

Abstract
Sildenafil citrate, a potent and selective inhibitor of phosphodiesterase type-5, is used clinically to treat erectile dysfunction and pulmonary arterial hypertension. We investigated the effect of sildenafil citrate on brain central fatigue through serotonin (5-hydroxytryptamine, 5-HT) synthesis after exhaustive swimming exercise in rats. The rats in the sildenafil citrate-treated groups received sildenafil citrate orally once a day for 14 consecutive days at respective dosage. On the 14 days after starting experiment, each animal was submitted to swimming test with intensity equivalent to overload. The exhaustion was defined as a state in which coordinated movements did not return to the water surface for breathing within 10 sec. Immunohistochemistry for 5-HT, tryptophan hydroxylase (TPH), and western blot for serotonergic type 1A (5-HT1A) receptor and 5-HT transporter (5-HTT) were performed. Exhaustive swimming exercise increased 5-HT and TPH expressions in the dorsal raphe and sildenafil citrate suppressed 5-HT and TPH expressions in the exhaustive swimming exercise rats. Exhaustive swimming exercise increased 5-HT1A receptor and 5-HTT expressions in the dorsal raphe and sildenafil citrate suppressed 5-HT1A receptor and 5-HTT expressions in the exhaustive swimming exercise rats. The significant suppressing effect appeared in the 20-mg/kg sildenafil citrate. Sildenafil citrate might be proposed as a potential ergogenic aid through anticentral fatigue.
Keywords: Sildenafil citrate, Central fatigue, Exhaustive exercise, Sero-tonin
INTRODUCTION
Central fatigue is associated with regulation of serotonergic neurotransmitter system, such as serotonin (5-hydroxytryptamine, 5-HT), tryptophan hydroxylase (TPH), 5-HT receptor, and 5-HT transporter (5-HTT) (Cordeiro et al., 2017). 5-HT is an important neurotransmitter, which acts as a biochemical messenger and regulator of the brain function (Kim et al., 2017). Increase in the concentration of 5-HT in the brain during prolonged exercise impairs central nervous system functions and thus bring about a deterioration in exercise performance (Newsholme et al., 1992). 5-HT is modulated by many factors involved in the intrinsic regulation of central neurotransmission, which include TPH and serotonergic type 1A (5-HT1A) receptor (Foley et al., 2006). TPH catalyzes the rate-limiting step of serotonin biosynthesis in the dorsal raphe (Rind et al., 2000). 5-HT1A receptor negatively regulates the activity of 5-HT neurons and is expressed in presynaptic autoreceptor on raphe neurons (Albert and Lemonde, 2004).
Sildenafil citrate, a potent and selective inhibitor of phosphodiesterase type-5, is used clinically to treat erectile dysfunction and pulmonary arterial hypertension (Mostafa, 2008). Sildenafil citrate increases local concentration of cyclic guanosine monophosphate, thus causing vasodilatation by relaxation of arterial wall smooth muscle fibers, especially in the corpus cavernosum and lungs (Harrold et al., 2000). Some studies have proposed the application of sildenafil citrate on several diseases, including myocardial infarction, heart failure, and stroke (Guazzi et al., 2011; Kukreja et al., 2005). In addition, sildenafil citrate has been shown to improve exercise capacity in subjects with cardiopulmonary diseases (Kloner, 2004) and in healthy subjects in hypoxia (Hsu et al., 2006).
As sildenafil citrate is an effective pulmonary vasodilator (Schermuly et al., 2004), sildenafil citrate is a potent ergogenic aid during exercise performed under hypoxic conditions. However, anti-fatigue effect of sildenafil citrate is the tendency to focus on only peripheral fatigue in hypoxic conditions. Therefore, we investigated effects of sildenafil citrate administration on brain central fatigue after exhaustive swimming exercise in rats.
MATERIALS AND METHODS
Animals
Adult male Sprague-Dawley rats (8 weeks old) were used in this experiment. The experimental procedure was performed in accordance with the animal care guidelines of the National Institutes of Health and the Korean Academy of Medical Sciences. The rats were randomly divided into four groups (n=10 in each group): the control group, the exhaustive exercise group, the exhaustive exercise and 20-mg/kg sildenafil citrate-treated group, and the exhaustive exercise and 40-mg/kg sildenafil citrate-treated group. The rats in the sildenafil citrate-treated groups received sildenafil citrate (Sigma Chemical Co., St. Louis, MO, USA) orally once a day for 14 consecutive days at respective dosage. The rats in the control and exercise groups received normal saline orally once a day for 14 consecutive days.
Exhaustive swimming exercise
On the 14 days after starting experiment, each animal was submitted to exhaustive swimming test with intensity equivalent to overload (a metal ring weight attached to the animal’s torso) of 10% of the body weight (Ding et al., 2009). The exhaustion was defined as a state in which coordinated movements did not return to the water surface for breathing within 10 sec.
Tissue preparation
After the exhaustive swimming exercise test, the rats were deeply anesthetized with Zoletil 50 anesthesia (40 mg/kg, intraperitoneally; Vibac Laboratories, Carros, France). After complete anesthesia, the rats were transcardially perfused with 0.05 M phosphate-buffered saline (PBS), followed by 4% paraformaldehyde in 0.5 M sodium phosphate buffer at pH 7.4. The brain was removed, postfixed in the same fixative overnight, and transferred to a 30% sucrose solution for cryoprotection. Serial 40-μm-thick coronal sections were cut with a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry for 5-HT and TPH
Immunohistochemistry for 5-HT and TPH was performed, according to the previously described method (Shin et al., 2017). An average of eight sections was selected in each brain region spanning from Bregma −7.30 to −8.00 mm. The sections were incubated in PBS for 10 min and they were next washed three times with PBS. The sections were then incubated in 1% H2O2 for 30 min, and then they were incubated overnight with rabbit anti-5-HT antibody (Oncogene Research Product, Cambridge, UK) at a dilution of 1:500 or with mouse anti-TPH antibody (Oncogene Research Product) at a dilution of 1:500. The sections were incubated for 1 hr with biotinylated anti-rabbit secondary antibody or with anti-mouse secondary antibody (Vector Laboratories, Burlingame, CA, USA), and they were subsequently incubated with avidin–biotin–peroxidase complex (Vector Laboratories) for 1 hr at room temperature. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% 3,3′-diaminobenzidine and 0.01% H2O2 in 50 mM Tris-buffer (pH, 7.6) for approximately 3 min. The sections were finally mounted on gelatin-coated glass slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount (Thermo Fisher Scientific Inc., Waltham, MA, USA).
Western blot analysis
Western analysis was performed, according to the previously described method (Shin et al., 2017). Dorsal raphe tissues were dissected. Sample tissues were stored at −70°C until analysis. The tissues were lysed in ice-cold lysate buffer containing 50 mM HEPES (pH, 7.5), 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1.5 mM magnesium chloride hexahydrate, 1 mM ethyleneglycol-bis-(β-aminoethyl ether)-N,N′-tetraacetic acid, 1 mM phenylmethylsulfonyl fluoride, 2-μg/mL leupeptin, 1-μg/mL pepstatin, 1 mM sodium orthovanadate, and 100 mM sodium fluoride, after which the mixture was incubated for 30 min at 4°C. The protein concentration was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein of 30 μg was separated on sodium dodecyl sulfate-polyacrylamide gels and transferred onto a nitrocellulose membrane (Whatman, Clifton, NJ, USA). Mouse anti-β actin (1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-5-HT1A receptor antibody (1:2,000; Abcam, Cambridge, UK) and rabbit anti-5-HTT (1:2,000; Abcam) were used as a primary antibody. A horseradish peroxidase-conjugated anti-mouse secondary antibody was used for β-actin, and an anti-rabbit secondary antibody was used for 5-HT1A receptor and 5-HTT. Band detection was performed using the enhanced chemiluminescence detection system (Santa Cruz Biotechnology). The bands were quantified using an Image-Pro Plus computer-assisted image analysis system (Media Cybernetics Inc., Bethesda, MD, USA).
Statistical analysis
Differences among the groups were evaluated using IBM SPSS Statistics ver. 23.0 (IBM Co., Armonk, NY, USA) by the one-way analysis of variance followed by Duncan post hoc test. All values are expressed as the mean±standard error of the mean. Statistically significant differences were established at P<0.05.
RESULTS
Effect of sildenafil citrate on exhaustive swimming time
Exhaustive swimming time is presented in Table 1. Exhaustive swimming time was increased in the exhaustive exercise and sildenafil citrate-treated groups than exhaustive swimming exercise group. The significant increasing effect on exhaustive swimming time appeared in the 20-mg/kg sildenafil citrate (P<0.05).
Table 1.
Effect of sildenafil citrate on exhaustive swimming time in rats
| Exhaustive exercise group | Exhaustive exercise and 20 mg sildenafil citrate-treated group | Exhaustive exercise and 40 mg sildenafil citrate-treated group |
|---|---|---|
| 18.85±0.69 min | 26.39±4.75 min* | 20.89±0.90 min |
The results are presented as the mean±standard error of the mean.
P<0.05 compared to the exhaustive exercise group.
Effect of sildenafil citrate on 5-HT expression in the dorsal raphe
Photomicrographs of 5-HT-positive cells in the dorsal raphe are presented in Fig. 1. Exhaustive swimming exercise increased 5-HT expression in the dorsal raphe (P<0.05) and sildenafil citrate suppressed 5-HT expression in the exhaustive swimming exercise rats (P<0.05). The significant suppressing effect appeared in the 20-mg/kg sildenafil citrate.
Fig. 1.
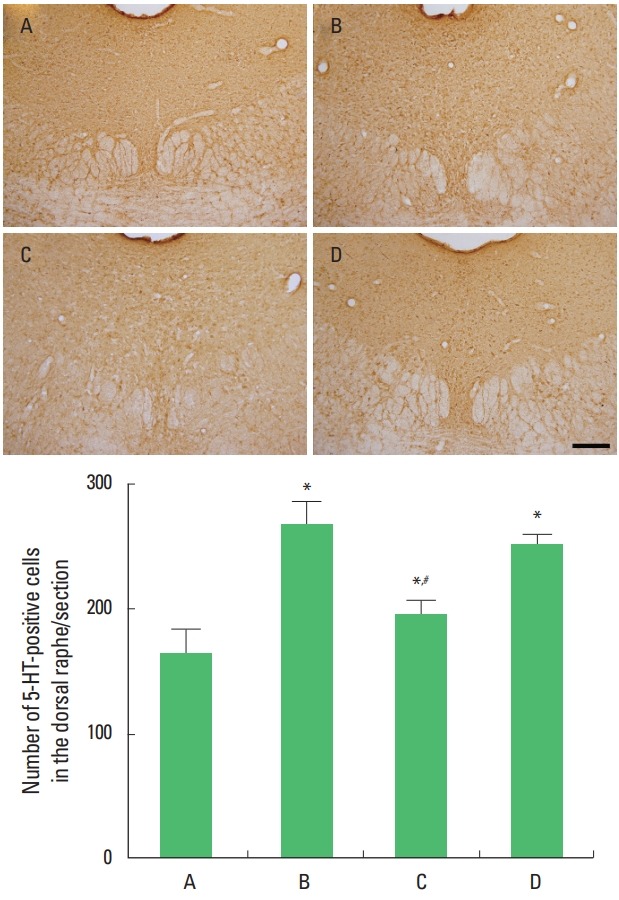
Effects of sildenafil citrate on 5-hydroxytryptamine (5-HT). Upper panel: Photomicrographs showing 5-HT expressions in the dorsal raphe. Lower panel: The results are presented as the mean±standard error of the mean. The scale bar represents 250 μm. A, control group; B, exhaustive exercise group; C, exhaustive exercise and 20-mg/kg sildenafil treatment group; D, exhaustive exercise and 40-mg/kg sildenafil treatment group. *P<0.05 compared to the control group. #P<0.05 compared to the exhaustive exercise group.
Effect of sildenafil citrate on TPH expression in the dorsal raphe
Photomicrographs of TPH-positive cells in the dorsal raphe are presented Fig. 2. Exhaustive swimming exercise increased TPH expression in the dorsal raphe (P<0.05) and sildenafil citrate suppressed TPH expression in the exhaustive swimming exercise rats (P<0.05). The significant suppressing effect appeared in the 20-mg/kg sildenafil citrate.
Fig. 2.
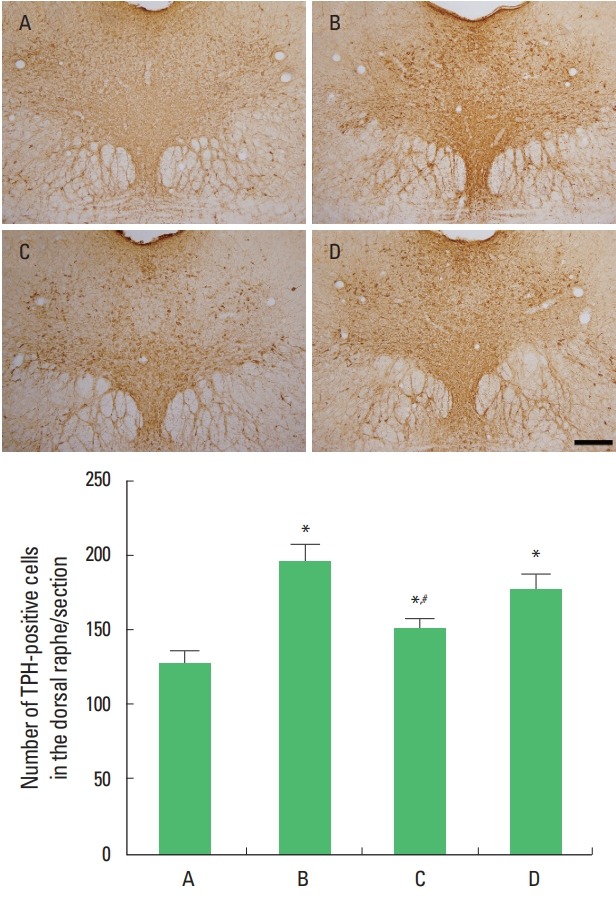
Effects of sildenafil citrate on tryptophan hydroxylase (TPH). Upper panel: Photomicrographs showing TPH expressions in the dorsal raphe. Lower panel: The results are presented as the mean±standard error of the mean. The scale bar represents 250 μm. A, control group; B, exhaustive exercise group; C, exhaustive exercise and 20-mg/kg sildenafil treatment group; D, exhaustive exercise and 40-mg/kg sildenafil treatment group. *P<0.05 compared to the control group. #P<0.05 compared to the exhaustive exercise group.
Effect of treadmill exercise on 5-HT1A receptor and 5-HTT expression in the dorsal raphe
The 5-HT1A receptor and 5-HTT expressions are shown in Fig. 3. Exhaustive swimming exercise increased 5-HT1A receptor and 5-HTT expressions in the dorsal raphe (P<0.05) and sildenafil citrate suppressed 5-HT1A receptor and 5-HTT expressions in the exhaustive swimming exercise rats (P<0.05). The significant suppressing effect appeared in the 20-mg/kg sildenafil citrate.
Fig. 3.
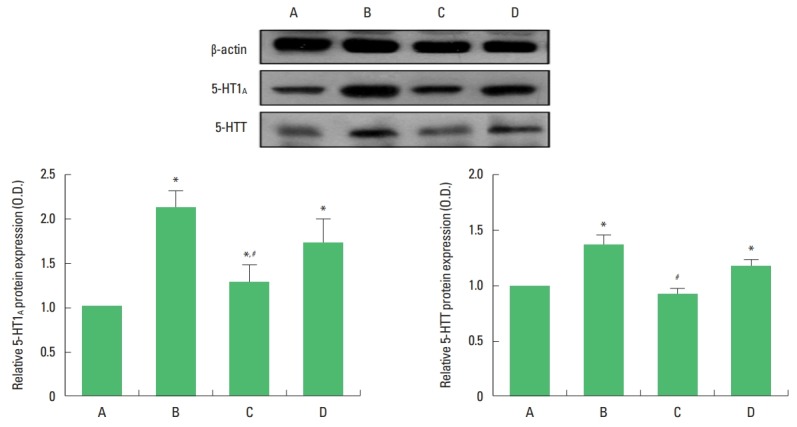
Effects of sildenafil citrate on 5-hydroxytryptamine-1A (5-HT1A) receptor and 5-hydroxytryptamine transporter (5-HTT) expressions in the dorsal raphe. Upper panel: The expressions of 5-HT1A and 5-HTT. Lower panel: The results are presented as the mean±standard error of the mean. A, control group; B, exhaustive exercise group; C, exhaustive exercise and 20-mg/kg sildenafil treatment group; D, exhaustive exercise and 40-mg/kg sildenafil treatment group. *P<0.05 compared to the control group. #P<0.05 compared to the exhaustive exercise group.
DISCUSSION
Sildenafil citrate decreases pulmonary vascular resistance and increases VO2max in hypoxic normal subjects (Ghofrani et al., 2004; Richalet et al., 2005). Galiè et al. (2005) reported that sildenafil citrate improved exercise capacity and pulmonary hemodynamic in patients with pulmonary arterial hypertension. Moreover, chronic sildenafil citrate treatment increased dopamine and 5-HT turnover rate in the medial preoptic area and nucleus accumben in the normal male rats (Kyratsas et al., 2013). In the present study, treatment of 20-mg sildenafil citrate prolonged exhaustive swimming time.
Central fatigue has been proposed the implication of various neurotransmitters such as serotonin, norepinephrine, and dopamine (Newsholme et al., 1992). Among these, serotonin is known to play the most important role in the pathogenesis of central fatigue (Kim et al., 2017). Increment in the 5-HT concentration in the brain is associated with mental fatigue (Davis et al., 2000). Increased concentration of 5-HT leads to the onset of fatigue (Soares et al., 2007), while decreased 5-HT concentration could delay the time to fatigue (Seo et al., 2011). In addition, exhaustive exercise increases 5-HT concentrations and then causes decrement of endurance performance (Caperuto et al., 2009). Inhibition of 5-HT production in the brain could increase endurance exercise performance (Seo et al., 2011). The synthesis of 5-HT is modulated by the hydroxylation of the amino acid tryptophan, which is the rate-limiting step catalyzed by TPH (Carkaci-Salli et al., 2006). Reduction in TPH expression decreases in 5-HT synthesis (Park et al., 2019). In the present study, treadmill exercise increased 5-HT synthesis and TPH expression in the dorsal raphe and 20-mg sildenafil citrate suppressed the exhaustive exercise-induced increase of 5-HT synthesis and TPH expression in the dorsal raphe.
Treadmill exercise upregulates the levels of 5-HT and 5-HT1A receptor in rats with permanent middle cerebral artery occlusion (Lan et al., 2014). 5-HT1A receptor expression in the dorsal raphe was reduced by olfactory bulbectomy and treadmill exercise increased 5-HT1A receptor expression in the olfactory bulbectomized rats (Shin et al., 2017). Exhaustive exercise increased 5-HT1A receptor and 5-HTT and colostrum serum treatment suppressed exhaustive exercise-induced 5-HT1A receptor and 5-HTT expression in the dorsal raphe (Kim et al., 2017). Primary pulmonary hypertension is caused by increased expression of the 5-HTT (Eddahibi et al., 2001). Degree of central fatigue depends on the function of the serotonin system (Kim et al., 2017; Maluchenko et al., 2009). In the present study, treadmill exercise increased 5-HT1A receptor and 5-HTT expression in the dorsal raphe and 20-mg sildenafil citrate suppressed the exhaustive exercise-induced increase of 5-HT1A receptor and 5-HTT expressions in the dorsal raphe.
These results may support sildenafil citrate as an ergogenic aid to improve exercise performance. Sildenafil citrate might be proposed as a potential ergogenic aid through anticentral fatigue.
ACKNOWLEDGMENTS
This study was supported by the Ministry of Education of the Republic of Korea and the National Research Foundation of Korea (NRF-2017S1A5A8022823).
Footnotes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
REFERENCES
- Albert PR, Lemonde S. 5-HT1A receptors, gene repression, and depression: guilt by association. Neuroscientist. 2004;10:575–593. doi: 10.1177/1073858404267382. [DOI] [PubMed] [Google Scholar]
- Caperuto EC, dos Santos RV, Mello MT, Costa Rosa LF. Effect of endurance training on hypothalamic serotonin concentration and performance. Clin Exp Pharmacol Physiol. 2009;36:189–191. doi: 10.1111/j.1440-1681.2008.05111.x. [DOI] [PubMed] [Google Scholar]
- Carkaci-Salli N, Flanagan JM, Martz MK, Salli U, Walther DJ, Bader M, Vrana KE. Functional domains of human tryptophan hydroxylase 2 (hTPH2) J Biol Chem. 2006;281:28105–28112. doi: 10.1074/jbc.M602817200. [DOI] [PubMed] [Google Scholar]
- Cordeiro LMS, Rabelo PCR, Moraes MM, Teixeira-Coelho F, Coimbra CC, Wanner SP, Soares DD. Physical exercise-induced fatigue: the role of serotonergic and dopaminergic systems. Braz J Med Biol Res. 2017;50:e6432. doi: 10.1590/1414-431X20176432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis JM, Alderson NL, Welsh RS. Serotonin and central nervous system fatigue: nutritional considerations. Am J Clin Nutr. 2000;72(2 Suppl):573S–578S. doi: 10.1093/ajcn/72.2.573S. [DOI] [PubMed] [Google Scholar]
- Ding X, Tang K, Lu P, Putheti R. Antifatigue effects of polydatin from Chinese herb Polygonum Cuspidatum in swimming mice. Afr J Microbiol Res. 2009;3:358–361. [Google Scholar]
- Eddahibi S, Humbert M, Fadel E, Raffestin B, Darmon M, Capron F, Simonneau G, Dartevelle P, Hamon M, Adnot S. Serotonin transporter overexpression is responsible for pulmonary artery smooth muscle hyperplasia in primary pulmonary hypertension. J Clin Invest. 2001;108:1141–1150. doi: 10.1172/JCI12805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foley TE, Greenwood BN, Day HE, Koch LG, Britton SL, Fleshner M. Elevated central monoamine receptor mRNA in rats bred for high endurance capacity: implications for central fatigue. Behav Brain Res. 2006;174:132–142. doi: 10.1016/j.bbr.2006.07.018. [DOI] [PubMed] [Google Scholar]
- Galiè N, Ghofrani HA, Torbicki A, Barst RJ, Rubin LJ, Badesch D, Fleming T, Parpia T, Burgess G, Branzi A, Grimminger F, Kurzyna M, Simonneau G, Sildenafil Use in Pulmonary arterial Hypertension (SUPER) Study Group Sildenafil citrate therapy for pulmonary arterial hypertension. N Engl J Med. 2005;353:2148–2157. doi: 10.1056/NEJMoa050010. [DOI] [PubMed] [Google Scholar]
- Ghofrani HA, Reichenberger F, Kohstall MG, Mrosek EH, Seeger T, Olschewski H, Seeger W, Grimminger F. Sildenafil increased exercise capacity during hypoxia at low altitudes and at Mount Everest basecamp: a randomized, double-blind, placebo-controlled crossover trial. Ann Intern Med. 2004;141:169–177. doi: 10.7326/0003-4819-141-3-200408030-00005. [DOI] [PubMed] [Google Scholar]
- Guazzi M, Vicenzi M, Arena R, Guazzi MD. Pulmonary hypertension in heart failure with preserved ejection fraction: a target of phosphodiesterase-5 inhibition in a 1-year study. Circulation. 2011;124:164–174. doi: 10.1161/CIRCULATIONAHA.110.983866. [DOI] [PubMed] [Google Scholar]
- Harrold LR, Gurwitz JH, Field TS, Andrade SE, Fish LS, Jarry PD, Yood RA. The diffusion of a novel therapy into clinical practice: the case of sildenafil. Arch Intern Med. 2000;160:3401–3405. doi: 10.1001/archinte.160.22.3401. [DOI] [PubMed] [Google Scholar]
- Hsu AR, Barnholt KE, Grundmann NK, Lin JH, McCallum SW, Friedlander AL. Sildenafil improves cardiac output and exercise performance during acute hypoxia, but not normoxia. J Appl Physiol. 2006;100:2031–2040. doi: 10.1152/japplphysiol.00806.2005. [DOI] [PubMed] [Google Scholar]
- Kim TW, Kim CJ, Seo J. Effects of colostrum serum on the serotonergic system in the dorsal raphe nuclei of exercised rats. J Exerc Nutrition Biochem. 2017;21:33–39. doi: 10.20463/jenb.2017.0047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kloner RA. Cardiovascular effects of the 3 phosphodiesterase-5 inhibitors approved for the treatment of erectile dysfunction. Circulation. 2004;110:3149–3155. doi: 10.1161/01.CIR.0000146906.42375.D3. [DOI] [PubMed] [Google Scholar]
- Kukreja RC, Salloum F, Das A, Ockaili R, Yin C, Bremer YA, Fisher PW, Wittkamp M, Hawkins J, Chou E. Pharmacological preconditioning with sildenafil: basic mechanisms and clinical implications. Vascul Pharmacol. 2005;42:219–232. doi: 10.1016/j.vph.2005.02.010. [DOI] [PubMed] [Google Scholar]
- Kyratsas C, Dalla C, Anderzhanova E, Polissidis A, Kokras N, Konstantinides K, Papadopoulou-Daifoti Z. Experimental evidence for sildenafil’s action in the central nervous system: dopamine and serotonin changes in the medial preoptic area and nucleus accumbens during sexual arousal. J Sex Med. 2013;10:719–729. doi: 10.1111/j.1743-6109.2012.03000.x. [DOI] [PubMed] [Google Scholar]
- Lan X, Zhang M, Yang W, Zheng Z, Wu Y, Zeng Q, Liu S, Liu K, Li G. Effect of treadmill exercise on 5-HT, 5-HT1A receptor and brain derived neurophic factor in rats after permanent middle cerebral artery occlusion. Neurol Sci. 2014;35:761–766. doi: 10.1007/s10072-013-1599-y. [DOI] [PubMed] [Google Scholar]
- Maluchenko NV, Schegolkova JV, Kulikova MA, Timofeeva MA, Shlepcova VA, Sysoeva OV, Tonevitsky AG. Gender effects on association of serotonin transporter gene polymorphism with symptoms of central fatigue. Bull Exp Biol Med. 2009;147:462–465. doi: 10.1007/s10517-009-0559-2. [DOI] [PubMed] [Google Scholar]
- Mostafa T. Oral phosphodiesterase type 5 inhibitors: nonerectogenic beneficial uses. J Sex Med. 2008;5:2502–2518. doi: 10.1111/j.1743-6109.2008.00983.x. [DOI] [PubMed] [Google Scholar]
- Newsholme EA, Blomstrand E, Ekblom B. Physical and mental fatigue: metabolic mechanisms and importance of plasma amino acids. Br Med Bull. 1992;48:477–495. doi: 10.1093/oxfordjournals.bmb.a072558. [DOI] [PubMed] [Google Scholar]
- Park SS, Park HS, Jeong H, Kwak HB, No MH, Heo JW, Yoo SZ, Kim TW. Treadmill exercise ameliorates chemotherapy-induced muscle weakness and central fatigue by enhancing mitochondrial function and inhibiting apoptosis. Int Neurourol J. 2019;23(Suppl 1):S32–9. doi: 10.5213/inj.1938046.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richalet JP, Gratadour P, Robach P, Pham I, Déchaux M, Joncquiert-Latarjet A, Mollard P, Brugniaux J, Cornolo J. Sildenafil inhibits altitude-induced hypoxemia and pulmonary hypertension. Am J Respir Crit Care Med. 2005;171:275–281. doi: 10.1164/rccm.200406-804OC. [DOI] [PubMed] [Google Scholar]
- Rind HB, Russo AF, Whittemore SR. Developmental regulation of tryptophan hydroxylase messenger RNA expression and enzyme activity in the raphe and its target fields. Neuroscience. 2000;101:665–677. doi: 10.1016/s0306-4522(00)00402-4. [DOI] [PubMed] [Google Scholar]
- Schermuly RT, Kreisselmeier KP, Ghofrani HA, Yilmaz H, Butrous G, Ermert L, Ermert M, Weissmann N, Rose F, Guenther A, Walmrath D, Seeger W, Grimminger F. Chronic sildenafil treatment inhibits monocrotaline-induced pulmonary hypertension in rats. Am J Respir Crit Care Med. 2004;169:39–45. doi: 10.1164/rccm.200302-282OC. [DOI] [PubMed] [Google Scholar]
- Seo JH, Sung YH, Kim KJ, Shin MS, Lee EK, Kim CJ. Effects of Phellinus linteus administration on serotonin synthesis in the brain and expression of monocarboxylate transporters in the muscle during exhaustive exercise in rats. J Nutr Sci Vitaminol (Tokyo) 2011;57:95–103. doi: 10.3177/jnsv.57.95. [DOI] [PubMed] [Google Scholar]
- Shin MS, Park SS, Lee JM, Kim TW, Kim YP. Treadmill exercise improves depression-like symptoms by enhancing serotonergic function through upregulation of 5-HT1A expression in the olfactory bulbectomized rats. J Exerc Rehabil. 2017;13:36–42. doi: 10.12965/jer.1734918.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soares DD, Coimbra CC, Marubayashi U. Tryptophan-induced central fatigue in exercising rats is related to serotonin content in preoptic area. Neurosci Lett. 2007;415:274–278. doi: 10.1016/j.neulet.2007.01.035. [DOI] [PubMed] [Google Scholar]