

ABSTRACT
During development, the ectoderm is patterned by a combination of BMP and WNT signaling. Research in model organisms has provided substantial insight into this process; however, there are currently no systems in which to study ectodermal patterning in humans. Further, the complexity of neural plate border specification has made it difficult to transition from discovering the genes involved to deeper mechanistic understanding. Here, we develop an in vitro model of human ectodermal patterning, in which human embryonic stem cells self-organize to form robust and quantitatively reproducible patterns corresponding to the complete medial-lateral axis of the embryonic ectoderm. Using this platform, we show that the duration of endogenous WNT signaling is a crucial control parameter, and that cells sense relative levels of BMP and WNT signaling in making fate decisions. These insights allowed us to develop an improved protocol for placodal differentiation. Thus, our platform is a powerful tool for studying human ectoderm patterning and for improving directed differentiation protocols.
This article has an associated ‘The people behind the papers’ interview.
KEY WORDS: BMP signaling, Ectoderm development, WNT signaling, Human embryonic stem cells, Self-organization
Highlighted Article: An in vitro system based on human embryonic stem cells recapitulates the medial-lateral patterning of human ectoderm and reveals how the ectoderm is spatially patterned by WNT and BMP signaling.
INTRODUCTION
The human ectodermal germ layer arises as a spatially organized epithelium composed of progenitors of the neural plate, neural crest, placodes and epidermis along the medial-lateral axis of the developing embryo. Establishment of this molecular pattern by upstream signaling events is necessary to properly coordinate the biomechanical processes of neurulation, in which the open neural plate folds along the midline as the neural folds fuse dorsally to form a tubular structure along the anterior-posterior axis (Mansouri et al., 2001; Ybot-Gonzalez et al., 2007). Studies from model organisms have identified key morphogens and corresponding inhibitors that coordinate the patterning of ectodermal fates. In several models, the emergence of the neural domain in the medial ectoderm results from the suppression of BMP, WNT and Nodal signaling by secreted inhibitors that include noggin, chordin, follistatin and DKK1 (del Barco Barrantes et al., 2003; Fainsod et al., 1997; Li et al., 2013; Sasai et al., 1995; Smith and Harland, 1992; Wilson et al., 1997). In contrast, BMP signaling is required at the time of gastrulation to initiate the formation of the non-neural ectoderm marked by the lateral expression of AP2α (TFAP2A), GATA3 and E-cadherin (ECAD; also known as cadherin 1) (Brugmann and Moody, 2005; Choi and Gumbiner, 1989; Li et al., 2013; Malaguti et al., 2013; Pieper et al., 2012; Schlosser, 2006).
Once established, the non-neural territory is further refined to distinct progenitor domains by spatiotemporally organized inductive signals. BMP activity in the anterior-proximal region (equivalent to lateral human ectoderm) of embryonic day (E)7 mouse embryos overlapped with epidermal precursor markers cytokeratin 8 (K8; KRT8) and 18 (K18; KRT18), and exogenous BMP signaling induced epidermal precursors in tissue grafts that would otherwise acquire neural fates (Li et al., 2013). FGF agonists, together with an attenuation of BMP and WNT signaling, initiate placodal emergence indicated by expression of SIX1 and EYA1/2 (Ahrens and Schlosser, 2005; Bhat et al., 2013; Kwon et al., 2010; Litsiou et al., 2005). Similarly, neural crest differentiation requires FGF signaling together with an intermediate BMP signaling level (Geary and LaBonne, 2018; Piacentino and Bronner, 2018; Stuhlmiller and García-Castro, 2012; Villanueva et al., 2002; Yardley and García-Castro, 2012). Unlike placodes, the specification of neural crest (marked by PAX3/7 and SOX9/10) at the neural plate border is dependent on a source of secreted WNT ligands from the paraxial mesoderm or the ectoderm itself (García-Castro et al., 2002; Litsiou et al., 2005; Sato et al., 2005; Simões-Costa et al., 2015).
Many studies have leveraged the knowledge gained from studies of model organisms to develop directed differentiation protocols using pluripotent human embryonic stem cells (hESCs). These protocols modulate the FGF, TGFβ, BMP and WNT signaling pathways, and aim to produce pure populations of desired cell fates within the ectoderm. As in vivo, inhibition of TGFβ superfamily signaling is essential for differentiation to neural fates (Chambers et al., 2009; Elkabetz et al., 2008; Zhang et al., 2010), and appropriately timed introduction of BMP4 to the media can induce epidermal and placodal fates (Dincer et al., 2013; Ealy et al., 2016; Leung et al., 2013; Qu et al., 2016; Tchieu et al., 2017). Alternatively, WNT pathway activation or precise levels of BMP4 have been shown to yield neural crest progenitors (Gomez et al., 2019; Hackland et al., 2017; Leung et al., 2016; Tchieu et al., 2017). Achieving pure populations of cells is likely essential in some applications; however, development involves the formation of self-organizing patterns of multiple cell types. Moreover, the patterning of human ectoderm differs notably from that of other mammals and model organisms (Barry et al., 2017; Böck et al., 2018). Studying human ectodermal patterning requires the development of in vitro models in which cells self-organize and spatiotemporal patterns emerge reproducibly in a fashion similar to that of the human embryo in vivo.
Recently, our lab and others have demonstrated that hESCs possess a remarkable ability to self-organize, and that this ability can be harnessed by confining them to micropatterned surfaces, so that embryonic fate patterns are generated in vitro (Chhabra et al., 2018 preprint; Etoc et al., 2016; Heemskerk et al., 2019; Knight et al., 2018; Warmflash et al., 2014; Xue et al., 2018). In this study, we developed a micropatterned platform in which cells self-organize to all four main fates within the ectoderm, and we use this system to dissect the spatiotemporal signaling events that pattern this model of human ectoderm. We demonstrate that when hESCs are directed to adopt the ectodermal germ layer fate by inhibiting Nodal signaling, the subsequent application of BMP4 to micropatterned colonies is sufficient to trigger medial-lateral patterning of multiple cell fates. Furthermore, we identified the duration of WNT signaling as a crucial control parameter that modulates the fate composition of ectodermal patterns. Finally, we used this system to expand our understanding of how BMP and WNT signaling pattern the ectoderm. In particular, we show that cells are sensitive to the relative, rather than absolute, levels of BMP and WNT, and that this system can be used to dissect interactions between signaling and transcription factor networks in neural crest differentiation. We also show that taking the knowledge gained from this system into account allows us to improve on current protocols for placodal differentiation. Taken together, this study presents a novel self-organized ectodermal patterning system, and shows the power of this system for understanding human ectoderm development and improving differentiation towards desired ectodermal fates.
RESULTS
Determining the window of competence for ectodermal patterning
We sought to create an in vitro model system in which to study the formation of patterned human ectodermal tissue. In analogy with our previous work creating gastrulation-stage patterns in vitro (Chhabra et al., 2018preprint; Heemskerk et al., 2019; Warmflash et al., 2014), we reasoned that geometrically confined hESCs treated with appropriately timed exogenous stimuli would create self-organized patterns of fates within the ectodermal germ layer. In both model organisms and embryonic stem cells, Nodal inhibition has been shown to be crucial for preventing mesendoderm differentiation, allowing cells to adopt ectodermal fates, whereas BMP signaling is responsible for generating the medial-lateral pattern within the ectoderm (Li et al., 2013; Liu et al., 2018). Thus, we followed a two-stage protocol where hESCs are initially induced by Nodal inhibition and subsequently stimulated by BMP4. Although we expected that performing this protocol in standard culture would generate ectodermal cells that are spatially disorganized, it would provide a starting point for micropatterning experiments that test whether geometric confinement leads to ordered emergence of the same set of fates. Previous work in mouse and human ESCs has shown that prolonged Nodal inhibition leads to commitment to the neural fate (Li et al., 2015; Liu et al., 2018; Smith et al., 2008), so we specifically sought a temporal window in which cells are committed to the ectoderm but not exclusively to neural fates.
Compared with pluripotent cells, ectodermal progenitors are characterized by lower levels of the pluripotency markers OCT4 (POU5F1) and NANOG, high levels of SOX2, and the absence of neural-specific genes such as PAX6 and NCAD (CDH2) (Chambers et al., 2009; Li et al., 2015; Liu et al., 2018; Wang et al., 2012). To determine when hESCs enter this state, we examined these markers as a function of the duration of Nodal inhibition, achieved by growing cells in N2B27 media supplemented with 10 μM of SB431542 (hereafter SB), a small molecule ALK4/5/7 (also known as ACVR1B, TGFBR1 and BMPR1B, respectively) receptor-kinase inhibitor (hereafter called ectodermal induction media). By day 2, OCT4 and NANOG were repressed, and SOX2 expression levels were elevated compared with those in the pluripotent state (Fig. 1C,D, Fig. S1B). On day 3, we observed low and heterogeneous expression levels of PAX6 and NCAD (Fig. 1A, Fig. S1A). By day 4, both PAX6 and NCAD expression increased in intensity and were homogeneously expressed by day 5 (Fig. 1A,B, Fig. S1A). Together, these data indicate that hESCs acquire a transient ectodermal progenitor state (SOX2+/NANOG−/OCT−/PAX6−/NCAD−) by day 2 of Nodal inhibition, but quickly differentiate towards the neural lineage upon continued Nodal inhibition.
Fig. 1.

During neural differentiation cells transition through an ectodermal progenitor state before commitment. (A,C) Representative images of cells fixed on the indicated days during the course of Nodal inhibition and immunostained for the neural progenitor marker PAX6 (A) or the pluripotent markers SOX2, OCT4 and NANOG (C). (B,D) Quantification of the fold change in average intensities of PAX6 (B) or SOX2, OCT4 and NANOG (D) normalized to DAPI for the indicated days relative to day 0. N=3 experiments. (E,F) Representative images of cells immunofluorescently co-stained for AP2α/PAX6 or SOX2/NANOG/OCT4 at the conclusion of a two-phase induction protocol. Cells were initially differentiated in Nodal inhibition media for either 2 (E) or 4 (F) days and then treated with the indicated culture conditions for the subsequent 6 (E) or 4 (F) days. Experiment replicated three times. (G) Representative images of cells immunofluorescently labeled for CDX2. All cells were treated with BMP4 for 2 days following either no prior treatment or 3 days of Nodal inhibition. Experiment replicated three times. (H) Representative images of cells immunofluorescently labeled for BRA at the conclusion of the experiment. All cells were initially induced for 3 days in Nodal inhibition media. Thereafter, the media was exchanged for treatment conditions indicated in the banner above the corresponding images. First row banners indicate the base media and the second banner row indicates the signals added to the base media. Experiment replicated three times. TGFβi, SB431542. Data are mean±s.e.m. Scale bars: 25 µm (C); 50 µm (A,E,F); 100 µm (G,H).
We next assessed the differentiation potential of hESCs as they transition from ectodermal to neural progenitors by exchanging the ectodermal induction media for a neutral (N2B27), pluripotency-maintaining (mTeSR), or non-neural differentiation (N2B27+50 ng/ml BMP4) media. Switching to a non-neural differentiation media on day 2 directed cells to non-neural ectoderm (AP2α+/PAX6−/SOX2−), whereas neutral media allowed most cells to adopt a neural fate (PAX6+/SOX2+/AP2α−) (Fig. 1E). Reverting to mTeSR on day 2 yielded a mixed population of fates consisting of neural, non-neural and pluripotent cells (Fig. 1E, Fig. S1C). In contrast, exchanging ectodermal for non-neural induction media on day 4 generated clear territories of neural (PAX6+/AP2α−) and non-neural fates (PAX6−/AP2α+) by day 8, whereas cells changed to neutral or mTeSR media generated an expression profile consistent with the acquisition of a neural progenitor fate (high PAX6, low AP2α) (Fig. 1F, Fig. S1D) (Chambers et al., 2009). Collectively, these data demonstrate that hESCs remained competent to revert to pluripotency or differentiate towards non-neural fates after 2 days of ectodermal induction, and after 4 days all cells had committed to the ectodermal lineage and a fraction of cells had committed towards the neural fate.
Pluripotent cells give rise to mesodermal and extraembryonic fates in response to BMP4 (Bernardo et al., 2011; Nemashkalo et al., 2017; Xu et al., 2002). We next tested whether cells lose competence to differentiate to these fates following 3 days of ectodermal induction. Pluripotent hESCs treated with BMP4 for 2 days upregulated CDX2; however, its expression was absent if cells were first grown in ectoderm induction media for 3 days (Fig. 1G). Additionally, we noted that following 3 days of ectodermal induction, 2 days of BMP4 treatment failed to upregulate mesodermal fates, defined by brachyury (BRA; also known as TBXT) expression, when introduced with or without the Nodal inhibitor SB in N2B27 media. BRA+ cells were detected in the case when cells were treated with BMP4 in mTeSR media following 3 days of ectoderm induction (Fig. 1H). However, addition of SB to mTeSR blocked mesodermal differentiation (Fig. 1H). This indicated that during the first 3 days of ectodermal differentiation, cells lose the potential to differentiate towards extra-embryonic fates, but maintain competence to differentiate to mesodermal fates. These findings are consistent with earlier reports that demonstrate BMP-induced mesodermal differentiation in pluripotent stem cells requires an exogenous source of TGFβ and/or FGF (Bernardo et al., 2011; Yu et al., 2011).
A two-step induction protocol generates patterns of human ectodermal fates in vitro
The previous experiments demonstrated that a properly timed two-step induction protocol with continuous Nodal inhibition could give rise to multiple ectodermal fates. We reasoned that carrying out this induction in geometrically confined colonies might lead to reproducible spatial patterning (Warmflash et al., 2014). We seeded hESCs on micropatterned surfaces and grew them for 72 h in ectodermal induction media. Patterning was initiated with the addition of 50 ng/ml BMP4 in the presence of SB (10 μM); cells were fixed following 72 h of this treatment. At the conclusion of the induction period, stem cell colonies routinely formed multilayered structures with a dense ring of cells aggregating at a reproducible radius within the colony. We evaluated the expression of lineage-specific markers within these micropatterned colonies.
Cells at the center of the micropatterned colonies differentiated to neural lineages (PAX6+/NCAD+/OTX2+/ECAD−/ISL1−/GATA3−/K8−) whereas those closer to the edge differentiated to non-neural ectoderm (PAX6−/NCAD–/ECAD+/ISL1+/GATA3+/KRT8+) (Fig. 2A, Fig. S2A-D,F) (Leung et al., 2013; Ozair et al., 2013; Pieper et al., 2012; Schlosser, 2006; Tchieu et al., 2017). In addition, we observed heterogeneous expression of SOX1, a late human neural progenitor marker, in the center of the colony co-expressed with NCAD and PAX6 (Fig. S2F) (Zhang et al., 2010). The neural crest markers SOX9 and PAX3 were expressed in a ring of cells between the neural and non-neural domains (Fig. 2A, Fig. S2E) (Betters et al., 2010; Bhat et al., 2013; Monsoro-Burq et al., 2005; Pieper et al., 2012; Plouhinec et al., 2017; Spokony et al., 2002). Next to these cells was a ring of cells positive for the placodal marker SIX1 (Fig. 2A, Fig. S2A,B) (Ahrens and Schlosser, 2005; Christophorou et al., 2009; Dincer et al., 2013; Plouhinec et al., 2017; Schlosser, 2006). These SIX1+ placodal cells were a subset of the ECAD+/ISL1+/GATA3+/K8+ population, and the SIX1− cells in this population represent epidermal precursors (Fig. S2A-D) (Dincer et al., 2013; Page, 1989; Pieper et al., 2012; Tchieu et al., 2017).
Fig. 2.

A two-phase protocol generates self-organized patterns of ectodermal fates. (A-C) Representative images of colonies stained with the indicated antibodies following treatment with either a two-step protocol consisting of 3 days in ectoderm induction media and 3 days in N2B27+BMP+SB (A), the same protocol except the first 3 days were in N2B27 alone (B), or 6 days of ectoderm induction media (C). The top row in (A) shows a schematic of the organization of fates in the anterior ectoderm. Each experiment was replicated at least three times. Scale bars: 100 μm.
We next evaluated the necessity of Nodal inhibition in the first phase and BMP stimulation in the second to achieve ectoderm patterns. Removing SB from the first phase resulted in downregulation of NCAD and SOX1 in the colony center, contraction of the PAX6+ domain, the expansion of SOX9 expression to the colony edge, and the co-expression of SOX9 in the remaining PAX6+ cells at the colony center (Fig. 2B, Fig. S2G,H). Thus, inhibition of Nodal signaling during the first 3 days is required to instruct future neural fates in the center of the colony upon BMP4 treatment.
Colonies that were grown for 6 days in ectoderm induction media without added BMP4 expressed PAX6 and NCAD throughout, whereas the surface ectoderm marker ISL1 was completely absent at the colony edge (Fig. 2C). We observed a small number of SOX9-expressing cells near the colony edge but without a clear pattern. Thus, exogenous BMP4 stimulation is required for surface ectoderm differentiation and self-organized patterning.
Duration of endogenous WNT signaling affects the composition of ectodermal tissue
Although the protocol above yielded self-organized patterns consisting of four ectodermal lineages, the neural crest frequently formed in a broad domain without sharp boundaries, and there was limited placodal differentiation. WNT signaling is crucial for neural crest differentiation, and we hypothesized that WNT ligands are endogenously produced and secreted, and that modulating these signals could alter patterning outcomes (Kurek et al., 2015). We therefore introduced IWP2, a small molecule inhibitor of WNT ligand secretion, at varying times during the two-step induction protocol (Chen et al., 2009). Introduction of IWP2 for all 6 days, or for the final 3 days, concurrently with BMP treatment, led to a dramatic increase in placodal SIX1+ cells and completely inhibited expression of the neural crest markers PAX3 and SOX9 (Fig. 3A, Fig. S3A,B). Delaying IWP2 treatment for 12 h following BMP4 addition was sufficient to initiate neural crest marker expression within the neural domain and adjacent to the emergent placodal cells (Fig. 3A, Fig. S3A,C,D). Allowing longer durations of WNT signaling by delaying the introduction of IWP2 caused an expansion of the neural crest territory from the edge of the neural domain inwards (Fig. 3A). Longer periods of endogenous WNT signaling also reduced the expression levels of the placodal marker SIX1 (Fig. 3A). Together, our results demonstrate that WNT ligands are endogenously secreted and operate to alter the composition of fates within ectodermal tissue. Continuous inhibition of WNT secretion results in colonies consisting of domains of neural, placodal and future epidermal lineages, and lacking neural crest. WNT signaling creates the domain of neural crest cells, and longer durations of signaling expand this domain at the expense of neural and placodal fates.
Fig. 3.

Endogenous WNT ligands drive differentiation to neural crest at the expense of placodal fates. (A) Representative images of colonies immunostained for PAX6, SOX9 or SIX1. Colonies were initially induced for 3 days in ectoderm induction media and then subsequently differentiated for 3 days in N2B27 media containing BMP4 and SB. The time between BMP4 and IWP2 addition is indicated in the induction schematic and linked in red with the banner above the corresponding image. Experiment replicated four times. (B) Quantification of the images in A represented as average nuclear intensities of the indicated markers normalized to DAPI as a function of radial position. N=3 colonies. (C) Representative images of cells in standard culture immunostained for SIX1. Cells were initially induced for 3 days in ectoderm induction media and then treated with either 5 or 50 ng/ml of BMP4 in media with (+) or without (−) IWP2 for the subsequent 3 days. (D) Fraction of cells expressing SIX1. N=3 experiments; P=0.011 (two-sided t-test). Data are mean±s.e.m. (E) Kymograph of β-catenin signaling activity over a 24-h period (on day 4) in micropatterned colonies initially treated in Nodal inhibition media for 3 days and then with the indicated signaling conditions. N=4 colonies and repeated twice. Colony diameter in A: 700 µm. Scale bars: 100 µm.
The above experiments demonstrated that delaying IWP2 addition for 12 h after stimulation with BMP4 was sufficient to initiate neural crest differentiation. We reasoned that WNT signaling must begin prior to this time point to drive this differentiation. To test this, we monitored WNT pathway dynamics using a cell line with GFP fused to β-catenin at the endogenous locus (Massey et al., 2019). The onset of WNT signaling occurs within 10 h of BMP4 treatment and its activity is strongly reduced by adding IWP2 simultaneously with BMP4 (Fig. 3E). WNT activity was strongest in the region of future neural crest and moved inward towards the center of the colony over time. Thus, WNT signaling required for neural crest differentiation is activated shortly after BMP4 addition.
WNT inhibition improves placode differentiation protocols
Placodes are unique cells of the ectoderm that give rise to the sensory organs. Previous reports have suggested that the specification of placodal fates by BMP is dose dependent with high levels (>10 ng/ml) inhibiting its expression and intermediate levels (∼5 ng/ml) optimally inducing SIX1 (Leung et al., 2013; Tchieu et al., 2017). An alternative hypothesis is that higher levels of BMP induce greater WNT signaling, which diverts potential placodal cells to neural crest. To distinguish between these hypotheses, hESCs were seeded in standard culture and initially induced for 3 days in ectoderm induction media. Thereafter, the cells were treated with 5 or 50 ng/ml of BMP4 either with or without IWP2 for 3 days. We did not observe a significant difference in placodal expression as a consequence of BMP4 dosage; however, including WNT inhibition with either dose of BMP4 elicited a large increase in placodal expression (Fig. 3C,D). Thus, preventing WNT secretion enables efficient placodal differentiation independently of the BMP4 dose.
The width of the domain of placodal and epidermal progenitors is controlled by BMP4 concentration
Next we sought to understand better the correspondence between the level of exogenous BMP4 and patterning outcomes. In all cases, we implemented a three-step induction protocol with IWP2 added 1 day after BMP4 so that we achieved robust patterns of all four ectodermal fates at sufficient BMP4 doses. At high concentrations (50 and 5 ng/ml), we observed the spatial organization of ectoderm progenitors described above (Fig. 4Ai,Aii). Unexpectedly, we observed expression of AP2α within the neural domain, whereas in vivo, its expression pattern is limited to the future neural crest and surface ectoderm (Fig. 4Ai,Aii) (Betters et al., 2010; de Crozé et al., 2011; Kwon et al., 2010; Luo et al., 2002, 2003; Mitchell et al., 1991). With lower doses of BMP4, AP2α expression was restricted to the edge of the colony and was mutually exclusive with PAX6 expression. The width of the AP2α-expressing territory depended on the BMP4 dose, with higher doses giving wider regions of expression (Fig. 4Aiii,Aiv). Consistent with this trend, lower doses of BMP4 led to an expansion of the PAX6+/NCAD+ neural domain at the expense of the ISL1+/ECAD+ surface ectoderm (Fig. 4A-C, Fig. S4A). Additionally, coordinated with this expansion of the neural domain, we observed a shift of both neural crest and placodal progenitors towards the edge of the colony (Fig. 4A-C). Lowering BMP4 concentration to 0.2 ng/ml led to patterns consisting only of neural in the center and neural crest at the edge of the colony. The loss of placodal fates (ISL1+/SIX1+) was not due to WNT signaling rather than insufficient BMP4 signaling, because inhibition of WNT secretion throughout the induction protocol did not rescue the expression of SIX1 (Fig. S4B). Together, these data suggest there is a critical level of BMP4 that is necessary to initiate surface ectodermal differentiation, and that the width of the territory of surface ectoderm differentiation at the edge of the colony increases with BMP4 concentration.
Fig. 4.

Cell fates are determined from the relative levels of BMP4 and WNT3a. (A,C) Representative images of colonies with the indicated single (A) or multiplexed (C) immunolabels following a three-step ectoderm induction protocol and patterned with (i) 50 ng/ml BMP4, (ii) 5 ng/ml BMP4, (iii) 1 ng/ml BMP4 or (iv) 0.20 ng/ml BMP4 in N2B27 media with SB. Experiment replicated three times. (B,D) Quantifications of images in A (B) and C (D) represented as the average nuclear intensities of the indicated markers normalized to DAPI as a function of radial position. N=3 colonies. (E) Representative images of colonies immunostained for SOX9 or SIX1. Cells were initially differentiated in ectoderm induction media for 3 days and subsequently induced in media containing IWP2 with the indicated levels of BMP4 and WNT3a in the overhead banner. Experiment replicated twice. (F) Quantification of images in E represent the intensities of the indicated markers normalized to DAPI as a function of distance from the colony center. N=3 colonies. Colony diameter: 700 µm. Scale bars: 100 µm.
To confirm that ectoderm fate patterns form in independent cell lines, we induced RUES2 and WTC-11 induced pluripotent stem cells (iPSCs) (Allen Institute) using a three-step induction protocol with 2 ng/ml of BMP4. In each cell line, fate organization was similar to ESI017 with neural fates positioned in the center of the colony, surface ectoderm fates at the edge and neural crest in between the neural and surface ectodermal domains (Fig. S4C,D).
The relative levels of BMP and WNT instruct the decision between neural crest and placode
The results described above argue that cells integrate information from the BMP and WNT pathways to create patterns within the ectoderm. We hypothesized that at the edge of the colony, a region competent to form neural crest and placodes (Fig. 4Aiii,Aiv), the decision between these fates depends on the relative, rather than absolute, levels of WNT and BMP signals. To test this, micropatterned hESC colonies were stimulated with different relative concentrations of exogenous BMP4 and WNT3a in the second phase of the induction protocol. To avoid endogenous WNT signaling complicating the interpretation of these experiments, IWP2 was used throughout the experiment for all treatments. We found that a dose of 1 ng/ml of BMP4 instructed the expression of placodal fates at the edge of the micropattern, whereas placodes were suppressed by 1 ng/ml of BMP4 with 300 ng/ml of WNT3a, and instead cells in a broad ring adopted a SOX9+ neural crest fate (Fig. 4E). Raising the BMP4 concentration to 50 ng/ml while maintaining 300 ng/ml of WNT3a rescued the expression of SIX1 at the edge of the micropattern and reduced that of SOX9 (Fig. 4E). These data show that cell fate decisions within the ectoderm are determined by the relative concentrations of BMP and WNT ligands.
Dynamics of cell fate decisions in micropatterned ectoderm
We next studied the spatiotemporal dynamics of key markers associated with fate acquisition during the course of ectodermal patterning. hESCs were seeded on a micropatterned surface, and differentiated using a three-step induction protocol with 1 ng/ml of BMP4 added on day 3 and IWP2 added 1 day later. Cells were fixed on days 4, 5 and 6 and the expression of fate markers evaluated. We found that the expression of SOX2, initially an ectoderm marker and later a neuroectoderm marker, was expressed throughout the colony on day 4; however, on days 5 and 6, high SOX2 levels were progressively restricted to the center of the colony whereas the edges maintained a lower level of expression (Fig. 5A) (Li et al., 2015; Papanayotou et al., 2008; Plouhinec et al., 2017; Zhang et al., 2010). NCAD and PAX6 formed a domain in the center of the colony with cells at the edge of this domain expressing the highest levels of these markers by day 4. Expression of neural markers was maintained in this territory throughout the rest of the induction protocol (Fig. 5A). The non-neural marker AP2α was found in a ring at the colony border throughout the induction period, whereas ISL1, a marker of surface ectoderm, was detected later on day 5 and maintained on day 6 in the same region (Fig. 5A,B). ECAD, a marker found in both pluripotent and surface ectodermal cells, was weakly expressed in the center of the colony whereas higher levels were observed at the colony edge on day 4. Thereafter, ECAD expression was lost in the center and maintained at the colony edge on days 5 and 6 (Fig. 5A). We observed a dynamic temporal pattern of GATA3 with its expression found at the colony border on days 4 and 5, but its levels reduced by day 6, recapitulating the transient GATA3 expression observed during placodal development in model organisms (Fig. 5A) (Bhatt et al., 2013; Groves and LaBonne, 2014; Schlosser, 2006). Following GATA3 and AP2α expression, cells at the colony edge were positive for the placodal marker SIX1 on day 5 of ectoderm patterning (Fig. 5A). The neural crest markers SOX9 and PAX3 were detected at low levels on day 4 and increased over time with the expression domain remaining in the same radial position (Fig. 5A). Co-expression analysis on day 6 colonies showed overlap of ISL1, GATA3 and SIX1 at the colony edge indicating the formation of surface ectoderm, which is mutually exclusive to the neural domain marked by PAX6 (Fig. 5B, Fig. S5C,D). We found AP2α expression spanning both the surface ectoderm (GATA3/ISL1) and neural crest (SOX9). The AP2α+/SOX9+ cells were found at the edge of the PAX6-expressing neural domain (Fig. 5B,C, Fig. S5A,B,D). Thus, most markers initially appear in their final domain of expression, and expression levels rise and cell fate boundaries are progressively sharpened in time.
Fig. 5.

Spatiotemporal characterization of human ectoderm pattern emergence. (A-C) Representative images of colonies fixed on the indicated days (A), or day 6 of induction (B,C), and immunostained with the indicated antibodies. Colonies were differentiated using a three-step ectoderm induction protocol and patterned with 1 ng/ml BMP4. Experiment replicated twice. Colony diameter: 700 µm. Scale bars: 100 µm.
Signaling history influences the interpretation of BMP4 by hESCs
During the first 3 days of induction, a small molecule Nodal inhibitor was applied without stimulating other pathways; however, endogenous signals may be required during this time. We first assessed BMP signaling through C-terminally phosphorylated SMAD1/5/8 levels (pSmad1/5/8), a proximal read-out of BMP activity (Hoodless et al., 1996; Kawai et al., 2000; Nishimura et al., 1998). We observed a prepattern with active endogenous BMP signaling at the colony edge. This prepattern was specific to BMP signaling as it was abolished by treatment with the BMP inhibitor LDN (Fig. 6A) (Yu et al., 2008). To assess the functional significance of this prepattern, cells were induced for 3 days with SB+LDN and then treated with BMP4 for 3 days. Under these conditions, colonies created patterns consisting only of neural crest at the colony edge and neural in the center without expression of the non-neural markers ISL1, GATA3 and SIX1 (Fig. 6B,C). However, both neural and surface ectodermal fates were observed if colonies were pretreated in SB+LDN for only 1 or 2 days prior to BMP stimulation (Fig. 6B) with greater reductions in the width of the surface ectoderm domain the longer BMP signaling was inhibited. Thus, a prepattern of endogenous BMP signaling is required to preserve competence for surface ectodermal and placodal differentiation at the colony edge.
Fig. 6.

Endogenous BMP signaling prior to BMP4 treatment is required for surface ectodermal differentiation. (A-C) Representative images of colonies immunostained for the indicated antibodies on day 3 following induction in media containing SB only or in combination with LDN (A), and on day 6 following 1, 2 or 3 days of SB+LDN treatment in the first phase of induction prior to BMP4 treatment for 3 days. Conditions that removed dual inhibition before day 3 had media replaced with N2B27+SB such that all conditions were in culture for the same duration prior to the introduction of BMP4 (B). Experiments were repeated three (A) and four (B) times. Colony diameter: 800 µm (A); 700 µm (B). Scale bars: 100 µm.
Combinatorial signaling logic guiding neural crest specification
Experimental evidence from model organisms suggests that the transcription factors SOX9, PAX3 and AP2α are required for the specification of neural crest (Betters et al., 2010; Bhatt et al., 2013; Garnett et al., 2012; Leung et al., 2013; Monsoro-Burq et al., 2005; Sato et al., 2005; Simões-Costa and Bronner, 2015). The regulatory links between these genes and others have been assembled into gene regulatory networks, but it remains unclear how these networks combinatorially interpret signaling from the BMP and WNT pathways. We aimed to leverage our in vitro model to dissect the contributions from BMP and WNT signaling pathways to the specification of neural crest fates. To accomplish this, cells were first induced with either SB alone or in combination with LDN for 3 days and then BMP4 or WNT3a was added alone or in combination with an inhibitor to the second pathway (Fig. 7A,B). Colonies treated with WNT3a (300 ng/ml) following 3 days of Nodal inhibition showed overlapping expression of SOX9, PAX3 and AP2α at the colony edge (Fig. 7Aii). However, inhibition of endogenous BMP (Fig. 6A) prior to WNT stimulation led to the expression of PAX3 without AP2α or SOX9 at the colony edge (Fig. 7Ai,Bi). Conversely, inhibiting WNT at the time of BMP treatment resulted in the upregulation of AP2α but not PAX3 or SOX9 (Fig. 7Aiii). Together, these data suggest that WNT signaling is necessary and sufficient for PAX3 expression whereas BMP is neither, although it may enhance the WNT-induced expression. For AP2α, the opposite is true: BMP is necessary and sufficient whereas WNT is not. For SOX9, both signals are necessary.
Fig. 7.
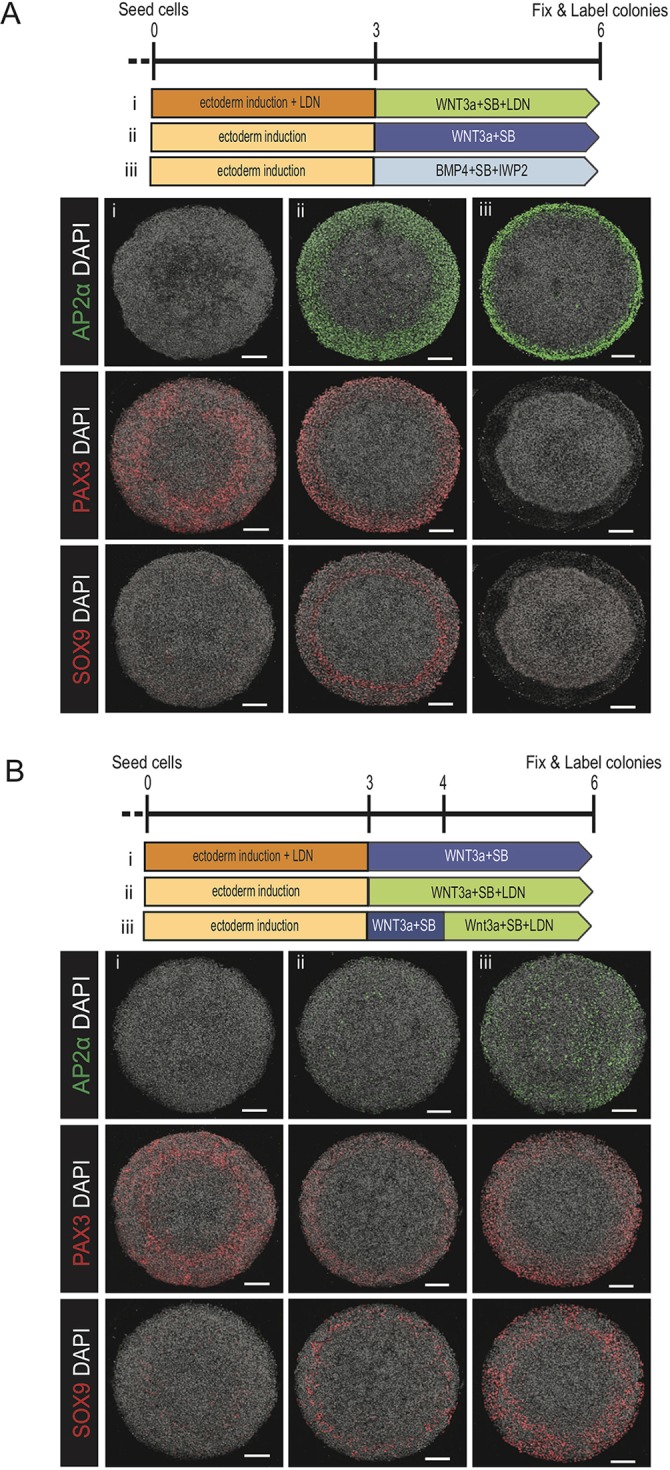
Dissecting the logic connecting BMP and WNT signaling to the transcription factors AP2a, PAX3 and SOX9. (A,B) Representative images of colonies immunostained for AP2α, PAX3 or SOX9. The induction schematic indicates the signaling conditions over the course of the experiment. In A: (i) 3 days SB+LDN followed by 3 days WNT3a+SB+LDN; (ii) 3 days SB followed by 3 days WNT3a+SB; (iii) 3 days SB followed by 3 days BMP4+SB+IWP2. In B: (i) 3 days SB+LDN followed by 3 days WNT3a+SB; (ii) 3 days SB followed by 3 days WNT3a+SB+LDN; (iii) 3 days SB followed by 1 day WNT3a+SB then 2 days WNT3a+LDN+SB. Each experiment was replicated three times. Colony diameter: 700 µm. Scale bars: 100 µm.
Lastly, we evaluated the temporal requirement for endogenous BMP signaling in generating neural crest at the colony borders in response to WNT stimulation (Fig. 7Bii,Biii). Introduction of the BMP inhibitor LDN concurrently or 24 h post-WNT3a stimulation reduced the expression of AP2α while maintaining that of PAX3 and SOX9 (Fig. 7Bii,Biii). Interestingly, the strength of AP2α reduction was dependent on the duration of BMP inhibition. These results suggest a previously unappreciated temporal requirement to specify neural crest fates: continuous endogenous BMP signaling is necessary to maintain AP2α expression; however, only a short duration in BMP signaling is required to initiate and maintain SOX9 once WNT is present. In the future, it will be interesting to determine whether the cells that are SOX9+PAX3+AP2α− are viable for further differentiation.
DISCUSSION
In this study, we developed a system in which hESCs create self-organized patterns of human neural, neural crest, placodal and epidermal progenitors on micropatterned surfaces with the same organization as found in vivo within the ectodermal germ layer along the medial-lateral axis (Table S1). Cells were first specified to ectoderm through an initial phase of SMAD2/3 inhibition, followed by a second phase in which patterning was initiated with BMP4, and a third phase in which a WNT inhibitor was combined with BMP4 stimulation to prevent widespread activation of WNT. We leveraged our in vitro system to dissect temporal aspects of signaling interactions that underlie patterning and to improve current directed differentiation protocols for placodal cells. We showed that increasing WNT signaling duration leads to the expansion of neural crest at the expense of placodal and neural fates, and that cells sense the relative amounts of WNT and BMP signaling in choosing between placodal and neural crest lineages. Finally, we used the in vitro system to dissect temporal relationships between WNT and BMP signaling and their link to governing the expression of neural crest, links that have been difficult to untangle precisely in model systems in vivo (Martik and Bronner, 2017). These results are summarized in the schematics found in Table S1.
Recently, it was shown that hESCs grown on micropatterns in neural differentiation media self-organize into two territories with neural cells in the center and neural crest cells at the periphery. The induction of neural crest at the colony edge relied on endogenous BMP signaling, and the authors suggested that BMP signaling is activated by mechanical effects at the colony edge (Xue et al., 2018). Micropatterns have also been used to study patterning within the neural cells themselves. During neural differentiation, cells organize themselves into rosette-like structures reminiscent of the neural tube (Elkabetz et al., 2008). This system provides an interesting opportunity to study the self-organizing properties of the neuro-epithelium; however, rosettes typically form in a variety of sizes in a disorganized fashion. A protocol was recently developed in which micropatterns are used so that a single rosette forms on each pattern, and the size of the patterns was shown to be a crucial control parameter (Knight et al., 2018). In the future, it will be interesting to investigate whether the center of our ectodermal micropatterns develops a rosette morphology with further differentiation, and whether self-organized mechanical cues play a role, especially as the WNT pathway plays a crucial role in our patterns and is a known mechanosensitive pathway (Benham-Pyle et al., 2015).
Although measurements of pSmad1/5/8 have proven difficult in mammalian embryos, experiments in zebrafish and Xenopus have revealed a dorsal-ventral gradient, comparable to human medial-lateral, in signaling activity (Schohl and Fagotto, 2002; Tucker et al., 2008). In Xenopus, experiments with dissociated animal cap cells have suggested that the concentration of BMP can elicit multiple fates in a dose-dependent manner (Wilson et al., 1997). Although here we observe a similar pattern of pSMAD1/5/8 along the radius of the micropatterned colonies, our results do not support a model in which cell fates along the medial-lateral axis are patterned exclusively by the BMP dose (Chhabra et al., 2018preprint; Nemashkalo et al., 2017). A minimum BMP4 concentration is necessary to initiate surface ectodermal differentiation; however, the decision to form neural crest or placodes requires the integration of BMP and WNT signals.
It is instructive to compare our results here with our previous work on self-organized germ layer differentiation in micropatterns. In both cases, patterning is initiated by an applied BMP4 signal, which combines with endogenous WNT signals. The edge fates (extra-embryonic for gastrulation micropatterns, surface ectoderm in this case) are upregulated by BMP signaling, whereas spatially intermediate fates (mesendoderm in the case of gastrulation, neural crest here), require the WNT signal. The fates at the center (ectoderm or neural) result from the inhibition of both signals. The essential difference in protocols is that the gastrulation patterns are generated directly from pluripotent hESCs whereas ectodermal patterns require a 3-day induction towards ectoderm before application of BMP4. The similarity in the signaling response in these two cases suggests the possibility that a core signaling network comprising the BMP and WNT pathways is responsible for both patterns while the competence of the cells shifts in time. This allows the same BMP-WNT signaling circuit to elicit different fates during the patterning of germ layers at gastrulation and the patterning of the ectoderm slightly later in development. A crucial difference, however, is the role of Nodal signaling. Although Nodal is essential for mesendoderm differentiation, it is inhibited throughout here. Given that cells are still capable of forming BRA-positive mesoderm after 3 days of ectodermal induction (Fig. 1H), it would be interesting to determine whether removing Nodal inhibition or exogenously adding Nodal ligands would convert some of the ectodermal cells to mesodermal fates, and whether cells would form patterns of ectoderm and mesoderm under these conditions.
We showed that cells do not respond to a particular level of BMP or WNT signaling, but instead determine fate as a function of these pathways combinatorially. Elevating WNT signaling drives neural crest differentiation at the expense of placodes, but the same levels of WNT are also compatible with placodal differentiation when BMP levels are also elevated. What is the role of this combinatorial control of cell fate? One possibility is that it is a means to allow diversification of cell fate by multiple pathways. This may be a more robust strategy than having cells read multiple levels off a single gradient. For example, rather than creating the four fates comprising the medial-lateral axis within the ectoderm by reading the precise levels of BMP4, cells may create the same four states by reading two pathways in a binary way. In agreement with this view, experimental and theoretical studies have questioned whether a single morphogen gradient or signaling pathway contains enough information to specify multiple fates (Cheong et al., 2011; Gregor et al., 2007). Additional sources of information may also be present in the signaling dynamics (Selimkhanov et al., 2014). More fine-grained mapping of the fate patterns as a function of signaling levels, as well as careful quantification of the signaling dynamics in each case will be required to understand fully this connection between signaling and cell fate.
The WNT pathway has been shown to play a crucial role in anterior-posterior (AP) axis formation in the gastrulating embryo (Arnold and Robertson, 2009; Kiecker and Niehrs, 2001; Yamaguchi, 2001). High WNT signaling is essential for primitive streak formation on the posterior side of the embryo, whereas WNT is inhibited anteriorly by secreted inhibitors from the anterior visceral endoderm (Arnold and Robertson, 2009; Kiecker and Niehrs, 2001; Yamaguchi, 2001). Later, ongoing WNT signaling is required in the tailbud, and in vitro studies have suggested that longer WNT exposure causes hESCs to adopt progressively more posterior fates (Knight et al., 2018). The shift in fate patterns that we observe when the duration of WNT signaling is modulated is consistent with this duration setting the position along the AP axis. When WNT signaling was inhibited throughout, the cell fate patterns resembled anterior ectoderm, and were composed of neural, placodal and future epidermal, but not neural crest, lineages. Permitting WNT activity generated ectodermal patterns consistent with the midbrain, which comprise all four of these lineages. As the principle markers we use here are for fates along the dorso-ventral (DV) axis, a more definitive assignment of AP position could be made by examining specific markers of AP position, such as OTX2, GBX2 and HOX genes. Recent studies support the idea that the posterior fates in the spinal cord are specified by a separate route from the anterior ones (Arnold and Robertson, 2009; Delile et al., 2019; Garriock et al., 2015; Gouti et al., 2014; Kiecker and Niehrs, 2001; Lippmann et al., 2015; Yamaguchi, 2001) even prior to germ layer specification, and it would be informative to determine protocols for generating ectodermal patterns that mimic the posterior of the embryo.
A large body of work has discovered numerous transcription factors that are involved in different stages of neural plate border development and neural crest differentiation. For example, knockdown experiments in model organisms have demonstrated a requirement for PAX3 and AP2α in the specification of neural crest fates as defined by SOX9/10 expression (Bae et al., 2014; Luo et al., 2003). It is believed that both the BMP and WNT signaling pathways are required for the maturation of neural crest, but it has been difficult to unravel how these pathways are embedded in the gene regulatory network that specifies the neural crest in vivo. We showed that the micropatterned system is an attractive platform for dissecting these interactions due to the ease of manipulation and the removal of confounding factors resulting from effects on earlier development or on other tissues. Specifically, that BMP and WNT signaling are each necessary and sufficient for the expression of particular genes, whereas others require both pathways. Markers of more mature neural crest fall into this last category. We also noted that as long as endogenous BMP signaling is allowed during the first phase of the protocol, WNT activation alone is sufficient to activate all of these genes, so the requirement for BMP signaling in neural crest differentiation is only transient, whereas that for WNT is ongoing. This approach can be extended to gain a fuller picture of how signaling pathways direct a complex network of transcription factors in the specification and function of the neural crest.
We have developed a novel system in which human embryonic stem cells generate the DV pattern within the ectoderm. This system can be used to gain an understanding of human ectodermal development and to improve directed differentiation protocols. Future studies can take advantage of this system together with live cell reporters of signaling and fate to understand in detail how ectodermal patterns are self-organized. Finally, our study provides proof of principle for extending the micropatterning approach beyond germ layer specification to later developmental events. As long as cells can be directed to adopt a progenitor cell fate, providing a reproducible geometric and chemical environment leads to patterning, and the patterns can then be tuned to yield a particular outcome. A similar approach could be used to generate patterns corresponding to endoderm or mesoderm, or to particular organs.
MATERIALS AND METHODS
Cell lines
All experiments were performed using ESI017 hESCS (obtained from ESI BIO, RRID: CVCL_B854, XX), RUES2 hESCs (a gift from Ali Brivanlou, Rockefeller University, NY, USA), or WTC-11 iPSCs (Paul Allen Institute, Cell Line ID: AICS-0058 cl.67). For live imaging of WNT signaling dynamics, the ESI017 GFP:β-catenin cell line as described by Massey et al. (2019) was used.
Routine cell culture
All cells were grown in the chemically defined medium mTeSR1 in tissue culture dishes and kept at 37°C, 5% CO2 as described by Nemashkalo et al. (2017). Cells were routinely passaged and checked for mycoplasma contamination also as described by Nemashkalo et al. (2017). In all experiments, cell passage number did not extend beyond 55.
Differentiation
Standard culture
ESI017 hESC cultures were washed twice with 1× PBS and disaggregated using accutase for 7 min at 37°C. Cells were subsequently harvested, centrifuged at 1000 rpm (300 g) for 4 min and seeded on laminin-521 coated Ibidi dishes at a density of 18,000 cells/cm2 in the presence of mTeSR media and ROCK inhibitor (ROCKi) (10 µM). On the following day ROCKi was withdrawn from the media, and hESCs were allowed to expand in culture for 2 days in mTeSR. All inductions were initiated on the third day in N2B27 medium, which we prepared by filtering a mixture of 250 ml DMEM/12, 250 ml Neurobasal media, 2.5 ml N2 supplement, 5 ml B27 supplement without vitamin A, 5 ml glutamax and 0.5 ml β-mercaptoethanol. N2B27 was supplemented with growth factors and small molecules as indicated. In all experiments, medium was replenished daily.
On micropatterns
For micropatterned cell culture experiments, micropatterned glass CYTOOchips or 96-well CYTOOplates were coated with 5 μg/ml laminin-521 in 1× PBS with calcium and magnesium (PBS++) for 2.5 h at 37°C. The laminin was then removed by first using serial dilutions without allowing the chip or well to dry (dilution 1:4 in PBS++, five times), and then using one complete wash with PBS++. The chip or plate was either used immediately or stored for up to 2 weeks at 4°C with PBS++ covering the micropatterned surfaces.
Seeding of hESCs onto micropatterned surfaces was performed as follows. Stem cell cultures were washed twice with 1× PBS and disaggregated using accurate for 7 min at 37°C. Cells were subsequently harvested, centrifuged at 1000 rpm (300 g) for 4 min and re-suspended in mTeSR containing ROCKi. Cells were then seeded to laminin-coated CYTOOchips (1×106 cells in 2 ml of media) or laminin-coated CYTOOplates (120,000 cell/well with 100 μl media/well), and then placed in the incubator for 45 min at 37°C. Cells were then washed twice with 1× PBS to remove ROCKi and cells bound nonspecifically to uncoated regions of the well or chip. To promote better adhesion to the micropatterned surfaces, all ectoderm inductions were initiated in the presence of mTeSR supplemented with SB. On the following day, mTeSR was withdrawn as the base media and replaced with N2B27 for the remainder of the experiment. Stem cell colonies were then further treated with reagents as described in the text. In all micropatterned experiments, the medium was replenished daily with the described signaling conditions and fixed at the indicated times within the text.
Immunostaining
Immunostaining followed standard protocols as described by Nemashkalo et al. (2017). Primary and secondary antibodies were diluted in the blocking solution as described by Warmflash et al. (2014) and Nemashkalo et al. (2017). Dilutions are listed in Table S2.
Imaging
Live cell imaging
GFP:β-catenin hESCs were seeded in multiple wells of a 96-well CYTOOplate and induced towards the ectoderm as described above. Four colonies (700 μm) per condition were imaged with a 20×, NA 0.75 objective on a Olympus/Andor spinning disk confocal microscope. Five z-sections were acquired per position every 30 min from 24 h pre- to 48 h post-BMP4 treatment. To remain consistent with fixed cell experiments, the imaging was halted for a brief period (∼1 h) every 24 h to replenish the media. In all cases, cells were maintained at 37°C and 5% CO2 throughout the duration of imaging.
Fixed cell imaging
All immunostaining data were acquired by imaging multiple z-sections at 10× (NA 0.40), 20× (NA 0.75) and 30× (NA 1.05) from fixed cell colonies on CYTOOchips or CYTOOplates with an Olympus FV1200 laser scanning confocal microscope.
Quantification and analyses
All micropatterning experiments were performed at least twice with consistent results whereas experiments in standard culture were performed three times. The data and analyses in each micropattern induction figure belong to one experiment whereas placode and ectoderm induction data belong to all three experiments. Sample size was not pre-determined and no statistical tests were used to determine significance of results on micropatterned colonies. Where indicated, statistical significance was determined using the two-sided t-test, implemented using the MATLAB function ttest2. Circular colonies with a non-radially symmetric cell density pattern at the end of treatment were excluded from analyses. The number of colonies included in each analysis (N) is mentioned in the figure legends. For images taken at 10× magnification with multiple z-slices, background subtraction, maximum z-projection and alignment were performed as described by Warmflash et al. (2014). The average intensity of a marker was calculated for each cell as the average of the immunofluorescence intensity in that cell normalized to the intensity of DAPI staining in the same cell. In cases in which the cells were too dense to perform cell segmentation effectively, averages were calculated over nuclear pixels rather than cells.
Software
MATLAB scripts for analyzing experimental data can be obtained from https://github.com/warmflasha/CellTracker.
Supplementary Material
Acknowledgements
We would like to thank Andrew Groves and members of the Warmflash lab for helpful discussions.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: G.B., A.W.; Methodology: G.B.; Software: I.H.; Validation: G.B.; Investigation: G.B., I.H., R.H.; Writing - original draft: G.B., A.W.; Writing - review & editing: G.B., I.H., R.H., A.A.Q., A.W.; Visualization: G.B.; Supervision: A.A.Q., A.W.; Project administration: A.A.Q., A.W.; Funding acquisition: A.A.Q., A.W.
Funding
This work was supported by the National Institute of General Medical Sciences (R01GM126122), the National Science Foundation (MCB-1553228 and ECCS-1533708), the Cancer Prevention and Research Institute of Texas (RR140073), the University of Texas STARS program and the Simons Foundation (511079). Deposited in PMC for release after 12 months.
Supplementary information
Supplementary information available online at http://dev.biologists.org/lookup/doi/10.1242/dev.179093.supplemental
References
- Ahrens K. and Schlosser G. (2005). Tissues and signals involved in the induction of placodal Six1 expression in Xenopus laevis. Dev. Biol. 288, 40-59. 10.1016/j.ydbio.2005.07.022 [DOI] [PubMed] [Google Scholar]
- Arnold S. J. and Robertson E. J. (2009). Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 10, 91-103. 10.1038/nrm2618 [DOI] [PubMed] [Google Scholar]
- Bae C.-J., Park B.-Y., Lee Y.-H., Tobias J. W., Hong C.-S. and Saint-Jeannet J.-P. (2014). Identification of Pax3 and Zic1 targets in the developing neural crest. Dev. Biol. 386, 473-483. 10.1016/j.ydbio.2013.12.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barry C., Schmitz M. T., Jiang P., Schwartz M. P., Duffin B. M., Swanson S., Bacher R., Bolin J. M., Elwell A. L., McIntosh B. E. et al. (2017). Species-specific developmental timing is maintained by pluripotent stem cells ex utero. Dev. Biol. 423, 101-110. 10.1016/j.ydbio.2017.02.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benham-Pyle B. W., Pruitt B. L. and Nelson W. J. (2015). Cell adhesion. Mechanical strain induces E-cadherin-dependent Yap1 and β-catenin activation to drive cell cycle entry. Science 348, 1024-1027. 10.1126/science.aaa4559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernardo A. S., Faial T., Gardner L., Niakan K. K., Ortmann D., Senner C. E., Callery E. M., Trotter M. W., Hemberger M., Smith J. C. et al. (2011). BRACHYURY and CDX2 mediate BMP-induced differentiation of human and mouse pluripotent stem cells into embryonic and extraembryonic lineages. Cell Stem Cell 9, 144-155. 10.1016/j.stem.2011.06.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betters E., Liu Y., Kjaeldgaard A., Sundström E. and García-Castro M. I. (2010). Analysis of early human neural crest development. Dev. Biol. 344, 578-592. 10.1016/j.ydbio.2010.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhat N., Kwon H.-J. and Riley B. B. (2013). A gene network that coordinates preplacodal competence and neural crest specification in zebrafish. Dev. Biol. 373, 107-117. 10.1016/j.ydbio.2012.10.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhatt S., Diaz R. and Trainor P. A. (2013). Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harb. Perspect. Biol. 5, a008326.. 10.1101/cshperspect.a008326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Böck J., Remmele C. W., Dittrich M., Müller T., Kondova I., Persengiev S., Bontrop R. E., Ade C. P., Kraus T. F. J., Giese A. et al. (2018). Cell type and species-specific patterns in neuronal and non-neuronal methylomes of human and chimpanzee cortices. Cereb. Cortex 28, 3724-3739. 10.1093/cercor/bhy180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brugmann S. A. and Moody S. A. (2005). Induction and specification of the vertebrate ectodermal placodes: precursors of the cranial sensory organs. Biol. Cell 97, 303-319. 10.1042/BC20040515 [DOI] [PubMed] [Google Scholar]
- Chambers S. M., Fasano C. A., Papapetrou E. P., Tomishima M., Sadelain M. and Studer L. (2009). Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat. Biotechnol. 27, 275-280. 10.1038/nbt.1529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen B., Dodge M. E., Tang W., Lu J., Ma Z., Fan C.-W., Wei S., Hao W., Kilgore J., Williams N. S. et al. (2009). Small molecule-mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nat. Chem. Biol. 5, 100-107. 10.1038/nchembio.137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheong R., Rhee A., Wang C. J., Nemenman I. and Levchenko A. (2011). Information transduction capacity of noisy biochemical signaling networks. Science 334, 354-358. 10.1126/science.1204553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chhabra S., Liu L., Goh R. and Warmflash A. (2018). Dissecting the dynamics of signaling events in the BMP, WNT, and NODAL cascade during self-organized fate patterning in human gastruloids. PLoS Biol. 10.1371/journal.pbio.3000498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi Y. S. and Gumbiner B. (1989). Expression of cell adhesion molecule E-cadherin in Xenopus embryos begins at gastrulation and predominates in the ectoderm. J. Cell Biol. 108, 2449-2458. 10.1083/jcb.108.6.2449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christophorou N. A. D., Bailey A. P., Hanson S. and Streit A. (2009). Activation of Six1 target genes is required for sensory placode formation. Dev. Biol. 336, 327-336. 10.1016/j.ydbio.2009.09.025 [DOI] [PubMed] [Google Scholar]
- de Crozé N., Maczkowiak F. and Monsoro-Burq A. H. (2011). Reiterative AP2a activity controls sequential steps in the neural crest gene regulatory network. Proc. Natl. Acad. Sci. USA 108, 155-160. 10.1073/pnas.1010740107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- del Barco Barrantes I., Davidson G., Gröne H.-J., Westphal H. and Niehrs C. (2003). Dkk1 and noggin cooperate in mammalian head induction. Genes Dev. 17, 2239-2244. 10.1101/gad.269103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delile J., Rayon T., Melchionda M., Edwards A., Briscoe J. and Sagner A. (2019). Single cell transcriptomics reveals spatial and temporal dynamics of gene expression in the developing mouse spinal cord. Development 146, dev173807 10.1242/dev.173807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dincer Z., Piao J., Niu L., Ganat Y., Kriks S., Zimmer B., Shi S.-H., Tabar V. and Studer L. (2013). Specification of functional cranial placode derivatives from human pluripotent stem cells. Cell Rep. 5, 1387-1402. 10.1016/j.celrep.2013.10.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ealy M., Ellwanger D. C., Kosaric N., Stapper A. P. and Heller S. (2016). Single-cell analysis delineates a trajectory toward the human early otic lineage. Proc. Natl. Acad. Sci. USA 113, 8508-8513. 10.1073/pnas.1605537113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elkabetz Y., Panagiotakos G., Al Shamy G., Socci N. D., Tabar V. and Studer L. (2008). Human ES cell-derived neural rosettes reveal a functionally distinct early neural stem cell stage. Genes Dev. 22, 152-165. 10.1101/gad.1616208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etoc F., Metzger J., Ruzo A., Kirst C., Yoney A., Ozair M. Z., Brivanlou A. H. and Siggia E. D. (2016). A balance between secreted inhibitors and edge sensing controls gastruloid self-organization. Dev. Cell 39, 302-315. 10.1016/j.devcel.2016.09.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fainsod A., Deissler K., Yelin R., Marom K., Epstein M., Pillemer G., Steinbeisser H. and Blum M. (1997). The dorsalizing and neural inducing gene follistatin is an antagonist of BMP-4. Mech. Dev. 63, 39-50. 10.1016/S0925-4773(97)00673-4 [DOI] [PubMed] [Google Scholar]
- García-Castro M. I., Marcelle C. and Bronner-Fraser M. (2002). Ectodermal Wnt function as a neural crest inducer. Science 297, 848-851. [DOI] [PubMed] [Google Scholar]
- Garnett A. T., Square T. A. and Medeiros D. M. (2012). BMP, Wnt and FGF signals are integrated through evolutionarily conserved enhancers to achieve robust expression of Pax3 and Zic genes at the zebrafish neural plate border. Development 139, 4220-4231. 10.1242/dev.081497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garriock R. J., Chalamalasetty R. B., Kennedy M. W., Canizales L. C., Lewandoski M. and Yamaguchi T. P. (2015). Lineage tracing of neuromesodermal progenitors reveals novel Wnt-dependent roles in trunk progenitor cell maintenance and differentiation. Development 142, 1628-1638. 10.1242/dev.111922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geary L. and LaBonne C. (2018). FGF mediated MAPK and PI3K/Akt signals make distinct contributions to pluripotency and the establishment of Neural Crest. eLife 7, e33845 10.7554/eLife.33845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez G. A., Prasad M. S., Sandhu N., Shelar P. B., Leung A. W. and García-Castro M. I. (2019). Human neural crest induction by temporal modulation of WNT activation. Dev. Biol. 449, 99-106. 10.1016/j.ydbio.2019.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gouti M., Tsakiridis A., Wymeersch F. J., Huang Y., Kleinjung J., Wilson V. and Briscoe J. (2014). In vitro generation of neuromesodermal progenitors reveals distinct roles for wnt signalling in the specification of spinal cord and paraxial mesoderm identity. PLoS Biol. 12, e1001937 10.1371/journal.pbio.1001937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gregor T., Tank D. W., Wieschaus E. F. and Bialek W. (2007). Probing the limits to positional information. Cell 130, 153-164. 10.1016/j.cell.2007.05.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groves A. K. and LaBonne C. (2014). Setting appropriate boundaries: fate, patterning and competence at the neural plate border. Dev. Biol. 389, 2-12. 10.1016/j.ydbio.2013.11.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackland J. O. S., Frith T. J. R., Thompson O., Marin Navarro A., Garcia-Castro M. I., Unger C. and Andrews P. W. (2017). Top-down inhibition of BMP signaling enables robust induction of hPSCs into neural crest in fully defined, xeno-free conditions. Stem Cell Rep. 9, 1043-1052. 10.1016/j.stemcr.2017.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heemskerk I., Burt K., Miller M., Chhabra S., Guerra M. C., Liu L. and Warmflash A. (2019). Rapid changes in morphogen concentration control self-organized patterning in human embryonic stem cells. eLife 8, e40526 10.7554/eLife.40526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoodless P. A., Haerry T., Abdollah S., Stapleton M., O'Connor M. B., Attisano L. and Wrana J. L. (1996). MADR1, a MAD-related protein that functions in BMP2 signaling pathways. Cell 85, 489-500. 10.1016/S0092-8674(00)81250-7 [DOI] [PubMed] [Google Scholar]
- Kawai S., Faucheu C., Gallea S., Spinella-Jaegle S., Atfi A., Baron R. and Roman S. R. (2000). Mouse smad8 phosphorylation downstream of BMP receptors ALK-2, ALK-3, and ALK-6 induces its association with Smad4 and transcriptional activity. Biochem. Biophys. Res. Commun. 271, 682-687. 10.1006/bbrc.2000.2704 [DOI] [PubMed] [Google Scholar]
- Kiecker C. and Niehrs C. (2001). A morphogen gradient of Wnt/beta-catenin signalling regulates anteroposterior neural patterning in Xenopus. Development 128, 4189-4201. [DOI] [PubMed] [Google Scholar]
- Knight G. T., Lundin B. F., Iyer N., Ashton L. M. T., Sethares W. A., Willett R. M. and Ashton R. S. (2018). Engineering induction of singular neural rosette emergence within hPSC-derived tissues. eLife 7, e37549 10.7554/eLife.37549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurek D., Neagu A., Tastemel M., Tüysüz N., Lehmann J., van de Werken H. J. G., Philipsen S., van der Linden R., Maas A., van IJcken W. F. J. et al. (2015). Endogenous WNT signals mediate BMP-induced and spontaneous differentiation of epiblast stem cells and human embryonic stem cells. Stem Cell Rep. 4, 114-128. 10.1016/j.stemcr.2014.11.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon H.-J., Bhat N., Sweet E. M., Cornell R. A. and Riley B. B. (2010). Identification of early requirements for preplacodal ectoderm and sensory organ development. PLoS Genet. 6, e1001133 10.1371/journal.pgen.1001133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung A. W., Kent Morest D. and Li J. Y. H. (2013). Differential BMP signaling controls formation and differentiation of multipotent preplacodal ectoderm progenitors from human embryonic stem cells. Dev. Biol. 379, 208-220. 10.1016/j.ydbio.2013.04.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung A. W., Murdoch B., Salem A. F., Prasad M. S., Gomez G. A. and García-Castro M. I. (2016). WNT/β-catenin signaling mediates human neural crest induction via a pre-neural border intermediate. Development 143, 398-410. 10.1242/dev.130849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L., Liu C., Biechele S., Zhu Q., Song L., Lanner F., Jing N. and Rossant J. (2013). Location of transient ectodermal progenitor potential in mouse development. Development 140, 4533-4543. 10.1242/dev.092866 [DOI] [PubMed] [Google Scholar]
- Li L., Song L., Liu C., Chen J., Peng G., Wang R., Liu P., Tang K., Rossant J. and Jing N. (2015). Ectodermal progenitors derived from epiblast stem cells by inhibition of Nodal signaling. J. Mol. Cell Biol. 7, 455-465. 10.1093/jmcb/mjv030 [DOI] [PubMed] [Google Scholar]
- Lippmann E. S., Williams C. E., Ruhl D. A., Estevez-Silva M. C., Chapman E. R., Coon J. J. and Ashton R. S. (2015). Deterministic HOX patterning in human pluripotent stem cell-derived neuroectoderm. Stem Cell Rep. 4, 632-644. 10.1016/j.stemcr.2015.02.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Litsiou A., Hanson S. and Streit A. (2005). A balance of FGF, BMP and WNT signalling positions the future placode territory in the head. Development 132, 4051-4062. 10.1242/dev.01964 [DOI] [PubMed] [Google Scholar]
- Liu C., Wang R., He Z., Osteil P., Wilkie E., Yang X., Chen J., Cui G., Guo W., Chen Y. et al. (2018). Suppressing nodal signaling activity predisposes ectodermal differentiation of epiblast stem cells. Stem Cell Rep. 11, 43-57. 10.1016/j.stemcr.2018.05.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo T., Matsuo-Takasaki M., Thomas M. L., Weeks D. L. and Sargent T. D. (2002). Transcription factor AP-2 is an essential and direct regulator of epidermal development in Xenopus. Dev. Biol. 245, 136-144. 10.1006/dbio.2002.0621 [DOI] [PubMed] [Google Scholar]
- Luo T., Lee Y.-H., Saint-Jeannet J.-P. and Sargent T. D. (2003). Induction of neural crest in Xenopus by transcription factor AP2alpha. Proc. Natl. Acad. Sci. USA 100, 532-537. 10.1073/pnas.0237226100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malaguti M., Nistor P. A., Blin G., Pegg A., Zhou X. and Lowell S. (2013). Bone morphogenic protein signalling suppresses differentiation of pluripotent cells by maintaining expression of E-Cadherin. eLife 2, e01197 10.7554/eLife.01197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansouri A., Pla P., Larue L. and Gruss P. (2001). Pax3 acts cell autonomously in the neural tube and somites by controlling cell surface properties. Development 128, 1995-2005. [DOI] [PubMed] [Google Scholar]
- Martik M. L. and Bronner M. E. (2017). Regulatory logic underlying diversification of the neural crest. Trends Genet. 33, 715-727. 10.1016/j.tig.2017.07.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massey J., Liu Y., Alvarenga O., Saez T., Schmerer M. and Warmflash A. (2019). Synergy with TGFβ ligands switches WNT pathway dynamics from transient to sustained during human pluripotent cell differentiation. Proc. Natl. Acad. Sci. USA 116, 4989-4998. 10.1073/pnas.1815363116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell P. J., Timmons P. M., Hébert J. M., Rigby P. W. and Tjian R. (1991). Transcription factor AP-2 is expressed in neural crest cell lineages during mouse embryogenesis. Genes Dev. 5, 105-119. 10.1101/gad.5.1.105 [DOI] [PubMed] [Google Scholar]
- Monsoro-Burq A.-H., Wang E. and Harland R. (2005). Msx1 and Pax3 cooperate to mediate FGF8 and WNT signals during Xenopus neural crest induction. Dev. Cell 8, 167-178. 10.1016/j.devcel.2004.12.017 [DOI] [PubMed] [Google Scholar]
- Nemashkalo A., Ruzo A., Heemskerk I. and Warmflash A. (2017). Morphogen and community effects determine cell fates in response to BMP4 signaling in human embryonic stem cells. Development 144, 3042-3053. 10.1242/dev.153239 [DOI] [PubMed] [Google Scholar]
- Nishimura R., Kato Y., Chen D., Harris S. E., Mundy G. R. and Yoneda T. (1998). Smad5 and DPC4 are key molecules in mediating BMP-2-induced osteoblastic differentiation of the pluripotent mesenchymal precursor cell line C2C12. J. Biol. Chem. 273, 1872-1879. 10.1074/jbc.273.4.1872 [DOI] [PubMed] [Google Scholar]
- Ozair M. Z., Noggle S., Warmflash A., Krzyspiak J. E. and Brivanlou A. H. (2013). SMAD7 directly converts human embryonic stem cells to telencephalic fate by a default mechanism. Stem Cells 31, 35-47. 10.1002/stem.1246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page M. (1989). Changing patterns of cytokeratins and vimentin in the early chick embryo. Development 105, 97-107. [DOI] [PubMed] [Google Scholar]
- Papanayotou C., Mey A., Birot A.-M., Saka Y., Boast S., Smith J. C., Samarut J. and Stern C. D. (2008). A mechanism regulating the onset of Sox2 expression in the embryonic neural plate. PLoS Biol. 6, e2 10.1371/journal.pbio.0060002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piacentino M. L. and Bronner M. E. (2018). Intracellular attenuation of BMP signaling via CKIP-1/Smurf1 is essential during neural crest induction. PLoS Biol. 16, e2004425 10.1371/journal.pbio.2004425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pieper M., Ahrens K., Rink E., Peter A. and Schlosser G. (2012). Differential distribution of competence for panplacodal and neural crest induction to non-neural and neural ectoderm. Development 139, 1175-1187. 10.1242/dev.074468 [DOI] [PubMed] [Google Scholar]
- Plouhinec J.-L., Medina-Ruiz S., Borday C., Bernard E., Vert J.-P., Eisen M. B., Harland R. M. and Monsoro-Burq A. H. (2017). A molecular atlas of the developing ectoderm defines neural, neural crest, placode, and nonneural progenitor identity in vertebrates. PLoS Biol. 15, e2004045 10.1371/journal.pbio.2004045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu Y., Zhou B., Yang W., Han B., Yu-Rice Y., Gao B., Johnson J., Svendsen C. N., Freeman M. R., Giuliano A. E. et al. (2016). Transcriptome and proteome characterization of surface ectoderm cells differentiated from human iPSCs. Sci. Rep. 6, 32007 10.1038/srep32007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sasai Y., Lu B., Steinbeisser H. and De Robertis E. M. (1995). Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus. Nature 377, 757 10.1038/377757a0 [DOI] [PubMed] [Google Scholar]
- Sato T., Sasai N. and Sasai Y. (2005). Neural crest determination by co-activation of Pax3 and Zic1 genes in Xenopus ectoderm. Development 132, 2355-2363. 10.1242/dev.01823 [DOI] [PubMed] [Google Scholar]
- Schlosser G. (2006). Induction and specification of cranial placodes. Dev. Biol. 294, 303-351. 10.1016/j.ydbio.2006.03.009 [DOI] [PubMed] [Google Scholar]
- Schohl A. and Fagotto F. (2002). Beta-catenin, MAPK and Smad signaling during early Xenopus development. Development 129, 37-52. [DOI] [PubMed] [Google Scholar]
- Selimkhanov J., Taylor B., Yao J., Pilko A., Albeck J., Hoffmann A., Tsimring L. and Wollman R. (2014). Systems biology. Accurate information transmission through dynamic biochemical signaling networks. Science 346, 1370-1373. 10.1126/science.1254933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simões-Costa M. and Bronner M. E. (2015). Establishing neural crest identity: a gene regulatory recipe. Development 142, 242-257. 10.1242/dev.105445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simões-Costa M., Stone M. and Bronner M. E. (2015). Axud1 integrates Wnt signaling and transcriptional inputs to drive neural crest formation. Dev. Cell 34, 544-554. 10.1016/j.devcel.2015.06.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith W. C. and Harland R. M. (1992). Expression cloning of noggin, a new dorsalizing factor localized to the Spemann organizer in Xenopus embryos. Cell 70, 829-840. 10.1016/0092-8674(92)90316-5 [DOI] [PubMed] [Google Scholar]
- Smith J. R., Vallier L., Lupo G., Alexander M., Harris W. A. and Pedersen R. A. (2008). Inhibition of Activin/Nodal signaling promotes specification of human embryonic stem cells into neuroectoderm. Dev. Biol. 313, 107-117. 10.1016/j.ydbio.2007.10.003 [DOI] [PubMed] [Google Scholar]
- Spokony R. F., Aoki Y., Saint-Germain N., Magner-Fink E. and Saint-Jeannet J.-P. (2002). The transcription factor Sox9 is required for cranial neural crest development in Xenopus. Development 129, 421-432. [DOI] [PubMed] [Google Scholar]
- Stuhlmiller T. J. and García-Castro M. I. (2012). FGF/MAPK signaling is required in the gastrula epiblast for avian neural crest induction. Development 139, 289-300. 10.1242/dev.070276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tchieu J., Zimmer B., Fattahi F., Amin S., Zeltner N., Chen S. and Studer L. (2017). A modular platform for differentiation of human PSCs into all major ectodermal lineages. Cell Stem Cell 21, 399-410.e7. 10.1016/j.stem.2017.08.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker J. A., Mintzer K. A. and Mullins M. C. (2008). The BMP signaling gradient patterns dorsoventral tissues in a temporally progressive manner along the anteroposterior axis. Dev. Cell 14, 108-119. 10.1016/j.devcel.2007.11.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villanueva S., Glavic A., Ruiz P. and Mayor R. (2002). Posteriorization by FGF, Wnt, and retinoic acid is required for neural crest induction. Dev. Biol. 241, 289-301. 10.1006/dbio.2001.0485 [DOI] [PubMed] [Google Scholar]
- Wang Z., Oron E., Nelson B., Razis S. and Ivanova N. (2012). Distinct lineage specification roles for NANOG, OCT4, and SOX2 in human embryonic stem cells. Cell Stem Cell 10, 440-454. 10.1016/j.stem.2012.02.016 [DOI] [PubMed] [Google Scholar]
- Warmflash A., Sorre B., Etoc F., Siggia E. D. and Brivanlou A. H. (2014). A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat. Methods 11, 847-854. 10.1038/nmeth.3016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson P. A., Lagna G., Suzuki A. and Hemmati-Brivanlou A. (1997). Concentration-dependent patterning of the Xenopus ectoderm by BMP4 and its signal transducer Smad1. Development 124, 3177-3184. [DOI] [PubMed] [Google Scholar]
- Xu R.-H., Chen X., Li D. S., Li R., Addicks G. C., Glennon C., Zwaka T. P. and Thomson J. A. (2002). BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat. Biotechnol. 20, 1261-1264. 10.1038/nbt761 [DOI] [PubMed] [Google Scholar]
- Xue X., Sun Y., Resto-Irizarry A. M., Yuan Y., Aw Yong K. M., Zheng Y., Weng S., Shao Y., Chai Y., Studer L. et al. (2018). Mechanics-guided embryonic patterning of neuroectoderm tissue from human pluripotent stem cells. Nat. Mater. 17, 633-641. 10.1038/s41563-018-0082-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi T. P. (2001). Heads or tails: Wnts and anterior-posterior patterning. Curr. Biol. 11, R713-R724. 10.1016/S0960-9822(01)00417-1 [DOI] [PubMed] [Google Scholar]
- Yardley N. and García-Castro M. I. (2012). FGF signaling transforms non-neural ectoderm into neural crest. Dev. Biol. 372, 166-177. 10.1016/j.ydbio.2012.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ybot-Gonzalez P., Gaston-Massuet C., Girdler G., Klingensmith J., Arkell R., Greene N. D. E. and Copp A. J. (2007). Neural plate morphogenesis during mouse neurulation is regulated by antagonism of Bmp signalling. Development 134, 3203-3211. 10.1242/dev.008177 [DOI] [PubMed] [Google Scholar]
- Yu P. B., Deng D. Y., Lai C. S., Hong C. C., Cuny G. D., Bouxsein M. L., Hong D. W., McManus P. M., Katagiri T., Sachidanandan C. et al. (2008). BMP type I receptor inhibition reduces heterotopic ossification. Nat. Med. 14, 1363-1369. 10.1038/nm.1888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu P., Pan G., Yu J. and Thomson J. A. (2011). FGF2 sustains NANOG and switches the outcome of BMP4-induced human embryonic stem cell differentiation. Cell Stem Cell 8, 326-334. 10.1016/j.stem.2011.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X., Huang C. T., Chen J., Pankratz M. T., Xi J., Li J., Yang Y., Lavaute T. M., Li X.-J., Ayala M. et al. (2010). Pax6 is a human neuroectoderm cell fate determinant. Cell Stem Cell 7, 90-100. 10.1016/j.stem.2010.04.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.