for both GFP and β-catenin (Figure 1G). Co-expression of GFP in cells with evidence of nuclear localization of β-catenin demonstrated that GFP+ve cells were responsive to an endogenous Wnt signal.
Wnt-Responsive Basal Cells Are Stem Cells in Both Hard and Soft Palates
With these tools in hand, we next examined the distribution of Wnt-responsive cells and followed their fates in the hard and soft palates. Tamoxifen was again delivered to 3-month-old-mice (Figure 2A). Within 1 day, analyses of the epithelium of the hard palate showed that GFP+ve, Wnt-responsive cells were primarily anchored to the basement membrane (Figure 2B). By day 5, GFP+ve cell clusters contained both cuboidal basal cells and flattened suprabasal cells (Figure 2C). By day 30, GFP+ve cells formed clones across all layers of the epithelium of the hard palate (Figure 2D). Clones continued to expand and were maintained for at least 180 days (Figures 2E and 2F). In the soft palate, Wnt-responsive cells also represented a subpopulation in the basal layer (Figure 2G). Fate analyses strongly suggested that GFP+ve Wnt-responsive cells were also stem cells (Figures 2H–2K). In terms of absolute number, the soft palate had more Wnt-responsive basal cells than the hard palate (Figure 2L), but in terms of the rate at which these populations grew, they were comparable (Figure S3).
Figure 2.

Wnt-Responsive Basal Cells Are Stem Cells in the Hard and Soft Palates
(A) A schematic diagram showing the lineage tracing strategy. Mice were injected with one-dose tamoxifen at the age of 3 months. The Wnt-responsive cells were analyzed 1 day (D), 5D, 30D, 90D, and 180D later.
(B–F) Wnt-responsive cells (green) were co-stained with β4 integrin (red, basement membrane). In the hard palate, (B) 1D chasing showing Wnt-responsive cells close to the basement membrane. These cells (C) expanded within 5D and (D) have established the clone in 30D. (E) 90D later, the clone enlarged and (F) the clone size was maintained in a 180D chasing. In the soft palate.
(G-K) (G) Wnt-responsive cells were first found in the basal layer and (H) the clones were quickly formed within 5D. (I) By 30D, the clone expanded and was maintained through (J) 90D and (K) 180D chasing.
(L) Quantification of GFP+ve basal cells, defined as GFP+ve cells that colocalized with the β4 integrin, in both the hard and soft palates at indicated time points (n = 3, 5 sections for each mouse). Abbreviation: lp, lamia proper. Scale bars: 25 μm. Data represent mean ± SD. See also Figures S2 and S3.
To confirm that Wnt-responsive basal cells were stem cells, hard and soft palate epithelia were harvested, from which single cells were isolated and examined in vitro (Figure S4A). First, we confirmed that GFP+ve cells were basal epithelial cells, by co-immunostaining of GFP (Figure S4B) with keratin 5 (Figure S4C), keratin 14 (Figure S4D), and β4 integrin (Figure S4E). GFP-expressing basal epithelial cells were immune-negative for the suprabasal markers Involucrin (Figure S4F) and loricrin (Figure S4G). The incorporation of EdU suggested that a high percentage of GFP+ve basal cells were proliferative (Figures S4H–S4J). Next, we examined the self-renewing ability of the GFP+ve basal cells using a colony forming unit (CFU) assay. GFP+ve clones were evident after 14 days (Figures S4K and S4L). Taken together, these in vivo and in vitro data support the conclusion that Wnt-responsive basal cells are stem cells in oral epithelia.
The Soft Palate Displays a Superior Healing Ability Compared with the Hard Palate
A wound in the oral cavity heals faster— and with less scarring—than other epithelial wounds (Iglesias-Bartolome et al., 2018, Politis et al., 2016). The stages of cutaneous wound healing have been well documented (Aragona et al., 2017, Dekoninck and Blanpain, 2019), whereas the same stages of healing in the oral cavity have not. In addition, it is not known whether different types of oral mucosa have different healing abilities. Therefore, we created a 2-mm full-thickness injury at the junction between the hard and soft palates (Figures 3A and 3B). This type of injury allowed us to study healing rates in both tissues. In the soft palate, the wound was clinically healed by post-surgery day (PSD) 3 (Figure 3C). In the hard palate, the injury site was not fully healed until PSD14 (Figures 3D and 3E). Histological analyses verified differences in healing rates. On PSD1, injuries spanning the hard and soft palates had visible wound edges (Figure 3F). By PSD3, the soft palate side of the injury was fully re-epithelialized, but the palate portion of the injury remained open (Figure 3G). By PSD7, the hard palate portion of the injury was re-epithelialized (Figure 3H), but the rugae were not fully reestablished until PSD14 (Figure 3I). Together, these data demonstrated that both hard and soft palates undergo repair in response to injury, but the soft palate does so much faster (Figures 3J and 3K).
Figure 3.

The Soft Palate Displays Superior Healing Ability Than the Hard Palate
(A) Intact hard and soft palates.
(B–E) (B) A 2-mm full-thickness injury (black dashed circle) was created in the area between the hard and soft palates. The center of the injury was located in the cross point of the middle line and the line along with the posterior ends of third molars. Yellow dashed circle indicates the unhealed part. Mice at age of 3 months were subjected to the injury and then monitored for (C) 3 days, (D) 7 days, and (E) 14 days.
(F-I) Representative Masson trichrome staining showing the healing of hard and soft palates on. (F) PSD1, (G) PSD3, (H) PSD7, and (I) PSD14. The black brackets indicate the injury site. The orange lines indicate the unhealed injury.
(J) Quantification of wound area shown in (B)–(E) (n = 6).
(K) Calculated repair rate between PSD1 and PSD3 (n = 6). Abbreviation: m1, maxillary first molar; m2, maxillary second molar; m3, maxillary third molar; PSD, post-surgery day; in, injury; hp, hard palate; sp, soft palate. Data represent mean ± SD. Scale bars: 200 μm.
Injury in the Soft Palate Induces More Robust Wnt and Proliferation Response
To explore the differential healing mechanisms in these two tissues, we examined the wound edges from the hard (Figure 4A) and soft palates (Figure 4B), focusing on cells that became Wnt responsive as a result of injury using Axin2LacZ/+ mice. On PSD3 in the hard palate, Xgal+ve, Wnt-responsive cells were abundant, distributed throughout the epithelial layers, and located at a distance (∼500 μm) from the wound edge (Figures 4C and 4C′). This represented a departure from the intact state (Figure 4D). In the soft palate, injury also boosted the number of Wnt-responsive cells on PSD3 (Figure 4E), and they were observed in almost all epithelial layers (Figure 4E’; compare with the intact state, Figure 4F). More Wnt-responsive cells were produced in response to soft palate injury versus hard palate injury (compare Figure 4E’ with Figure 4C’).
Figure 4.

In the Early Stage, Injury Induces More Robust Wnt and Proliferation Response in the Soft Palate
(A–B) On PSD3, representative Masson trichrome staining showing the healing front in (A) the hard palate and (B) the soft palate.
(C-F) In Axin2LacZ/+ mice, Xgal staining was performed to detect Wnt-responsive cells in (C) the healing hard palate on PSD3 and (D) the intact hard palate. The orange box area in (C) has been enlarged in (C′). Xgal staining was performed to detect Wnt-responsive cells in (E) the healing soft palate on PSD3 and (F) the intact soft palate. The orange box area in (E) has been enlarged in (E′).
(G-I) EdU staining was used to examine cell proliferation in (G) the healing hard palate on PSD3 and (H) the intact hard palate. The orange box area in (G) has been enlarged in (G′). EdU staining was used to examine cell proliferation in (I) the healing soft palate on PSD3 and (J) the intact soft palate. The orange box area in (I) has been enlarged in (I′).
(K) Quantification of EdU+ve cells (n = 6, 5 sections of each mouse). Abbreviation: lp, lamia proper; in, injury; PSD, post-surgery day; hp, hard palate; sp, soft palate. Scale bars: 100 μm.
Data represent mean ± SD. See also Figure S4.
We next tested whether injury-induced Wnt-responsive cells contributed to tissue repair. On PSD3, the majority of basal cells in the hard palate were proliferating, as indicated by EdU incorporation (Figures 4G and 4G′). This represented a dramatic departure from the intact hard palate, where only ∼10% of basal cells were actively proliferating (Figure 4H). In the soft palate, injury also induced a massive cell proliferation (Figures 4I and 4I′). Similar to what we observed in the hard palate, this proliferative burst stood in contrast to the intact state in the soft palate (Figure 4J). The number of proliferating cells was quantified (Figure 4K). Compared with the intact state, injury induced ∼5 times more EdU+ve cells in the hard palate and ∼11 times more EdU+ve cells in the soft palate (Figure 4K). Collectively, palatal injury stimulated a vigorous Wnt response that was accompanied by a vigorous proliferative response.
Substantial Wnt-Responsive Cells Persist in the Hard Palate after Re-Epithelization
To correlate Wnt-responsive cells with tissue healing, we examined injuries between PSD7 and PSD28. PSD7 was chosen because in the hard palate, the wound was re-epithelialized but not fully regenerated (Figure 5A). In contrast, by PSD7 in the soft palate, the tissue's original architecture was fully restored (Figure 5B). In the hard palate, Wnt-responsive cells were still abundant (Figure 5C). Conversely in the soft palate, the number of Wnt-responsive cells had returned to baseline (Figure 5D).
Figure 5.

Substantial Wnt-Responsive Cells Persist in the Hard Palate During and Post-Injury Healing
(A–D) Representative Masson trichrome staining showing the healed (A) hard palate and (B) soft palate on PSD7. Xgal staining was performed to detect the Wnt-responsive cells in (C) the healed hard palate and (D) the healed soft palate on PSD7.
(E-H) Representative Masson trichrome staining showing the healed (E) hard palate and (F) soft palate on PSD14. Xgal staining was performed to detect Wnt-responsive cells in (G) the healed hard palate and (H) the healed soft palate on PSD14.
(I) Xgal staining showing Wnt-responsive cells in the healed hard palate on PSD28.
(J) Quantification of Xgal+ve cells in the hard and soft palates at indicated time points (n = 6, 5 sections of each mouse). Abbreviation: lp, lamia proper; PSD, post-surgery day. Scale bars: 100 μm.
Data represent mean ± SD. See also Figure S4.
By PSD14 in the hard palate, the epithelium was thinner (Figure 5E) and rugae were present. The soft palate remained unchanged (Figure 5F). In the hard palate, the population of Wnt-responsive cells remained high in the repaired tissue (Figure 5G). In the soft palate, the number and distribution of Wnt-responsive cells were indistinguishable from that in the intact state (Figure 5H). On PSD28, the number and distribution of Xgal+ve Wnt-responsive cells in the hard palate finally returned to levels seen in the intact state (Figure 5I).
We quantified these changes in the number of Xgal+ve cells over time, and this value was normalized to total basal cell number using nuclear fast red co-staining (Figure S5). In both tissues, Wnt-responsive cells initially escalated in number, followed by their decline over time as the tissue healed. The primary difference between the hard and soft palates was in the magnitude of the response (Figure 5J). In the hard palate, the injury-induced expansion of Wnt-responsive cells was milder and more durable, whereas in the soft palate the increase in Wnt-responsive cells was considerable but transient (Figure 5J). Precisely how this expansion and contraction in Wnt-responsive cells was achieved became the focus of our next experiments.
The Wnt-Responsive Population Enlarges in Response to Injury but Prunes after Homeostasis Is Reestablished
The expansion in Wnt-responsive cells after injury could have been achieved via recruitment of cells. Alternatively, this could reflect the activation of stem cells. To distinguish between these possibilities, we returned to Axin2CreErt2/+; R26mTmG/+ strain. Our control group consisted of animals that received tamoxifen but did not sustain any injuries (Figure S6). Our test group consisted of reporter mice that received tamoxifen 2 days before palatal injury (Figure 6A). Cells that are responsive to an endogenous Wnt signal and simultaneously exposed to tamoxifen will stop expressing membrane TdTomato and start expressing membrane GFP (Figure S7). Tamoxifen stops label basal cells 1 day after injection (Figure S2F); therefore, we hypothesized that delivering tamoxifen 2 days before injury would only label resting Wnt-responsive cells in the basal layer and specifically avoid labeling cells that became Wnt-responsive as a result of the tissue damage. Analyses conducted immediately after injury confirmed that the labeling strategy worked: only small clones of GFP+ve cells were in evidence (Figures S6A and S6B), equivalent to the control (intact) group (Figures S6C and S6D).
Figure 6.

Wnt-Responsive Population Enlarges in Response to Proliferation but Prunes after Homeostasis Is Reestablished
(A) A schematic diagram showing the lineage tracing strategy. Mice were injected with one dose of tamoxifen at the age of 3 months. Two days later, a 2-mm full-thickness injury was generated in the hard and soft palates. Wnt-responsive cells were analyzed 3D, 7D, 10D, 14D, and 21D after surgery.
(B-C) On PSD3, GFP+ve Wnt-responsive cells were examined in the healing (B) hard palate and (C) soft palate.
(D-E) On PSD7, GFP+ve Wnt-responsive cells were examined in the healed (D) hard palate and (E) soft palate.
(F) On PSD14, abundant GFP+ve Wnt-responsive cells still existed in the healed hard palate.
(G) By PSD14, GFP+ve Wnt-responsive cells were returned to normal level.
(H) On PSD10, TUNEL were co-stained with GFP+ve Wnt-responsive cells in the soft palate.
(I) EdU staining showing the proliferation of cells in both hard and soft palates on PSD14. Orange bracket indicates the injury site.
(J) On PSD21, TUNEL were co-stained with GFP+ve Wnt-responsive cells in the hard palate.
(K) GFP+ve Wnt-responsive cells in the hard palate 28D after injury. Wnt-responsive cells (green) were co-immunostained with β4 integrin (β4, red, basement membrane). Yellow arrows indicate Wnt-responsive cells in suprabasal layers. White arrowhead indicates Wnt-responsive cells in basal layer. Abbreviation: lp, lamia proper; hp, hard palate; sp, soft palate; PSD, post-surgery day. Scale bars: 100 μm.
See also Figures S5–S9.
On PSD3, the GFP+ve population was clustered close to the edge of the injury (Figures S6E and S6F). Compared with the distribution of GFP+ve cells in the intact group (Figures S6G and S6H), there was a dramatic increase in GFP+ve population in injury group (Figures 6B and 6C). Besides the difference in number, suprabasal GFP+ve cells lack the continuity with basal GFP+ve cells during injury, indicating the injury-induced cell migration.
On PSD7, re-epithelialization of the wound was complete and, in this newly formed tissue, progeny of the initial Wnt-responsive stem population had re-established their position in the basal layer and had given rise to much larger clones (compare Figures 6D and 6E with the intact state, Figures S6I and S6J). Given the timing of the labeling (before injury), we conclude that the regenerated tissue arose from the progeny of Wnt-responsive cells residing in the basal layer of the intact palatal epithelia.
In most respects, the hard and soft palates responded similarly to injury, and in both anatomical locations, the Wnt-responsive populations behaved equivalently. What differed significantly, however, was the timing of events: in the hard palate, Wnt-responsive descendants persisted until PSD14 (Figure 6F), but in the soft palate, the GFP+ve population had declined by nearly 90% by PSD14 (Figure 6G, quantified in Figure S9) to its intact stage (Figure S6L). These data demonstrated that a massive pruning event was underway.
Although the reduction in GFP+ve epithelial clone size could be attributed to a higher level of turnover in the regenerated tissue, this would not explain the drastic reduction in GFP+ve cells in the basal layer (from 85% to 17%, Figures 6E and 6G, quantified in Figure S9). We wondered if the GFP+ve basal cells were undergoing apoptosis to re-establish homeostasis. Staining on PSD10 revealed only sporadic TUNEL+ve cells, and none of them were in the basal layer (Figure 6H) as they were observed in the intact state (Figure S8). Thus, the pruning of Wnt-responsive basal cells was apparently fulfilled by halting self-renewal.
The same sequence of events also occurred in the hard palate, just at later time points: for example, on PSD14, when abundant Wnt-responsive cells were still present in the hard palate (Figure 5G), and cell proliferation remained high (Figure 6I), indicating the tissue remodeling was ongoing. On PSD21, most GFP+ve cells were in suprabasal layers (Figure 6J) and homeostasis was ultimately re-established on PSD28 (Figure 6K, compared with Figure S6M).
Taken together, these data demonstrated that a stem population of GFP+ve Wnt-responsive cells residing in the basal layer dramatically expanded in response to injury. Part of the response was to re-establish a stem cell pool in the new basal layer overlying the wound, and another part of the response was to then directly differentiate and regenerate the missing tissue. Thereafter, through mechanisms that do not appear to involve apoptosis, the stem cell pool was pruned back to the level seen during homeostasis.
Discussion
Here, we report that Wnt-responsive basal cells in the hard and soft palates have a self-renewing capacity, give rise to differentiated suprabasal cells, and participate in tissue homeostasis and tissue regeneration. The hard and soft palates share similar homeostatic mechanisms and comparable injury responses; the main difference is that Wnt-responsive cells in the soft palate are more mitotically active, which correlates with a superior healing outcome.
To put these behaviors of oral epithelial stem cells in context, we considered other types of epithelial tissue. For several decades, researchers have characterized stem cells and described their self-renewing and differentiation capacities in skin. Multiple stem cell populations have been identified that contribute to both homeostasis and injury repair (Dekoninck and Blanpain, 2019). In comparison with skin, relatively little is known about stem cell populations in the oral mucosa. Mitotically active, long-lived BMI1+ve oral mucosal progenitor cells in the basal layer of the lining mucosa participate in homeostasis (Jones et al., 2019). Selective cell ablation leads to expansion of BMI1+ve clones, but their overall contribution to injury repair remains elusive (Jones et al., 2019). In our study, the Axin2+ve population in the basal layer of the oral mucosa is also long-lived. This Axin2+ve population maintains oral mucosa homeostasis (Figure 2), is activated by injury, and directly contributes to the regenerated tissue (Figures 4, 5, and 6). These similarities suggest that the two populations may overlap. In fact, several studies have linked BMI1 activity with Wnt signaling. BMI encodes a ring finger protein in the polycomb group complex that controls chromatin remodeling (Cho et al., 2013, Li et al., 2018), and at least one study shows that BMI1 promotes Wnt signaling in epithelial cells and cancer cells, perhaps through repressing the Wnt antagonist, Dickkopf (Cho et al., 2013, Li et al., 2018). Activating Wnt signaling via over-expression of Wnt1 promotes BMI1 expression as well (Cho et al., 2013). In ongoing studies, we are exploring the extent to which the Axin2+ve Wnt-responsive population of stem cells overlaps with the BMI1+ve population.
In our study, we also noticed that the number of Wnt-responsive basal cells is tightly controlled. During homeostasis, Wnt-responsive basal cells increased in a very slow rate (Figure 2). In response to injury, however, abundant new cells are required to quickly close a wound. We demonstrated that, during the repair process, Wnt-responsive cells produced ample progeny (Figure 6). These cells initially occupied a large percentage of the basal layer in the newly formed oral epithelia, but this state was transient. Shortly after re-epithelialization was complete, the percentage of Wnt-responsive basal cells was reduced (Figure 5) by halting their self-renewing capacity (Figure 6). We speculate that this “pruning” process represents a potential cancer-preventing mechanism based on the undisputed link between chronic injury and oncogenic transformation of the oral mucosa. We demonstrated that a stem cell population exists in the basal layer (Figure 2) and this stem cell population dramatically expands in response to injury (Figure 4). Most cancers arise from the accumulation of DNA mutations due to infidelity of the DNA replication and failure of DNA repair (Behrens et al., 2014, Vassilev and DePamphilis, 2017) and the risk of a stem cell undergoing oncogenic transformation is strongly correlated with the total number of divisions it undergoes (Tomasetti and Vogelstein, 2015). Statistically, the chance that a random mutation arises during the rapid proliferation process in our model is high. If cells with such DNA mutations are not eliminated, they can initiate carcinogenesis (Batlle and Clevers, 2017, Lin et al., 2008). Currently, it is unclear how the body selects Wnt-responsive cells to stay in the basal layers as well as what the pruning mechanism is.
In our study, we examined the healing in both the hard and soft palate. Our data demonstrated that Wnt-responsive cells in the soft palate response to the injury more quickly (Figures 4 and 5). The hard palate and the soft palate are distinctive in their morphology (Figure 1) and, therefore, cells in these two tissues are in different mechanical environments. For example, the hard palate is exposed to compressive and shear forces during the mastication of food (Nanci, 2017) and the junction between its epithelium and underlying connective tissue is convoluted, to prevent the epithelium from detaching under shear force. The soft palate, on the other hand, is flexible and has a loose layer of elastic fibers underneath (Nanci, 2017). Tissue damage alters the mechanical environment of cells (Begnaud et al., 2016, Ladoux and Mege, 2017, Rousselle et al., 2018), and stem cells translate a change in the mechanical environment into molecular signals that activate cellular functions related to tissue repair. For example, changes in mechanical forces stimulate stem cell proliferation (Elbediwy et al., 2016, Vining and Mooney, 2017). We hypothesize that the robust cell proliferation that characterizes soft palate healing (Figure 4) may be activated by wound contracture and changes in extracellular matrix stiffness as part of the repair process (Li et al., 2014). One molecular pathway that is implicated in regulating stem cell function is the Hippo pathway, which integrates cytoskeletal changes in stem cells with alterations in the extracellular environment (Varelas, 2014, Zhao et al., 2011) during both tissue homeostasis and repair (Elbediwy et al., 2016). Whether TAZ/YAP also serves as a mechano-sensor/damage-sensor in oral mucosa is a focus of future studies.
Limitations of the Study
Although this study provided insights into the stem cell function of Wnt-responsive cells in oral epithelia, subsequent studies are necessary to demonstrate whether these cells are indispensable to maintain tissue integrity and support repair. One approach may be to ablate Axin2+ve cells in epithelia, then study how the homeostasis and repair response is affected. It is also worth noting that we did not investigate the source of Wnt, neither during homeostasis nor during injury repair. Identifying the Wnt-secreting cells might help to understand the mechanism of the “trimming process” we observed after repair.
Methods
All methods can be found in the accompanying Transparent Methods supplemental file.
Acknowledgment
We thank Christina Mac, Ryan Ma, and Peter Chew for their assistance in cell culture, tissue processing, and staining. The study was funded by NIH grant R01DE024000-14 to J.A.H. and K99DE028585-01 to X.Y.
Author Contributions
Conceptualization, X.Y. and J.A.H.; Methodology, X.Y., Q.X, X.Z, and J.A.H.; Investigation, X.Y., Q.X, X.Z, L.A.v.B., P.T., and J.A.H.; Writing – Original Draft, X.Y. and J.A.H.; Writing – Review & Editing X.Y., L.A.v.B., P.T., and J.A.H.; Funding Acquisition, X.Y. and J.A.H.; Resources, J.A.H.; Supervision, X.Y. and J.A.H.
Declaration of Interests
J.A.H. is an employee of Ankasa Regenerative Therapeutics, a biopharmaceutical company developing L-WNT3A for commercial use. The authors declare no competing financial interests.
Published: November 22, 2019
Footnotes
Supplemental Information can be found online at https://doi.org/10.1016/j.isci.2019.10.016.
Contributor Information
Xue Yuan, Email: xueyuan@stanford.edu.
Jill A. Helms, Email: jhelms@stanford.edu.
Supplemental Information
References
- Aragona M., Dekoninck S., Rulands S., Lenglez S., Mascre G., Simons B.D., Blanpain C. Defining stem cell dynamics and migration during wound healing in mouse skin epidermis. Nat. Commun. 2017;8:14684. doi: 10.1038/ncomms14684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Batlle E., Clevers H. Cancer stem cells revisited. Nat. Med. 2017;23:1124–1134. doi: 10.1038/nm.4409. [DOI] [PubMed] [Google Scholar]
- Begnaud S., Chen T., Delacour D., Mege R.M., Ladoux B. Mechanics of epithelial tissues during gap closure. Curr. Opin. Cell Biol. 2016;42:52–62. doi: 10.1016/j.ceb.2016.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens A., van Deursen J.M., Rudolph K.L., Schumacher B. Impact of genomic damage and ageing on stem cell function. Nat. Cell Biol. 2014;16:201–207. doi: 10.1038/ncb2928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowman A.N., van Amerongen R., Palmer T.D., Nusse R. Lineage tracing with Axin2 reveals distinct developmental and adult populations of Wnt/beta-catenin-responsive neural stem cells. Proc. Natl. Acad. Sci. U S A. 2013;110:7324–7329. doi: 10.1073/pnas.1305411110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho J.H., Dimri M., Dimri G.P. A positive feedback loop regulates the expression of polycomb group protein BMI1 via WNT signaling pathway. J. Biol. Chem. 2013;288:3406–3418. doi: 10.1074/jbc.M112.422931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clevers H., Loh K.M., Nusse R. Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science. 2014;346:1248012. doi: 10.1126/science.1248012. [DOI] [PubMed] [Google Scholar]
- Dekoninck S., Blanpain C. Stem cell dynamics, migration and plasticity during wound healing. Nat. Cell Biol. 2019;21:18–24. doi: 10.1038/s41556-018-0237-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elbediwy A., Vincent-Mistiaen Z.I., Thompson B.J. YAP and TAZ in epithelial stem cells: a sensor for cell polarity, mechanical forces and tissue damage. Bioessays. 2016;38:644–653. doi: 10.1002/bies.201600037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iglesias-Bartolome R., Uchiyama A., Molinolo A.A., Abusleme L., Brooks S.R., Callejas-Valera J.L., Edwards D., Doci C., Asselin-Labat M.L., Onaitis M.W. Transcriptional signature primes human oral mucosa for rapid wound healing. Sci. Transl. Med. 2018;10 doi: 10.1126/scitranslmed.aap8798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones K.B., Furukawa S., Marangoni P., Ma H., Pinkard H., D'Urso R., Zilionis R., Klein A.M., Klein O.D. Quantitative clonal analysis and single-cell transcriptomics reveal division kinetics, hierarchy, and fate of oral epithelial progenitor cells. Cell Stem Cell. 2019;24:183–192.e8. doi: 10.1016/j.stem.2018.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones K.B., Klein O.D. Oral epithelial stem cells in tissue maintenance and disease: the first steps in a long journey. Int. J. Oral Sci. 2013;5:121–129. doi: 10.1038/ijos.2013.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kretzschmar K., Clevers H. Wnt/beta-catenin signaling in adult mammalian epithelial stem cells. Dev. Biol. 2017;428:273–282. doi: 10.1016/j.ydbio.2017.05.015. [DOI] [PubMed] [Google Scholar]
- Ladoux B., Mege R.M. Mechanobiology of collective cell behaviours. Nat. Rev. Mol. Cell Biol. 2017;18:743–757. doi: 10.1038/nrm.2017.98. [DOI] [PubMed] [Google Scholar]
- Li J., Johnson C.A., Smith A.A., Shi B., Brunski J.B., Helms J.A. Molecular mechanisms underlying skeletal growth arrest by cutaneous scarring. Bone. 2014;66:223–231. doi: 10.1016/j.bone.2014.06.007. [DOI] [PubMed] [Google Scholar]
- Li X.G., Wang Z., Chen R.Q., Fu H.L., Gao C.Q., Yan H.C., Xing G.X., Wang X.Q. LGR5 and BMI1 increase pig intestinal epithelial cell proliferation by stimulating WNT/beta-Catenin signaling. Int. J. Mol. Sci. 2018;19(4):1036. doi: 10.3390/ijms19041036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lien W.H., Fuchs E. Wnt some lose some: transcriptional governance of stem cells by Wnt/beta-catenin signaling. Genes Dev. 2014;28:1517–1532. doi: 10.1101/gad.244772.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin E.H., Jiang Y., Deng Y., Lapsiwala R., Lin T., Blau C.A. Cancer stem cells, endothelial progenitors, and mesenchymal stem cells: “seed and soil” theory revisited. Gastrointest. Cancer Res. 2008;2:169–174. [PMC free article] [PubMed] [Google Scholar]
- Nanci A. Elsevier Health Sciences; 2017. Ten Cate's Oral Histology-E-Book: Development, Structure, and Function. [Google Scholar]
- Politis C., Schoenaers J., Jacobs R., Agbaje J.O. Wound healing problems in the mouth. Front. Physiol. 2016;7:507. doi: 10.3389/fphys.2016.00507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rousselle P., Braye F., Dayan G. Re-epithelialization of adult skin wounds: cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2018 doi: 10.1016/j.addr.2018.06.019. in press. [DOI] [PubMed] [Google Scholar]
- Squier C.A., Kremer M.J. Biology of oral mucosa and esophagus. J. Natl. Cancer Inst. Monogr. 2001:7–15. doi: 10.1093/oxfordjournals.jncimonographs.a003443. [DOI] [PubMed] [Google Scholar]
- Tomasetti C., Vogelstein B. Variation in cancer risk among tissues can be explained by the number of stem cell divisions. Science. 2015;347:78–81. doi: 10.1126/science.1260825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Amerongen R., Bowman A.N., Nusse R. Developmental stage and time dictate the fate of Wnt/beta-catenin-responsive stem cells in the mammary gland. Cell Stem Cell. 2012;11:387–400. doi: 10.1016/j.stem.2012.05.023. [DOI] [PubMed] [Google Scholar]
- Varelas X. The Hippo pathway effectors TAZ and YAP in development, homeostasis and disease. Development. 2014;141:1614–1626. doi: 10.1242/dev.102376. [DOI] [PubMed] [Google Scholar]
- Vassilev A., DePamphilis M.L. Links between DNA replication, stem cells and cancer. Genes. 2017;8:45. doi: 10.3390/genes8020045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vining K.H., Mooney D.J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 2017;18:728–742. doi: 10.1038/nrm.2017.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang B., Zhao L., Fish M., Logan C.Y., Nusse R. Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of the liver. Nature. 2015;524:180–185. doi: 10.1038/nature14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winning T.A., Townsend G.C. Oral mucosal embryology and histology. Clin. Dermatol. 2000;18:499–511. doi: 10.1016/s0738-081x(00)00140-1. [DOI] [PubMed] [Google Scholar]
- Yuan X., Pei X., Zhao Y., Li Z., Chen C.H., Tulu U.S., Liu B., Van Brunt L.A., Brunski J.B., Helms J.A. Biomechanics of immediate postextraction implant osseointegration. J. Dent Res. 2018;97:987–994. doi: 10.1177/0022034518765757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan X., Pei X., Zhao Y., Tulu U.S., Liu B., Helms J.A. A Wnt-responsive PDL population effectuates extraction socket healing. J. Dent Res. 2018;97:803–809. doi: 10.1177/0022034518755719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao B., Tumaneng K., Guan K.L. The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat. Cell Biol. 2011;13:877–883. doi: 10.1038/ncb2303. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.