

Abstract
By sequencing 523 ancient humans, we show that the primary source of ancestry in modern South Asians is a prehistoric genetic gradient between people related to early hunter-gatherers of Iran and southeast Asia. Following the Indus Valley Civilization’s decline, they mixed with people in the southeast to form one of the two main ancestral populations of South Asia whose direct descendants live in southern India. Simultaneously, they mixed with descendants of Steppe pastoralists who spread via Central Asia after 4000 years ago to form the other main ancestral population. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages.
Graphical Abstract
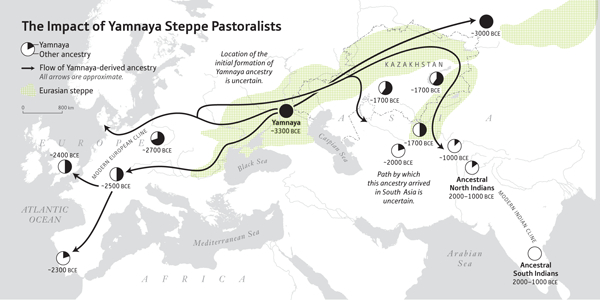
The Bronze Age spread of Yamnaya steppe pastoralist ancestry into two subcontinents, Europe and South Asia. Pie charts reflect the proportion of Yamnaya ancestry, and dates reflect the earliest available ancient DNA with Yamnaya ancestry in each region. There is no ancient DNA yet for the ANI and ASI, so for these the range is inferred statistically.
One Sentence Summary:
Genome wide ancient DNA from 523 ancient individuals sheds light on genetic exchanges between the Steppe, Iran and South Asia, and highlights the parallel demographic histories of two subcontinents: Europe and South Asia.
One Page Summary
Introduction and Rationale:
To elucidate the extent to which the major cultural transformations of farming, pastoralism and shifts in the distribution of languages in Eurasia were accompanied by movement of people, we report genome-wide ancient DNA data from 523 individuals spanning the last 8000 years mostly from Central Asia and northernmost South Asia.
Results:
Movements of people following the advent of farming resulted in genetic gradients across Eurasia that can be modeled as mixtures of seven deeply divergent populations. A key gradient formed in southwestern Asia beginning in the Neolithic and continuing into the Bronze Age, with more Anatolian farmer-related ancestry in the west and more Iranian farmer-related ancestry in the east. This cline extended to the desert oases of Central Asia and was the primary source of ancestry in peoples of the Bronze Age Bactria Margiana Archaeological Complex (BMAC). This supports the idea that the archaeologically documented dispersal of domesticates was accompanied by the spread of people from multiple centers of domestication.
The main population of the BMAC carried no ancestry from Steppe pastoralists and did not contribute substantially to later South Asians. However, Steppe pastoralist ancestry appeared in outlier individuals at BMAC sites by the turn of the second millennium BCE around the same time as it appeared on the southern Steppe. Using data from ancient individuals from the Swat Valley of northernmost South Asia, we show that Steppe ancestry then integrated further south in the first half of the second millennium BCE, contributing up to 30% of the ancestry of modern groups in South Asia. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages.
The primary ancestral population of modern South Asians is a mixture of people related to early Holocene populations of Iran and South Asia that we detect in outlier individuals from two sites in cultural contact with the Indus Valley Civilization (IVC), making it plausible that it was characteristic of the IVC. After the IVC’s decline, this population mixed with northwestern groups with Steppe ancestry to form the “Ancestral North Indians” (ANI) and with southeastern groups to form the “Ancestral South Indians” (ASI) whose direct descendants live today in tribal groups in southern India. Mixtures of these two post-IVC groups--the ANI and ASI--drive the main gradient of genetic variation in South Asia today.
Conclusion:
Earlier work recorded massive population movement from the Steppe into Europe early in the 3rd millennium BCE, likely spreading Indo-European languages. We reveal a parallel series of events leading to the spread of Steppe ancestry to South Asia, thereby documenting movements of people that were likely conduits for the spread of Indo-European languages.
Introduction
The past ten thousand years have witnessed profound economic changes driven by the transition from foraging to food production, and have also witnessed dramatic changes in cultural practice evident from archaeology, the distribution of languages, and the written record. The extent to which these changes were associated with movements of people has been a mystery in Central Asia and South Asia in part because of a paucity of ancient DNA. We report genome-wide data from 523 individuals from Central Asia and northernmost South Asia from the Mesolithic period onward (1), and co-analyze them with previously published ancient DNA from across Eurasia and with data from diverse present-day people.
In Central Asia, we studied the extent to which the spread of farming and herding practices from the Iranian plateau to the desert oases south of the Steppe was accompanied by movements of people or adoption of ideas from neighboring groups (2–4). For the urban communities of the Bactria Margiana Archaeological Complex (BMAC) in the Bronze Age, we assessed whether the people buried in its cemeteries descended directly from earlier smaller scale food producers, and also documented their genetic heterogeneity (5). Further to the north and east, we showed that the Early Bronze Age spreads of crops and domesticated animals between southwest Asia and eastern Eurasia along the Inner Asian Mountain Corridor (6) was accompanied by movements of people. Finally, we examined when descendants of the Yamnaya, who spread across the Eurasian Steppe beginning around 3300 BCE (7–9), began to appear in Central Asia south of the Steppe.
In northernmost South Asia, we report a time transect of more than one hundred individuals beginning ~1200 BCE, which we co-analyze along with modern data from hundreds of present-day South Asian groups, as well as ancient DNA from neighboring regions (10). Previous analyses place the majority of present-day South Asians along a genetic cline (11) that can be modeled as having arisen from mixture of two highly divergent populations after 4000 years ago: the Ancestral North Indians (ANI) who harbor large proportions of ancestry related to West Eurasians, and the Ancestral South Indians (ASI) who are much less closely related to West Eurasians (12). We leveraged ancient DNA to place constraints on the genetic structure of the ANI and ASI and, in conjunction with other lines of evidence, to make inferences about when and where they formed. By modeling modern South Asians along with ancient individuals from sites in cultural contact with the IVC, we inferred a likely genetic signature for people of the Indus Valley Civilization (IVC) which reached its maturity in northwestern South Asia 2600–1900 CE. We also examined when Steppe pastoralist-derived ancestry (9) mixed with groups in South Asia, and placed constraints on whether Steppe-related ancestry or Iranian-related ancestry is more plausibly associated with the spread of Indo-European languages in South Asia.
Dataset and Analysis Strategy
We generated whole-genome ancient DNA data from 523 previously unsampled ancient individuals and increased the quality of data from 19 previously sequenced individuals. The individuals derive from three broad geographical regions: 182 from Iran and the southern part of Central Asia that we call Turan (present-day Turkmenistan, Uzbekistan, Tajikistan, Afghanistan and Kyrgyzstan), 209 from the Steppe and northern forest zone mostly in present-day Kazakhstan and Russia, and 132 from northern Pakistan. The ancient individuals are from 1) Mesolithic, Copper, Bronze and Iron Age Iran and Turan (12000–1 BCE from 19 sites) including the Bactria Margiana Archaeological Complex (BMAC); 2) early ceramic-using hunter-gatherers from the western Siberian forest zone who we show represent a point along an early Holocene cline of North Eurasians and who emerge as a valuable source population for modeling the ancestry of Central and South Asians (6400–3900 BCE from 2 sites); 3) Copper Age and Bronze Age pastoralists from the central Steppe, including from Bronze Age Kazakhstan (3400–800 BCE from 56 sites); and 4) northernmost South Asia, specifically Late Bronze Age, Iron Age and historical settlements in the Swat and Chitral districts of present-day Pakistan (~1200 BCE - 1700 CE from 12 sites) (Fig. 1, Table S1, (1, 13)). We prepared samples in dedicated clean rooms, extracted DNA (14, 15), and constructed libraries for Illumina sequencing (16, 17). We enriched the libraries for DNA overlapping around 1.2 million single nucleotide polymorphisms (SNPs) (7, 18, 19), sequenced the products on Illumina instruments, and performed quality control (Table S2) (7, 19, 20). Our final dataset after merging with previously reported data (7–9, 16, 18, 19, 21–31) spans 837 ancient individuals that passed all our analysis filters, which included removing individuals determined genetically to be first-degree relatives of other higher coverage individuals (Table S3), and restricting to the 92% of individuals (Table S1) that were represented by at least 15,000 of the targeted SNPs which we found was the number at which we began to be able to reliably estimate proportions of the deeply divergent ancestry sources. The median number of SNPs analyzed per individual was 617,000. We also merged with previously reported whole genome sequencing data from 686 present-day individuals (Table S1), and co-analyzed with 1,789 present-day people from 246 ethnographically-distinct groups in South Asia genotyped at ~600,000 SNPs (Table S5; (13)) (10, 32, 33).
Fig. 1. Overview of ancient DNA data.

(A) Distribution of sites and associated archeological or radiocarbon dates along with the number of individuals meeting our analysis thresholds from each site. (B) Locations of ancient individuals for whom we generated ancient DNA that passed our analysis thresholds along with the locations of individuals from 140 groups from present-day South Asia that we analyzed as forming the Modern Indian Cline. Shapes distinguish the individuals from different sites. Data from 106 South Asian groups that do not fit along the Modern Indian Cline as well as AHG are not shown. (C) PCA analysis of ancient and modern individuals projected onto a basis formed by 1,340 present day Eurasians reflects clustering of individuals that mirrors their geographical relationships. An interactive version of this figure is presented in the Online Data Visualizer.
We grouped individuals based on archaeological and chronological information, taking advantage of 269 direct radiocarbon dates generated on skeletal material from the individuals from whom we report DNA (Table S4). We further clustered individuals that were genetically indistinguishable within these groupings, and labeled outliers with ancestry that was significantly different from others at the same site and time period (13). For our primary analyses, we did not include individuals that were the sole representatives of their ancestry profiles, thereby reducing the chance that our conclusions were being driven by single individuals with contaminated DNA or misattributed archaeological context. This also ensured that each major analysis grouping was represented by many more SNPs that our minimum cutoff of 15,000 per individual. Thus, all but one analysis cluster included at least one individual covered by >200,000 SNPs, sufficient to support high resolution analysis of population history (19) (the exception is a pair of genetically similar outliers from the site of Gonur who are not the focus of any main analyses). We use Italic font to refer to genetic groupings and plain font to indicate archaeological cultures or sites.
To make inferences about population structure, we began by carrying out principal component analysis (PCA) projecting ancient individuals onto the patterns of genetic variation in present-day Eurasians, a procedure that allowed us to obtain meaningful constraints on ancestry even of ancient individuals with limited coverage because each SNP from each individual can be compared to a large reference data set (34–36). This revealed three major clusters strongly correlating to the geographic regions of the Forest Zone/Steppe, Iran/Turan, and South Asia (Fig. 1), a pattern we replicate in ADMIXTURE unsupervised clustering (37). To test if groups of ancient individuals were heterogeneous in their ancestry, we used f4-statistics to measure whether different partitions of these groups into two subgroups differed in their degree of allele sharing to a third group (using a distantly related outgroup as a baseline). We also used f3-statistics to test for admixture (33). To model the ancestry of each group, we used qpAdm, which evaluates whether a tested group is consistent with deriving from a pre-specified number of source populations (relative to a set of outgroups), and if so estimates proportions of ancestry (7). We first used qpAdm to attempt to model groups from the Copper Age and afterward as a mixture of seven “distal” sources related deeply to pre-Copper Age or distantly related modern populations for which we have data (Box 1). In this paper we use the term ‘farmers’ to refer to people who either cultivated crops, or herded animals, or both; this definition covers not only large settled communities but also smaller and probably less sedentary communities like the early herders of the Zagros Mountains of western Iran from the site of Ganj Dareh. The latter kept domesticated animals but did not cultivate crops, and are a key reference population for this study as they had a distinctive ancestry profile that spread widely after the Neolithic (9, 24, 38). We also identified “proximal” models for each group as mixtures of temporally preceding groups (10). We implemented an algorithm, DATES, for estimating the age of population mixtures by measuring the average size of segments of ancestry derive from the admixing populations, an approach whose reliability we verified by computer simulation (10) and that is an improvement relative to methods not optimized for analysis of ancient DNA (33, 39) (the approach’s robustness reflects the fact that it relies for its molecular clock on the accurately measured rate of meiotic recombination in humans (40)). In Box 2, we summarize the findings of these analyses (we use the same headings in Box 2 and the main text to allow cross-referencing), while the Online Data Visualizer (1) allows an interactive exploration of the data.
Box 1. Seven Source Populations Used for Distal Ancestry Modeling.
| Anatolia_N | Anatolian farmer-related: Represented by 7th millennium BCE western Anatolian farmers (19) |
| Ganj_Dareh_N | Iranian early farmer-related: Represented by 8th millennium BCE early goat herders from the Zagros Mountains of Iran (9, 24) |
| WEHG | Western European Hunter-Gatherer-related: represented by 9th millennium BCE Western Europeans (7, 19, 32, 69). (WEHG and EEHG discussed below were denoted WHG and EHG in previous studies, but as we co-analyze them with hunter-gatherers from Asia we modify the names to specify a European origin.) |
| EEHG | Eastern European Hunter-Gatherer-related: represented by 6th millennium BCE hunter-gatherers from Eastern Europe (19, 32) |
| WSHG | West Siberian Hunter-Gatherer-related: a previously undescribed deep source of Eurasian ancestry represented in this study by three individuals from the Forest Zone of Central Russia dated to the 6th millennium BCE. |
| ESHG | East Siberian Hunter-Gather-related: represented by 6th millennium BCE hunter-gatherers from the Lake Baikal region with ancestry deeply related to East Asians (26) |
| AHG | Andamanese Hunter-Gatherer-related: represented by present day indigenous Andaman Islanders (55) who we hypothesize are related to unsampled indigenous South Asians (Ancient Ancestral South Indians - AASI) |
Box 2. Summary of Key Findings.
Iran and Turan
A West-to-East Cline of Decreasing Anatolian Farmer-Related Ancestry. There was a west-to-east gradient of ancestry across Eurasia in the Copper Age and Bronze Ages—the Southwest Asian Cline—with more Anatolian farmer-related ancestry in the west and more WSHG- or AASI-related ancestry in the east, superimposed on primary ancestry related to early Iranian farmers. The establishment of this gradient correlates in time to the spread of plant-based agriculture across this region, raising the possibility that people of Anatolian ancestry spread this technology east just as they helped spread it west into Europe.
People of the BMAC Were Not a Major Source of Ancestry for South Asians. The primary BMAC population largely derived from preceding local Copper Age peoples who were in turn closely related to people from the Iranian plateau, and had little of the Steppe ancestry that is ubiquitous in South Asia today.
Steppe Pastoralist-Derived Ancestry Arrived in Turan by 2100 BCE. We find no evidence of Steppe pastoralist-derived ancestry in groups at BMAC sites prior to 2100 BCE, but multiple outlier individuals buried at these sites show that by ~2100–1700 BCE, BMAC communities were regularly interacted with peoples carrying such ancestry.
An Ancestry Profile Widespread During the Indus Valley Civilization. We document a distinctive ancestry profile—45–82% Iranian farmer-related and 11–50% AASI (with negligible Anatolian farmer-related admixture)—that was present at two sites in cultural contact with the Indus Valley Culture (IVC). Combined with our detection of this same ancestry profile (in mixed form) about a millennium later in the post-IVC Swat Valley, this documents an Indus Periphery Cline during the flourishing of the IVC. Ancestors of this group formed by admixture ~5400–3700 BCE. There is little if any Anatolian farmer-related ancestry in the Indus Periphery Cline.
The Steppe and Forest Zone
Ancestry Clines in North Eurasia Established After the Advent of Farming. Prior to the spread of farmers and herders, northern Eurasia was characterized by a west-to-east gradient of very divergent hunter-gatherer populations with increasing proportions of relatedness to present-day East Asians: from Western European Hunter-Gatherers (WEHG), to Eastern European Hunter-Gatherers (EEHG), to West Siberian Hunter-Gatherers (WSHG), to Eastern Siberian Hunter-Gatherers (ESHG). Mixture of people along this ancestry gradient and its counterpart to the south formed five later clines following the advent of farming, the three northern ones of which are the European Cline, the Caucasus Cline, and the Central Asian Cline.
A Distinctive Ancestry Profile Stretching from Eastern Europe to Kazakhstan in the Bronze Age. We add more than one hundred samples from the previously described Western_Steppe_MLBA genetic cluster, including individuals associated with the Corded Ware, Srubnaya, Petrovka, and Sintashta archaeological complexes, and characterized by a mixture of about two-thirds ancestry related to Yamnaya Steppe pastoralists (from the Caucasus Cline), and European farmers (from the European Cline) suggesting that this population formed at the geographic interface of these two groups in Eastern Europe. Our analysis suggests that in the central Steppe and Minusinsk Basin in the Middle to Late Bronze Age, Western_Steppe_MLBA ancestry mixed with about 9% ancestry from previously established people from the region carrying WSHG-related to form a distinctive Central_Steppe_MLBA cluster that was the primary conduit for spreading Yamnaya Steppe pastoralist-derived ancestry to South Asia.
Bidirectional Mobility Along the Inner Asian Mountain Corridor. Beginning in the 3rd millennium BCE and intensifying in the 2nd millennium BCE, we observe multiple individuals in the Central Steppe who lived along the Inner Asian Mountain Corridor and who harbored admixture from Turan, documenting northward movement into the Steppe in this period. By the end of the 2nd millennium BCE, these people were later joined by numerous outlier individuals with East Asian-related admixture which become ubiquitous in the region by the Iron Age (25, 52). This ancestry is also seen in later groups with known cultural impacts on South Asia including Huns, Kushans and Sakas and is hardly present in the two primary ancestral populations of South Asia, suggesting that the Steppe ancestry widespread in South Asia derived from pre-Iron Age Central Asians.
South Asia
Three Ancestry Clines That Succeeded Each Other in Time in South Asia. We identify a unique trio of source populations that fits geographically and temporally diverse South Asians since the Bronze Age: a mixture of AASI, an Indus Periphery Cline group with predominantly Iranian farmer-related ancestry, and Central_Steppe_MLBA. Two-way clines driven more by populations that were mixtures of these three sources succeeded each other in time: prior to 2000 BCE the Indus Periphery Cline had no detectable Steppe ancestry, beginning after 2000 BCE the Steppe Cline, and finally the Modern Indian Cline.
The ASI and ANI Arose as Indus Periphery Cline People Mixed with Groups to the North and East. An ancestry gradient of which the Indus Periphery Cline individuals were a part played a pivotal role in the formation of both the two proximal sources of ancestry in South Asia: a minimum of ~55% Indus Periphery Cline ancestry for the ASI and ~70% for the ANI. Today there are groups in South Asia with very similar ancestry to the statistically reconstructed ASI suggesting that they have essentially direct descendants today. Much of the formation of both the ASI and ANI occurred in the 2nd millennium BCE. Thus, the events that formed both the ASI and ANI overlapped the time of the decline of the IVC.
Steppe Ancestry in South Asia is Primarily from Males and Disproportionately High in Brahmins. Most of the Steppe ancestry in South Asia derives from males, pointing to asymmetric social interaction between descendants of Steppe pastoralists and peoples of the Indus Periphery Cline. Groups that view themselves as being of traditionally priestly status, including traditional custodians of liturgical texts in the early Indo-European language Sanskrit, tend (with exceptions) to have more Steppe ancestry than expected based on ANI-ASI mixture, providing an independent line of evidence for a Steppe origin for South Asia’s Indo-European languages.
Iran and Turan
A West-to-East Cline of Decreasing Anatolian Farmer-Related Ancestry
We studied the genetic transformations accompanying the spread of agriculture eastward from Iran beginning in the 7th millennium BCE (41, 42). We replicate previous findings that 9th to 8th millennium BCE herders from the Zagros Mountains of western Iran harbored a distinctive West Eurasian-related ancestry profile (9, 27), while later groups across a broad region were admixed between this ancestry and that related to early Anatolian farmers. Our analysis reveals a west-to-east cline of decreasing Anatolian farmer-related admixture in the Copper Age and Bronze Age ranging from ~70% in Anatolia to ~31% in eastern Iran, to ~7% in far eastern Turan (Fig. 1; (13), Fig. S10, Table S8–S16). This suggests that the archaeologically documented spread of a shared package of plants and animal domesticates from diverse locations across this region was accompanied by bi-directional spread of people and mixture with the local groups they encountered (3, 41, 43, 44). We call this the Southwest Asian Cline. In the far east of the Southwest Asian Cline (eastern Iran and Turan) in individuals from the 3rd millennium BCE, we detect not only the smallest proportions of Anatolian farmer-related admixture but also admixture related to West Siberian Hunter Gatherers (WSHG) (plausibly reflecting admixture from unsampled hunter-gatherer groups who inhabited this region prior to the spread of Iranian farmer-related ancestry into it). This shows that North Eurasian-related ancestry impacted Turan well before the spread of descendants of Yamnaya Steppe pastoralists into the region. We can exclude the possibility that the Yamnaya were the source of this North Eurasian-related ancestry, as they had more EEHG-than WSHG-related ancestry, and also carried high frequencies of mitochondrial DNA haplogroup type U5a as well as Y chromosome haplogroup types R1b or R1a not represented in Iran and Turan in this period ((13), Table S93-S94).
People of the BMAC Were Not a Major Source of Ancestry for South Asians
From Bronze Age Iran and Turan, we obtained genome-wide data for 84 ancient individuals (3000–1400 BCE) who lived in four urban sites of the Bactria Margiana Archaeological Complex (BMAC) and its immediate successors. The great majority of these individuals fall in a cluster genetically similar to the preceding groups in Turan, consistent with the hypothesis that the BMAC coalesced from preceding pre-urban populations (5). We infer three primary genetic sources: early Iranian farmer-related ancestry (~60–65%), and smaller proportions of Anatolian farmer- (~20–25%) and WSHG-related ancestry (~10%). Unlike preceding Copper Age individuals from Turan, people of the BMAC cluster also harbored an additional 2–5% ancestry related (deeply in time) to Andamanese Hunter-Gatherers (AHG). This evidence of north-to-south gene flow from South Asia is consistent with the archaeological evidence of cultural contacts between the Indus Valley Civilization and the BMAC and the existence of an IVC trading colony in northern Afghanistan (although we lack ancient DNA from that site) (45), and stands in contrast to our qpAdm analyses showing that a reciprocal north-to-south spread is undetectable. Specifically, our analyses reject the BMAC and the people who lived before them in Turan as plausible major sources of ancestry for diverse ancient and modern South Asians by showing that their ratio of Anatolian farmer-related to Iranian farmer-related ancestry is too high for them to be a plausible source for South Asians (p<0.0001, χ2 test; (13), Fig S50–S51). A previous study (26) fit a model in which a population from Copper Age Turan was used a source of the Iranian farmer-related ancestry in present-day South Asians, thus raising the possibility that the people of the BMAC whom the authors correctly hypothesized were primarily derived from the groups that preceded them in Turan were a major source population for South Asians. However, that study only had access to 2 samples from this period compared to the 36 we report with this study, and it lacked ancient DNA from individuals from the BMAC period or from any ancient South Asians. With additional samples, we have the resolution to show that none of the large number of Bronze and Copper Age populations from Turan for which we have ancient DNA fit as a source for the Iranian farmer-related ancestry in South Asia.
Steppe Pastoralist-Derived Ancestry Arrived in Turan by 2100 BCE
Our large sample sizes from Central Asia, including individuals from BMAC sites, are a particular strength of this study, allowing us to detect outlier individuals with ancestry different from those living at the same time and place, and revealing cultural contacts that would be otherwise difficult to appreciate (Fig. 2). Around ~2300 BCE, we observe three outliers in BMAC-associated sites carrying WSHG-related ancestry and we report data from the third millennium BCE from three sites in Kazakhstan and one in Kyrgyzstan that fit as sources for them (related ancestry has been found in ~3500 BCE Botai culture individuals (26)). Yamnaya-derived ancestry arrived by 2100 BCE, since from 2100–1700 BCE we observe outliers from three BMAC-associated sites carrying ancestry ultimately derived from Western_Steppe_EMBA pastoralists, in the distinctive admixed form typically carried by many Middle to Late Bronze Age Steppe groups (with roughly two thirds of the ancestry being of Western_Steppe_EMBA origin, and the rest consistent with deriving from European farmers). Thus, our data document a southward movement of ancestry ultimately descended from Yamnaya Steppe pastoralists that spread into Central Asia by the turn of the 2nd millennium BCE.
Fig. 2. Outlier analysis reveals ancient contacts between sites.

We plot the average of Principal Component 1 (x-axis) and Principal Component 2 (y-axis) for the West Eurasian and All Eurasian PCA plots, as we found that this aids visual separation of the ancestry profiles. (A) In the Middle to Late Bronze Age Steppe, we observe in addition to the Western_Steppe_MLBA and Central_Steppe_MLBA clusters (indistinguishable in this projection), outliers admixed with other ancestries. The BMAC-related admixture in Kazakhstan documents northward gene flow onto the Steppe and confirms the Inner Asian Mountain Corridor as a conduit for movement of people. (B) At Shahr-i-Sokhta in eastern Iran, there are two primary groupings: one with ~20% Anatolian farmer-related ancestry and no detectable AHG-related ancestry, and the other with ~0% Anatolian farmer-related ancestry and substantial AHG-related ancestry (Indus Periphery Cline). (C) In individuals of the BMAC and successor sites, we observe a main cluster as well as numerous outliers: outliers >2000 BCE with admixture related to WSHG, outliers >2000 BCE on the Indus Periphery Cline (with an ancestral similar similar to the outliers at Shahr-i-Sokhta), and outliers after 2000 BCE that reveal how Central_Steppe_MLBA ancestry had arrived. (D) In the Late Bronze Age and Iron Age of northernmost South Asia, we observe a main cluster consistent with admixture between peoples of the Indus Periphery Cline and Central_Steppe_MLBA, and variable Steppe pastoralist-related admixture.
An Ancestry Profile Widespread During the Indus Valley Civilization
We document 11 outliers—3 with radiocarbon dates between 2500–2000 BCE from the BMAC site of Gonur, and 8 with radiocarbon dates or archaeological context dates between 3300 BCE to 2000 BCE from the eastern Iranian site of Shahr-i-Sokhta—that harbored elevated proportions of AHG-related ancestry (range of 11–50%) and the remainder from a distinctive mixture of Iranian farmer- and WSHG-related ancestry (~50–89%). These outliers had no detectable Anatolian farmer-related ancestry, in contrast with the main BMAC (~20–25% Anatolian-related) and Shahr-i-Sokhta (~16–21%) clusters, allowing us to reject both the BMAC and Shahr-i-Sokhta main clusters as sources for them (p<10−7, χ2 test; (13), Table S83). Without ancient DNA from individuals buried in IVC cultural contexts, we cannot make a definitive statement that the genetic gradient represented by these 11 outlier individuals, which we call the Indus Periphery Cline, was also an ancestry profile common in the IVC. Nevertheless, our result provides six circumstantial lines of evidence for this hypothesis. (i) These individuals had no detectable Anatolian farmer-related ancestry suggesting they descend from groups further east along the Anatolia-to-Iran cline of decreasing Anatolian farmer-related ancestry than any individuals we sampled from this period. (ii) All 11 outliers had elevated proportions of AHG-related ancestry, and two carried Y chromosome haplogroup H1a1d2 which today is primarily found in southern India. (iii) At both Gonur and Shahr-i-Sokhta there is archaeological evidence of exchange with the IVC (46, 47), and all the outlier individuals we directly dated fall within the time frame of the mature IVC. (iv) Several outliers at Shahr-i-Sokhta were buried with artifacts stylistically linked to Baluchistan in South Asia whereas burials associated with the other ancestries did not have these linkages (13). (v) In our modeling, the 11 outliers fit as a primary source of ancestry for 86 ancient individuals from post-IVC cultures living near the headwaters of the Indus River ~1200–800 BCE as well as diverse present-day South Asians, whereas no other ancient genetic clusters from Turan fit as sources for all these groups ((13), Fig S50). (vi) The estimated date of admixture between Iranian farmer-related and AHG-related ancestry in the outliers is several millennia before the time they lived (71 ± 15 generations, corresponding to a 95% confidence interval of ~5400–3700 BCE assuming 28 years per generation (13, 48). Thus, AHG-and Iranian farmer-related groups were in contact well before the time of the mature IVC at ~2600–1900 BCE as might be expected if the ancestry gradient was a major feature of a group that was living in the Indus Valley during the IVC.
The Steppe and Forest Zone
Ancestry Clines in Eurasia Established After the Advent of Farming
The late hunter-gatherer individuals from northern Eurasia lie along a west-to-east hunter-gatherer gradient of increasing relatedness to East Asians (Fig. 3). In the Neolithic and Copper Ages, hunter-gatherers at different points along this cline mixed with people with ancestry at different points along a southern cline to form five later clines, two of which were in the south (the Southwest Asian Cline and the Indus Periphery Cline which are described in the previous section), and three of which were in northern Eurasia (Fig. 3). Furthest to the west in the Steppe and Forest Zone there was the European Cline, established by the spread of farmers from Anatolia after ~7000 BCE and mixture with Western European Hunter-Gatherers (19). In far eastern Europe at latitudes spanning the Black and Caspian Seas there was the Caucasus Cline, consisting of a mixture of Eastern European Hunter-Gatherers and Iranian farmer-related ancestry with additional Anatolian farmer-related ancestry in some groups (49). East of the Urals we detect a Central Asian Cline, with WSHG individuals at one extreme and Copper Age and Early Bronze Age individuals from Turan at the other.
Fig. 3. Ancestry Transformations in Holocene Eurasia.

(A) Ancestry clines before and after the advent of farming. We document a South Eurasian Early Holocene Cline of increasing Iranian farmer- and West Siberian hunter-gatherer related ancestry moving west-to-east from Anatolia to Iran, and a North Eurasian Early Holocene Cline of increasing relatedness to East Asians moving west-to-east from Europe to Siberia. Mixtures of peoples along these two clines following the spread of farming formed five later gradients (shaded): moving west-to-east: the European Cline, the Caucasus Cline from which the Yamnaya formed, the Central Asian Cline which characterized much of Central Asia in the Copper and Bronze Ages, the Southwest Asian Cline established by spreads of farmers in multiple directions from several loci of domestication, and the Indus Periphery Cline. (B) Following the appearance of the Yamnaya Steppe pastoralists, Western_Steppe_EMBA (Yamnaya-like) ancestry then spread across this vast region. We use arrows to show plausible directions of spread of increasingly diluted ancestry (the arrows are not meant as exact routes which we do not have enough sampling to determine at present). Rough estimates of the timing of the arrival of this ancestry and estimated ancestry proportions are shown.
A Distinctive Ancestry Profile Stretching from Eastern Europe to Kazakhstan in the Bronze Age
Beginning around 3000 BCE, the ancestry profiles of many groups in Eurasia were transformed by the spread of Yamnaya Steppe pastoralist ancestry (Western_Steppe_EMBA) from its source in the Caucasus Cline (9, 49) to a vast region stretching from Hungary in the west to the Altai mountains in the east (7, 8) (Fig. 3). Over the next two millennia this ancestry spread further while admixing with local groups, eventually reaching the Atlantic shores of Europe in the west and South Asia in the southeast. The source of the Western_Steppe_EMBA ancestry that eventually reached Central Asia and South Asia was not the initial eastward expansion but instead a secondary expansion, which involved a group that had ~67% Western_Steppe_EMBA ancestry and ~33% ancestry from a point on the European Cline (8) (Fig. 3). We replicate previous findings that this group included people of the Corded Ware, Srubnaya, Petrovka, and Sintashta archaeological complexes spreading over a vast region from the border of eastern Europe to northwestern Kazakhstan (8, 19, 21), and our dataset adds more than one hundred individuals from this Western_Steppe_MLBA cluster. We also detect a further cluster, Central_Steppe_MLBA, which is differentiated from Western_Steppe_MLBA (p=7×10−6 by qpAdm), due to carrying ~9% additional ancestry derived from Bronze Age pastoralists of the central Steppe of primarily of WSHG-related ancestry (Central_Steppe_EMBA). Thus, individuals with Western_Steppe_MLBA ancestry admixed with local populations as they integrated eastward and southward.
Bidirectional Mobility Along the Inner Asian Mountain Corridor
As in Iran/Turan, the outlier individuals provide crucial information about human interaction.
First, our analysis of 50 individuals from the Sintashta culture cemetery of Kamennyi Ambar V reveals multiple groups of outliers whom we directly radiocarbon dated to be contemporaries of the main cluster but were genetically distinctive, indicating that this was a cosmopolitan site (Fig. 2). One set of outliers had elevated proportions of Central_Steppe_EMBA (largely WSHG-related) ancestry, another had elevated Western_Steppe_EMBA (Yamnaya-related), and a third had elevated EEHG-related ancestry.
Second, in the central Steppe (present-day Kazakhstan), an individual from one site dated to 2800–2500 BCE, and individuals from three sites dated to ~1600–1500 BCE, show significant admixture from Iranian farmer-related populations that is well-fit by the main BMAC cluster, demonstrating northward gene flow from Turan into the Steppe at the same time as there was southward movement of Central_Steppe_MLBA-related ancestry through Turan to South Asia. Thus, the archaeologically documented spreads of material culture and technology both north and south along the Inner Asian Mountain Corridor (50, 51), which began as early as the middle 3rd millennium BCE, were associated with substantial movements of people (Fig. 2).
Third, we observe individuals from Steppe sites (Krasnoyarsk) dated to ~1700–1500 BCE that derive up to ~25% ancestry from a source related to East Asians (well-modeled as ESHG), with the remainder best modeled as Western_Steppe_MLBA. By the Late Bronze Age, ESHG-related admixture became ubiquitous as documented by our time transect from Kazakhstan, and ancient DNA data from the Iron Age and from later periods in Turan and the central Steppe including Scythians, Sarmatians, Kushans, and Huns (25, 52). Thus, these 1st millennium BCE to 1st millennium CE archaeological cultures with documented cultural and political impacts on South Asia cannot be important sources for the Steppe pastoralist-related ancestry widespread in South Asia today (since present-day South Asians have too little East Asian-related ancestry to be consistent with deriving from these groups), providing an example of how genetic data can rule out scenarios that are plausible based on the archaeological and historical evidence alone ((13), Fig S52). Instead, our analysis shows that the only plausible source for the Steppe ancestry is Steppe Middle to Late Bronze Age groups, who not only fit as a source for South Asia but who we also document as having spread into Turan and mixed with BMAC-related individuals at sites in Kazakhstan in this period. Taken together, these results identify a narrow time window (first half of the second millennium BCE) when the Steppe ancestry that is widespread today in South Asia must have arrived.
The Genomic Formation of Human Populations in South Asia
Three Ancestry Clines That Succeeded Each Other in Time in South Asia
Previous work has shown that South Asians harbor ancestry from peoples related to ancient groups in northern Eurasia and Iran, East Asians, and Australasians (9). Here we document the process through which these deep sources of ancestry mixed to form later groups.
We begin with the pre-2000 BCE Indus Periphery Cline, described in an earlier section and detected in 11 outliers from two sites in cultural contact with the Indus Valley Civilization (Fig. 4). We can jointly model all individuals in this cline as a mixture of two source populations: one end of the cline is consistent with being entirely AHG-related, and the other is consistent with being 90% Iranian farmer-related and 10% WSHG-related (Fig. 4, (13)). People fitting on the Indus Periphery Cline form the majority of the ancestors of present-day South Asians. Through formal modeling, we demonstrate that it is this contribution of Indus Periphery Cline people to later South Asians, rather than westward gene flow bringing an ancestry unique to South Asia onto the Iranian plateau, that explains the high degree of shared ancestry between present-day South Asians and early Holocene Iranians (9, 13, 27).
Fig. 4. The Genomic Formation of South Asia.

(A) The degree of allele sharing with southern Asian hunter-gatherers (AASI) measured by f4(Ethiopia_4500BP, X; Ganj_Dareh_N, AHG) and with Steppe pastoralists measured by f4(Ethiopia_4500BP, X; Central_Steppe_MLBA, Ganj_Dareh_N) reveals three ancestry clines that succeeded each other in time: the Indus Periphery Cline prior to ~2000 BCE, the Steppe Cline represented by northern South Asian individuals after ~2000 BCE, and the Modern Indian Cline. (B) Modeling South Asians as a mixture of Central_Steppe_MLBA, AHG (as a proxy for AASI), and Indus_Periphery_West (the individual from the Indus Periphery Cline with the least AASI ancestry). Groups along the edges of the triangle fit a two-way model, and in the interior only fit a three-way model. The 140 present-day South Asian groups on the Modern Indian Cline are shown as small dots. (C) Groups that traditionally view themselves as being of priestly status in this and the preceding panel are shown in red (“Brahmin,” “Pandit,” and “Bhumihar” but excluding “Catholic Brahmins”), and tend to have a significantly higher ratio of Central_Steppe_MLBA to Indus_Periphery_Cline ancestry than other groups. (D) Plot of the proportion of Central_Steppe_MLBA ancestry on the autosomes (x-axis) and the Y chromosome (y-axis) shows that the source of this ancestry is primarily from females in Late Bronze Age and Iron Age individuals from the Swat District of northernmost South Asia, and primarily from males in most present-day South Asians.
We next characterized the 2000 BCE Steppe Cline, represented in our analysis by 117 individuals dating to 1400 BCE - 1700 CE from the Swat and Chitral districts of northernmost South Asia (Fig. 2, Fig. 4). We found that we could jointly model all individuals on the Steppe Cline as a mixture of two sources albeit different from the two sources in the earlier cline. One end is consistent with a point along the Indus Periphery Cline. The other end is consistent with a mixture of about 41% Central_Steppe_MLBA ancestry and 59% from a subgroup of the Indus Periphery Cline with relatively high Iranian farmer-related ancestry ((13), Fig S50).
To understand the formation of the Modern Indian Cline, we searched for triples of populations that could fit as sources for diverse present-day South Asians groups as well as peoples of the Steppe Cline. All fitting models include as sources Central_Steppe_MLBA (or a group with a similar ancestry profile), a group of Indus Periphery Cline individuals, and either AHG or a subgroup of Indus Periphery Cline individuals with relatively high AHG-related ancestry (13), Fig S51). Co-analyzing 140 diverse South Asian groups (10) that fall on a gradient in PCA (13), we show that while there are three deep sources, just as in the case of the earlier two clines the great majority of groups on the Modern Indian Cline can be jointly modeled as a mixture of two populations that are mixed from the earlier three. While we do not have ancient DNA data from either of the two statistically reconstructed source populations for the Modern Indian Cline, the ASI or the ANI, in what follows we co-analyze our ancient DNA data in conjunction with modern data to characterize the exact ancestry of the ASI, and to provide constraints on the ANI.
The ASI and ANI Arose as Indus Periphery Cline People Mixed with Groups to the North & East
To gain insight into the formation of the ASI, we extrapolated to the least West Eurasian-related theoretical extreme of the Modern Indian Cline by setting the Central_Steppe_MLBA ancestry proportion to zero in our model. We estimate a minimum of 55% ancestry from people on the Indus Periphery Cline (by representing the Indus Periphery Cline by the individual on it with the most Iranian farmer-related ancestry, which we call Indus_Periphery_West), and modeling the reminder of the ancestry as deriving from an AHG-related group (13). We find that several tribal groups from southern India are consistent with ~0% Central_Steppe_MLBA ancestry (13). The fact that these individuals match the most extreme possible position for the ASI not only reveals that nearly direct descendants of the ASI live today in South Asia, but also allows us to make a precise statement about the ancestry profile of the ASI. In particular, the fact that they harbor substantial Iranian farmer-related ancestry (via the Indus Periphery Cline), disproves earlier suggestions that the ASI might not have any ancestry related to West Eurasians (11). Using the DATES software, we estimate an average of 107 ± 11 generations since admixture of the Iranian farmer-related and AHG-related groups in one of these groups: Palliyar. This corresponds to a 95% confidence interval of 1700–400 BCE assuming 28 years per generation (53). Thus, the ASI were note fully formed at the time of the IVC, and instead must have continued to form through mixture after its decline as material culture typical of the IVC spread eastward (54) and Indus Periphery Cline ancestry mixed with people of less West Eurasian relatedness.
We also obtained additional evidence for a late (Bronze Age) formation of the ASI by building an admixture graph using qpGraph, co-modeling Palliyar and Juang (an Austroasiatic-speaking group in India with low West Eurasian-relatedness) (Fig. 5). The graph fits the component of South Asian ancestry with no West Eurasian relatedness (AASI - “Ancestral Ancient South Asians”) as an Asian lineage that split off around the time that East Asian, Andaman Islander, and Australian aboriginal ancestors separated from each other, consistent with the hypothesis that eastern and southern Asian lineages derives from an eastward spread that in a short span gave rise to lineages leading to AASI, East Asians, Andamanese Hunter Gatherers, and Australians (55) (Fig. 5). The Juang cannot be fit through a mixture of ASI ancestry and ancestry related to Austroasiatic language speakers, and instead can only be fit by modeling additional ancestry from AASI, showing that at the time Austroasiatic groups formed in South Asia, groups with less Iranian farming-related ancestry than in the ASI were also present. Austroasiatic languages are hypothesized to have spread into South Asia in the 3rd millennium BCE (based on hill cultivation systems hypothesized to be associated with the spread of Austroasiatic languages (42), and thus the ancestry profile of the Juang provides an independent line of evidence for a late (Bronze Age and plausibly post-IVC) formation of the ASI.
Fig. 5. Admixture Graph Model.

The largest deviation between empirical and theoretical f-statistics is |Z|=2.9, indicating a good fit considering the large number of f-statistics analyzed. Admixture events are shown as dotted lines labeled by proportions, with the minor ancestry in gray. The present-day groups are shown in orange ovals, the ancient ones in blue, and unsampled groups in white. (The ovals and admixture events are positioned according to guesses about their relative dates to help in visualization, although the dates are in no way meant to be exact.) In this graph we do not attempt to model the contribution of WSHG and Anatolian farmer-related ancestry, and thus cannot model Central_Steppe_EMBA, the proximal source of Steppe ancestry in South Asia (instead we model the Steppe ancestry in South Asia through the more distally related Yamnaya). However, the admixture graph does highlight several key findings of the study, including the deep separation of the AASI from other Eurasian lineages, and the fact that some Austroasiatic-speaking groups in South Asia (e.g. Juang) harbor ancestry from a South Asian group with a higher ratio of AASI-related to Iranian farmer-related ancestry than any groups on the Modern Indian Cline, thus revealing that groups with substantial Iranian farmer-related ancestry were not ubiquitous in peninsular South Asia in the 3rd millennium BCE when Austroasiatic languages likely spread across the subcontinent.
To shed light on the formation of the statistically reconstructed ANI, we return to the Swat Valley time transect that formed the Steppe Cline after 2000 BCE. The Modern Indian Cline intersects the Steppe Cline at a position close to the position of the Kalash, the group in northwest South Asia with the highest ANI ancestry proportion (56) (Fig. 4). The DATES-based estimate of admixture in the Kalash is 110 ± 12 generations (56), suggesting a post-IVC date of formation of the ANI paralleling the post-IVC date of formation of the ASI. Further evidence for a post-IVC integration of Steppe ancestry into South Asia comes from ancient individuals on the Steppe Cline (along which the ANI could theoretically have formed) whose admixture date for Steppe ancestry is also post-IVC. Specifically, we estimate the date of admixture into the Late Bronze Age and Iron Age individuals from the Swat District of northernmost South Asia to be on average 26 generations before the date that they lived, corresponding to a 95% confidence interval of ~1900–1500 BCE. This time scale for the arrival of Steppe ancestry in the region is consistent with our observation of 6 outlier individuals in Turan who lived between ~2000–1500 BCE and who carry this ancestry in mixed form (Fig. 2), and with our finding that the R1a Y chromosome associated with Central_Steppe_MLBA ancestry in South Asia is also present in the Swat District Late Bronze and Iron Age individuals (2 copies).
Taken together, these results show neither of the two primary source populations of the Modern Indian Cline, the ANI and ASI, was fully formed before the turn of the 2nd millennium BCE.
Steppe Ancestry in South Asia is Primarily from Males and Disproportionately High in Brahmins
In the Late Bronze Age and Iron Age individuals of the Swat Valley, we detect a significantly lower proportion of Steppe admixture on the Y chromosome (only 5% of the 44 Y chromosomes of the R1a-Z93 subtype that occurs at 100% frequency in the Central_Steppe_MLBA males) compared to 20% on the autosomes (Z = −3.9 for a deficiency from males under the simplifying assumption that all the Y chromosomes are unrelated to each other since admixture and thus statistically independent), documenting how Steppe ancestry was incorporated into these groups largely through females (Fig. 4). However, sex bias varied in different parts of South Asia, as in present-day South Asians we observe a reverse pattern of excess Central_Steppe_MLBA-related ancestry on the Y chromosome compared to the autosomes (Z = 2.7 for an excess from males) (13, 57) (Fig. 4). Thus, the introduction of lineages from Steppe pastoralists into the ancestors of present-day South Asians was mediated mostly by males. This bias is similar in direction to what has been documented for the introduction of Steppe ancestry into Iberia in far western Europe, although the bias is less extreme than reported in that case (58).
Our analysis of Steppe ancestry also identified 6 groups with a highly significantly elevated ratio of Central_Steppe_MLBA-to-Indus_Periphery_West-related ancestry compared to the expectation for the model at the Z < −4.5 level. The strongest two signals were in Brahmin_Tiwari (Z = −7.9) and Bhumihar_Bihar (Z = −7.0). More generally, there is a notable enrichment in groups that consider themselves to be of traditionally priestly status: 5 of the 6 groups with Z < −4.5 were Brahmins or Bhumihars even though they comprise only 7–11% of the 140 groups analyzed (p<10−12 by a χ2 test assuming all the groups evolved independently). We caution that this is not a formal test as there is an unknown degree of shared ancestry among groups since they formed by mixture, and because our decisions about which groups to include in the analysis was not made in a blinded way; for example, we excluded four “Catholic Brahmin” groups with strong evidence of substantial shared ancestry in the last millennium (10) which makes them not statistically independent (Table S5, Fig. 4 (13)). Nevertheless, the fact that traditional custodians of liturgy in Sanskrit (Brahmins) tend to have more Steppe ancestry than is predicted by a simple ASI-ANI mixture model provides an independent line of evidence, beyond the distinctive ancestry profile shared between South Asia and Bronze Eastern Europe mirroring the shared features of Balto-Slavic and Indo-Iranian languages (59), for a Steppe origin for South Asia’s Indo-European languages prior to ~2000 BCE.
Discussion
Our analysis reveals that the ancestry of the greater South Asian region in the Holocene was characterized by at least three genetic gradients. Prior to ~2000 BCE, there was the Indus Periphery Cline consisting of people with different proportions of Iranian farmer- and AASI-related ancestry, which we hypothesize was a characteristic feature of many IVC people. The ASI formed after 2000 BCE as a mixture of a point along this cline with South Asians with higher proportions of AASI-related ancestry. Between about 2000 and 1000 BCE, people of largely Central_Steppe_MLBA ancestry expanded toward South Asia, mixing with people along the Indus Periphery Cline to form the Steppe Cline. Multiple points along the Steppe Cline are represented by individuals of the Swat Valley time transect and statistically we find that the ANI, one of the two primary source population of South Asia, can fit along the Steppe Cline. After 2000 BCE, mixtures of mixed populations—the ASI and ANI—mixed themselves to form the Modern Indian Cline, which is represented today in diverse groups in South Asia (Fig. 4).
Our finding based on admixture linkage disequilibrium ((13), Fig S59) that the mixture that formed the Indus Periphery Cline occurred by ~5400–3700 BCE—at least a millennium before the formation of the mature IVC—raises two possibilities. One is that Iranian farmer-related ancestry in this group was characteristic of the Indus Valley hunter-gatherers in the same way as it was characteristic of northern Caucasus and Iranian plateau hunter-gatherers. The presence of such ancestry in hunter-gatherers from Belt and Hotu Caves in northeastern Iran increases the plausibility that this ancestry could have existed in hunter-gatherers further east. An alternative is that this ancestry reflects movement into South Asia from the Iranian plateau of people accompanying the eastward spread of wheat and barley agriculture and goat and sheep herding as early as the 7th millennium BCE and forming early farmer settlements such as those at Mehrgarh in the hills flanking the Indus Valley (60, 61). However, this is in tension with the observation that the Indus Periphery Cline people had little if any Anatolian farmer-related ancestry, which is strongly correlated with the eastward spread of crop-based agriculture in our dataset. Thus, while our analysis supports the idea that eastward spread of Anatolian farmer-related ancestry was associated with the spread of farming to the Iranian plateau and Turan, our results do not support large-scale movements of ancestry from the Near East into South Asia following ~6000 BCE (the time after which all ancient individuals from Iran in our data have Anatolian farmer-related ancestry even though South Asians have very little). Languages in pre-state societies usually spread through movements of people (62), and thus the absence of much Anatolian farmer-related ancestry in the Indus Periphery Cline suggests that the Indo-European languages spoken in South Asia today are unlikely to owe their origin to the spread of farming from West Asia.
Our results not only provide negative evidence against an Iranian plateau origin for Indo-European languages in South Asia, but also positive evidence for the theory that these languages spread from the Steppe. While ancient DNA has documented westward movements of Steppe pastoralist ancestry providing a likely conduit for the spread of many Indo-European languages to Europe (7, 8), the chain-of-transmission into South Asia has been unclear because of a lack of relevant ancient DNA. Our observation of the spread of Central_Steppe_MLBA ancestry into South Asia in the first half of the 2nd millennium BCE provides this evidence, and is particularly striking as it provides a plausible genetic explanation for the linguistic similarities between the Balto-Slavic and Indo-Iranian sub-families of Indo-European, which despite their vast geographic separation, share the Satem innovation and Ruki sound laws (63). If the spread of people from the Steppe in this period was a conduit for the spread of South Asian Indo-European languages, then it is striking that there are so few material culture similarities between the central Steppe and South Asia in the Middle to Late Bronze Age (i.e. after the middle of the 2nd millennium BCE). Indeed, the material culture differences are so substantial that some archaeologists recognize no evidence of a connection. However, lack of material culture connections does not provide evidence against spread of genes, as has been demonstrated in the case of the Beaker Complex, which originated largely in western Europe, but in Central Europe was associated with skeletons that harbored ~50% ancestry related to Yamnaya Steppe pastoralists (18). Thus, in Europe we have an unambiguous example of people with ancestry from the Steppe making profound demographic impacts on the regions into which they spread while adopting important aspects of local material culture. Our findings document a similar phenomenon in South Asia, with the locally acculturated population harboring up to ~20% Western_Steppe_EMBA-derived ancestry according to our modeling (via Central_Steppe_MLBA groups) (Fig. 3). Our analysis also provides a second line of evidence for a linkage between Steppe ancestry and Indo-European languages. Steppe ancestry enrichment in groups that view themselves as being of traditionally priestly status is striking as some of these groups including Brahmins are traditional custodians of literature composed in early Sanskrit. A possible explanation is that the influx of Central_Steppe_MLBA ancestry into South Asia in the mid-2nd millennium BCE created a meta-population with varied proportions of Steppe ancestry, with people of more Steppe ancestry (or admixing less with Indus Periphery Cline groups) tending to be more strongly associated with Indo-European culture. Due to strong endogamy, which kept groups generally isolated from neighbors for thousands of years (7), some of this population substructure persists in South Asia among present-day custodians of Indo-European texts.
Our findings also shed light on the origin of its second-largest language group in South Asia, Dravidian. The strong correlation between ASI ancestry and present-day Dravidian languages suggests that the ASI, which we have shown formed as groups with ancestry typical of the Indus Periphery Cline moved south and east after the decline of the IVC to mix with groups with more AASI ancestry, most likely spoke an early Dravidian language. A possible scenario combining genetic data with archaeology and linguistics is that proto-Dravidian was spread by peoples of the IVC along with the Indus Periphery Cline ancestry component of the ASI. Non-genetic support for an IVC origin for Dravidian languages includes the present-day geographic distribution of these languages (in southern India and southwestern Pakistan), and a suggestion that some symbols on ancient Indus Valley seals denote Dravidian words or names (64, 65). An alternative possibility is that proto-Dravidian was spread by the approximately half of the ASI’s ancestry that was not from the Indus Periphery Cline, and instead derived from the south and the east (peninsular South Asia). The southern scenario is consistent with reconstructions of Proto-Dravidian terms for flora and fauna unique to peninsular India (66, 67).
We finally highlight a remarkable parallel between the prehistory of South Asia and Europe. In both regions there were exchanges between people related to Southwest Asian people and local people; mixtures of these groups led to the Indus Periphery Cline in South Asia and the European Cline in Europe. In both subcontinents, people arriving in the 3rd and 2nd millennia BCE who descended from mixtures of people related to Yamnaya Steppe pastoralists and European farmers mixed further with local populations: in South Asia forming the ANI, and in Europe forming groups like that of the Beaker Complex. In both cases, mixtures of these mixed populations—those with Steppe pastoralist-related admixture and those without—drive the modern ancestry clines in both regions (Fig. 3). However, there are also profound differences between the Bronze Age and Neolithic spreads of ancestry across the two subcontinents. One is that the maximum proportion of local ancestry is higher in South Asia (AASI ancestry of up to ~60%) than Europe (WEHG ancestry of up to ~30%) (7), which could reflect stronger ecological or cultural barriers to the spreads of people in South Asia than in Europe, allowing the previously established groups more time to adapt and mix with incoming groups. A second difference is the smaller proportion of Steppe pastoralist-related ancestry in South Asia than in Europe, its later arrival by ~500–1000 years, and a lower male sex bias in the admixture, factors that help to explain the continued persistence of a large fraction of non-Indo-European speakers amongst people of present-day South Asia today. The situation in South Asia is somewhat reminiscent of Mediterranean Europe where the proportion of Steppe ancestry is considerably lower than that of northern and central Europe (Fig. 3), and where many non-Indo European languages are attested in classical times (68). Further studies of ancient DNA from South Asia and the linguistically related Iranian world will extend and add nuance to the model presented here.
Materials and Methods
Ancient DNA Laboratory Work.
For the skeletal elements that we were not able to transport from field sites, we drilled directly into bone, for the most part focusing on inner ear portions of petrous bones using a method for sampling from the cranial base (CBD) (70). The great majority of skeletal elements were prepared in dedicated ancient DNA clean rooms at Harvard Medical School, University College Dublin, the University of Vienna, or the Max Planck Institute for Evolutionary Anthropology in Leipzig Germany, either by drilling, or by sandblasting to isolate a bone piece followed by milling (Table S1, Table S2).
All the molecular work except for that on a single individual (Darra-i-Kur) was carried out at Harvard Medical School (HMS). At HMS, we extracted DNA using a method that is optimized to retain small DNA fragments. We implemented this method both using a manual method based on silica spin columns (565 libraries) (14, 15), and with the assistance of robotic liquid handlers using silica coated magnetic beads and Buffer D (149 libraries) (71). We converted the DNA into a form that could be sequenced using a double-stranded library preparation protocol (711 libraries) and a single stranded library preparation protocol (3 libraries) (72). For all but four of the double stranded libraries, we pre-treated with a mixture of the enzymes Uracil-DNA Glycosylase (UDG) and Endo VIII (USER, New England Biolabs) to reduce characteristic cytosine-to-thymine errors in ancient DNA except in the terminal base (17). The remaining four libraries were not pre-treated with USER (73). The three single-stranded libraries were also pre-treated with USER in a way that results in a similar damage pattern of inefficient uracil removal in the terminal base (72). We prepared most double stranded libraries (n=524) with the assistance of a robotic liquid handler, substituting the MinElute columns used for cleaning up reactions in manual processing with silica coated magnetic beads in robotic processing, and the MinElute column-based PCR cleanup at the end of library preparation with SPRI beads (74, 75). We enriched all libraries both for sequences overlapping mitochondrial DNA (76), and for sequences overlapping about 1.24 million nuclear targets. We carried out two rounds of enrichment for these targets (7, 19, 20) either in two independent capture experiments or together. After indexing the enrichment products in a way that assigned a unique index and combination to each library (77), we sequenced the enriched products on an Illumina NextSeq500 instrument using v.2 150 cycle kits for 2×76 cycles and 2×7 cycles (2×8 for single-stranded libraries), and sequenced up to the point that the expected number of additional SNPs covered per 100 additional read pairs sequenced was less than about 1. We also shotgun sequenced double-stranded libraries to assess the fraction of sequences that mapped to the human genome.
To analyze the data, we began by sorting the read pairs by searching for the expected identification indices and barcodes for each library, allowing up to one mismatch from the expected sequence in each case. We removed adapters and merged together sequences requiring a 15 base pair overlap (allowing up to one mismatch), taking the highest quality base in the merged segment to represent the allele. We mapped the resulting sequences to the hg19 human reference (GRCh37, the version used for the 1000 Genomes project (78)) using the samse command of BWA (79) (version 0.6.1). We removed duplicate sequences (mapping to the same position in the genome and having the same barcode pair), and merged libraries corresponding to the same sample (merging across samples when the genetic data revealed multiple samples from the same individual). For each individual, we restricted to sequences passing filters (not overlapping known insertion/deletion polymorphisms, and having a minimum mapping quality 10), and trimmed two nucleotides from the end of each sequence to reduce deamination artifacts. We also further restricted to sequence data with a minimum base quality of 20. To represent each individual at each SNP position, we randomly selected a single sequence (if one was available).
For Darra-i-Kur, we analyzed a single-stranded DNA library (L5082) at the Max-Planck-Institute for Evolutionary Anthropology (MPI-EVA) in Leipzig, Germany, generated as part of a previous study (80). The previous study only analyzed mitochondrial DNA, and for the current study, we enriched the library for sequences overlapping the same panel of about 1.2 million nuclear targets using two rounds of hybridization capture (7, 19, 20). We sequenced the enriched libraries on 2 lanes of an Illumina HiSeq2500 platform in a double index configuration (2×76 cycles) (77), and we determined alleles using FreeIbis (81). We merged overlapping paired-end and trimmed using leeHom (82). We used BWA to align the sequences to the human reference genome hg19 (GRCh37) (79). We retained sequences showing a perfect match to the expected index combination for downstream analyses.
We assessed evidence for ancient DNA authenticity by measuring the rate of damage in the first nucleotide, flagging individuals as potentially contaminated if they had less than a 3% cytosine-to-thymine substitution rate in the first nucleotide for a UDG-treated library and less than a 10% substitution rate for a non-UDG-treated library. We used contamMix to test for contamination based on polymorphism in mitochondrial DNA (83), and ANGSD to test for contamination based on polymorphism on the X chromosome in males (84).
Radiocarbon dating.
We generated 269 radiocarbon (14C) dates on bone using accelerator mass spectrometry (AMS) (Table S3). Most of these (n=242) were generated at the Pennsylvania State University (PSU) Radiocarbon Laboratory, and here we excerpt a description of the sample preparation methodology at PSU (the methods used at the other laboratories are publicly available and we refer readers to the literature for those methodologies). Possible contaminants (conservants and adhesives) were removed by sonicating all bone samples in successive washes of ACS grade methanol, acetone, and dichloromethane for 30 minutes each at room temperature, followed by three washes in Nanopure water to rinse. Bone collagen for 14C was extracted and purified using a modified Longin method with ultrafiltration (>30kDa gelatin; (85)). If collagen yields were low and amino acids poorly preserved we used a modified XAD process (XAD Amino Acids; (86)). For quality assurance we measured carbon and nitrogen concentrations and C/N ratios of all extracted and purified collagen/amino acid samples with a Costech elemental analyzer (ECS 4010). We evaluated quality based on % crude gelatin yield, %C, %N and C/N ratios before AMS 14C dating. C/N ratios for all directly radiocarbon samples fell between 2.9 and 3.6, indicating excellent preservation (87). Collagen/amino acid samples (~2.1 mg) were then combusted for 3 h at 900°C in vacuum-sealed quartz tubes with CuO and Ag wires. Sample CO2 was reduced to graphite at 550°C using H2 and a Fe catalyst, with reaction water drawn off with Mg(ClO4)2 (88). Graphite samples were pressed into targets in Al boats and loaded on a target wheel with OX-1 (oxalic acid) standards, known-age bone secondaries, and a 14C-free Pleistocene whale blank. All 14C measurements were made on a modified National Electronics Corporation compact spectrometer with a 0.5 MV accelerator (NEC 1.5SDH-1). The 14C ages were corrected for mass-dependent fractionation with measured δ13C values (89) and compared with samples of Pleistocene whale bone (backgrounds, 48,000 14C BP), late Holocene bison bone (~1,850 14C BP), late 1800s CE cow bone, and OX-2 oxalic acid standards. All calibrated 14C ages were calculated using OxCal version 4.3 (Ramsey and Lee 2013) using the IntCal13 northern hemisphere curve (90), and we quote 95% confidence intervals (2-sigma ranges).
Principal component analysis (PCA)
We carried out PCA using the smartpca package of EIGENSOFT 7.2.1 (36). We used default parameters and added two options (lsqproject:YES and numoutlieriter:0) to project the ancient individuals onto the PCA space. We used two basis sets for the projection: the first based on 1,340 present-day Eurasians genotyped on the Affymetrix Human Origins array, and the second based on a subset of 991 present-day West Eurasians (7, 32, 33). These projections are shown repeatedly in (13) and are used in the Online Data Visualizer. We also computed FST between groups using the parameters inbreed:YES and fstonly:YES. We restricted these analyses to the dataset obtained by merging our ancient DNA data with the modern DNA data on the Human Origins array and restricting to 597,573 SNPs. We treated positions where we did not have sequence data as missing genotypes.
ADMIXTURE clustering
Using PLINK2 (91), we first pruned our dataset using the --geno 0.7 option to ensure that we only performed our analysis on sites where at least 70% of individuals were covered by at least one sequence. This resulted in 892,613 SNPs. Individuals without coverage on specific SNPs were assigned missing data at those sites. We ran ADMIXTURE (37) with 10 replicates, reporting the replicate with the highest likelihood. We show results for K=5 in (13), as we found that this provides good resolution for disambiguating the sources of pre-Copper Age ancestry in the ancient individuals.
f-statistics
We used the qp3pop and qpDstat packages in ADMIXTOOLS to compute f3-statistics and f4-statistics. We used the inbreed:YES parameter to compute f3-statistics as a test for admixture with an ancient population as a target, with all ancient genomes as sources. Using the f4Mode:YES parameter in qpDstat, we also computed two sets of f4-symmetry statistics to evaluate if pairs of populations are consistent with forming a clade relative to a comparison population. The first is a “Two-population comparison” statistic where we compare all possible pairs of ancient groups (the Test populations) to a panel of populations that encompasses diverse pre-Copper Age and more widespread genetic variation. Thus, we compute a statistic of the form f4(Test 1, Test 2; Pre-Copper Age, Mbuti). The second is a “Pre-Copper Age affinity” statistic that compares each ancient group in turn against all possible pairs of Pre-Copper Age populations, using statistics of the form f4(Pre-Copper Age 1, Pre-Copper Age 2; Test, Mbuti).
Modeling admixture history
We used qpAdm (33) in the ADMIXTOOLS software package to estimate the proportions of ancestry in a Test population deriving from a mixture of N ‘reference’ populations by leveraging (but not explicitly modeling) shared genetic drift with a set of ‘Outgroup’ populations. We set the details:YES parameter, which reports a normally distributed Z-score for the goodness of fit of the model (estimated with a Block Jackknife).
Hierarchical modeling.
For each group on a proposed cline, we used qpAdm to obtain estimates for the proportion of ancestry from hypothesized source populations, along with the covariance matrix across groups. We jointly modeled these estimates using a bivariate normal model (forcing the three proportions to sum to 100%) and estimated the mean and covariance of the two parameters using maximum likelihood. With this inferred matrix, we tested whether the cline could be modeled by a mixture of two primary source populations. First, we tested if the covariance matrix is consistent with being singular, implying that knowledge of the proportion of ancestry from one of the mixing components was consistent with being fully predictive of the other two, as expected for two-way mixture. Second, if we were able to establish that this was the case, we examined the difference between the expected and observed ratios of the ancestry proportions of the analyzed groups within this generative model by fitting all the groups simultaneously. This resulted in a handful of groups deviating from expectation.
Method for dating admixture events.
We implemented an algorithm, DATES, which leverages ancestry covariance patterns that can be measured in a single individual (instead of admixture LD that requires multiple individuals). Full details of the approach and simulations documenting its efficacy in modern as well as ancient data are presented in (13). The software implementing DATES is available at https://zenodo.org/record/3263997#.XRnebJNKj6A (DOI: 10.5281/zenodo.3263997).
Supplementary Material
Acknowledgments:
We acknowledge the people past and present whose samples we analyzed in this study. We thank Oliver Uberti for the design of Figure 3 and the figure associated with the One Page Summary. We thank the Minusinsk Regional Museum of N. M. Martyanov for sharing skeletal samples. We thank Orazak Ismagulov, Ainagul Ismagulova, and Akmatov Kunbolot Toktonsunovich for facilitating access to skeletal material.We thank Orazak Ismagulov, Ainagul Ismagulova, and Akmatov Kunbolot Toktonsunovich for facilitating access to skeletal material We thank the Department of Archaeology and Museums, Government of Pakistan, the Directorate of Archaeology and Museums, Government of Khyber-Pakhtunkhwa Province (Pakistan), and the Dipartimento di Biologia Ambientale, Sapienza Università (Rome) for facilitating access to the materials from Swat excavated by the Italian Archaeological Mission (now ISMEO).
Funding: N.P. carried out this work while a fellow at the Radcliffe Institute for Advanced Study at Harvard University. P.M. was supported by a Burroughs Wellcome Fund CASI award. N.N. is supported by a NIGMS (GM007753) fellowship. T.C. and A.D. were supported by the Russian Science Foundation (project no. 14–50-00036). T.S. was supported by the Russian Foundation for Basic Research (project no. grant 8–09-00779) “Anthropological and archaeological aspects of ethnogenesis of the population of the southern part of Western and Central Siberia in the Neolithic and Early Bronze Age.” D.P., S.S. and D.L. were supported by European Research Council ERC-2011-AdG 295733 grant (Langelin). O.M. was supported by a grant from the Ministry of Education and Sciences of the Russian Federation No 33. 1907, 2017/ П4 “Traditional and innovational models of a development of ancient Volga population”. A.E. was supported by a grant from the Ministry of Education and Sciences of the Russian Federation No 33.5494, 2017/BP “Borderlands of cultural worlds (Southern Urals from Antiquity to Early Modern period)”. Radiocarbon dating work supported by the NSF Archaeometry program BCS-1460369 to D.Ken. and B.C. and by the NSF Archaeology program BCS-1725067 to D.Ken. K.Th. was supported by NCP fund (MLP0117) of the Council of Scientific and Industrial Research (CSIR), Government of India, New Delhi. N.Bo., A.N., and M.Z.. were supported by the Max Planck Society. D.Re. is an Investigator of the Howard Hughes Medical Institute and his ancient DNA laboratory work was supported by National Science Foundation HOMINID grant BCS-1032255, by National Institutes of Health grant GM100233, by an Allen Discovery Center grant, and by grant 61220 from the John Templeton Foundation.
Footnotes
Competing interests: The authors declare no competing interests.
Data and materials availability: All newly reported sequencing data are available from the European Nucleotide Archive, accession number PRJEB32466, and the software for dating admixture events in ancient samples is available at https://zenodo.org/record/3263997#.XRnebJNKj6A (DOI: 10.5281/zenodo.3263997). The Online Data Visualizer is available at: https://public.tableau.com/views/TheGenomicFormationofSouthandCentralAsia/Fig_1.
Supplementary Materials:
Materials and Methods
Online Data Visualizer for exploring data: https://public.tableau.com/views/TheGenomicFormationofSouthandCentralAsia/Fig_1.
Tables S1-S93
Figures S1-S61
Genotypes for newly reported individuals
References and Notes
- 1.Online Data Visualizer, (available at https://public.tableau.com/views/TheGenomicFormationofSouthandCentralAsia/Fig_1).
- 2.Fuller DQ, Lucas L, in Human Dispersal and Species Movement, Boivin N, Petraglia M, Crassard R, Eds. (Cambridge University Press, Cambridge, 2017), pp. 304–331. [Google Scholar]
- 3.Stevens CJ et al. , Between China and South Asia: A Middle Asian corridor of crop dispersal and agricultural innovation in the Bronze Age. The Holocene. 26, 1541–1555 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Allaby RG, Stevens C, Lucas L, Maeda O, Fuller DQ, Geographic mosaics and changing rates of cereal domestication. Philos. Trans. R. Soc. B Biol. Sci 372, 20160429 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dani AH et al. , History of Civilizations of Central Asia: The Development of Sedentary and Nomadic Civilizations, 700 B. C. to A (UNESCO Publishing, 1994). [Google Scholar]
- 6.Frachetti MD, Smith CE, Traub CM, Williams T, Nomadic ecology shaped the highland geography of Asia’s Silk Roads. Nature. 543, 193–198 (2017). [DOI] [PubMed] [Google Scholar]
- 7.Haak W et al. , Massive migration from the steppe was a source for Indo-European languages in Europe. Nature. 522, 207–211 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Allentoft ME et al. , Population genomics of Bronze Age Eurasia. Nature. 522, 167–172 (2015). [DOI] [PubMed] [Google Scholar]
- 9.Lazaridis I et al. , Genomic insights into the origin of farming in the ancient Near East. Nature. 536, 419–424 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nakatsuka N et al. , The promise of discovering population-specific disease-associated genes in South Asia. Nat. Genet 49, 1403–1407 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reich D, Thangaraj K, Patterson N, Price AL, Singh L, Reconstructing Indian population history. Nature. 461, 489–494 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moorjani P et al. , Genetic evidence for recent population mixture in India. Am. J. Hum. Genet 93, 422–438 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Supplementary Materials.
- 14.Dabney J et al. , Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. U. S. A 110, 15758–15763 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Korlević P et al. , Reducing microbial and human contamination in DNA extractions from ancient bones and teeth. Biotechniques. 59, 87–93 (2015). [DOI] [PubMed] [Google Scholar]
- 16.Meyer M et al. , A high-coverage genome sequence from an archaic Denisovan individual. Science. 338, 222–226 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rohland N, Harney E, Mallick S, Nordenfelt S, Reich D, Partial uracil-DNA-glycosylase treatment for screening of ancient DNA. Philos. Trans. R. Soc. B Biol. Sci 370, 20130624 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Olalde I et al. , The Beaker phenomenon and the genomic transformation of northwest Europe. Nature. 555, 190–196 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mathieson I et al. , Genome-wide patterns of selection in 230 ancient Eurasians. Nature. 528, 499–503 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fu Q et al. , The genetic history of Ice Age Europe. Nature. 534, 200–205 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang MA et al. , 40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia. Curr. Biol 27, 3202–3208 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Saag L et al. , Extensive Farming in Estonia Started through a Sex-Biased Migration from the Steppe. Curr. Biol 27, 2185–2193 (2017). [DOI] [PubMed] [Google Scholar]
- 23.Raghavan M et al. , Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans. Nature. 505, 87–91 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mittnik A et al. , The genetic prehistory of the Baltic Sea region. Nat. Commun 9, 442 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mathieson I et al. , The genomic history of southeastern Europe. Nature. 555, 197–203 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lipson M et al. , Parallel palaeogenomic transects reveal complex genetic history of early European farmers. Nature. 551, 368–372 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lazaridis I et al. , Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature. 513, 409–13 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gallego-Llorente M et al. , The genetics of an early Neolithic pastoralist from the Zagros, Iran. Sci. Rep 6, 31326 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.de B. Damgaard P et al. , 137 ancient human genomes from across the Eurasian steppes. Nature. 557, 369–374 (2018). [DOI] [PubMed] [Google Scholar]
- 30.de Barros Damgaard P et al. , The first horse herders and the impact of early Bronze Age steppe expansions into Asia. Science. 360, eaar7711 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Broushaki F et al. , Early Neolithic genomes from the eastern Fertile Crescent. Science. 353, 499–503 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lazaridis I et al. , Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature. 513, 409–413 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Patterson N et al. , Ancient admixture in human history. Genetics. 192, 1065–93 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Skoglund P et al. , Origins and genetic legacy of Neolithic farmers and hunter-gatherers in Europe. Science. 336, 466–469 (2012). [DOI] [PubMed] [Google Scholar]
- 35.Patterson N, Price AL, Reich D, Population Structure and Eigenanalysis. PLoS Genet 2, e190 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Galinsky KJ et al. , Fast Principal-Component Analysis Reveals Convergent Evolution of ADH1B in Europe and East Asia. Am. J. Hum. Genet 98, 456–472 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Alexander DH, Novembre J, Lange K, Fast model-based estimation of ancestry in unrelated individuals. Genome Res 19, 1655–1664 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.McNeill J, Pomeranz K, Eds., The Cambridge World History (Cambridge University Press, Cambridge, 2015). [Google Scholar]
- 39.Loh P-R et al. , Inferring admixture histories of human populations using linkage disequilibrium. Genetics. 193, 1233–54 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hinch AG et al. , The landscape of recombination in African Americans. Nature. 476, 170–175 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Petraglia MD, Allchin B, in The Evolution and History of Human Populations in South Asia (Springer Netherlands, Dordrecht, 2007), pp. 1–20. [Google Scholar]
- 42.Bellwood PS, Renfrew C, M. I. for Archaeological Research, Examining the Farming/language Dispersal Hypothesis (University of Cambridge, 2002), McDonald Institute monographs. [Google Scholar]
- 43.Daly KG et al. , Ancient goat genomes reveal mosaic domestication in the Fertile Crescent. Science. 361, 85–88 (2018). [DOI] [PubMed] [Google Scholar]
- 44.Ammerman A, Cavalli-Sforza LL, The Neolithic Transition and the Genetics of Populations in Europe (Princeton University Press, 1984). [Google Scholar]
- 45.Dupree L, Notes on Shortugai : an Harappan site in northern Afghanistan (Centre for the Study of the Civilization of Central Asia, Quaid-i-Azam University, Islamabad, Pakistan, 1981). [Google Scholar]
- 46.Minniti C, Sajjadi SMS, New data on non‐human primates from the ancient Near East: The recentdiscovery of a rhesus macaque burial at Shahr‐i Sokhta(Iran). Int J Osteoarchaeol. 2019;1–11. Int. J. Osteoarchaeol 1 (2019). [Google Scholar]
- 47.Vidale M, A “Priest King” at Shahr-i Sokhta? Archaeol. Res. Asia 15, 110–115 (2018). [Google Scholar]
- 48.Moorjani P et al. , A genetic method for dating ancient genomes provides a direct estimate of human generation interval in the last 45,000 years. Proc. Natl. Acad. Sci. U. S. A 113, 5652–7 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang C-C et al. , Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions. Nat. Commun 10, 590 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Frachetti MD, Multiregional Emergence of Mobile Pastoralism and Nonuniform Institutional Complexity across Eurasia. Curr. Anthropol 53, 2–38 (2012). [Google Scholar]
- 51.Frachetti MD, Pastoralist landscapes and social interaction in bronze age Eurasia (University of California Press, 2008). [Google Scholar]
- 52.Unterländer M et al. , Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat. Commun 8, 14615 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Moorjani P et al. , A genetic method for dating ancient genomes provides a direct estimate of human generation interval in the last 45,000 years. Proc. Natl. Acad. Sci. U. S. A 113, 5652–7 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Giosan L et al. , Fluvial landscapes of the Harappan civilization. Proc. Natl. Acad. Sci. U. S. A 109, 1688–1694 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mallick S et al. , The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature. 538, 201–206 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hellenthal G et al. , The Kalash Genetic Isolate? The Evidence for Recent Admixture. Am. J. Hum. Genet 98, 396–397 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Silva M et al. , A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals. BMC Evol. Biol 17, 88 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Olalde I et al. , The genomic history of the Iberian Peninsula over the past 8000 years. Science (80-. ). 363, 1230–1234 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ringe D, Warnow T, Taylor A, Indo‐European and Computational Cladistics. Trans. Philol. Soc 100, 59–129 (2002). [Google Scholar]
- 60.Lister DL et al. , Barley heads east: Genetic analyses reveal routes of spread through diverse Eurasian landscapes. PLoS One. 13, e0196652 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Costantini L, The first farmers in Western Pakistan: the evidence of the Neolithic agropastoral settlement of Mehrgarh. Pragdhara. 18, 167–178 (2008). [Google Scholar]
- 62.Bellwood P, in The Encyclopedia of Global Human Migration (Blackwell Publishing Ltd, Oxford, UK, 2013). [Google Scholar]
- 63.B. W. Fortson, Indo-European Language and Culture: An Introduction (Wiley, 2011), Blackwell Textbooks in Linguistics. [Google Scholar]
- 64.Parpola A, The Roots of Hinduism : the Early Aryans and the Indus Civilization (Oxford University Press, 2015). [Google Scholar]
- 65.Mahadevan I, Bhaskar MV, in Walking with the Unicorn: Social Organization and Material Culture in Ancient South Asia, Frenez D, Jamison G, Law R, Vidale M, Meadow RH, Eds. (Archeopress, Oxford, 2018), pp. 359–376. [Google Scholar]
- 66.Southworth F, in 7th ESCA Harvard-Kyoto Roundtable (2005).
- 67.Krishnamurti B, The Dravidian Languages (Cambridge University Press, Cambridge, 2003). [Google Scholar]
- 68.Anthony DW, Ringe D, The Indo-European Homeland from Linguistic and Archaeological Perspectives. Annu. Rev. Linguist 1, 199–219 (2015). [Google Scholar]
- 69.Olalde I et al. , Derived immune and ancestral pigmentation alleles in a 7,000-year-old Mesolithic European. Nature. 507, 225–228 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sirak KA et al. , A minimally-invasive method for sampling human petrous bones from the cranial base for ancient DNA analysis. Biotechniques. 62, 283–289 (2017). [DOI] [PubMed] [Google Scholar]
- 71.Rohland N, Glocke I, Aximu-Petri A, Meyer M, Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc 13, 2447–2461 (2018). [DOI] [PubMed] [Google Scholar]
- 72.Gansauge M-T et al. , Single-stranded DNA library preparation from highly degraded DNA using T4 DNA ligase. Nucleic Acids Res 45, e79 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Briggs AW, Heyn P, in Methods in molecular biology (Clifton, N.J.) (2012), vol. 840, pp. 143–154. [DOI] [PubMed] [Google Scholar]
- 74.Rohland N, Reich D, Cost-effective, high-throughput DNA sequencing libraries for multiplexed target capture. Genome Res 22, 939–946 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.DeAngelis MM, Wang DG, Hawkins TL, Solid-phase reversible immobilization for the isolation of PCR products. Nucleic Acids Res 23, 4742–4743 (1995). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Maricic T, Whitten M, Pääbo S, Multiplexed DNA Sequence Capture of Mitochondrial Genomes Using PCR Products. PLoS One. 5, e14004 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kircher M et al. , A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet 46, 310–5 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Auton A et al. , A global reference for human genetic variation. Nature. 526, 68–74 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Li H, Durbin R, Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics. 26, 589–595 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Douka K et al. , Direct radiocarbon dating and DNA analysis of the Darra-i-Kur (Afghanistan) human temporal bone. J. Hum. Evol 107, 86–93 (2017). [DOI] [PubMed] [Google Scholar]
- 81.Renaud G, Kircher M, Stenzel U, Kelso J, freeIbis: an efficient basecaller with calibrated quality scores for Illumina sequencers. Bioinformatics. 29, 1208–1209 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Renaud G, Stenzel U, Kelso J, leeHom: adaptor trimming and merging for Illumina sequencing reads. Nucleic Acids Res 42, e141 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Fu Q et al. , A Revised Timescale for Human Evolution Based on Ancient Mitochondrial Genomes. Curr. Biol 23, 553–559 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Korneliussen TS, Albrechtsen A, Nielsen R, ANGSD: Analysis of Next Generation Sequencing Data. BMC Bioinformatics. 15, 356 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.McClure SB, Puchol OG, Culleton BJ, Ams Dating of Human Bone from Cova De La Pastora: New Evidence of Ritual Continuity in the Prehistory of Eastern Spain. Radiocarbon. 52, 25–32 (2010). [Google Scholar]
- 86.Lohse JC, Culleton BJ, Black SL, Kennett DJ, A Precise Chronology of Middle to Late Holocene Bison Exploitation in the Far Southern Great Plains. J. Texas Archeol. Hist 1, 94–126 (2014). [Google Scholar]
- 87.van Klinken GJ, Bone Collagen Quality Indicators for Palaeodietary and Radiocarbon Measurements. J. Archaeol. Sci 26, 687–695 (1999). [Google Scholar]
- 88.Santos GM, Southon JR, Druffel-Rodriguez KC, Griffin S, Mazon M, Magnesium Perchlorate as an Alternative Water Trap in AMS Graphite Sample Preparation: A Report On Sample Preparation at Kccams at the University of California, Irvine. Radiocarbon. 46, 165–173 (2004). [Google Scholar]
- 89.Stuiver M, Polach HA, Discussion Reporting of 14C Data. Radiocarbon. 19, 355–363 (1977). [Google Scholar]
- 90.Reimer PJ et al. , IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years cal BP. Radiocarbon. 55, 1869–1887 (2013). [Google Scholar]
- 91.Chang CC et al. , Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience. 4, 7 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ramsey C. Bronk, Bayesian Analysis of Radiocarbon Dates. Radiocarbon. 51, 337–360 (2009). [Google Scholar]
- 93.Enshin DN, Skochina SN, Zakh VA, On settling rites in the Neolithic of the Low Ishim basin (basing on materials of Mergen 6 settlement). Bull. Archeol. Anthropol. Ethnogr 4, 43–52 (2012). [Google Scholar]
- 94.Enshin DN, Neolithic dwellings from settlements of the Mergen lake. Bull. Archeol. Anthropol. Ethnogr 1, 14–23 (2014). [Google Scholar]
- 95.Kosintsev PA, Nekrasov AE, in Ecology of ancient and modern societies. (Publishing House IPOS SB RAS, Tyumen, 1999), pp. 100–104. [Google Scholar]
- 96.Enshin DN, A pottery complex from the settlement of Mergen’ 7 (Low Ishim basin): description and interpretation. Bull. Archeol. Anthropol. Ethnogr 29, 15–27 (2015). [Google Scholar]
- 97.Skochina SN, Enshin DN, Bone inventory of the Neolithic burial of the settlement of Mergen 7. Br. reports Inst. Archeol, 242–251 (2017).
- 98.Larin OV, Afanasievo culture of Altai Mountains: burial ground Saldyar-1. Barnaul (2005).
- 99.Vadetskaya NF, E. B; Polyakov AV; Stepanova, Corpus of the Afanasyevo Culture sites. Azbuka, Barnaul, 301 (2014). [Google Scholar]
- 100.Gryaznov MP, Afanasievo Culture in the Yenisei Basin. Nauk. Saint-petersbg (1999).
- 101.Teploukhov SA, Classification of the ancient metal cultures of the Minusinks region. (1929), vol. IV (2). [Google Scholar]
- 102.Stepanova NF, On the question of the chronology of the Afanasievo culture of Gorny Altai. Archaeol. Stud. steppe Eurasia, 61–70 (2013).
- 103.Stepanova NF, The burial grounds of the Afanasievo culture of the surroundings of the village of Elo in the Altai Mountains (materials to the arch of monuments). Antiq. Sib. Cent. Asia. Collect. Sci. Pap 16, 8–26 (2012). [Google Scholar]
- 104.Abdulganeev MT, Posrednikov VA, Stepanova N, in Sources on the history of the Altai Republic (Gorno Altaisk, 1997), pp. 69–90. [Google Scholar]
- 105.Parzinger H, Nagler A, Leont’ev N, Zubkov V, The multi-period burial ground of Suchanicha near Minusinsk. On the position of the Tagar culture in the context of the Bronze Age and Iron Age culture in the Minusinsk Basin. Eurasia Antiq. 15, 67–208 (2009). [Google Scholar]
- 106.Dobeš M, Budinský P, Buchvaldek M, Muška J, The catalog of the Corded Ware in Bohemia V. Bílina region. Praehistorica. 17, 75–146 (1991). [Google Scholar]
- 107.Velemínský P, The skeleton from the grave 4/81 from Radovesice. Arch. Anthropol. Dep. Natl. Museum, Prague (1997).
- 108.Stloukal M, Eneolithic skeletons from Radovesice. Arch. Anthropol. Dep. Natl. Museum, Prague (1981).
- 109.Černý V, Anthropology of the Chalcolithic in Central Europe: variability of chronological, geographical and sexual dimorphism. PhD Dissertation. University of Bordeaux I; (1999). [Google Scholar]
- 110.Hanáková H, Stloukal M, Muška J, Anthropological findings from northwest Bohemia taken from older finds of the Regional Museum in Teplice. Proc. Natl. Museum Prague B33/3–4, 159–264 (1977).
- 111.Zápotocký M, Muška J, in Archaeological research in Northwest Bohemia in 1993–1997. (1999), pp. 7–43.
- 112.von Weinzierl R, The Neolithic settlement near Velké Žernoseky on the Elbe. Mitth. der Anthropol. Gesellschaft Wien 25, 29–49 (1895). [Google Scholar]
- 113.Epimakhov A, Early complex societies in the North of Central Eurasia (based on the materials of the burial ground Kamennyi Ambar-5 (Chelyabinsk Publishing House, Chelyabisnk, 2005), vol. 1. [Google Scholar]
- 114.Krause R, L. N. (Liudmila N. Koriakova, Multidisciplinary investigations of the Bronze Age settlements in the Southern Trans-Urals (Russia) (Habelt, 2013). [Google Scholar]
- 115.Evdokimov VV, Works of the Karagands Detachment (Nauka, Moscow, 1980). [Google Scholar]
- 116.Tkachev AA, Central Kazakhstan in the Bronze Age (Publishing House of the Tyumen Oil and Gas Institute, Tyumen, 2002). [Google Scholar]
- 117.Tkachev AA, Central Kazakhstan in the Bronze Age (Publishing House of the Tyumen Oil and Gas Institute, 2002). [Google Scholar]
- 118.Doumani PN et al. , Burial ritual, agriculture, and craft production among Bronze Age pastoralists at Tasbas (Kazakhstan). Archaeol. Res. Asia 1–2, 17–32 (2015). [Google Scholar]
- 119.Mar’Yashev AN, New Materials on the Settlements of the Bronze Epoch in the Bayan Zurek Mountains. Izvestiya. 1, 23–30 (2002). [Google Scholar]
- 120.Usmanova ER, Burial ground Lisakovsky-1: facts and parallels (TENGRI Ltd. Publ., Karaganda, Lisakovsk, 2005). [Google Scholar]
- 121.Epimakhov AV, Revisiting radiocarbon argumentation of the early dating of Alakul’ antiquities. Bull. Archeol. Anthropol. Ethnogr 34, 60–67 (2016). [Google Scholar]
- 122.Panyushkina IP, Mills BJ, Usmanova ER, Cheng L, Calendar Age of Lisakovsky Timbers Attributed to Andronovo Community of Bronze Age in Eurasia. Radiocarbon. 50, 459–469 (2008). [Google Scholar]
- 123.Kuzmina EE, Aryans – way to the South. (Moscow - St.Petersburg, Letniy sad, 2008). [Google Scholar]
- 124.Molodin VI, Epimakhov AV, Marchenko Zh V, Radiocarbon chronology of the Bronze Age of the Urals and south of the Western Siberia: Principles and Approaches, achievements and challenges. Bull. Novosib. State Univ. Ser. Hist. Philol, 136–167 (2014).
- 125.Kozintsev AG, Craniometric Evidence of the Early Caucasoid Migrations to Sibera and Eastern Central Asia, with Reference to the Indo-European Probem. Archaeol. Ethnol. Anthropol. Eurasia 37, 125–136 (2009). [Google Scholar]
- 126.Zubova DV, A. V; Chikisheva TA; Pozdnyakov, in The Aryans in the Eurasian steppes and neighboring territories: the Bronze and Early Iron Ages (Altai State University Press, Barnaul, 2014). [Google Scholar]
- 127.Zubova AV, Populations of Western Siberia of the II millennium BC according to anthropological data. (IAET SO RAN, Novosibirsk, 2014). [Google Scholar]
- 128.Leontyev NV, Archive of the N. M. Martyanov Minusinsk Regional Museum. Minusinsk. 1 (1989).
- 129.Fribus AV, Grushin SP, in Proceedings of the V (XXI) All-Russian Archaeological Congress in Barnaul (Barnaul, 2017), p. 370. [Google Scholar]
- 130.Kiselev SV, Ancient history of Southern Siberia (Nauka, Moscow, 1951). [Google Scholar]
- 131.Molodin VI, Baraba in the Bronze Age (Novosibirsk: Nauka, 1985). [Google Scholar]
- 132.Kushaev G, Unpublished Archive Report. Inst. Archaeol. Almaty, Kazakhstan (1983).
- 133.Isakov TM, A. I; Potemkina, The Cemetery of the Bronze Age Tribes in Tajikistan. Sov. Arkheologija 1 1, 145–167 (1989). [Google Scholar]
- 134.Potemkina TM, The Archeoastronomical Aspect in the Reconstruction of the Worldviews of Ancient Peoples. Anthropol. Archeol. Eurasia 45, 7–28 (2006). [Google Scholar]
- 135.Mar’yashev AN, Goryachev AA, About the Question of the Typology and Chronology of the Bronze Age Sites of Semirech’e. Russ. Archaeol, 5–20 (1993).
- 136.Mar’Yashev AA, A. N; Goryachev, Questions of periodization and chronology of the Bronze Age monuments шт Semirechye. Ross. Arkheologiya 1, 5–19 (1993). [Google Scholar]
- 137.Mar’yashev AN, “Unpublished Archive Report number 2172” (Almaty, Kazakhstan, 1988). [Google Scholar]
- 138.Mar’Yashev KM, A N; Karabaspakova, in Ancient Memorials of North Asia and their Guard excavations Novosibirsk, Nauka, Medvedev VE, Khudyakov YS, Eds. (Nauka, Novosibirsk, 1988), pp. 24–39. [Google Scholar]
- 139.Mar’yashev AN, “Unpublished Archive Report Number 2044” (Almaty, Kazakhstan, 1984). [Google Scholar]
- 140.Ivanov GP, Kashkarchinsky burial ground - a new monument of the late Bronze Age in Fergana. Soc. Sci. Uzb. Tashkent 10, 44–47 (1988). [Google Scholar]
- 141.Varfolomeev V, in Questions of archeology of Central and Northern Kazakhstan (Karaganda, 1989), pp. 76–84. [Google Scholar]
- 142.Tkachev AA, Chansharsky archaeological microdistrict in the Late Bronze Age of the Aktobe Urals. Ufa Archeol. Her 9, 72–83 (2009). [Google Scholar]
- 143.Arslanova FK, Essays in Medieval Archeology of the Upper Irtysh region. Arch. A.Kh. Margulan Inst. Archeol (2013).
- 144.Arslanova FK, in “In the depths of centuries”. Archeological collection, Akishev KA, Ed. (Nauka, Alma-Ata, 1974), pp. 46–60. [Google Scholar]
- 145.Arslanova FK, “Unpublished Archive Report 1175” (Almaty, Kazakhstan, 1969). [Google Scholar]
- 146.Doumani P, Bronze Age Potters in Regional Context: Long-Term Development of Ceramic Technology in the Eastern Eurasian Steppe Zone. Arts Sci. Electron. Theses Diss (2014), doi: 10.7936/K79S1P69. [DOI]
- 147.Spengler RN, Frachetti MD, Doumani PN, Late Bronze Age agriculture at Tasbas in the Dzhungar Mountains of eastern Kazakhstan. Quat. Int 348, 147–157 (2014). [Google Scholar]
- 148.Coon CS, The seven caves : archaeological explorations in the Middle East / by Carleton S. Coon (Knopf New York, [1st ed.]., 1957). [Google Scholar]
- 149.Coon CS, Cave explorations in Iran, 1949. (University Museum, University of Pennsylvania, Philadelphia, 1951), Museum monographs. [Google Scholar]
- 150.Gregg M, Thornton CP, A Preliminary Analysis of Prehistoric Pottery from Carleton Coon’s Excavations of Hotu and Belt Caves in Northern Iran: Implications for Future Research into the Emergence of Village Life in Western Central Asia. International Journal of the Humanities. 19, 56–94 (2012), [Google Scholar]
- 151.Libby WF, Radiocarbon dating. Univ. Chicago Press; Chicago, (1951). [Google Scholar]
- 152.Ralph EK, University of Pennsylvania Radiocarbon Dates I. Science (80-. ). 121, 149–151 (1955). [DOI] [PubMed] [Google Scholar]
- 153.Zeder MA, Hesse B, The initial domestication of goats (Capra hircus) in the Zagros mountains 10,000 years ago. Science. 287, 2254–7 (2000). [DOI] [PubMed] [Google Scholar]
- 154.Meiklejohn C, Merrett DC, Reich D, Pinhasi R, Direct dating of human skeletal material from Ganj Dareh, Early Neolithic of the Iranian Zagros. J. Archaeol. Sci. Reports 12, 165–172 (2017). [Google Scholar]
- 155.Merrett DC, Bioarchaeology in Early Neolithic Iran: assessment of health status and subsistence strategy (University of Manitoba, 2004). [Google Scholar]
- 156.Meiklejohn C, Agelarakis A, Akkermans PMMG, Smith PEL, Solecki R, Artificial cranial deformation in the Proto-neolithic and Neolithic Near East and its possible origin : Evidence from four sites. Paléorient. 18, 83–97 (1992). [Google Scholar]
- 157.Smith PEL, Architectural Innovation and Experimentation at Ganj Dareh, Iran. World Archaeol. 21 (1990), pp. 323–335. [Google Scholar]
- 158.Voigt MM, Meadow RH, Hasanlu, Volume I: Hajji Firuz Tepe, Iran--The Neolithic Settlement (University of Pennsylvania Press, 1983), Hasanlu excavation reports. [Google Scholar]
- 159.Spengler RN, Willcox G, Archaeobotanical results from Sarazm, Tajikistan, an Early Bronze Age Settlement on the edge: Agriculture and exchange. Environ. Archaeol 18, 211–221 (2013). [Google Scholar]
- 160.Lyonnet B, Isakov AE, Ceramics, (Chalcolithic and Bronze Age Ancient). Mem. French Archaeol. Mission Cent. Asia 7 (1996). [Google Scholar]
- 161.Avanessova NA, Dzhurakulova DM, in Proceeding of the International Conferrnece on The Culture of Nomads of Central Asia (2007), p. Pp. 13–33.
- 162.Rice DT, Excavations at Tepe Hissar, Damghan. By Erich F. Schmidt. With an additional chapter on the Sasanian building by Fiske Kimball. Philadelphia: University of Pennsylvania Press, 1937. pp. XXI, 478, 177 figs. and 79 plates. £3 7s 6d. Antiquity. 12, 494–495 (1938). [Google Scholar]
- 163.Gursan-Salzmann A, The new chronology of the Bronze Age settlement of Tepe Hissar, Iran (University of Pennsylvania Press, 2016). [Google Scholar]
- 164.Bobomulloyev S, Vinogradova NM, Bobomulloyev B, Excavations of the Middle Bronze Age burial ground of Farkhor in the south of Tajikistan. Zap. IIMK 11, 47–66 (2015). [Google Scholar]
- 165.Bobomulloyev S, Vinogradova NM, Bobomulloyev B, Monuments of the Middle Bronze Age in Southwest Tajikistan. Reports Acad. Sci. Repub. Tajikistan 3, 39–47 (2014). [Google Scholar]
- 166.Vinogradova NM, Monuments of the Middle Bronze Age in Southwest Tajikistan. VDI. 4, 84–98 (2011). [Google Scholar]
- 167.Vinogradova NM, Preliminary Results of the Excavations at the Necropolis of Farkhor, a Middle Bronze Age Site in Southern Tajikistan. Archeol. Rev. Anc. east. News Archaeol. Res 2, 21–27 (2015). [Google Scholar]
- 168.Tosi M, Excavations at Shahr-i Sokhta, a Chalcolithic Settlement in the Iranian Sīstān. Preliminary Report on the Second Campaign. East West, 283–386 (1969).
- 169.Tosi M, Excavations at Shahr-i Sokhta, a Chalcolithic Settlement in the Iranian Sīstān. Preliminary Report on the First Campaign, October-December. East West. 18 (1967), pp. 9–66. [Google Scholar]
- 170.Lamberg-Karlovsky CC, Tosi M, Shahr-i Sokhta and Tepe Yahya: Tracks on the Earliest History of the Iranian Plateau. East West. 23 (1973), pp. 21–57. [Google Scholar]
- 171.S. V.I, The Pottery of Shahr-i Sokhta I and its Southern Turkmenian Connections. In: Tosi M (ed.). Prehist. Sistan 1. IsMEO Reports Mem. 19.1. IsMEO, Rome 19.1, 183–198 (1983). [Google Scholar]
- 172.Bonora GL, Domani C, Salvatori S, Soldini A, The oldest graves of the Shahr-i Sokhta graveyard. South Asian Archeol. Isiao, Ser. Orient. Rome, 485–94 (1997).
- 173.Cortesi E, Tosi M, Lazzari A, Vidale M, Cultural Relationships beyond the Iranian Plateau: The Helmand Civilization, Baluchistan and the Indus Valley in the 3rd Millennium BCE. Paléorient. 34, 5–35 (2008). [Google Scholar]
- 174.Salvatori M, Tosi S, Shahr-i-Sokhta Revised Sequence. South Asian Archeol. 2001, Paris, ERC, 281–292.
- 175.Jarrige J-F, Didier A, Quivron G, Shahr-i Sokhta and the chronology of the Indo-Iranian regions. Paléorient. 37, 7–34 (2011). [Google Scholar]
- 176.Salvatori S, Vidale M, Shahr-i Sokhta 1975 – 1978: central quarters excavations ; preliminary report (IsIAo, 1997), Reports and memoirs. Series minor. [Google Scholar]
- 177.F. T. (Fredrik T. Hiebert, K. (Kakamurad) Kurbansakhatov, H. Schmidt, A Central Asian village at the dawn of civilization, excavations at Anau, Turkmenistan (University of Pennsylvania Museum of Archaeology and Anthropology, 2003). [Google Scholar]
- 178.Denisov EP, The Ksirov burial ground in the context of the funeral monuments of the nomads of North Bastria-Tokharistan (On the ethnic history of the North Bactria of the Kushan time)]. (Satvropol, 1999). [Google Scholar]
- 179.Vidale M, Micheli R, Protohistoric graveyards of the Swat Valley, Pakistan: new light on funerary practices and absolute chronology. Antiquity. 91, 389–405 (2017). [Google Scholar]
- 180.Vidale M, Micheli R, Olivieri LM, Excavations at the protohistoric graveyards of Gogdara and Udegram (ACT Field School Repors and Memoirs, III, Sang-e-Meel, Lahore, 2016). [Google Scholar]
- 181.Antonini C. Silvi, Stacul G, The proto-historic graveyards of Swāt (Pakistan). 1, Description of graves and finds (IsMEO Reports and Memoirs; (Rome: ) VII, 1972). [Google Scholar]
- 182.Stacul G, Preliminary Report on the Pre-Buddhist Necropolises in Swat (W. Pakistan). East West. 16 (1966), pp. 37–79. [Google Scholar]
- 183.Antonini CS, Preliminary Notes on the Excavation of the Necropolises found in Western Pakistan. East West. 14 (1964), pp. 13–26. [Google Scholar]
- 184.Tucci G, The Tombs of the Asvakayana-Assakenoi. East West. 14 (1964), pp. 27–28. [Google Scholar]
- 185.Vinogradova N, Towards the Question of the Relative Chronology for Protohistoric Swat Sequence (on the Basis of the Swat Graveyards). East West. 51, 9–36 (2001). [Google Scholar]
- 186.Salvatori S, Analysis of the Association of Types in the Protohistoric Graveyards of the Swāt Valley (Loebanr I, Kātelai I, Butkara II). East West. 25, 333–351 (1975). [Google Scholar]
- 187.Castaldi E, The graveyard of Katelai I in the Swat. Excavation report of the tombs 46–80. Accad. Naz. dei Lincei, Anno CCCLXV, Roma (1968).
- 188.Passarello P, Macchiarelli R, The proto-historic graveyard of Katelai (Swat, Pakistan): Anthropological analyses of skeletal material with reference to the paleobiological human context of the Middle East. Riv. di Antropol. LXV, 5–104 (1984). [Google Scholar]
- 189.Tucci G, Preliminary report on an archaeological survey in Swat. East West. 9 (1958), pp. 279–328. [Google Scholar]
- 190.Tusa S, Notes on Some Protohistoric Finds in the Swāt Valley (Pakistan). East West. 31 (1981), pp. 99–120. [Google Scholar]
- 191.Stacul G, Bīr-kōṭ-ghuṇḍai (Swāt, Pakistan) 1978 Excavation Report. East West. 30 (1980), pp. 55–65. [Google Scholar]
- 192.Stacul G, Excavation at Bīr-kōṭ-ghuṇḍai (Swāt, Pakistan). East West. 28 (1978), pp. 137–150. [Google Scholar]
- 193.Olivieri LM et al. , The last phases of the urban site of Bir-Kot-Ghwandai (Barikot) : The Buddhist sites of Gumbat and Amluk-Dara (Barikot) (ACT Reports and Memoirs, II, Sang-e-Meel, Lahore, 2014). [Google Scholar]
- 194.Olivieri LM, Iori E, Early-historic Data from the 2016 Excavation Campaigns at the Urban Site of Barikot, Swat (Pakistan): A Shifting Perspective. Proc. 29th South Asian Art Archeol. Conf. (2016). [Google Scholar]
- 195.Olivieri LM, Urban Defenses at Bīr-koṭ-ghwaṇḍai, Swat (Pakistan). Anc. Civilizations from Scythia to Sib 21, 183–199 (2015). [Google Scholar]
- 196.Olivieri LM, Marzaioli F, A new revised chronology and cultural sequence of the Swat valley, Khyber Pakhtunkhwa (Pakistan) in the light of current excavations at Barikot (Bir-kot-gwandai). Proc. 14th Int. Conf. AMS, Nucl. Instruments Methods Phys. Res. Sect. B Beam Interact. with Mater. Atoms (2017). [Google Scholar]
- 197.Stacul G, Tusa S, Report on the Excavations at Aligrāma (Swāt, Pakistan) 1966, 1972. East West. 25 (1975), pp. 291–321. [Google Scholar]
- 198.Stacul G, Tusa S, Report on the Excavations at Aligrāma (Swāt, Pakistan) 1974. East West. 27 (1977), pp. 151–205. [Google Scholar]
- 199.Zahir M, in A Companion to South Asia in the Past (John Wiley & Sons, Inc, Hoboken, NJ, 2016), pp. 274–293. [Google Scholar]
- 200.Alciati G, Human skeletal remains of the necropoli of the Swat (W. Pakistan). 1. Butkara II (IsMEO Reports and Memoirs; (Rome: ), VIII, 1, 1967). [Google Scholar]
- 201.Genna G, First Anthropological Investigations of the Skeletal Remains of the Necropolis of Butkara II (Swat, West Pakistan). East West. 15 (1965), pp. 161–167. [Google Scholar]
- 202.Fritsch CC, Butkara II Revisited. East West. 47, 41–66 (1997). [Google Scholar]
- 203.Pardini E, The Human Remains from Aligrāma Settlement (Swāt, Pakistan). East West. 27 (1977), pp. 207–226. [Google Scholar]
- 204.Callieri P, Saidu Sharif I (Swat, Pakistan).: The Buddhist sacred area : the Monastery (IsMEO Reports and Memoirs (Rome), XXIII,1, Rome, 1989). [Google Scholar]
- 205.Noci F, Faccenna D, Macchiarelli R, Saidu Sharif I (Swat, Pakistan) : The Graveyard, IsIao Reports and Memoirs (Rome) XXIII, 3, 1997. [Google Scholar]
- 206.Faccenna D, Saidu Sharif I (Swat, Pakistan). The Buddhist sacred area : the Stupa terrace (1995), IsMEO Reports and memoirs (Rome)XXIII, 2, 1995.
- 207.Olivieri LM et al. , The Graveyard and the Buddhist shrine at Saidu Sharif I (Swat, Pakistan): fresh chronological and stratigraphic evidence. J. Anc. Hist 76, 559–578 (2016). [Google Scholar]
- 208.Tucci G, On Swāt. The Dards and Connected Problems. East West. 27 (1977), pp. 9–103. [Google Scholar]
- 209.Olivieri LM, Afridi A, Iori E, Urban defenses at Bīr-koṭ-ghwaṇḍai, Swat (Pakistan). Data from the 2015 excavation campaign. Pakistan Herit. 7, 73–94 (2015). [Google Scholar]
- 210.Callieri P, Filigenzi A, Stacul G, Bir-Kot-Ghwandai, Swat: 1987 Excavation Campaign. Pakistan Archeol. 25, 183–192 (1990). [Google Scholar]
- 211.Callieri P et al. , Bīr-koṭ-ghwaṇḍai, Swat, Pakistan. 1998–1999 Excavation Report. East West. 50 (2000), pp. 191–226. [Google Scholar]
- 212.Bagnera A, The Ghaznavid Mosque and the Islamic Settlement at Mt. Rāja Gīrā, Udegram (ACT Reports and Memoirs, V, Sang-e-Meel, Lahore, 2015). [Google Scholar]
- 213.Chaubey G et al. , Population genetic structure in Indian Austroasiatic speakers: the role of landscape barriers and sex-specific admixture. Mol. Biol. Evol 28, 1013–24 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Basu A, Sarkar-Roy N, Majumder PP, Genomic reconstruction of the history of extant populations of India reveals five distinct ancestral components and a complex structure. Proc. Natl. Acad. Sci. U. S. A 113, 1594–9 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Narasimhan VM et al. , The Genomic Formation of South and Central Asia. bioRxiv, 292581 (2018).
- 216.Reich D et al. , Reconstructing Native American population history. Nature. 488, 370–374 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Lazaridis I et al. , Genetic origins of the Minoans and Mycenaeans. Nature. 548, 214 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Frenez D, Manufacturing and trade of Asian elephant ivory in Bronze Age Middle Asia. Evidence from Gonur Depe (Margiana, Turkmenistan). Archaeol. Res. Asia 15, 13–33 (2018). [Google Scholar]
- 219.Jarrige J-F, Didier A, Quivron G, Shahr-i Sokhta and the chronology of the Indo-Iranian regions. Paléorient. 37, 7–34 (2011). [Google Scholar]
- 220.Kunsch HR, The Jackknife and the Bootstrap for General Stationary Observations. Ann. Stat 17, 1217–1241 (1989). [Google Scholar]
- 221.Seguin-Orlando A et al. , Paleogenomics. Genomic structure in Europeans dating back at least 36,200 years. Science. 346, 1113–8 (2014). [DOI] [PubMed] [Google Scholar]
- 222.Metspalu M et al. , Shared and unique components of human population structure and genome-wide signals of positive selection in South Asia. Am. J. Hum. Genet 89, 731–44 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Chaubey G, Kadian A, Bala S, Rao VR, Genetic Affinity of the Bhil, Kol and Gond Mentioned in Epic Ramayana. PLoS One. 10, e0127655 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.O’Connell JF et al. , When did Homo sapiens first reach Southeast Asia and Sahul? Proc. Natl. Acad. Sci. U. S. A 115, 8482–8490 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Poznik GD et al. , Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences. Nat. Genet 48, 593–599 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Moorjani P et al. , The History of African Gene Flow into Southern Europeans, Levantines, and Jews. PLoS Genet. 7, e1001373 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Hellenthal G et al. , A genetic atlas of human admixture history. Science. 343, 747–751 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.1000 Genomes Project Consortium et al. , An integrated map of genetic variation from 1,092 human genomes. Nature. 491, 56–65 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.