

Abstract
Cancer stem cells (CSC) are a subpopulation of tumor cells with properties of high tumorigenicity and drug resistance, which lead to recurrence and poor prognosis. Although a better understanding of CSC is essential for developing cancer therapies, scarcity of the CSC population has hindered such analyses. The aim of the present study was to elucidate whether the E‐cadherin‐Fc chimera protein (E‐cad‐Fc) enhances cancer stem‐like properties because studies show that soluble E‐cadherin stimulates human epithelial growth factor receptor (EGFR) and downstream signaling pathways that are reported to play a crucial role in CSC. For this purpose, we used ornithine decarboxylase (ODC)‐degron–transduced (Degron(+)) KM12SM cells as a CSC model that retains relatively low CSC properties. Compared to cultures without E‐cad‐Fc treatment, we found that E‐cad‐Fc treatment further suppressed proteasome activity and largely enhanced cancer stem‐like properties of ODC‐degron–transduced KM12SM cells. These results include increased expression of stem cell markers Lgr5, Bmi‐1, SOX9, CD44, and CD44v9, aldehyde dehydrogenase (ALDH), and enhancement of robust spheroid formation, and chemoresistance to 5‐fluorouracil (5‐FU) and oxaliplatin (L‐OHP). These effects could be attributed to activation of the EGFR pathway as identified by extensive phosphorylation of EGFR, ERK, PI3K, AKT, and mTOR. In SW480 cells, E‐cad‐Fc matrix induced some CSC markers such as CD44v9 and ALDH. We also found that E‐cad‐Fc matrix showed high efficiency of inducing mesenchymal changes in colon cancer cells. Our data suggest that the E‐cad‐Fc matrix may enhance CSC properties such as enhancement of chemoresistance and sphere formation.
Keywords: cancer stem cell, colon cancer, E‐cadherin‐Fc, epithelial‐mesenchymal transition, extracellular matrix
1. INTRODUCTION
Epithelial‐mesenchymal transition (EMT) is a cellular process first discovered in embryogenesis1 and is increasingly recognized as the main factor in cancer progression.2, 3 Cancer cells with mesenchymal‐like phenotype present a loss of tight cell‐cell adhesion and show cellular polarity reversal4, 5 with enhanced expression of EMT‐activating transcription factors such as SNAIL, TWIST and ZEB families.6, 7 The process of EMT facilitates invasiveness and migration capability and is thus considered a trigger of cancer metastasis and recurrence.8, 9
Cumulative evidence suggests that EMT is linked to cancer stem cells (CSC). EMT process is considered to be involved in altering the microenvironment and regulating CSC initiation.3, 10, 11, 12 CSC undergo self‐renewal and produce differentiated cells by asymmetric division.13 Because CSC have properties such as drug resistance and high tumorigenicity, they are associated with cancer recurrence and poor prognosis.14 Decades of research have shown that the CSC niche microenvironment plays a pivotal role in their development,15 offering the possibility of supporting their stem cell status. Several studies have indicated that CSC stem cell properties could be enhanced through the extracellular matrix (ECM).16 For instance, endothelial‐derived ECM propagates the population of mesenchymal stem cells (MSC) by providing MSC with a perivascular niche.17 Cancer cells cultured on hepatocyte‐derived ECM show increased expression of stem cell markers Lgr5 and CD133.18 Additionally, feeder cells such as fibroblasts or laminin‐511–coated matrix support growth of human pluripotent embryonic stem cells (hPSC).19, 20 These findings indicate an essential role of the ECM in supporting stem cells by cell‐cell or cell‐matrix interactions.
Recently, E‐cadherin Fc chimera protein (E‐cad‐Fc) has been noted as cell culture material for stem cell maintenance.21 E‐cadherin is an epithelial cell‐cell adhesion molecule that is frequently downregulated in many cancers. Its expression is inversely correlated with EMT.22 Culturing with the E‐cad‐Fc matrix satisfactorily retains the undifferentiated state of embryonic stem (ES) cells and maintains isolation of adhered cells without colony formation.23 It is notable that E‐cad‐Fc matrix promotes adhesion and proliferation of MSC compared with Matrigel.24 Studies have also shown that cell adherence and junctions between cancer cells are disturbed by addition of the soluble fragment of E‐cadherin, leading to malignancy in skin squamous cell carcinoma and breast cancer, possibly by activation of EGFR and its downstream signaling pathways.25, 26, 27
Based on the above findings, in the present study, we hypothesized that the E‐cad‐Fc matrix might serve as potential material for enhancement or maintenance of CSC properties. In parallel, we investigated the effect of E‐cad‐Fc matrix on EMT. For this purpose, we used colon cancer SW480 cells because the cells readily undergo EMT with transforming growth factor‐beta (TGF‐β)1 and EGF cocktail.28, 29, 30 We also used ornithine decarboxylase (ODC)‐degron–transduced KM12SM colon cancer cells that show relatively weak CSC properties in this system. Low proteasome activity (LPA) is considered a hallmark of CSC in human cancers, including gastrointestinal cancer,31, 32 cervical cancer,33 and osteosarcoma.34 By using the ZsGreen‐labeled degronODC (Gdeg) proteasome reporter system, we and other groups showed that compared with non‐LPA cells, colon and pancreatic cancer cells with LPA showed strong stem cell potential, enhanced chemo‐ or radio‐resistance, and upregulated expression of stem cell markers such as CD44, CD133, or ALDH.32
2. MATERIALS AND METHODS
2.1. Cell culture
KM12SM cells acquire highly metastatic ability through mouse serial transplantation of primary colon cancer KM12C cells.35 KM12SM was a kind gift from Professor T. Minamoto (Cancer Research Institute, Kanazawa University, Ishikawa Japan). Human colon cancer cell line SW480 was purchased from ATCC. Cells were maintained in DMEM supplemented with 10% FBS and Myco‐Zap Plus CL (Lonza) at 37°C in a humidified 5% CO2 atmosphere. The E‐cad‐Fc fusion protein was purchased from Somar Corporation, Ltd and partially donated by Prof. T. Akaike (FAIS, Ibaraki, Japan).
2.2. Epithelial‐mesenchymal transition induction
Epithelial‐mesenchymal transition induction assay was conducted according to a previous study.28 Briefly, after cells were seeded, they were treated with TGF‐β1 (2.5 ng/mL; Sigma‐Aldrich) and epithelial growth factor (EGF) (10 ng/mL; Sigma‐Aldrich) for 48 hours. To form the E‐cad‐Fc matrix, culture dishes were coated with the E‐cad‐Fc fusion protein at a concentration of 10 μg/mL for 2 hours at room temperature. Dishes were washed gently with PBS twice, then cells were seeded and grown for the indicated time.
2.3. Transduction of the degron reporter
The degron sequence of ODC is recognized directly by proteasomes, which leads to immediate destruction of the involved protein. The retroviral expression vector pQCXIN‐ZsGreen‐cODC, containing green fluorescence Gdeg, was kindly provided by Dr Frank Pajonk (Jonsson Comprehensive Cancer Center, UCLA, CA, USA) to S. Tanaka. The vector was transfected into platinum retroviral packaging cells, and the retrovirus collected from the supernatant was used to infect KM12SM cells. Stable transfectants were selected with G418 solution (Roche). The top 0.86% of cell populations of the EGFP channel were enriched by sorting three times using flow cytometry (Cell Sorter SH800; SONY) and maintained in 0.1 mg/mL G418 solution.
2.4. Cell morphology
Cells were cultured in polystyrene tissue culture (TC) dishes with a TC‐treated surface (IWAKI) or in E‐cad‐Fc coated dishes. After treatment for 24, 48, and 72 hours, microscopy observation was carried out using BZ‐X700 (Keyence).
2.5. Western blot analysis
Cell lysates were extracted by RIPA buffer with 1% Halt Protease Inhibitor Cocktail Kit (Thermo Fisher Scientific). For detection of phosphorylation, 1% Halt Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific) was added to the lysates. Cell extracts were loaded onto Mini‐Protean TGX 4%‐15% gels (Bio‐Rad) and transferred using the Trans‐Blot Turbo Blotting System (Bio‐Rad). Primary antibodies used in this study are shown in Table S1. Secondary antibodies were incubated with ECL substrate (Bio‐Rad), and bands were visualized using the ChemiDoc Touch Imaging System (Bio‐Rad). Images were processed with Image Lab 5.2.1 software.
2.6. RNA extraction and qRT‐PCR
Total RNA was collected from cultured cells using TRIzol Reagent (Thermo Fisher Scientific), and complementary DNA was synthesized from 1.0 μg total RNA using oligo dT primer and a Reverse Transcription System (Promega) according to the manufacturer's instructions. Quantitative real‐time PCR (q‐PCR) was carried out using LightCycler FastStart DNA Master SYBR Green I (Roche) or LightCycler TaqMan Master (Roche) on a LightCycler 2.0 II (Roche).
Expression of the target gene was normalized relative to β2M mRNA expression using the 2−ΔΔCt method. Primers are shown in Table S2.
2.7. Flow cytometric analysis
Cells were suspended in FluoroBrite DMEM (Thermo Fisher Scientific) with 10% FBS. For analysis of CSC markers, rat antihuman CD44v/RV3 (LKG‐M001; Cosmo Bio Co., Ltd) was used as the primary antibody and phycoerythrin (PE)‐conjugated mouse antirat IgG2a (Becton Dickinson) as the secondary antibody. BV421‐conjugated mouse antihuman CD44 (Becton Dickinson) and PE‐conjugated mouse antihuman CD24 (Becton Dickinson) were used. Data were collected from three independent experiments. Stem cell marker ALDH was detected using ALDEFLUOR Kit (STEMCELL Technologies) according to the manufacturer's protocol. All samples included a test group and negative control treated with ALDH inhibitor, diethylaminobenzaldehyde (DEAB). Immunofluorescent intensity was measured by flow cytometry.
2.8. Spheroid culture
Serum‐free DMEM/F12 medium was added to ultra‐low attachment 96‐well plates. Cells were then suspended in DMEM and added to wells at a density of 1000 cells per well. In the E‐cad‐Fc–treated group, cells were transferred to ultra‐low attachment plates 48 hours after incubation. Cells were cultured for 18 days. Images were acquired digitally using BX‐Z analysis software.
2.9. Proliferation assay
Cells were suspended with DMEM, and 5000 cells of each group were plated into a 96‐well plate. Medium was changed every 2 days. Cell proliferation was measured every 24 hours with CCK‐8 (Dojindo) according to the manufacturer's protocol. Absorbance (450 nm) was measured using a spectrophotometer.
2.10. Chemosensitivity assay
A total of 8000 cells per group was seeded onto a 96‐well plate and treated with chemotherapeutic drugs at concentrations of 0.5‐256 μmol/L for 5‐FU and 0.25‐100 μmol/L for oxaliplatin (L‐OHP). Cells were treated for 72 hours, then measured with CCK‐8 (Dojindo).
2.11. Proteasome activity assay
One million cells of each group were suspended with 500 μL of 0.5% NP‐40, then centrifuged at 15 300 g for 15 minutes at 4°C to remove insoluble cellular debris. Cell lysates were collected to detect proteasome activity according to the manufacturer's protocol (Abcam). Each reaction well (96‐well plate [opaque white; Thermo Fisher Scientific]) was mixed with 30 μL sample lysate and 70 μL assay buffer provided with the kit, and negative control cell lysate was measured in the presence of proteasome inhibitor. Proteasome activity (mU) per one million cells was calculated.
2.12. Statistical analysis
Data are presented as mean ± SEM. Statistical significance of differences was calculated using one‐way ANOVA followed by Bonferroni's correction for multiple comparisons. Statistical analyses were carried out with GraphPad Prism version 6.00 for Mac (GraphPad Software). P < .05 was considered to indicate significance.
3. RESULTS
3.1. Effects of E‐cad‐Fc on EMT and cancer stem cell markers in SW480 colon cancer cells
We first examined EMT‐related changes in morphology and related molecular events in SW480 cells. Because EGF plus TGF‐β1 cocktail is widely recognized as an EMT inducer, we used this treatment as a positive control. We found that compared with control cells, both E‐cad‐Fc matrix–treated cells and EGF + TGF‐β1–treated cells showed spindle‐like morphology, a hallmark of EMT and this change became evident as the time passed (Figure 1A). Western blot analyses showed that EGF + TGF‐β1–treated cells showed increased expression of mesenchymal markers, N‐cadherin, Snail1, and Vimentin to some extent (Figure 1B). Because SW480 cells expressed a relatively low level of E‐cadherin, apparent reduction in E‐cadherin was not noted by E‐cad‐Fc treatment. Quantitative analysis of band intensity in three independent experiments indicated that N‐cadherin protein expression was significantly upregulated in the E‐cad‐Fc–treated group (P < .05, Figure 1B). RT‐PCR assay in two independent experiments indicated concordant results to those obtained by western blotting. Thus, N‐cadherin mRNA was significantly upregulated by E‐cad‐Fc treatment (P < .001, Figure 1C).
Figure 1.
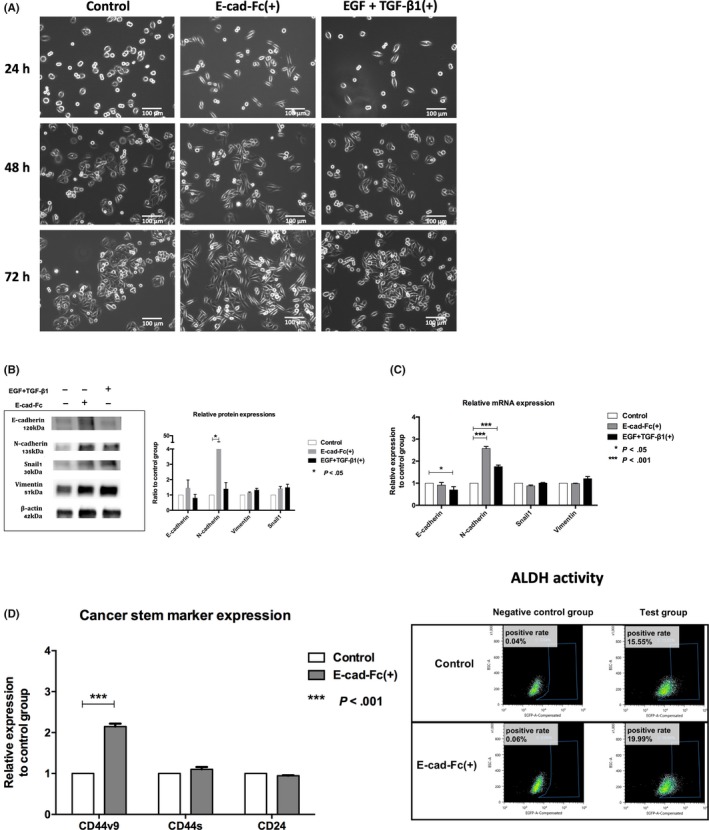
Effects of E‐cadherin‐Fc chimera protein (E‐cad‐Fc) on epithelial‐mesenchymal transition (EMT) and cancer stem cell markers in SW480 colon cancer cells. SW480 cells were divided into three groups, control group, E‐cad‐Fc(+) group, and epithelial growth factor (EGF) + transforming growth factor (TGF)β1(+) group. A, Time course study of cell morphology (24, 48, and 72 h). Cells were cultured on normal tissue culture plate (left), cultured on a non‐treated plate coated with E‐cad‐Fc matrix (middle), or cultured on normal tissue culture plate with medium supplemented by EGF and TGF‐β1 (right). Mesenchymal‐like cells that showed a spindle‐like shape appeared as time passed in the E‐cad‐Fc(+) and EGF + TGF‐β1(+) groups. B, Expressions of epithelial or mesenchymal markers at the protein level when treated for 48 h with E‐cad‐Fc matrix or EGF and TGF‐β1. Quantitative analysis in three independent experiments is shown in the right panel. N‐cadherin expression was significantly upregulated in the E‐cad‐Fc–treated group (*P < .05). C, Expression of epithelial or mesenchymal markers at the mRNA level was detected after 24 h treatment using quantitative real‐time PCR assay. Expression of the target gene was normalized relative to β2M mRNA expression using the 2−ΔΔCt method. Data were collected from two independent experiments. *P < .05, ***P < .001. D, Expression of cancer stem cell markers. Left panel: CD44v9, CD44s, and CD24 were detected by immunostaining using flow cytometry after 48 h treatment. Results are shown from two independent experiments as a ratio of immunofluorescence intensity of Degron(+) E‐cad‐Fc(+) group to that of Degron(−) control group. ***P < .001. Right panel: Stem cell marker aldehyde dehydrogenase (ALDH) expression. Cells of each group were divided into test group and negative control group blocked by DEAB (ALDH inhibitor). ALDH positive rate: Degron(−) control, 15.55%; Degron(+) E‐cad‐Fc(+), 19.99%
We next examined expression of CSC‐associated cell surface markers including CD44v9, CD44s,36, 37 and CD24.38 Flow cytometric analysis showed that the E‐cad‐Fc–treated group showed an increase in CD44v9 expression (P < .001) and ALDH activity (from 15.55% to 19.99%), another hallmark of CSC 39 (Figure 1D).
3.2. Effects of E‐cad‐Fc on EMT and cancer stem cell markers in ODC‐degron–transduced KM12SM cells
As the second cell line, we used ODC‐degron–transduced KM12SM cells because these cells acquire insufficient CSC properties even with the ODC‐degron system. Time‐course study showed that only the Degron(+) E‐cad‐Fc(+) group showed spindle‐like morphology (Figure 2A). Western blot analysis indicated that the Degron(+) E‐cad‐Fc(+) group, compared to the other groups, highly expressed mesenchymal markers, including N‐cadherin, Vimentin, ZEB1, Snail1, Slug, and Twist‐1 while E‐cadherin expression was modestly reduced (Figure 2B). Notably, both latent (45 kDa) and active (26 kDa) forms of TGF‐β1 increased in the Degron(+) E‐cad‐Fc(+) group. Quantitative analysis in two independent experiments indicated that there was >twofold increase in protein expression of Vimentin, Snail1, Twist1, TGF‐β1 in the Degron(+) E‐cad‐Fc(+) group (Figure 2B). These changes of mesenchymal markers were also noted in the Degron(−) E‐cad‐Fc(+) group (Figure S1). The RNA study showed concordant results except for vimentin and TGF‐ β1 (Figure 2C). Time‐course study showed that Vimentin mRNA increased at 48 hours, but TGF‐β1 mRNA did not increase throughout the time point examined (Figure S2).
Figure 2.
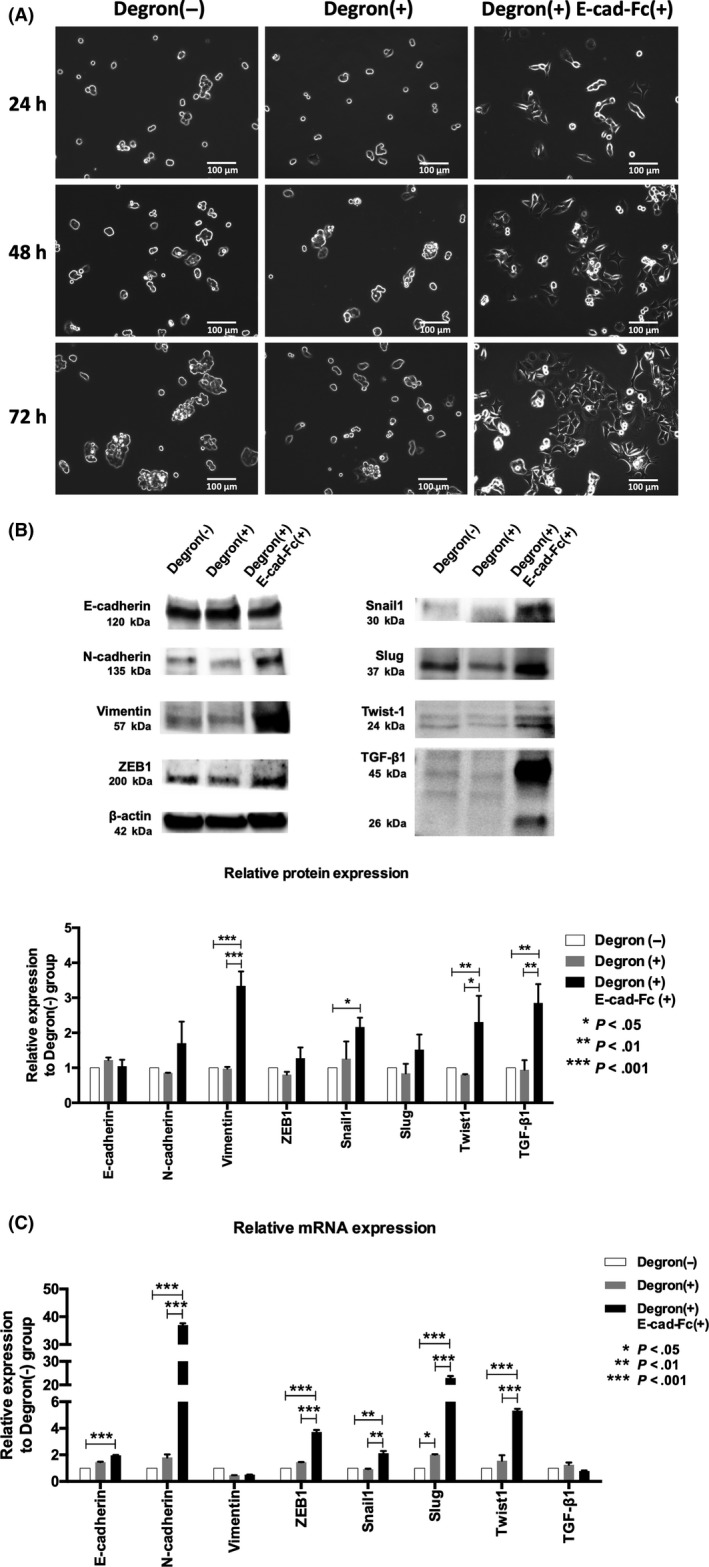
Mesenchymal‐like characteristics in human colon cancer cell line KM12SM. A, Cells were cultured for 24, 48, and 72 h. Degron(−) group was cultured on normal tissue culture plate, Degron(+) group was cultured on normal tissue culture plate, and Degron(+) E‐cad‐Fc(+) group was cultured on a non‐treated plate coated with E‐cad‐Fc matrix. Degron(+) E‐cad‐Fc(+) group cells showed a spindle‐like shape as time passed. B, Degron(+) E‐cad‐Fc(+) group showed upregulation of mesenchymal markers at the protein level after 48 h treatment. Latent (45 kDa) and active (26 kDa) forms of transforming growth factor (TGF)‐β1 protein also increased. Quantitative analysis in two independent experiments are shown. C, RNA study was done after 24 h treatment. *P < .05, **P < .01, ***P < .001. E‐cad‐Fc, E‐cadherin‐Fc chimera protein
We then examined the expression of the CSC markers Lgr5, Bmi‐1,40, 41 and SOX942 by RT‐PCR. ODC‐degron–transduced cells increased SOX9 mRNA expression (Figure 3A; P < .05), whereas Degron(+) E‐cad‐Fc(+) cells showed a significant increase in Lgr5, Bmi‐1, and SOX9 mRNA expression compared with the Degron(−) control cells (Figure 3A; *P < .05, **P < .01). Flow cytometric analysis showed that the Degron(+) group showed no increase in CSC surface markers; however, the Degron(+) E‐cad‐Fc(+) group had significantly higher expression of CD44v9 and CD44s (but not CD24) (Figure 3B, left panel, *P < .05, **P < .01). Flow cytometric analysis indicated that ALDH activity increased from Degron(−) control cells to ODC‐degron–transduced cells to Degron(+) E‐cad‐Fc(+) cells in a stepwise method (6.79% ~ 11.65% ~ 22.50%; Figure 3B, right panel).
Figure 3.
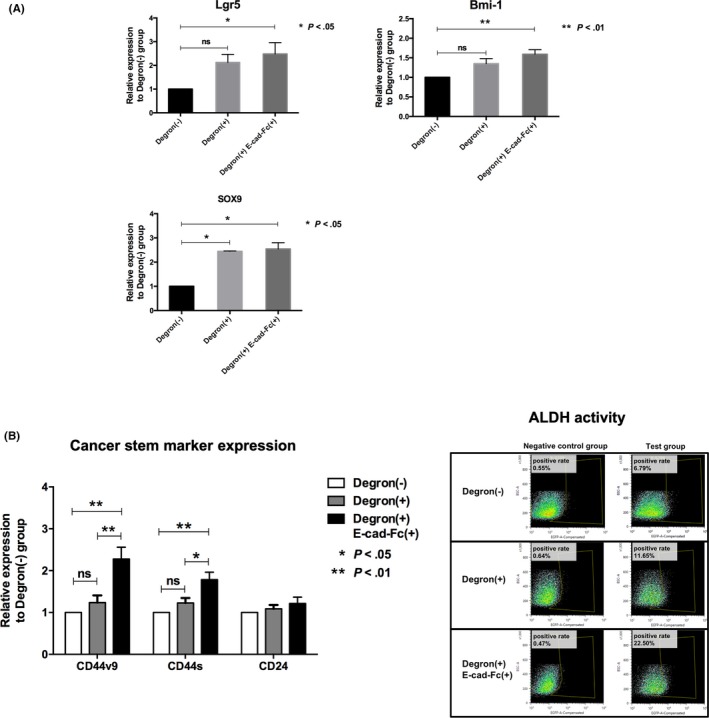
Stem cell marker expressions. A, Quantitative real‐time PCR analysis. Results are shown as a ratio of Degron(−) group to Degron(+) or Degron(+) E‐cad‐Fc(+) group. Data were collected from three independent experiments. *P < .05, **P < .01. B, Left panel: Cancer stem cell markers CD44v9, CD44s, and CD24 were detected by immunostaining using flow cytometry. Results are shown as a ratio of immunofluorescence intensity of Degron(−) group to Degron(+) or Degron(+) E‐cad‐Fc(+) group. Data were collected from three independent experiments. *P < .05, **P < .01. Right panel: Stem cell marker aldehyde dehydrogenase (ALDH) expression of Degron(−), Degron(+) and Degron(+) E‐cad‐Fc(+) was detected by flow cytometry. ALDH positive rate: Degron(−), 6.79%; Degron(+), 11.65%; Degron(+) E‐cad‐Fc(+), 22.50%. E‐cad‐Fc, E‐cadherin‐Fc chimera protein
3.3. E‐cadherin‐Fc chimera protein enhanced stem cell properties
To investigate whether the E‐cad‐Fc matrix increased stem cell properties, we carried out a sphere‐formation assay. When we assessed the number of spheres with a diameter ≥100 μm, the Degron(+) E‐cad‐Fc(+) group, but not the Degron(+) group, showed significantly increased sphere formation compared with the Degron(−) control cells (Figure 4A, B‐a; *P < .05). Detailed observation under microscopy further showed that the Degron(+) E‐cad‐Fc(+) group had a higher potential to form smooth‐edged, robust spheroids and presented fewer rough‐edged and disrupted spheroids compared to the Degron(+) group (Figure 4A, B‐b; *P < .05, **P < .01).
Figure 4.
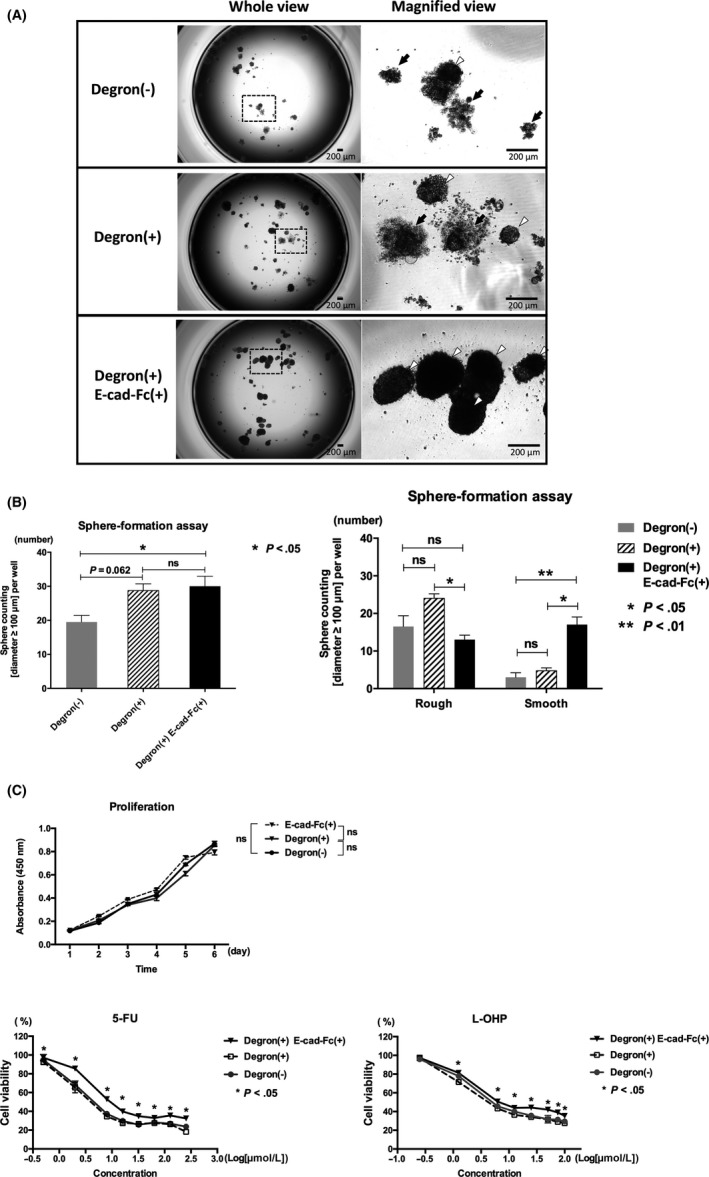
Stem cell properties endowed by E‐cadherin‐Fc chimera protein (E‐cad‐Fc). A, Sphere formation of each group seeded at a density of 1000 cells per well cultured for 18 days. In the Degron(+) E‐cad‐Fc(+) group, cells were cultured with E‐cad‐Fc–coated plates for 48 h and then transferred to ultra‐low attachment plates. Spheroid morphology is shown in whole view (left) and magnified view (right). Rough‐edged spheres are marked by black arrows, and smooth spheres by white arrowheads. Scale bar, 200 μm. B‐a, Sphere size ≥100 μm in diameter was counted to assess sphere‐formation ability. *P < .05. B‐b, Sphere size ≥100 μm in diameter was counted to assess sphere‐formation ability, and smooth‐edged spheres and rough‐edged spheres were separately counted. *P < .05, **P < .01. C‐a, Proliferation assay and chemoresistance assay. In the Degron(+) E‐cad‐Fc(+) group, cells were cultured with E‐cad‐Fc–coated plates for 48 h and then transferred to plates at a density of 5000 cells per well, and cultured with DMEM supplemented with 10% FBS. Proliferation was measured by CCK‐8 every 24 h. C‐b, Cells of each group were treated with fluorouracil (5‐FU) or oxaliplatin (L‐OHP) for 72 h. After drug treatment, cell survival rates were measured. *P < .05
We then examined drug resistance to the anticancer agents 5‐FU and oxaliplatin (L‐OHP). Proliferative activity did not differ among the three groups (Figure 4C‐a), and the Degron(+) E‐cad‐Fc(+) group alone showed chemoresistance compared with Degron(−) control cells (*P < .05 for both drugs; Figure 4C‐b). IC50 value of 5‐FU against Degron(−) control cells and Degron(+) E‐cad‐Fc(+) group was 7.10 μmol/L and 17.95 μmol/L, respectively, and the IC50 value of L‐OHP against Degron(−) control cells and Degron(+) E‐cad‐Fc(+) group was 9.46 and 16.84 μmol/L, respectively.
3.4. E‐cadherin‐Fc chimera protein activated the EGFR signaling pathway
It is reported that soluble E‐cadherin binds to EGFR and activates PI3K/AKT/mTOR signaling which is one of the major mechanisms involved in cancer stemness as well as the Wnt signaling pathway.43, 44, 45 Therefore, we attempted to investigate whether a E‐cad‐Fc coating would enhance this pathway. In the Degron(+) group, as shown in Figure 5, quantitative analysis in two independent western blotting experiments indicated that the EGFR activity was significantly upregulated through phosphorylation, and AKT, ERK1/2 and mTOR activities were also increased to some extent. Notably E‐cad‐Fc treatment of ODC‐degron‐transduced cells further enhanced phosphorylation of EGFR, AKT, ERK1/2, mTOR, PI3K (*P < .05, ** P < .01, *** P < .001) although the total expression of each molecule was not affected.
Figure 5.
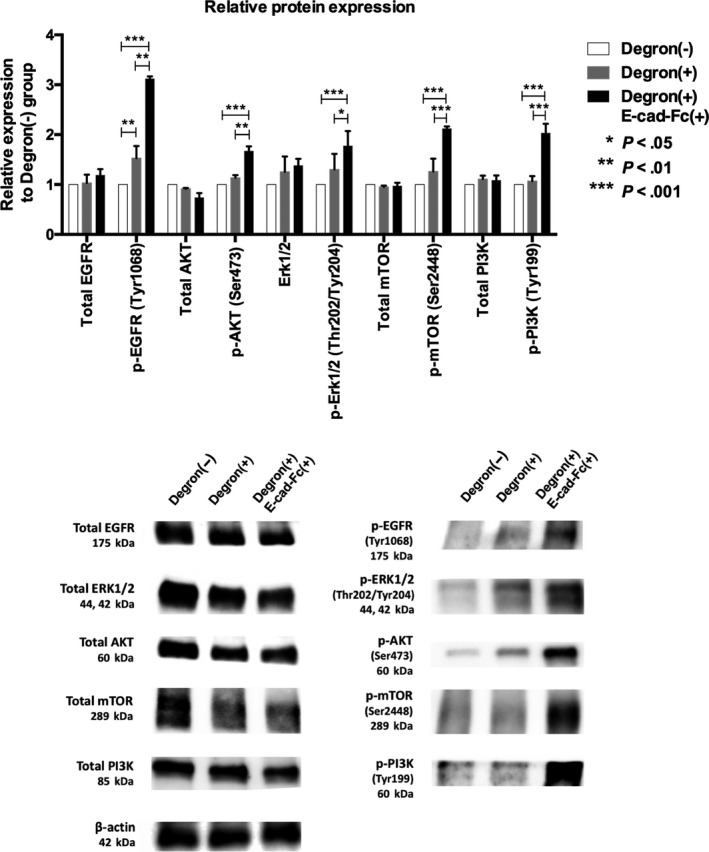
Activation of the epithelial growth factor receptor (EGFR) signaling pathway with E‐cadherin‐Fc chimera protein (E‐cad‐Fc) treatment. Upper panel: Quantitative analysis in two independent western blotting experiments. E‐cad‐Fc treatment of ornithine decarboxylase‐degron–transduced cells markedly enhanced phosphorylation of EGFR, AKT, ERK1/2, mTOR, and PI3K, although total expression of each molecule was not affected. *P < .05, **P < .01, ***P < .001. Lower panel: Representative blots are shown
3.5. E‐cadherin‐Fc chimera protein matrix suppressed proteasome activity
Because earlier studies showed that LPA is a hallmark of CSC,31, 32, 33, 34 we measured proteasome activity in KM12SM cells. Our results showed that the Degron(+) group significantly inhibited proteasome activity compared to Degron(−) control cells (Figure 6; *P < 0.05, **P < .001). We also found that the proteasome activity tended to be further suppressed when Degron(+) cells were cultured on the E‐cad‐Fc matrix (Degron(+) E‐cad‐Fc(+)) compared with the Degron(+) group (Figure 6, P = .068).
Figure 6.
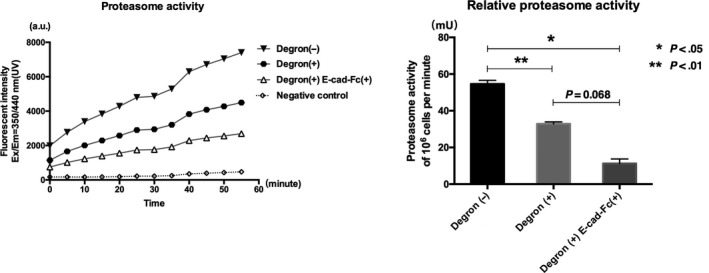
Proteasome activity measured in cell lysates. Proteasome activity was observed every 5 min until 55 min. Negative control was measured in the presence of proteasome inhibitor. Proteasome activity (mU) per one million cells was determined. *P < .05, **P < .01
4. DISCUSSION
Despite rigorous studies, an effective treatment targeting CSC has still not been established. One reason is that CSC are relatively rare among tumor cells and readily lose their properties through dilution with rapidly expanding differentiated daughter cells.46 This scarcity hinders analyses involving CSC. One solution is development of tools that enhance CSC properties and allow for efficient analysis of these cells.
Homeostatic proteasome activity plays a critical role in regulating the physiological activities of cancer cells and contributes to CSC initiation and development.47, 48 During experiments, we were aware that the ODC‐degron system worked well to show CSC properties in some cells such as colon cancer cell lines HCT116 and DLD132 and pancreatic cancer cell line Panc‐1,31 but did not work in other cancer cell types. Therefore, it is assumed that the success of the ODC‐degron system may depend on the degree to which LPA in the cell affects CSC properties. In the present study, we used KM12SM cells because ODC‐degron–transduced KM12SM cells did not acquire sufficient CSC properties as shown in the present study. Therefore, we viewed these cells as suitable for evaluating an enhanced effect of cancer stemness.
We showed that culturing with E‐cad‐Fc–coated plates raised CD44v9 expression significantly and ALDH expression to some extent in SW480 cells and enhanced various CSC‐related molecular in the ODC‐degron–transduced KM12SM cells. In the latter cells, Lgr5, Bmi‐1, and SOX9 mRNA were increased by E‐cad‐Fc treatment. They are reported to play an important role in regulating Wnt signaling and are associated with stem cell maintenance.49, 50, 51 Cell surface markers for CSC, CD44v9 and CD44s but not CD24 were also increased. These changes could lead to cancer stem‐like properties such as sphere formation and chemoresistance to 5‐FU and L‐OHP. In sphere‐formation assays, Degron(+) E‐cad‐Fc(+) cells showed enhanced formation of smooth and round robust spheres. This finding is consistent with a report by Frankel et al52 showing that smooth and round spheroids have a higher spheroidal quality, whereas rough spheroids begin to disintegrate and lose cell junctions. Tumorigenicity assay is a valid method to estimate CSC properties, but we could not judge the results because the ODC‐degron–transduced KM12SM cells alone readily accomplished in vivo tumor formation (6/6 by injection of either 104 or 105 cells, our unpublished observations by HY, YQ).
Low proteasome activity and EGFR signaling are both considered important to exert cancer stem properties31, 32, 33, 34, 43, 44, 45 and they could be the possible mechanism of how E‐cad‐Fc facilitates cancer stemness. It is reported that soluble E‐cadherin fragment can bind to EGFR and activate the PI3K/AKT/mTOR pathway in breast and skin cancer cells.26, 27 Conversely it was also shown that an antibody against the ectodomain of E‐cadherin selectively reduces tumor growth by downregulating EGFR and components of MAPK‐PI3K/AKT/mTOR.53 Mechanistic studies indicated that ODC‐degron–transduced KM12SM cells indeed showed low proteasome activity and partially activated the EGFR downstream pathway, which was insufficient to induce a variety of CSC cell surface markers and chemoresistance, although it may contribute to some increase in SOX9 mRNA and ALDH expression (Figure 3A,B). In contrast, when ODC‐degron–transduced KM12SM cells were cultured on E‐cad‐Fc–coated plates, proteasome activity was further lowered and the EGFR signaling pathway was markedly activated as identified by intense phosphorylation of the constituent molecules. It is assumed that these drastic changes led to induction of the broad range of CSC markers (Lgr5, Bmi‐1, SOX9, CD44v9, CD44s, and ALDH), increased chemoresistance to 5‐FU and oxaliplatin and smooth‐edged, robust spheroid formation. These findings suggest that E‐cad‐Fc, like soluble E‐cadherin, could affect EGFR and enhanced cancer stem‐like properties.
During the experiments, we found that E‐cad‐Fc–treated cells had a spindle‐like cell morphology, a characteristic of EMT. Analyses at protein and RNA levels showed the underlying molecular events. Thus, N‐cadherin was highly induced by E‐cad‐Fc in SW480 cells. In particular, KM12SM cells showed a large increase in a variety of mesenchymal markers with E‐cad‐Fc treatment although E‐cadherin, an epithelial marker, was not reduced much (Figure 2B). KM12SM cells acquire highly metastatic ability through mouse serial transplantation of primary colon cancer cells.35 Therefore, we postulate that this cell line may have potential to transform in the mesenchymal direction. RT‐PCR analyses indicated that most mesenchymal molecules were upregulated by E‐cad‐Fc at the mRNA level. However, TGF‐β1, a typical inducer of EMT, did not increase at the mRNA level whereas both latent and active forms of TGF‐β1 were highly induced at the protein level, suggesting the existence of certain post‐translational mechanisms.
Several studies have shown that low proteasome activity enhances EMT through stabilization of the EMT‐related transcription factor Snail1 in gastric cancer and mammary epithelial cells.54, 55 However, this is not likely to be the current case because ODC‐degron–transduced cells without E‐cad‐Fc treatment showed no change in EMT‐related protein expression despite significant downregulation of proteasome activity. However, considering that various mesenchymal markers were similarly induced by E‐cad‐Fc even in control KM12SM cells (Figure S1), it is assumed that E‐cad‐Fc directly facilitates mesenchymal changes, irrespective of cancer stemness. Because currently used E‐cad‐Fc is a fusion protein consisting of human E‐cadherin and mouse Fc domain, we examined whether a fusion protein of human E‐cadherin and human Fc domain would directly induce mesenchymal changes in ODC‐degron‐transduced KM12SM cells. As shown in Figure S3, recombinant human E‐cadherin‐human Fc chimera protein caused spindle‐like morphology at 48 hours and highly induced mesenchymal markers, Snail1, Slug, Twist1 expression and TGF‐β1. These findings emphasize that human E‐cadherin fragment is essentially important to induce mesenchymal changes and that this unique effect is not likely to be a non‐specific reaction caused by the mouse Fc domain.
Cumulative evidence further emphasizes the links between EMT and CSC13, 14, 15, 16 including their similar characteristics, such as tumorigenesis, enhanced survival signals, and metastatic ability. Mesenchymal markers including ZEB1, Snail1, and Slug are thought to be involved in CSC maintenance and tumorigenesis.56, 57, 58 SOX9 is stabilized through combination with the transcription factor Slug, promoting CSC properties and metastasis.59 Mesenchymal change may also contribute to spheroid formation and chemoresistance. Recent research on ovarian cancer cells showed that N‐cadherin–positive mesenchymal‐type cells form stable, highly cohesive solid spheroids with a smooth shape, and that their adhesive, migratory, and invasive cell properties were promoted.60 It is also reported that CSC markers such as CD44, CD44v9, and ALDH, along with EMT markers including Snail1, Slug, and ZEB1, contribute to resistance for chemotherapy and radiotherapy.61, 62, 63
In conclusion, we showed that the E‐cad‐Fc matrix facilitated CSC properties and induced mesenchymal changes. Our data suggest that the E‐cad‐Fc matrix may be efficient for providing better access to CSC research as well as EMT investigation.
DISCLOSURE
Authors declare no conflicts of interest for this article.
Supporting information
ACKNOWLEDGMENTS
This work was supported by the Rotary Yoneyama Memorial Foundation and a grant from the Kagoshima Shinsangyo Sousei Investment Limited Partnership (its general partner is Kagoshima Development Co., Ltd). We are grateful to Dr Frank Pajonk (Jonsson Comprehensive Cancer Center, UCLA, CA, USA) for providing the retroviral expression vector pQCXIN‐ZsGreen‐cODC containing green fluorescence ZsGreen‐labeled degronODC (Gdeg). We also thank Emeritus Prof. Toshihiro Akaike (FAIS, Ibaraki, Japan) and Prof. Toshinari Minamoto (Division of Translational and Clinical Oncology, Cancer Research Institute, Kanazawa University, Ishikawa, Japan) for providing the initial test samples of the E‐cad‐Fc fusion protein and KM12SM cells, respectively.
Qian Y, Wu X, Yokoyama Y, et al. E‐cadherin‐Fc chimera protein matrix enhances cancer stem‐like properties and induces mesenchymal features in colon cancer cells. Cancer Sci. 2019;110:3520–3532. 10.1111/cas.14193
REFERENCES
- 1. Hay Elizabeth D. An overview of epithelio‐mesenchymal transformation. Cells Tissues Organs. 1995;154(1):8‐20. [DOI] [PubMed] [Google Scholar]
- 2. Kim DH, Xing T, Yang Z, Dudek R, Lu Q, Chen YH. Epithelial mesenchymal transition in embryonic development, tissue repair and cancer: a comprehensive overview. J Clin Med. 2017;7(1):1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Chen T, You Y, Jiang H, Wang ZZ. Epithelial–mesenchymal transition (EMT): a biological process in the development, stem cell differentiation, and tumorigenesis. J Cell Physiol. 2017;232(12):3261‐3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Moreno‐Bueno G, Portillo F, Cano A. Transcriptional regulation of cell polarity in EMT and cancer. Oncogene. 2008;27(55):6958. [DOI] [PubMed] [Google Scholar]
- 5. Huang RY, Guilford P, Thiery JP. Early events in cell adhesion and polarity during epithelial‐mesenchymal transition. J Cell Sci. 2012;125(Pt 19):4417‐4422. [DOI] [PubMed] [Google Scholar]
- 6. Kalluri R, Weinberg RA. The basics of epithelial‐mesenchymal transition. J Clin Investig. 2008;119(6):1420‐1428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kalluri R. EMT: when epithelial cells decide to become mesenchymal‐like cells. J Clin Investig. 2009;119(6):1417‐1419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Sánchez‐Tilló E, Liu Y, de Barrios O, et al. EMT‐activating transcription factors in cancer: beyond EMT and tumor invasiveness. Cell Mol Life Sci. 2012;69(20):3429‐3456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Yilmaz M, Christofori G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009;28(1‐2):15‐33. [DOI] [PubMed] [Google Scholar]
- 10. Mani SA, Guo W, Liao MJ, et al. The epithelial‐mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704‐715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Shibue T, Weinberg RA. EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat Rev Clin Oncol. 2017;14(10):611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Fantozzi A, Gruber DC, Pisarsky L, et al. VEGF‐mediated angiogenesis links EMT‐induced cancer stemness to tumor initiation. Can Res. 2014;74(5):1566‐1575. [DOI] [PubMed] [Google Scholar]
- 13. Gil J, Stembalska A, Pesz KA, Sasiadek MM. Cancer stem cells: the theory and perspectives in cancer therapy. J Appl Genet. 2018;49(2):193‐199. [DOI] [PubMed] [Google Scholar]
- 14. Zhao J. Cancer stem cells and chemoresistance: the smartest survives the raid. Pharmacol Ther. 2016;160:145‐158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16(3):225‐238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Ye J, Wu D, Wu P, Chen Z, Huang J. The cancer stem cell niche: cross talk between cancer stem cells and their microenvironment. Tumour Biol. 2014;35(5):3945‐3951. [DOI] [PubMed] [Google Scholar]
- 17. Lee MK, Lin SP, HuangFu WC, Yang DS, Liu IH. Endothelial‐derived extracellular matrix ameliorate the stemness deprivation during ex vivo expansion of mouse bone marrow‐derived mesenchymal stem cells. PLoS ONE. 2017;12(8):e0184111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Zvibel I, Wagner A, Pasmanik‐Chor M, et al. Transcriptional profiling identifies genes induced by hepatocyte‐derived extracellular matrix in metastatic human colorectal cancer cell lines. Clin Exp Metas. 2013;30(2):189‐200. [DOI] [PubMed] [Google Scholar]
- 19. Hovatta O, Mikkola M, Gertow K, et al. A culture system using human foreskin fibroblasts as feeder cells allows production of human embryonic stem cells. Hum Reprod. 2003;18(7):1404‐1409. [DOI] [PubMed] [Google Scholar]
- 20. Rodin S, Domogatskaya A, Ström S, et al. Long‐term self‐renewal of human pluripotent stem cells on human recombinant laminin‐511. Nat Biotechnol. 2010;28(6):611. [DOI] [PubMed] [Google Scholar]
- 21. Nagaoka M, Si‐Tayeb K, Akaike T, Duncan SA. Culture of human pluripotent stem cells using completely defined conditions on a recombinant E‐cadherin substratum. BMC Dev Biol. 2010;10(1):60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA. Loss of E‐cadherin promotes metastasis via multiple downstream transcriptional pathways. Can Res. 2008;68(10):3645‐3654. [DOI] [PubMed] [Google Scholar]
- 23. Nagaoka M, Koshimizu U, Yuasa S, et al. E‐cadherin‐coated plates maintain pluripotent ES cells without colony formation. PLoS ONE. 2006;1(1):e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zhang Y, Mao H, Qian M, et al. Surface modification with E‐cadherin fusion protein for mesenchymal stem cell culture. J Mater Chem B. 2016;4(24):4267‐4277. [DOI] [PubMed] [Google Scholar]
- 25. Hu QP, Kuang JY, Yang QK, Bian XW, Yu SC. Beyond a tumor suppressor: soluble E‐cadherin promotes the progression of cancer. Int J Cancer. 2016;138(12):2804‐2812. [DOI] [PubMed] [Google Scholar]
- 26. Brouxhon SM, Kyrkanides S, Teng X, et al. Soluble E‐cadherin: a critical oncogene modulating receptor tyrosine kinases, MAPK and PI3K/Akt/mTOR signaling. Oncogene. 2014;33(2):225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Brouxhon SM, Kyrkanides S, Teng X, et al. Soluble‐E‐cadherin activates HER and IAP family members in HER2 + and TNBC human breast cancers. Mol Carcinog. 2014;53(11):893‐906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hamabe A, Konno M, Tanuma N, et al. Role of pyruvate kinase M2 in transcriptional regulation leading to epithelial‐mesenchymal transition. Proc Natl Acad Sci. 2014;111(43):15526‐15531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Docherty NG, O'Sullivan OE, Healy DA, et al. TGF‐β1‐induced EMT can occur independently of its pro‐apoptotic effects and is aided by EGF receptor activation”. Am J Physiol Renal Physiol. 2006;290(5):1202‐1212. [DOI] [PubMed] [Google Scholar]
- 30. Alipio Zaida A, Jones N, Liao W, et al. Epithelial to mesenchymal transition (EMT) induced by bleomycin or TFGb1/EGF in murine induced pluripotent stem cell‐derived alveolar Type II‐like cells. Differentiation. 2011;82(2):89‐98. [DOI] [PubMed] [Google Scholar]
- 31. Adikrisna R, Tanaka S, Muramatsu S, et al. Identification of pancreatic cancer stem cells and selective toxicity of chemotherapeutic agents. Gastroenterology. 2012;143(1):234‐245. [DOI] [PubMed] [Google Scholar]
- 32. Munakata K, Uemura M, Tanaka S, et al. Cancer stem‐like properties in colorectal cancer cells with low proteasome activity. Clin Cancer Res. 2016;22(21):5277‐5286. [DOI] [PubMed] [Google Scholar]
- 33. Hayashi K, Tamari K, Ishii H, et al. Visualization and characterization of cancer stem‐like cells in cervical cancer. Int J Oncol. 2014;45(6):2468‐2474. [DOI] [PubMed] [Google Scholar]
- 34. Tamari K, Hayashi K, Ishii H, et al. Identification of chemoradiation‐resistant osteosarcoma stem cells using an imaging system for proteasome activity. Int J Oncol. 2014;45(6):2349‐2354. [DOI] [PubMed] [Google Scholar]
- 35. Ogata Y, Hara Y, Akagi Y, Ohkita A, Morodomi T, Shirouzu K. Metastatic model of human colon cancer constructed using orthotopic implantation in nude mice. Kurume Med J. 1998;45(1):121‐125. [DOI] [PubMed] [Google Scholar]
- 36. Nagano O, Okazaki S, Saya H. Redox regulation in stem‐like cancer cells by CD44 variant isoforms. Oncogene. 2013;32(44):5191. [DOI] [PubMed] [Google Scholar]
- 37. Zavros Y. Initiation and maintenance of gastric cancer: a focus on CD44 variant isoforms and cancer stem cells. Cell Mol Gastroenterol Hepatol. 2017;4(1):55‐63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Jaggupilli A, Elkord E. Significance of CD44 and CD24 as cancer stem cell markers: an enduring ambiguity. Clin Dev Immunol. 2012;2012:708036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Shenoy A, Butterworth E, Huang EH. ALDH as a marker for enriching tumorigenic human colonic stem cells. Methods Mol Biol. 2012:916;373‐385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Yanai H, Atsumi N, Tanaka T, et al. Intestinal cancer stem cells marked by Bmi1 or Lgr5 expression contribute to tumor propagation via clonal expansion. Sci Rep. 2017;7:41838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Yan KS, Chia LA, Li X, et al. The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations. Proc Natl Acad Sci. 2012;109(2):466‐471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Roche KC, Gracz AD, Liu XF, Newton V, Akiyama H, Magness ST. SOX9 maintains reserve stem cells and preserves radioresistance in mouse small intestine. Gastroenterology. 2015;149(6):1553‐1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Moon BS, Jeong WJ, Park J, Kim TI, Min do S, Choi KY. Role of oncogenic K‐Ras in cancer stem cell activation by aberrant Wnt/β‐catenin signaling. J Natl Cancer Inst. 2014;106(2):djt373. [DOI] [PubMed] [Google Scholar]
- 44. Ma L, Zhang G, Miao XB, et al. Cancer stem‐like cell properties are regulated by EGFR/AKT/β–catenin signaling and preferentially inhibited by gefitinib in nasopharyngeal carcinoma. FEBS J. 2013;280(9):2027‐2041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Singh S, Trevino J, Bora‐Singhal N, et al. EGFR/Src/Akt signaling modulates Sox2 expression and self‐renewal of stem‐like side‐population cells in non‐small cell lung cancer. Mol Cancer. 2012;11(1):73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. O'Brien CA, Kreso A, Jamieson CH. Cancer stem cells and self‐renewal. Clin Cancer Res. 2010;16(12):3113‐3120. [DOI] [PubMed] [Google Scholar]
- 47. Lenos KJ, Vermeulen L. Cancer stem cells don't waste their time cleaning—low proteasome activity, a marker for cancer stem cell function. Ann Transl Med. 2016;4(24):519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Voutsadakis IA. Proteasome expression and activity in cancer and cancer stem cells. Tumour Biol. 2017;39(3):1010428317692248. [DOI] [PubMed] [Google Scholar]
- 49. Yu T, Chen X, Zhang W, et al. Regulation of the potential marker for intestinal cells, BMI1, by β‐catenin and the zinc finger protein KLF4 implications for colon cancer. J Biol Chem. 2012;287(6):3760‐3768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Basu S, Haase G, Ben‐Ze'ev A. Wnt signaling in cancer stem cells and colon cancer metastasis. F1000Res. 2016;5:699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ma F, Ye H, He HH, et al. SOX9 drives WNT pathway activation in prostate cancer. J Clin Investig. 2016;126(5):1745‐1758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Frankel A, Man S, Elliott P, Adams J, Kerbel RS. Lack of multicellular drug resistance observed in human ovarian and prostate carcinoma treated with the proteasome inhibitor PS‐341. Clin Cancer Res. 2000;6(9):3719‐3728. [PubMed] [Google Scholar]
- 53. Brouxhon SM, Kyrkanides S, Teng X, et al. Monoclonal antibody against the ectodomain of E‐cadherin (DECMA‐1) suppresses breast carcinogenesis: involvement of the HER/PI3K/Akt/mTOR and IAP pathways. Clin Cancer Res. 2013;19(12):3234‐3246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Zou S, Ma C, Yang F, Xu X, Jia J, Liu Z. FBXO31 suppresses gastric cancer EMT by targeting snail1 for proteasomal degradation. Mol Cancer Res. 2018;16(2):286‐295. [DOI] [PubMed] [Google Scholar]
- 55. Banno A, Garcia DA, van Baarsel ED, et al. Downregulation of 26S proteasome catalytic activity promotes epithelial‐mesenchymal transition. Oncotarget. 2016;7(16):21527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Li Y, Wang L, Pappan L, Galliher-Beckley A, Shi J. IL‐1β promotes stemness and invasiveness of colon cancer cells through Zeb1 activation. Mol Cancer. 2012;11(1):87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Fan F, Samuel S, Evans KW, et al. Overexpression of Snail induces epithelial–mesenchymal transition and a cancer stem cell–like phenotype in human colorectal cancer cells. Cancer Med. 2012;1(1):5‐16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Sun Y, Song GD, Sun N, Chen JQ, Yang SS. Slug overexpression induces stemness and promotes hepatocellular carcinoma cell invasion and metastasis. Oncol Lett. 2014;7(6):1936‐1940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Luanpitpong S, Li J, Manke A, et al. SLUG is required for SOX9 stabilization and functions to promote cancer stem cells and metastasis in human lung carcinoma. Oncogene. 2016;35(22):2824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Klymenko Y, Johnson J, Bos B, et al. Heterogeneous cadherin expression and multicellular aggregate dynamics in ovarian cancer dissemination. Neoplasia. 2017;19(7):549‐563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Tomono T, Yano K, Ogihara T. Snail‐induced epithelial‐to‐mesenchymal transition enhances P‐gp‐Mediated multidrug resistance in HCC827 cells. J Pharm Sci. 2017;106(9):2642‐2649. [DOI] [PubMed] [Google Scholar]
- 62. Findlay VJ, Wang C, Nogueira LM, et al. SNAI2 modulates colorectal cancer 5‐fluorouracil sensitivity through miR145 repression. Mol Cancer Ther. 2014;13(11):2713‐2726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Guo C, Ma J, Deng G, et al. ZEB1 promotes oxaliplatin resistance through the induction of epithelial‐mesenchymal transition in colon cancer cells. J Cancer. 2017;8(17):3555. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials