

Abstract
Multiple sclerosis (MS) is a demyelinating disease that causes chronic inflammation in the central nervous system. The aim of this study was to investigate the effects of apamin administration on myelination process. MS was induced by feeding cuprizone pellets (0.2%) for 6 weeks (demyelination phase) followed by normal feeding for additional 2 weeks (remyelination phase). Briefly, C57BL/6 male mice were randomly divided into six groups. Group 1, received the regular food pellets. Group 2 contained two subgroups of 6 animals each (n = 2 × 6). First group received cuprizone for 6 weeks and the sacrificed while the second group after 6 weeks of cuprizone, received no treatment for additional 2 weeks. Group 3 (co-treatment group) was composed of two subgroups of 6 animals each (n = 2 × 6). Both subgroups received apamin (100 μg/kg) intraperitoneally twice a week for 6 weeks. First subgroup terminated at this time and the second subgroup was fed normal diet for two additional weeks. Group 4 (post-treatment, n = 6) received apamin (100 μg/kg) intraperitoneally twice a week for 2 weeks after cuprizone secession. Groups 5 and 6 (vehicle, n = 6 in each group) received phosphate buffered saline as the vehicle of apamin during demyelination and remyelination phase. At the end of each phase, mice were deeply anesthetized and perfused. Groups 5 and 6 (vehicle) received PBS as the vehicle during both phases. Mice were anesthetized, perfused with PBS through their heart, and their brains were removed. Brain sections stained with luxol fast blue and the images were analyzed. Apamin co-treatment significantly increased the myelin content as compared to the cuprizone group. Also, mild elevation in the myelinated areas was observed with apamin post-treatment in comparison with remyelination phase. Our results revealed that apamin prevents myelin destruction more significantly as compared to remyelination process. This observation explains the possible role of apamin in inhibiting the activation of the microglia cells than stimulation of the oligodendrocytic precursor cells.
Keywords: Apamin, C57BL/6, Cuprizone, Multiple sclerosis, Myelin
INTRODUCTION
Multiple sclerosis (MS) is a demyelinating disease that causes chronic inflammation in the central nervous system (1,2). MS especially affects young adults and specified by lots of inflammatory cells and demyelinated lesions in the white matter of brain (1,3). The pathophysiology of MS is related to a functional disturbance in the immune system which leads to axonal damages (4,5). Although the exact mechanism of MS has not been yet determined (6), oligodendrocytes are involved in repairing and regenerating myelin in lesions. It is likely that the failure in myelin restoration is caused by the lack of sufficiently formed new oligodendrocytes (7).
Bee venom is a clear liquid that is soluble in water with a pH around 5 to 5.5 (8). It contains various compounds such as peptide and non-peptide components, enzymes, biogenic amines, and phospholipids.
Melittin, the most prevalent substance of bee venom, is a peptide with anti-inflammatory and cytotoxic effects (9). Apamin is the second most abundant peptide in bee venom (2-3% of dry bee venom) which is characterized by neuroprotective effects and ability to cross the blood-brain barrier (9). Apamin selectively blocks axonal potassium channels named SK channels, a type of Ca2+ activated K+ channels that is especially expressed in central nervous system. These channels play an important role in controlling repetitive activity in central nervous system (10). Furthermore, apamin inhibits Kv1.3 channels that are expressed in various tissues such as immune system and microglia cells (11).
Recent researches demonstrated that treatment with whole bee venom could be effective against MS through anti- inflammatory properties in experimental autoimmune encephalomyelitis (12). Considering proven effects of apamin on the ion channels and its ability to cross blood-brain barrier, in this research we studied the effects of apamin administration on myelin content in cuprizone-induced MS animal model.
MATERIAL AND METHODS
Animals
C57BL/6 mice were obtained from Royan institute (Isfahan, I.R. Iran) and kept in individually ventilated cages under standard conditions of 12/12-h light/dark cycles and controlled humidity with ad libitum access to food and water. All procedures were approved by Iran National committee for Ethics in Biomedical Research (ethical approval ID: IR.MUI.RESEARCH.REC.1397.387) which was performed in accordance with the Guidlines for the Care and Use of Laboratory Animals. Possible efforts were made to decrease animal number and distress.
Experimental design
Male eight-week old C57BL/6 mice were randomly divided into six groups. Group 1 (negative control, n = 6) received the regular food pellets during the study and served as a healthy group. Rest of the animals were fed with pellets containing 0.2% (w/w) cuprizone (Sigma-Aldrich, USA) for 6 weeks to induce MS model of demyelination in the corpus callosum (demyelination phase) and those not sacrificed were fed with normal diet pellets for 2 additional weeks (remyelination phase). Group 2 composed of two subgroups of 6 animals each (n = 2 × 6) was recruited to evaluate the effect of cuprizone administration on demyelination after 6 weeks (first group sacrificed at this point) and remyelination (other group) after 2 weeks without any particular treatment. Group 3 (co-treatment group) composed of two subgroups of 6 animals each (n = 2 × 6). Both subgroups received apamin (100 μg/kg) intraperitoneally twice a week for 6 weeks. First subgroup terminated at this time and the second subgroup was fed normal diet for two additional weeks. Group 4 (post-treatment, n = 6) received apamin (100 μg/kg) intraperitoneally twice a week for 2 weeks after cuprizone secession. Groups 5 and 6 (vehicle, n = 6 in each group) received phosphate buffered saline as the vehicle of apamin during demyelination and remyelination phases. Changes in body weight, water, and food intake were measured weekly in each group. At the end of each phase, mice were deeply anesthetized and perfused.
Cardiac perfusion
Mice were anesthetized with ketamine/ xylazine (80/10 mg/kg, intraperitoneally), and cardiac perfusion was done with 5 mL phosphate buffered saline to clear the blood from the circulatory system and continued with 5 mL of 4% neutral formalin. At the end of perfusion, the animal was decapitated and the brain was removed immediately from cranium and stored in cold fixative for 24 h (13).
Tissue processing
Samples were placed in tissue processor (Sakura Tissue Tek VIP E150, Sakura, Japan) and dehydrated with growing concentration alcohol baths followed by toluene clearing step to replace trapped alcohols inside the tissue.
Finally, tissue samples were solidified through a hot (44-60 °C) wax chamber. Then serial sections (5 μm) from the mid part of the corpus callosum were obtained and the best one was used for staining (14).
Myelin staining
Tissue sections were analyzed using luxol fast blue staining (Sigma, USA). Samples were placed in luxol fast blue overnight at 60 °C and washed with 95% alcohol followed by distilled water to remove the excess blue stain. The blue color was trapped in myelinated white matter after lithium carbonate exposure step (20 sec). Slides were washed three times with 75% alcohol and the background was stained with cresyl violet for 1 min. With this technique myelinated fibers (blue), neutrophils (pink) and nerve cells (purple) were visualized with different colors (15).
Imaging and myelin quantification
Images were taken using Nikon Eclipse 55i light microscope (Nikon, Japanese) with a 4× objective magnification and were processed with NIH Fiji software (National Institute of Health, Bethesda, Maryland, USA). To evaluate myelin changes, at first background was subtracted from images and then colors were separated through a defined region of interest using color deconvolution plugin. Density of all samples median corpus callusom was quantified three times with equation below:
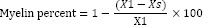
where, X1 is the mean of measured integrity density of normal control and Xs is the same region of interest integrated density of each sample (16).
Statistical analysis
All data are presented as means ± standard deviation and analyzed with Shapiro-wilk for normal distribution. Statistical differences between various groups were analyzed by analysis of variance (ANOVA) followed by Bonferroni’s Multiple Comparison post hoc test using GraphPad Prism software (La Jolla, CA, USA). Spearman correlation test was used to determine the relationship between measured variables. Values of P ≤ 0.05 were defined as statistically significant.
RESULTS
To set up the model of axonal damage over the time, we performed 6-week cuprizone exposure in C57BL/6 mouse model followed by the remyelination process for 2 weeks. Signs of disease induction including loss of appetite, reduced physical activity, tail or limb paralysis, and further physical changes were not seen throughout the study. Interestingly more physical activity and climbing behavior were seen at the beginning of the exposure.
A total weight change between animals during the course of the study is shown in Fig. 1A. The weight of animals in cuprizone group was significantly decreased as compared with negative control during the first week after the beginning of the study. However, during the 8 weeks of study a positive correlation between time and weight gain in all groups was observed (r, 0.93-0.98). During the study the average food (g/g BW) and water intake (mL/g BW) of animals in each group were measured per week. As it is shown in Fig. 1B and 1C food and water intake in cuprizone-exposed animals were lower than negative control during the first three weeks.
Fig. 1.
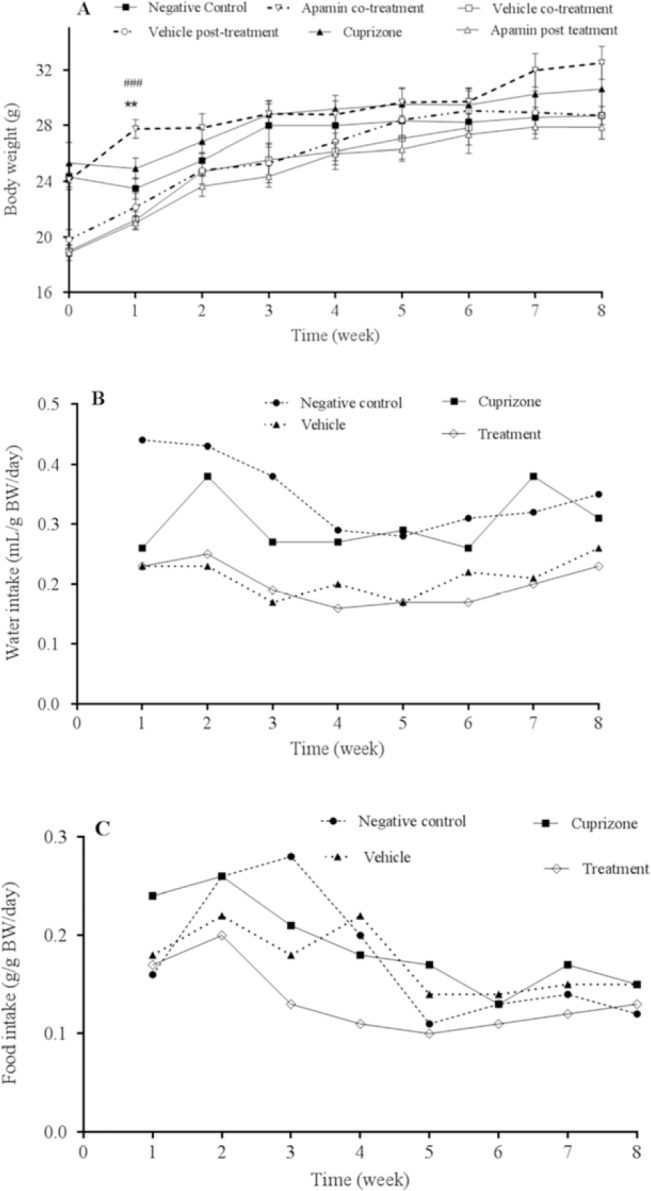
Effect of apamin (100 μg/kg, intraperitoneally) administration during and after cuprizone exposure on (A) body weight, (B) water, and (C) food intake of C57BL/6 mice multiple sclerosis model. Mice were weighed once a week during the study. **Shows significant difference (P < 0.01) between apamin co-treated animals and the vehicle group; and ### indicates significant difference (P < 0.001) between apamin co-treated and apamin post-treated subjects.
Axonal myelination content was determined after deconvolution of the loxul fast blue stained images of brain sections (Fig. 2A-2G). A significant decrease in median corpus callusom density of white matter fibers was observed after 6 weeks exposure to cuprizone compared with the negative control (Fig. 2A, 2B). Apamin treatment (Fig. 2D, 2H) inhibited the excessive demyelination during the demyelination phase (P < 0.001). Apamin also improved the remyelination and increased myelinated axons (Fig. 2G) in comparison with the cuprizone group (Fig. 2E, 2I) in remyelination phase (P < 0.05). Animals in demyelination phase after 6 weeks exposure to cuprizone had about 83% demyelination in median corpus callosum compared with the control group (Fig. 2H). This was followed by a rapid remyelination within 2 weeks after cuprizone withdrawal (Fig. 2I).
Fig. 2.

Effect of apamin (100 μg/kg, intraperitoneally) treatment or phosphate buffered saline (vehicle group) on cuprizone-induced demyelination after 6 weeks (demyelination phase) or 2 weeks of remyelination (remyelination phase). In comparison with the (A) negative control (healthy group), significant myelin loss reveals in the (B) cuprizone-exposed animals at the end of demyelination phase. (C) Indicates the vehicle-administered animals at the end of demyelination phase. Significant restoration of myelin was observed in apamin treated groups during (D) demyelination phase and (G) remyelination phase. (E and F) Show cuprizone and vehicle group corpus callusom after remyelination phase respectively. Also, the calculated means of myelin contents in demyelination and remyelination phase between groups are shown in part H and I. Data represented as mean ± SD (n = 6 in each group). *** Indicates significant differences in comparison with control group, P ≤ 0.001. #P ≤ 0.05 and ###P < 0.001 show significant differences compared with apamin-treated groups.
DISCUSSION
The cuprizone-induced MS is one of the best animal models for evaluation of the de- and remyelination process. Several factors such as the strain and age of the mouse (8-10 weeks), duration of the treatment (5-6 weeks), and the selected dose of cuprizone play critical roles in this model (17). In this study, body weight of animals in all groups increased over time. The average food intake in cuprizone exposed animals was decreased during the time of exposure.
Contrary to our results, Morell et al. reported that the activity of the animal and their body weight during the first week of the exposure to cuprizone was reduced but did not cause any problems with the animal eating or drinking and despite the persistent cuprizone exposure, the animals retained their body weight and recovered their activity at the end of the study (18). In addition, an increase in the relative rate of movement and a decrease in feces excretion was observed after 2 weeks of cuprizone exposure. These findings are consistent with previous published study indicating that cuprizone increased the activity of animals and decreased anxiety during the first three weeks of exposure. Also, it was suggested that the mice movement and distance traveled was significantly reduced following treatment with cuprizone (19).
Cuprizone is a copper chelator which inhibits cytochrome oxidase and monoamine oxidase functions which leads to megamitochondria induction. It is assumed that mitochondrial disturbances in the energy-producing cycle is the main cause of oligodendrocytes apoptosis in which oligodendrocytes main function is insulation of the neural cells with myelin sheath (20). Procaccini et al. confirmed this hypothesis and reported that the death of oligodendrocytes started at the end of the first week of cuprizone exposure and lasted for up to 6 weeks (21). Our findings are in accordance with the previous works showing that in the positive control group myelin content in the corpus callosum was reduced to 19% after 6 weeks of cuprizone exposure.
It was suggested that two weeks after initiation of demyelination, proliferation and aggregation of oligodendrocytic precursor cells in the subventricular zone and demyelinated areas are visible resulting in a beginning of a remyelination process (22). Similarly, we observed that after cuprizone withdrawal, the proportion of myelin content was increased after two weeks from 17% to 70% approximately.
Dry bee venom consists of about 2-3% apamin peptide with an 18 amino acids rigid sequence of H-Cys-Asn-Cys-Lys-Ala-Pro-Glu- Thr-Ala-Leu-Cys-Ala-Arg-Arg-Cys-Gln-Gln- His-NH2. The venom contains two disulfide bonds between Cys1-Cys11 and Cys3-Cys15 with great stability at various pH, temperature, and serum protease exposure conditions (23,24). Unlike other components of the bee venom, this peptide has the ability to cross the blood-brain barrier (25). In this regard, Fischer et al. stated that apamin plays a pivotal protecting role on dopaminergic neurons while the rest of the venom components increase such effects (23). Also, in 2013 a study showed the protective effect of honey bee venom and its apamin peptide in Parkinson’s induced model with 1-methyl-4- phenyl,1,2,3, and 6-tetrahydropyridine.
Furthermore, apamin significantly reduces the activity of inflammatory cytokines and adhesion molecules, such as vascular cell adhesion molecular-1 and intracellular cell adhesion molecule-1 in THP-1 derived macrophages. In addition, apamin suppressed the expression of vascular cell adhesion molecular-1, intracellular cell adhesion molecule-1, transforming growth factor beta, and fibronectin in mice with atherosclerosis and reduced the penetration of macrophages into the vessel wall. Apamin also decreases the phosphorylation of IκB dose-dependently and inhibits nuclear factor kappa B activation at nuclear level (26).
Several studies have indicated the presence of calcium-dependent potassium channels (SK/KCa2) in microglia and T cells, indicating the role of these channels in inflammation and neuronal degeneration processes. As a result, apamin may exert its neuroprotective effect through inhibition of these channels (27). Apamin inhibited Kv1.3 channels, in addition to the SK channels. It has also been proven that the concentration of 10 nM of this peptide is enough to inhibit half of the Kv1.3 channels (11). Fordyce et al. suggested that Kv1.3 channels play an important role in T cells, platelets, microglia and macrophages. As a result, Kv1.3 channels inhibition can cause depolarization and inhibition of the activity of immune responses (28). Active microglia appear about two weeks before detectable demyelination in the corpus callosum, cortex, and hippocampus. With this regard, Benton et al. have shown that apamin at 12 nM-100 pM can inhibit 50% of SK channels function. Blocking of these channels in microglia also reduces microglial activation and prevents neurotoxicity (29). These findings are in accordance with our results showing that apamin treatment decreased demyelination after 6 weeks of cuprizone exposure Fig. 2D.
On the other hand, oligodendrocytes express different types of potassium channels at their surface such as Kv 1.3 channels and the reduction of the expression of these potassium channels at the level of oligodendrocytes is essential for the synthesis of myelin structural proteins because inhibiting the outward potassium ion current leads to differentiation and survival of oligodendrocytes. It should be noted that oligodendrocytes are supportive cells in the central nervous system and are responsible for the production of myelin. It has been proven that the activation of these channels is essential for the planned cell death, and an increase in the removal of potassium ions from the neuronal cell causes neuronal apoptosis (30). Most likely inhibition of the above mentioned channels with apamin could enhance myelin content that could be the result of microglia cells inhibition or oligodendrocytes stimulation and needs further evaluation.
CONCLUSION
Our findings suggest the protective role of apamin against neurotoxicity following exposure to cuprizone with an emphasis on corpus callosum myelin content. Apamin exhibited higher activity in preventing myelin reduction compared with the administration at the remyelination process. This observation explains the critical role of apamin in inhibiting the activation of the microglia cells than stimulation of the oligodendrocytic precursor cells for increasing myelination in the recovery phase. Further investigations on the expression of the involved proteins and the migration of oligodendrocytic precursor cells during apamin exposure are warranted.
ACKNOWLEDGMENTS
This research was financially supported by the Vice Chancellery of Rsearch of Isfahan University of Medical Sciences, Isfahan, I.R. Iran, through the Grant No. 396818. Authors would like to thank Dr. Mehdi Eftekhari (Azarmehr pathology laboratory, Isfahan, I.R. Iran) who greatly assisted this research in brain section processing.
REFERENCES
- 1.Olsson T, Barcellos LF, Alfredsson L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat Rev Neurol. 2017;13(1):25–36. doi: 10.1038/nrneurol.2016.187. [DOI] [PubMed] [Google Scholar]
- 2.Aliomrani M, Sahraian MA, Shirkhanloo H, Sharifzadeh M, Khoshayand MR, Ghahremani MH. Correlation between heavy metal exposure and GSTM1 polymorphism in Iranian multiple sclerosis patients. Neurol Sci. 2017;38(7):1271–1278. doi: 10.1007/s10072-017-2934-5. [DOI] [PubMed] [Google Scholar]
- 3.Rezapour-Firouzi S, Shahabi S, Mohammadzadeh A, Tehrani AA, Kheradmand F, Mazloomi E. The potential effects of hemp seed/evening primrose oils on the mammalian target of rapamycin complex 1 and interferon-gamma genes expression in experimental autoimmune encephalomyelitis. Res Pharm Sci. 2018;13(6):523–532. doi: 10.4103/1735-5362.245964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gold R, Wolinsky JS. Pathophysiology of multiple sclerosis and the place of teriflunomide. Acta Neurol Scand. 2011;124(2):75–84. doi: 10.1111/j.1600-0404.2010.01444.x. [DOI] [PubMed] [Google Scholar]
- 5.Aliomrani M, Sahraian MA, Shirkhanloo H, Sharifzadeh M, Khoshayand MR, Ghahremani MH. Blood concentrations of cadmium and lead in multiple sclerosis patients from Iran. Iran J Pharm Res. 2016;15(4):825–833. [PMC free article] [PubMed] [Google Scholar]
- 6.Greenstein JI. Current concepts of the cellular and molecular pathophysiology of multiple sclerosis. Dev Neurobiol. 2007;67(9):1248–1265. doi: 10.1002/dneu.20387. [DOI] [PubMed] [Google Scholar]
- 7.Dawson MR, Levine JM, Reynolds R. NG2-expressing cells in the central nervous system: are they oligodendroglial progenitors. J Neurosci Res? 2000;61(5):471–479. doi: 10.1002/1097-4547(20000901)61:5<471::AID-JNR1>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 8.Eze OBL, Nwodo OFC, Ogugua VN. Therapeutic effect of honey bee venom. Res J Pharm Biol Chem Sci. 2016;4(1):48–53. [Google Scholar]
- 9.Khoei NS, Atashpaz S, Ghabili K, Khoei NS, Omidi Y. Melittin and hyaluronidase compound derived from Bee venom for the treatment of multiple sclerosis. J Med Hypotheses Ideas. 2009;3(3):24–28. [Google Scholar]
- 10.Moreno M, Giralt E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: melittin, apamin and mastoparan. Toxins (Basel) 2015;7(4):1126–1150. doi: 10.3390/toxins7041126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Voos P, Yazar M, Lautenschläger R, Rauh O, Moroni A, Thiel G. The small neurotoxin apamin blocks not only small conductance Ca2+ activated K+ channels (SK type) but also the voltage dependent Kv1.3 channel. Eur Biophys J. 2017;46(6):517–523. doi: 10.1007/s00249-016-1196-0. [DOI] [PubMed] [Google Scholar]
- 12.Shimpi R, Chaudhari P, Deshmukh R, Devare S, Bagad Y, Bhurat M. A review: pharmacotherapeutics of bee venom. Word J Pharm Pharm Sci. 2016;5(7):656–567. [Google Scholar]
- 13.Gage GJ, Kipke DR, Shain W. Whole animal perfusion fixation for rodents. J Vis Exp. 2012;(65) doi: 10.3791/3564. pii: 3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.McCormick JB. Apparatus and method for preparing tissue samples for histological examination. United State Patents US7771992B2. 2010. http://www.freepatentsonline.com/7771992.html .
- 15.Laule C, Leung E, Li DK, Traboulsee AL, Paty DW, MacKay AL, et al. Myelin water imaging in multiple sclerosis: quantitative correlations with histopathology. Mult Scler. 2006;12(6):747–753. doi: 10.1177/1352458506070928. [DOI] [PubMed] [Google Scholar]
- 16.Ruifrok AC, Johnston DA. Quantification of histochemical staining by color deconvolution. Anal Quant Cytol Histol. 2001;23(4):291–299. [PubMed] [Google Scholar]
- 17.Zhen W, Liu A, Lu J, Zhang W, Tattersall D, Wang J. An alternative cuprizone-induced demyelination and remyelination mouse model. ASN Neuro. 2017;9(4):1–9. doi: 10.1177/1759091417725174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Morell P, Barrett CV, Mason JL, Toews AD, Hostettler JD, Knapp GW, et al. Gene expression in brain during cuprizone-induced demyelination and remyelination. Mol Cell Neurosci. 1998;12(4-5):220–227. doi: 10.1006/mcne.1998.0715. [DOI] [PubMed] [Google Scholar]
- 19.Torkildsen O, Brunborg LA, Myhr KM, Bø L. The cuprizone model for demyelination. Acta Neurol Scand Suppl. 2008;188:72–76. doi: 10.1111/j.1600-0404.2008.01036.x. [DOI] [PubMed] [Google Scholar]
- 20.Hochstrasser T, Exner GL, Nyamoya S, Schmitz C, Kipp M. Cuprizone-containing pellets are less potent to induce consistent demyelination in the corpus callosum of C57BL/6 mice. J Mol Neurosci. 2017;61(4):617–624. doi: 10.1007/s12031-017-0903-3. [DOI] [PubMed] [Google Scholar]
- 21.Procaccini C, De Rosa V, Pucino V, Formisano L, Matarese G. Animal models of multiple sclerosis. Eur J Pharmacol. 2015;759:182–191. doi: 10.1016/j.ejphar.2015.03.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gudi V, Gingele S, Skripuletz T, Stangel M. Glial response during cuprizone-induced de-and remyelination in the CNS: lessons learned. Front Cell Neurosci. 2014;8:73–96. doi: 10.3389/fncel.2014.00073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Alvarez-Fischer D, Noelker C, Vulinovic F, Grünewald A, Chevarin C, Klein C, et al. Bee venom and its component apamin as neuroprotective agents in a Parkinson disease mouse model. PLoS One. 2013;8(4):e61700. doi: 10.1371/journal.pone.0061700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dempsey CE, Sessions RB, Lamble NV, Campbell SJ. The asparagine-stabilized ß-turn of apamin: contribution to structural stability from dynamics simulation and amide hydrogen exchange analysis. Biochemistry. 2000;39(51):15944–15952. doi: 10.1021/bi002044q. [DOI] [PubMed] [Google Scholar]
- 25.Silva J, Monge-Fuentes V, Gomes F, Lopes K, dos Anjos L, Campos G, et al. Pharmacological alternatives for the treatment of neurodegenerative disorders: wasp and bee venoms and their components as new neuroactive tools. Toxins (Basel) 2015;7(8):3179–3209. doi: 10.3390/toxins7083179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kim SJ, Park JH, Kim K-H, Lee W-R, Pak SC, Han S-M, et al. The protective effect of apamin on LPS/fat-induced atherosclerotic mice. Evid Based Complement Alternat Med. 2012;2012 doi: 10.1155/2012/305454. Article ID 305454, 10 pages. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ferreira R, Schlichter LC. Selective activation of KCa3.1 and CRAC channels by P2Y2 receptors promotes Ca(2+) signaling, store refilling and migration of rat microglial cells. PLoS One. 2013;8(4):e62345. doi: 10.1371/journal.pone.0062345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fordyce CB, Jagasia R, Zhu X, Schlichter LC. Microglia Kv1.3 channels contribute to their ability to kill neurons. J Neurosci. 2005;25(31):7139–7149. doi: 10.1523/JNEUROSCI.1251-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Benton DC, Garbarg M, Moss GW. The relationship between functional inhibition and binding for K(Ca)2 channel blockers. PloS One. 2013;8(9):e73328. doi: 10.1371/journal.pone.0073328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu H, Liu J, Xu E, Tu G, Guo M, Liang S, et al. Human immunodeficiency virus protein Tat induces oligodendrocyte injury by enhancing outward K+ current conducted by KV1.3. Neurobiol Dis. 2017;97(Pt A):1–10. doi: 10.1016/j.nbd.2016.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]