

In this issue, Bru et al. connect Pseudomonas aeruginosa PQS signaling secretion during stress response to swarming behavior (J.-L. Bru, B. Rawson, C. Trinh, K. Whiteson, et al., J Bacteriol 201:e00383-19, 2019, https://doi.org/10.1128/JB.00383-19).
Keywords: quorum sensing, Pseudomonas aeruginosa, stress response, swarming, bacteriophages
ABSTRACT
In this issue, Bru et al. connect Pseudomonas aeruginosa PQS signaling secretion during stress response to swarming behavior (J.-L. Bru, B. Rawson, C. Trinh, K. Whiteson, et al., J Bacteriol 201:e00383-19, 2019, https://doi.org/10.1128/JB.00383-19). Phage-infected or antibiotic-treated bacterial cells secrete PQS to repel healthy, unexposed cells away from the source of the stress. Thus, the collective stress response mechanism driven by PQS signaling influences spatial organization and population dynamics in P. aeruginosa that may provide competitive advantages in certain niches.
TEXT
Pseudomonas aeruginosa is a complicated bacterium with intricate regulatory systems, a wealth of interesting physiological, pathogenic, and behavioral attributes, and clinically important interactions with other bacteria; of course, this is why it is so fascinating to study. Cell-cell communication studies of P. aeruginosa have revealed four quorum-sensing systems: Las and Rhl (both of which are LuxI/R-type N-acylhomoserine lactone systems), PQS (Pseudomonas quinolone signal), and IQS (integrated quorum sensing), as well as an orphan LuxR-type regulator, QscR (1–5). The LasR and RhlR regulators as well as IQS influence synthesis of PQS and the function of the PQS receptor PqsR. In addition, LasR regulates RhlR and QscR regulates LasR (2, 3). This cross talk network of signaling systems controls virulence gene expression, swarming motility, and biofilm formation. PQS is particularly interesting because its synthesis is controlled by the other three quorum-sensing systems as well as in response to nutrient starvation (the stringent response) and antibiotic stress (6, 7). Recent work has shown that PQS binds to many more proteins in P. aeruginosa in addition to its cognate receptor, PqsR, thereby directly influencing virulence via pathways such as phenazine and hydrogen cyanide production (8). This broadly regulative molecule influences iron acquisition, cytotoxicity via oxidative stress, outer membrane vesicle production, and immune responses in the host. Thus, it is perhaps not surprising that PQS also is involved in control of swarming motility.
Swarming is a group behavior in which cells utilize flagella and surfactants to move as multicellular groups across surfaces. P. aeruginosa swarms develop colonies with tendril-like morphology that form from the site of inoculation outward (illustrated in Fig. 1). In P. aeruginosa, as in many other bacteria, motility and biofilm formation (specifically attachment) are inversely related (9), and both are controlled by intracellular levels of cyclic di-GMP and quorum sensing, among other cues, such as the stringent response (10). Swarming enables P. aeruginosa cells to move through mucosal layers in the lung and is postulated to be one of many factors that contributes to resistance to antibiotic treatment (11). Thus, the intersection of cues that connect swarming and antibiotic resistance is of great interest to P. aeruginosa biologists and in the clinic.
FIG 1.
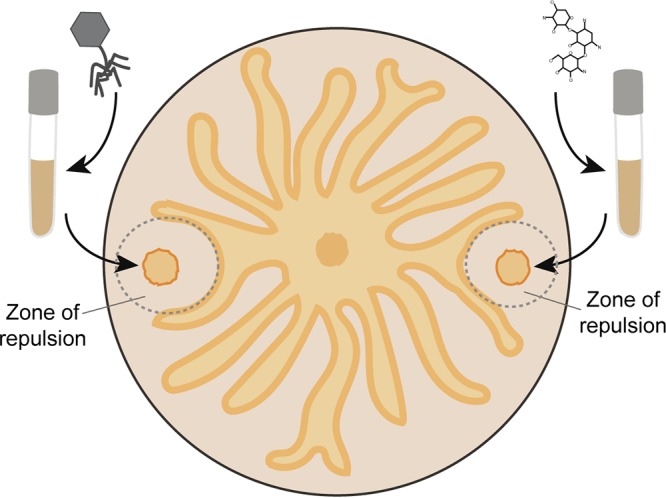
Illustration of Pseudomonas aeruginosa swarming on an agar plate, mimicking the experiments performed by Bru et al. (12). Satellite colonies were generated by spotting cultures treated with either bacteriophage or antibiotic. Wild-type P. aeruginosa is spotted in the center.
In the paper by Bru et al. (12), the authors use swarm assays to monitor spatial organization and population dynamics in the face of two stresses: infection by bacteriophages and exposure to antibiotics (Fig. 1). It had been previously shown that the bacteriophage DMS3 inhibits swarming when integrated as a lysogen (13), and PQS is known to be produced in response to stresses. A population of P. aeruginosa cells infected with a lytic version of DMS3 is able to repel swarms of uninfected cells. This is not due to the presence of phage alone, because cell-free lysate containing active phage does not repel swarms. Instead, cells respond to phage attack by producing PQS. As an example of one of the benefits of community access to preprints, the researchers were able to learn from work performed in the Lavigne laboratory that expression of the PQS synthesis genes is increased during phage infection (14). This result gave Bru and colleagues a clue that the mechanism behind the repulsion of swarms is PQS production. Indeed, adding increasing concentrations of PQS to agar plates results in increased radii of swarming repulsion. Importantly, the repulsion observed by the phage-treated cells was not dependent on the surfactants produced in P. aeruginosa [rhamnolipids and 3-(3-hydroxyalkanoyloxy) alkanoic acid]. Thus, the authors demonstrated that PQS inhibits swarming, and PQS production is a consequence of phage infection. An interesting side observation is that bacteria surviving phage infection are not all phage resistant; approximately half of the P. aeruginosa population that grows after phage infection remains sensitive to phage. It is intriguing to speculate on the mechanism behind the uninheritable phage resistance, but minimally it indicates that the resistance incurred is not genetic in nature (i.e., not a spontaneous suppressor mutant).
To test whether other stresses would influence swarming, the authors treated P. aeruginosa cells with antibiotic (gentamicin). When gentamicin is spotted directly onto the agar medium, swarming is not inhibited. However, when a bacterial culture treated with gentamicin is spotted on the agar, the result is repulsion of untreated P. aeruginosa swarms. The authors showed that this repulsion is due to secretion of PQS into the medium from the gentamicin-treated cells. This result really drives home the point that multiple stressors result in PQS secretion to repel other cells away from a stressed population. The “bow” connecting the “ribbons” of their manuscript is the demonstration that a hypervirulent clinical isolate is capable of repelling swarms of P. aeruginosa cells from the laboratory strain PA14. These findings provide important insights into the role of PQS as a multifunctional signal that can act across long distances on surfaces. The ability of P. aeruginosa cells to respond to stressors and impact a group behavior like swarming is unique, such that the authors coin the term the “collective stress response.”
The core novelty of their manuscript lies in the discovery that stressed P. aeruginosa cells restrict swarming of healthy cells via PQS signaling. This confinement of phage-infected or antibiotic-treated cells and simultaneous repulsion of unaffected cells organize the bacteria into two populations and are a demonstrative example of interpopulation signaling. The next question is the following: can interpopulation signaling confer a benefit to P. aeruginosa cells? Likely the answer is yes, given the number of environmental assaults encountered during infection of host tissue and/or under mixed-species conditions and the presence of stresses such as nutrient deprivation, production of antibiotics from neighboring microbes, and host-produced molecules. One could imagine exploiting this relatively simple swarm assay to model how the collective stress response affects bacterial population structures in mixed-species cultures and, therefore, competition under these conditions.
ACKNOWLEDGMENT
Research on quorum sensing in the van Kessel laboratory is supported by National Institutes of Health grant R35GM124698.
The views expressed in this article do not necessarily reflect the views of the journal or of ASM.
REFERENCES
- 1.Lee J, Wu J, Deng Y, Wang J, Wang C, Wang J, Chang C, Dong Y, Williams P, Zhang LH. 2013. A cell-cell communication signal integrates quorum sensing and stress response. Nat Chem Biol 9:339–343. doi: 10.1038/nchembio.1225. [DOI] [PubMed] [Google Scholar]
- 2.Turkina MV, Vikstrom E. 2019. Bacteria-host crosstalk: sensing of the quorum in the context of Pseudomonas aeruginosa infections. J Innate Immun 11:263–279. doi: 10.1159/000494069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Papenfort K, Bassler BL. 2016. Quorum sensing signal-response systems in Gram-negative bacteria. Nat Rev Microbiol 14:576–588. doi: 10.1038/nrmicro.2016.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ding F, Oinuma KI, Smalley NE, Schaefer AL, Hamwy O, Greenberg EP, Dandekar AA. 2018. The Pseudomonas aeruginosa orphan quorum sensing signal receptor QscR regulates global quorum sensing gene expression by activating a single linked operon. mBio 9:e01274-18. doi: 10.1128/mBio.01274-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fuqua C. 2006. The QscR quorum-sensing regulon of Pseudomonas aeruginosa: an orphan claims its identity. J Bacteriol 188:3169–3171. doi: 10.1128/JB.188.9.3169-3171.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schafhauser J, Lepine F, McKay G, Ahlgren HG, Khakimova M, Nguyen D. 2014. The stringent response modulates 4-hydroxy-2-alkylquinoline biosynthesis and quorum-sensing hierarchy in Pseudomonas aeruginosa. J Bacteriol 196:1641–1650. doi: 10.1128/JB.01086-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Morales-Soto N, Dunham SJB, Baig NF, Ellis JF, Madukoma CS, Bohn PW, Sweedler JV, Shrout JD. 2018. Spatially dependent alkyl quinolone signaling responses to antibiotics in Pseudomonas aeruginosa swarms. J Biol Chem 293:9544–9552. doi: 10.1074/jbc.RA118.002605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dandela R, Mantin D, Cravatt BF, Rayo J, Meijler MM. 2018. Proteome-wide mapping of PQS-interacting proteins in Pseudomonas aeruginosa. Chem Sci 9:2290–2294. doi: 10.1039/c7sc04287f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ha D-G, O'Toole GA. 2015. c-di-GMP and its effects on biofilm formation and dispersion: a Pseudomonas aeruginosa review. Microbiol Spectr 3:MB-0003-2014. doi: 10.1128/microbiolspec.MB-0003-2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Daniels R, Vanderleyden J, Michiels J. 2004. Quorum sensing and swarming migration in bacteria. FEMS Microbiol Rev 28:261–289. doi: 10.1016/j.femsre.2003.09.004. [DOI] [PubMed] [Google Scholar]
- 11.Breidenstein EBM, de la Fuente-Núñez C, Hancock REW. 2011. Pseudomonas aeruginosa: all roads lead to resistance. Trends Microbiol 19:419–426. doi: 10.1016/j.tim.2011.04.005. [DOI] [PubMed] [Google Scholar]
- 12.Bru JL, Rawson B, Trinh C, Whiteson K, Hoyland-Kroghsbo NM, Siryaporn A. 26 August 2019. PQS produced by the Pseudomonas aeruginosa stress response repels swarms away from bacteriophage and antibiotics. J Bacteriol 201:e00383-19. doi: 10.1128/JB.00383-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zegans ME, Wagner JC, Cady KC, Murphy DM, Hammond JH, O'Toole GA. 2009. Interaction between bacteriophage DMS3 and host CRISPR region inhibits group behaviors of Pseudomonas aeruginosa. J Bacteriol 191:210–219. doi: 10.1128/JB.00797-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Blasdel B, Ceyssens P-J, Chevallereau A, Debarbieux L, Lavigne R. 2018. Comparative transcriptomics reveals a conserved bacterial adaptive phage response (BAPR) to viral predation. bioRxiv https://www.biorxiv.org/content/10.1101/248849v1.full. doi: 10.1101/248849. [DOI]