

Abstract
Vascular pericytes provide critical contributions to the formation and integrity of the blood vessel wall within the microcirculation. Pericytes maintain vascular stability and homeostasis by promoting endothelial cell junctions and depositing extracellular matrix (ECM) components within the vascular basement membrane (vBM), among other vital functions. As their importance in sustaining microvessel health within various tissues and organs continues to emerge, so does their role in a number of pathological conditions including cancer, diabetic retinopathy, and neurological disorders. Here, we review vascular pericyte contributions to the development and remodeling of the microcirculation, with a focus on the local microenvironment during these processes. We discuss observations of their earliest involvement in vascular development and essential cues for their recruitment to the remodeling endothelium. Pericyte involvement in the angiogenic sprouting context is also considered with specific attention to crosstalk with endothelial cells such as through signaling regulation and ECM deposition. We also address specific aspects of the collective cell migration and dynamic interactions between pericytes and endothelial cells during angiogenic sprouting. Lastly, we discuss pericyte contributions to mechanisms underlying the transition from active vessel remodeling to the maturation and quiescence phase of vascular development.
Keywords: pericytes, endothelial cells, vascular morphogenesis
Introduction
Vascular pericytes are essential components of the microcirculation. These specialized cells wrap around and ensheath microvessels, promoting endothelial cell junction stability and depositing extracellular matrix (ECM), among many key functions. Eberth and Rouget first described these cells in the 1870s as perivascular cells that differed in morphology from vascular smooth muscle cells (vSMCs); pericytes appeared to be elongated along microvessels in perivascular locations, while vSMCs were concentrically wrapped around the endothelium [1–3]. Despite being nearly ubiquitous in the microcirculation, research into vascular pericytes lagged behind their endothelial counterparts following their initial identification. More recently, however, pericytes have attracted significant attention across an array of biological disciplines. Potential new functions are being reported for these cells in vascular development and tissue homeostasis, as well as in a broad range of disease states [4, 5]. Contributions to microvessel stability, and to overall vascular barrier function are well-accepted roles for pericytes. Intriguing nuances within these more “canonical” roles are still being discovered in health [6–8] and in diseases such as proliferative diabetic retinopathy [9–12], cancer and metastatic progression [13, 14], and Alzheimer’s disease [15]. Pericyte contractility, or the modulation of microvessel diameter, is an area of on-going investigation, particularly in the central nervous system (CNS) [16–23]. In addition, the tissue regeneration capacity of pericytes, acting as a pool of perivascular mesenchymal stem cells (MSCs) [24–26], has also been described as a potential role for these cells [27–29]. This particular role may be context-and/or model-dependent however [27, 30–39]. These established and emerging functions for pericytes, among others, are still being elucidated and have been the subject of many insightful reviews [4, 40–46]. We acknowledge the wide breadth of intriguing studies that have addressed pericyte biology thus far. Here, we narrow the focus of this review on pericytes in their engagement with the vascular endothelium during blood vessel development and remodeling. We give a specific consideration to the elements within the pericyte microenvironment that are critical for their contributions to these processes.
Pericyte Origins and Identification
During development, pericytes arise from a wide-range of embryonic and extra-embryonic regions. Neural crest and primordial mesenchyme are the most commonly described origins, though specific tissues and organs likely derive vascular pericytes from unique cellular niches [23, 43, 47, 48]. Because of their physical proximity and proposed functional overlap, pericyte lineage has frequently been inferred from tracing the differentiation of vascular smooth muscle cells (vSMCs) [4]. While pericytes and vSMCs may share a common derivation to a certain point, these cells ultimately occupy unique regions of the vasculature and are morphologically and functionally distinct [49]. Additional validation studies will therefore be necessary, especially as pericyte-specific tools and markers continue to evolve alongside the advancement of single-cell analysis and next-generation sequencing techniques [50–55]. Volz et al. for example recently observed vascular pericytes present in the mouse heart as early as embryonic day 11.5 (E11.5), and these cells provided a source for coronary artery vSMCs [56]. Interestingly, this elegant study found that Notch pathway signals, known to be important for vSMC differentiation and development [57–61], were not required for the differentiation, recruitment, or retention of microvascular pericytes. These observations are consistent with CNS-focused [62, 63] and developmental studies [64]. However, these results may be context-dependent [65] and could be confounded by the fact that, although platelet-derived growth factor receptor-β (PDGFRβ) and neural-glial antigen-2 (NG2, gene name: Cspg4) are useful markers for pericytes, they are also expressed by other cell types such as vSMCs and brain glia [61, 63, 66], and oligodendrocyte precursor cells (OPCs) [67–69], respectively.
To clarify the interpretation of these markers, vascular endothelial cells are often co-labeled within a given specimen, providing proximity to the endothelium as another criteria for positive identification of vascular pericytes [4]. French et al. utilized this approach in the developing mouse yolk sac, and found PDGFRβ-positive pericytes adjacent to distinct Tie2-expressing endothelial cells as early as E8.5 [70]. Recently, Jung et al. coupled a powerful double-labeled pericyte transgenic mouse line (Pdgfrβ-EGFP and Cspg4-DsRed) with endothelial immunostaining. The authors visualized vascular-associated pericytes in the brain at E10.5, with increasing abundance throughout the remainder of embryonic and early postnatal development [71].
Previous studies have suggested that the endothelium acquires pericyte associations primarily, if not exclusively, after endothelial sprouting events begin to establish basic vascular networks, perhaps allowing greater plasticity in endothelial remodeling [66, 72–74]. We, and others [75–78], have found that mouse embryonic stem cells (ESCs) give rise not only to primitive endothelial cell networks, but also to presumptive pericytes (or pericyte precursors) during the earliest stages of cardiovascular development [64]. Pericytes seem to emerge at approximately the same time as, or even prior to, endothelial cell differentiation, homing to endothelium engaged in both vasculogenic (Payne et al., Unpublished observations) and angiogenic processes [64, 75–78]. Nevertheless, whether pericytes, or their precursors, directly engage with the developing endothelium to actively coordinate the formation of primitive blood vessels remains an ongoing area of investigation. Insight from these studies could shed light on the involvement of pericytes in pathological conditions associated with vessel dysmorphogenesis and mis-patterning such as arterio-venous malformations (AVMs) and cerebral cavernous malformations (CCMs) [58, 79, 80].
Pericyte Recruitment and Contribution to Early Vessel Formation
As endothelial cells form primitive tubes and more complex vascular networks, they simultaneously release molecular cues that facilitate pericyte recruitment to the developing vessel wall [66, 81, 82]. Platelet-derived growth factor-BB (PDGF-BB) is one of the most potent regulators of pericyte recruitment, investment, and retention. Genetic loss or mutation of this ligand [e.g. altering its extracellular matrix (ECM) retention motif] and its primary signaling receptor tyrosine kinase, PDGFRβ, lead to substantial loss of pericytes and subsequent vascular complications [8, 66, 83–87]. In an in vitro model of vasculogenic tube formation, endothelial cells also secrete high levels of Heparin-Binding EGF-like Growth Factor (HB-EGF), which engages Epidermal Growth Factor Receptor (EGFR, or ErbB1) and ErbB4 on pericytes to promote their recruitment [88]. A pericyte recruitment factor that can be released from non-endothelial sources appears to be Stromal Derived Factor-1α (SDF-1α), which binds to CXCR4 on pericytes [89]. SDF1α may in fact work synergistically with PDGF-BB to elicit pericyte migration to growing vessels [90]. These factors, as well as others, can facilitate pericyte recruitment to developing vessels, perhaps during the earliest stages of vessel formation. Comparable to observations from extra-embryonic yolk sac vessels [70], our recent observations suggest that this recruitment may occur during vasculogenic formation of primitive vessels from nascent endothelial cells (Payne et al., Unpublished observations). Pericytes may also physically participate in endothelial cell coalescence [75] and additional processes that are critical for vasculogenesis, though more work remains to further support these findings.
As vessels develop, specialized ECM known as the vascular basement membrane (vBM) is deposited around the cells within the blood vessel wall, conferring stability and providing a scaffold for signaling proteins, among other functions [91, 92]. Pericytes make important contributions to this ECM deposition and vBM formation [93, 94]. In an in vitro co-culture model with endothelial cells mimicking developmental vasculogenic tube formation, pericytes deposited ECM components such as fibronectin, laminin isoforms, perlecan, and nidogen-1 [95]. These observations, along with ultra-structural imaging techniques such as scanning electron microscopy (SEM) [96, 97], have revealed that their synthesis, remodeling of, and location within the vBM is another key characteristic of vascular pericytes [98]. This feature is likely an important biomarker for distinguishing pericytes from other cell types that may be positioned adjacent to blood vessels but may not be physically incorporated within the vessel wall proper i.e. encased within the vBM. Type IV collagen (col-IV) for instance is an important component of the vBM that pericytes and endothelial cells synthesize or degrade depending on the phase of vascular remodeling, pruning, or stabilization [88, 92, 98–100]. In the developing and adult mouse CNS, pericytes have also been found to secrete vitronectin into the vBM [50]. Pericyte-derived vitronectin may in fact exacerbate pathologies such as Cerebral Autosomal Dominant Arteriopathy with Sub-cortical Infarcts and Leukoencephalopathy (CADASIL) by contributing to white matter lesion severity in aged mice [101]. There may also be unique signaling relationships between pericytes and their surrounding ECM, as recent studies suggest that ECM components secreted by adjacent cell types such as astrocytes may activate pericytes and even promote their differentiation [102]. While pericytes ultimately provide important regulation of the vBM, it is unclear to what extent pericytes deposit each of these vBM components during: (i) early blood vessel formation, (ii) the later stages of vascular development and maturation when vessel remodeling nears completion, and (iii) aging and in vascular-related pathologies. Mis-regulated synthesis of ECM components may also implicate pericytes in fibrotic disease processes.
Pericyte Involvement in Angiogenic Sprouting
During angiogenic remodeling, endothelial cells sprout and migrate outward from existing vessels such as those formed during vasculogenesis. Sprouting angiogenesis can also occur from quiescent microvasculature stimulated with endogenous, pro-angiogenic cues like Vascular Endothelial Growth Factor-A (VEGF-A) (Figure 1). In the adult brain (Figure 1) and early postnatal retina [103], for example, pericyte are found directly adjacent to endothelial “tip” cells that are leading the elongation of nascent vessel branches. This scenario has also been described previously in a variety of vascular formation models and assays [76, 96, 97, 104–106]. This close spatial proximity between sprouting endothelial cells and neighboring pericytes suggests a crosstalk, and potentially a reciprocal signaling relationship, between these two cell types during angiogenic remodeling. Hellstrom et al. provided support for this idea from observations of VEGF-A-stimulated endothelial tip cells synthesizing PDGF-BB as they migrated outward from a parent vessel [107]. Because PDGF-BB possesses a critical ECM retention motif [84], this ligand is thought to localize to the endothelial cell surface and/or become anchored within the ECM alongside endothelial cells. This deposition also appears to require appropriate configuration of heparan sulfate proteoglycans (HSPGs) such as perlecan (i.e. Hspg2) [83].
Figure 1.

Confocal image of a sprouting endothelial cell labeled for platelet-endothelial cell adhesion molecule-1 (PECAM-1; A and green in D, indicated by light green arrows) with an adjacent pericyte labeled for platelet-derived growth factor receptor-β (PDGFRβ; B and red in D, indicated by pink arrows) extending from a microvessel in the adult mouse brain. Cell nuclei are labeled with DAPI (C, and blue in D). The pericyte appears to engage the sprouting endothelial tip cell from the base of the filopodial extensions, along the length of the sprout and the parent vessel. Scale bar, 10 microns.
We recently visualized NG2-DsRed-positive pericytes migrating along sprouting endothelial cells in an ex vivo model of mouse embryonic (E14.5) back skin (Figure 2 and Supplemental Movie 1) [64]. Throughout remodeling vascular networks, pericyte migration appeared to be tightly entrained to the endothelial sprout. These observations were consistent with the notion that endothelial-derived PDGF-BB, and perhaps other recruitment and retention factors, guide and focus pericyte migration to the abluminal microvessel surface [84]. Interestingly, we also noted a relatively frequent occurrence in the dynamic positioning of endothelial tip cells and adjacent pericytes. Specifically, as a tip cell retracted, the corresponding pericyte remained extended well beyond the endothelial cell soma (Figure 2 and Supplemental Movie 1). Over time, the pericyte pulled back to match the apparent leading front of the endothelial cell. The tip cell would eventually resume its outward migration, and the pericyte would continue to keep pace with vessel sprouting (Figure 2 and Supplemental Movie 1). Considering these anecdotal observations alongside current published data [64, 74] suggests a model in which endothelial-secreted cues such as PDGF-BB are critical for synchronizing pericyte migration along growing sprouts. As endothelial cells extend and retract in a dynamic competition for the tip cell position [108, 109], pericytes likely engage recruitment ligands via receptor binding and dynamically reduce or deplete local concentrations of these chemo-attractants at the leading front.
Figure 2.
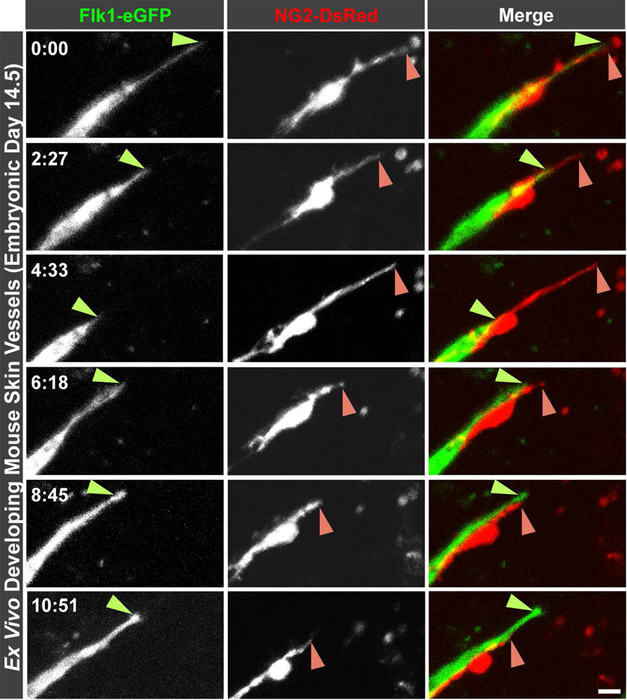
Time-lapse images of a sprouting endothelial cell visualized by eGFP expression from the Flk-1 promoter (left column, and green in right column) and an associated pericyte expressing the fluorescent reporter DsRed from the Ng2/Cspg4 promoter (middle column, and red in the right column) in an ex vivo model of vessel formation in the developing back skin from embryonic day 14.5 mice. Light green arrowheads indicate the apparent front of the endothelial tip cell, and pink arrowheads indicate the apparent leading edge of the associated pericyte. Time in the upper left corner of the left column indicates hours and minutes as hh:mm. Scale bar, 10 microns. Non-consecutive images were taken from the time-lapse sequence provided in Supplemental Movie 1. See Darden et al. Angiogenesis 2019 [64] for full experimental details and animal use certifications.
Our observations (Figure 2 and Supplemental Movie 1) suggest that, if these migration cues are not rapidly replenished to maintain a concentration gradient [84], pericytes may retract and pause their migration until their endothelial counterparts advance and deposit additional cues such as PDGF-BB. The spatial distribution of these cues may also be shaped by the pericytes themselves, as pericytes are capable of generating soluble isoforms of critical receptors such as PDGFRβ [64, 110–114]. This hypothetical model also suggests that a baseline level of retention/guidance factors must be present to maintain pericyte position alongside the sprouting endothelial cell. This proposed mechanism may also keep pericytes from detaching and migrating into the interstitium away from a growing vessel. While more studies will be necessary to better understand the existence of these and potentially other pericyte-retention mechanisms, pericyte loss by detachment and interstitial migration has been described for numerous disease conditions [24, 25, 94, 115–123]. It will therefore be important to further elucidate this phenomenon in healthy and pathological conditions.
This dynamic positioning of pericytes alongside endothelial cells during vessel formation raises several questions about the pericyte-endothelial cell interface as angiogenic sprouting unfolds. For instance, as the pericyte remains extended (Figure 2 and Supplemental Movie 1), it must adhere to and migrate along a permissive substrate, which might be the endothelial cell surface itself. Alternatively, pericytes may engage an intermediate form of the vBM that is being jointly produced by the endothelium and associated pericytes. Pericytes appear to prefer only “touch-point” contacts with the endothelium (and with other pericytes [124]), as seen in “peg-and-socket” connections between the pericyte and endothelial cell compartments in more mature vessels [29, 125–127]. It is therefore intriguing to speculate that pericytes migrate primarily along a unique configuration of the vBM that must be composed of specific ECM components, deposited at appropriate concentrations. Col-IV for example is known to be a major constituent of the mature vBM and may mediate adequate pericyte adhesion. Col-IV levels must be maintained within a narrow physiological range however, as substrates dominated by Col-IV have been shown to be non-permissive for cell adhesion in a number of in vitro systems [128, 129]. Laminins, which can integrate within the Col-IV scaffold [92, 130, 131], are also important elements of the vBM [91, 95, 132]. In addition, both pericytes and endothelial cells can secrete distinct isoforms of these ECM proteins [50, 94, 120]. Pericytes must also utilize a corresponding set of integrins such as the α2, αV, β1, and β3 subunits to facilitate their interaction with this pro-migration vBM [50, 133–135]. As vessel formation progresses, pericytes synthesize and deposit an additional, and likely molecularly distinct, “outer layer” of the vBM that contributes to the eventual stabilization of nascent vessels (Figure 3). This outer-most layer of the vBM may limit non-vascular cell types (e.g. fibroblasts) from penetrating the vBM and may therefore restrict their inclusion within the vessel wall [136]. With the vBM being established by both endothelial cells and pericytes throughout sprouting angiogenesis [109], it is critical therefore that the activities of these two cell types are well integrated such that pericyte coverage and vBM/ECM deposition keep pace with endothelial cell sprouting and new vessel branch formation.
Figure 3.

Schematic illustrating the interactions between a sprouting endothelial cell (EC, green) and an associated pericyte (PC, orange) during angiogenic remodeling. (A) Pericyte recruitment and retention factors (yellow) are secreted by endothelial tip cells to maintain pericyte coverage during vessel formation, while endothelial cell-derived extracellular matrix (ECM) proteins are also deposited (purple). (B) As pericytes establish coverage along endothelial sprouts, they secrete additional ECM components (brown) adjacent to the vessel wall to form the vascular basement membrane supporting cell migration as well as vessel stability.
Pericytes in the Transition from Angiogenic Sprouting to Vessel Maturation
Along with vBM production, pericytes have been described as playing additional roles in endothelial cell sprouting and facilitating the transition of new blood vessels from immature and highly plastic towards stable and quiescent conduits. Angiopoietin-1 (Ang-1), for instance, is produced by pericytes, along with other mesenchymal cells. Ang-1 stabilizes microvascular endothelial cells via Tie2 signaling [137], limits vessel permeability [138], and promotes quiescence [139, 140] within a vascular network [4]. In addition, pericytes may influence endothelial tip cell competition and Notch pathway signaling dynamics by physically and spatially restricting the distribution of these signals [103]. Pericytes may also influence endothelial Jagged1 (Jag1) availability through the binding of Notch receptors on the pericyte surface. These cell-cell interactions may in turn impact Delta-like 4 (Dll4)/Jag1 binding of Notch1 receptors on adjacent endothelial cells [141]. Pericytes have also been described as regulators of VEGF-A signaling through the production of VEGF Receptor-1 (VEGFR-1/Flt-1) [142, 143], though pericyte expression of VEGF-A receptors may be context-dependent. Several reports from a variety of experimental models support the notion that pericytes generally lack VEGF receptor expression [50, 51, 53–55, 144–147]. In the context of pro-angiogenic cues stimulating the remodeling of established vessels, pericytes likely contribute to the transient degradation of the vBM to permit sprouting endothelial cells to migrate outward from existing vessels [148, 149] (as well as in certain inflammation states [150]). This process must be tightly regulated, as widespread or elevated levels of matrix-degrading enzymes secreted by pericytes would lead to pathological conditions. For instance, pericyte-mediated vessel destabilization and rupture [151] has been implicated in age-related retinopathy [152] and in neonatal intraventricular hemorrhage [98, 153]. Therefore the crosstalk between pericytes and endothelial cells at multiple levels ensures that their coordinated activities yield blood vessels that are (i) patterned correctly and (ii) capable of transitioning quickly and efficiently into stable conduits for blood flow.
As with angiogenic sprouting, pericytes may contribute to other phases of vessel formation in ways that have previously been underappreciated. While endothelial cells are likely capable of completing several of these stages unilaterally, pericytes may be more involved than initially thought. Pericyte-focused tools and models are continuing to emerge, and it is therefore worth revisiting many of these processes with an eye towards understanding unique pericyte contributions [154]. For example, the mechanisms underlying vessel anastomosis and stabilization [155–162] may entail a component of pericyte regulation. Specifically, pericytes may influence precisely where endothelial cells may connect and form branch points, similar to mechanisms described for macrophages [158]. Pericytes have been described as important regulators of vessel pruning and regression [163, 164], thus contributing to the refinement of an initial microvascular network towards its final configuration. Mechanisms underlying blood vessel lumen formation may also be uniquely influenced by the presence of pericytes. Apical-basal polarity of microvascular endothelial cells may be enhanced or reinforced through contact with pericytes and/or pericyte-derived vBM proteins on their abluminal surface. In particular, pericytes form unique junctions with endothelial cells. For example, gap junctions between pericytes and endothelial cells are likely composed of hemi-channels containing Connexin43 (Cx43) and Connexin45 (Cx45) [165, 166], though other connexins may also be involved. Pericytes also form adherens junctions such as through N-cadherin [78, 127, 167] that may orient the polarity of endothelial cells as well as pericytes themselves. These intercellular junctions may enhance pericyte-specific processes, including differentiation into vascular-specific mural cells [165, 166]. They may also provide important luminal-abluminal polarity cues to the developing endothelium. As nascent microvessels acquire perfusion, the biochemical and mechanical inputs from blood flow reinforce these processes and likely provide additional vessel maturation cues to the endothelium as well as to the pericyte compartment.
As blood vessels mature into a stable and hierarchical network, they acquire contractile cells capable of shifting blood flow to regions of high metabolic activity though coordinated vasoconstriction and vasodilation. Vascular SMCs on arteries and arterioles provide this regulation of perfusion to downstream tissues. In certain instances, such as for the coronary arteries of the developing mouse heart, pericytes provide a source for these vSMCs [56]. Within the microcirculation, regulation of blood vessel diameter has been ascribed in part to vascular pericytes [17, 20, 22, 29, 168–172]. Recent work by Hill et al., however, cautions that advances in experimental approaches are needed to more fully resolve this question, specifically in the CNS [18]. One concern raised in this study is that microvascular pericytes appear to lack the morphological and biochemical features necessary to constrict or dilate the vessel lumen. In particular, it is not entirely clear whether pericytes sufficiently “wrap around” microvessels (i.e. concentrically) and/or express α-smooth muscle actin (αSMA) [173] or other contractile machinery proteins necessary to elicit vessel diameter changes [18]. Advances in imaging modalities, single-cell transcriptional profiling, and tissue-processing techniques [23] will certainly shed more light on this potential pericyte function. Furthermore, resolving this question may inform clinical approaches to managing serious pathological conditions such as coronary no-reflow [169, 174], ischemic stroke [16], and spinal cord injury [121]. In each of these situations, maintaining adequate tissue perfusion following the primary insult remains a challenge and could involve targeting pericytes. We recently found that disrupting oxygen-sensing mechanisms via induced genetic Vhl mutations accelerated vessel maturation and specifically caused pericytes within developing retinal vasculature to ectopically express αSMA [175]. Taken together with previous studies, it is intriguing to speculate that the hypoxic conditions within several of the aforementioned disease states might not only antagonize pericyte constriction mechanisms [16, 170, 176], but also cause an aberrant increase in the expression of αSMA and other contractile machinery proteins. This “hyper-muscularization” of pericytes has also been reported in other contexts [177]. This working model requires further studies to determine if such relationships exist. In summary, in addition to their potential role in modulating capillary tone, pericytes within the microcirculation contribute to a number of important processes that promote the transition of microvessels from sprouting and remodeling to mature, quiescent networks.
Current and Forward-looking Perspectives on Pericytes
The recent surge in studies exploring the potential roles for pericytes in developmental and disease processes allows us to revise and extend existing models for pericyte function within the microcirculation. New tools and models will provide a more detailed understanding of the well-established roles of pericytes in vessel stabilization during angiogenic sprouting and barrier function [6–8, 15] as well as their contributions within specific tissues and organs and potential influence on diverse cell populations [178]. Though certain challenges remain in studying pericytes in a variety of biological contexts such as during angiogenesis, it will be important to overcome these hurdles and further clarify the roles that pericytes play in the formation and homeostasis of the microcirculation. Pericytes are implicated in the onset and progression of numerous pathological conditions such as tumor vascularization and dysfunctional angiogenesis in diabetes-and mutation-associated (e.g. VHL) retinopathies. Their connection to these and other disease states warrants extensive investigation into their behaviors and the underlying signaling mechanisms. Such insight will inspire new ideas for targeting pericytes in the treatment and management of these and other vascular-related conditions such as tissue fibrosis, blood-brain barrier dysfunction, and metastatic disease. Clinical strategies are continuing to emerge wherein pericytes are being considered as viable therapeutic targets, and this trend is likely to persist well into the future.
Our knowledge of the pericyte microenvironment during vascular development will also continue to build upon the solid foundation of the studies discussed herein, as well as many others. Nevertheless, fundamental challenges and questions within this line of research remain to be fully addressed, making it an emerging topic that is gaining considerable interest and momentum in the field of vascular biology. In particular, the criteria for confident identification of a bona fide vascular pericyte must continue to evolve and strengthen to enhance the rigor and utility of pericyte-focused studies. Maintaining and/or restoring pericyte coverage within the microcirculation has gained significant attention as a therapeutic target for many pathological conditions. It will therefore be critical to investigate pericytes within the sprouting angiogenesis context to better understand the molecular determinants for establishing and sustaining pericyte investment within the microvessel wall. An equally important goal will be to address the converse relationship, identifying how pericytes might influence their endothelial cell counterparts during each stage of vessel formation. Furthermore, across discrete vascular beds, the genetic and molecular mechanisms underlying pericyte-mediated vessel stability may vary during vascular development. Elucidating the similarities and differences will shed light on how specific processes such as vBM deposition and junction formation are regulated in promoting vessel quiescence. As the tools and models for studying pericytes continue to advance, so too will our insight into these and many other open questions regarding pericytes and their essential contributions to building and maintaining the microcirculation.
Supplementary Material
Acknowledgements
We thank the Chappell and Smyth Labs for critical and extensive discussions of published data.
Sources of Funding.
This work was supported by NIH grants R00HL105779 and R56HL133826 and NSF grant 1752339 (to JCC).
Abbreviations:
- ECM
Extracellular Matrix
- CNS
Central Nervous System
- vSMC
vascular Smooth Muscle Cell
- PDGFRβ
Platelet-Derived Growth Factor Receptor-β
- NG2
Neural Glial Antigen-2
- AVM
Arterio-Venous Malformation
- CCM
Cerebral Cavernous Malformation
- vBM
vascular Basement Membrane
- SEM
Scanning Electron Microscopy
- Col-IV
Type IV Collagen
- VEGF-A
Vascular Endothelial Growth Factor-A
- αSMA
α-Smooth Muscle Actin
- MSC
mesenchymal stem cell
- ESC
embryonic stem cell
Footnotes
COI Statement: The authors have declared that no conflict of interest exists.
Disclosures.
None
REFERENCES
- 1.Sims DE, The pericyte--a review. Tissue Cell, 1986. 18(2): p. 153–74. [DOI] [PubMed] [Google Scholar]
- 2.Rouget C, Memoire sur le developpement, la structure et les proprietes physiologiques des capillaires sanguins. Archives Physiol. Normale Pathol, 1873. 5: p. 603–661. [Google Scholar]
- 3.Eberth CJ, Handbuch der-Lehre von den Gewebwn des Menschen und der Tiere 1871. Vol 1(Leipzig: Engelmann; ). [Google Scholar]
- 4.Armulik A, Genove G, and Betsholtz C, Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev Cell, 2011. 21(2): p. 193–215. [DOI] [PubMed] [Google Scholar]
- 5.Zhao Z, et al. , Establishment and Dysfunction of the Blood-Brain Barrier. Cell, 2015. 163(5): p. 1064–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Armulik A, et al. , Pericytes regulate the blood-brain barrier. Nature, 2010. 468(7323): p. 557–61. [DOI] [PubMed] [Google Scholar]
- 7.Daneman R, et al. , Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature, 2010. 468(7323): p. 562–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bell RD, et al. , Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron, 2010. 68(3): p. 409–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kern TS, et al. , Response of capillary cell death to aminoguanidine predicts the development of retinopathy: comparison of diabetes and galactosemia. Invest Ophthalmol Vis Sci, 2000. 41(12): p. 3972–8. [PubMed] [Google Scholar]
- 10.Barber AJ, Gardner TW, and Abcouwer SF, The significance of vascular and neural apoptosis to the pathology of diabetic retinopathy. Invest Ophthalmol Vis Sci, 2011. 52(2): p. 1156–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Romeo G, et al. , Activation of nuclear factor-kappaB induced by diabetes and high glucose regulates a proapoptotic program in retinal pericytes. Diabetes, 2002. 51(7): p. 2241–8. [DOI] [PubMed] [Google Scholar]
- 12.Geraldes P, et al. , Activation of PKC-delta and SHP-1 by hyperglycemia causes vascular cell apoptosis and diabetic retinopathy. Nat Med, 2009. 15(11): p. 1298–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen Z, Xu XH, and Hu J, Role of pericytes in angiogenesis: focus on cancer angiogenesis and anti-angiogenic therapy. Neoplasma, 2016. 63(2): p. 173–82. [DOI] [PubMed] [Google Scholar]
- 14.Hanahan D and Weinberg RA, Hallmarks of cancer: the next generation. Cell, 2011. 144(5): p. 646–74. [DOI] [PubMed] [Google Scholar]
- 15.Sagare AP, et al. , Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nat Commun, 2013. 4: p. 2932. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 16.Hall CN, et al. , Capillary pericytes regulate cerebral blood flow in health and disease. Nature, 2014. 508(7494): p. 55–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mishra A, et al. , Imaging pericytes and capillary diameter in brain slices and isolated retinae. Nat Protoc, 2014. 9(2): p. 323–36. [DOI] [PubMed] [Google Scholar]
- 18.Hill RA, et al. , Regional Blood Flow in the Normal and Ischemic Brain Is Controlled by Arteriolar Smooth Muscle Cell Contractility and Not by Capillary Pericytes. Neuron, 2015. 87(1): p. 95–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hillman EM, Coupling mechanism and significance of the BOLD signal: a status report. Annu Rev Neurosci, 2014. 37: p. 161–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fernandez-Klett F, et al. , Pericytes in capillaries are contractile in vivo, but arterioles mediate functional hyperemia in the mouse brain. Proc Natl Acad Sci U S A, 2010. 107(51): p. 22290–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sweeney PW, Walker-Samuel S, and Shipley RJ, Insights into cerebral haemodynamics and oxygenation utilising in vivo mural cell imaging and mathematical modelling. Sci Rep, 2018. 8(1): p. 1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Peppiatt CM, et al. , Bidirectional control of CNS capillary diameter by pericytes. Nature, 2006. 443(7112): p. 700–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Alarcon-Martinez L, et al. , Capillary pericytes express alpha-smooth muscle actin, which requires prevention of filamentous-actin depolymerization for detection. Elife, 2018. 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Birbrair A, et al. , Pericytes at the intersection between tissue regeneration and pathology. Clin Sci (Lond), 2015. 128(2): p. 81–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Birbrair A, et al. , Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner. Stem Cell Res Ther, 2014. 5(6): p. 122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Birbrair A, et al. , Role of pericytes in skeletal muscle regeneration and fat accumulation. Stem Cells Dev, 2013. 22(16): p. 2298–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Crisan M, et al. , A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell, 2008. 3(3): p. 301–13. [DOI] [PubMed] [Google Scholar]
- 28.Caplan AI, Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J Cell Physiol, 2007. 213(2): p. 341–7. [DOI] [PubMed] [Google Scholar]
- 29.Diaz-Flores L, et al. , Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol Histopathol, 2009. 24(7): p. 909–69. [DOI] [PubMed] [Google Scholar]
- 30.Davidoff MS, et al. , Progenitor cells of the testosterone-producing Leydig cells revealed. J Cell Biol, 2004. 167(5): p. 935–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Crisan M, et al. , Purification and long-term culture of multipotent progenitor cells affiliated with the walls of human blood vessels: myoendothelial cells and pericytes. Methods Cell Biol, 2008. 86: p. 295–309. [DOI] [PubMed] [Google Scholar]
- 32.Feng J, et al. , Dual origin of mesenchymal stem cells contributing to organ growth and repair. Proc Natl Acad Sci U S A, 2011. 108(16): p. 6503–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Olson LE and Soriano P, PDGFRbeta signaling regulates mural cell plasticity and inhibits fat development. Dev Cell, 2011. 20(6): p. 815–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tang W, et al. , White fat progenitor cells reside in the adipose vasculature. Science, 2008. 322(5901): p. 583–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Dore-Duffy P, et al. , CNS microvascular pericytes exhibit multipotential stem cell activity. J Cereb Blood Flow Metab, 2006. 26(5): p. 613–24. [DOI] [PubMed] [Google Scholar]
- 36.Dellavalle A, et al. , Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat Cell Biol, 2007. 9(3): p. 255–67. [DOI] [PubMed] [Google Scholar]
- 37.Paquet-Fifield S, et al. , A role for pericytes as microenvironmental regulators of human skin tissue regeneration. J Clin Invest, 2009. 119(9): p. 2795–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yianni V and Sharpe PT, Molecular programming of perivascular stem cell precursors. Stem Cells, 2018. [DOI] [PubMed]
- 39.Guimaraes-Camboa N, et al. , Pericytes of Multiple Organs Do Not Behave as Mesenchymal Stem Cells In Vivo. Cell Stem Cell, 2017. 20(3): p. 345–359 e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Berthiaume AA, et al. , Pericyte Structural Remodeling in Cerebrovascular Health and Homeostasis. Front Aging Neurosci, 2018. 10: p. 210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Winkler EA, Bell RD, and Zlokovic BV, Central nervous system pericytes in health and disease. Nat Neurosci, 2011. 14(11): p. 1398–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kelly-Goss MR, et al. , Targeting pericytes for angiogenic therapies. Microcirculation, 2014. 21(4): p. 345–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Trost A, et al. , Brain and Retinal Pericytes: Origin, Function and Role. Front Cell Neurosci, 2016. 10: p. 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Armulik A, Abramsson A, and Betsholtz C, Endothelial/pericyte interactions. Circ Res, 2005. 97(6): p. 512–23. [DOI] [PubMed] [Google Scholar]
- 45.Gerhardt H and Betsholtz C, Endothelial-pericyte interactions in angiogenesis. Cell Tissue Res, 2003. 314(1): p. 15–23. [DOI] [PubMed] [Google Scholar]
- 46.Balabanov R and Dore-Duffy P, Role of the CNS microvascular pericyte in the blood-brain barrier. J Neurosci Res, 1998. 53(6): p. 637–44. [DOI] [PubMed] [Google Scholar]
- 47.Trost A, et al. , Neural crest origin of retinal and choroidal pericytes. Invest Ophthalmol Vis Sci, 2013. 54(13): p. 7910–21. [DOI] [PubMed] [Google Scholar]
- 48.Korn J, Christ B, and Kurz H, Neuroectodermal origin of brain pericytes and vascular smooth muscle cells. J Comp Neurol, 2002. 442(1): p. 78–88. [DOI] [PubMed] [Google Scholar]
- 49.Hartmann DA, et al. , Pericyte structure and distribution in the cerebral cortex revealed by high-resolution imaging of transgenic mice. Neurophotonics, 2015. 2(4). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.He L, et al. , Analysis of the brain mural cell transcriptome. Sci Rep, 2016. 6: p. 35108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hrvatin S, et al. , Single-cell analysis of experience-dependent transcriptomic states in the mouse visual cortex. Nat Neurosci, 2018. 21(1): p. 120–129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang Y, et al. , An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci, 2014. 34(36): p. 11929–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Daneman R, et al. , The mouse blood-brain barrier transcriptome: a new resource for understanding the development and function of brain endothelial cells. PLoS One, 2010. 5(10): p. e13741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zeisel A, et al. , Molecular Architecture of the Mouse Nervous System. Cell, 2018. 174(4): p. 999–1014 e22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zeisel A, et al. , Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science, 2015. 347(6226): p. 1138–42. [DOI] [PubMed] [Google Scholar]
- 56.Volz KS, et al. , Pericytes are progenitors for coronary artery smooth muscle. Elife, 2015. 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.High FA, et al. , Endothelial expression of the Notch ligand Jagged1 is required for vascular smooth muscle development. Proc Natl Acad Sci U S A, 2008. 105(6): p. 1955–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kofler NM, et al. , Combined deficiency of Notch1 and Notch3 causes pericyte dysfunction, models CADASIL, and results in arteriovenous malformations. Sci Rep, 2015. 5: p. 16449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Liu H, Kennard S, and Lilly B, NOTCH3 expression is induced in mural cells through an autoregulatory loop that requires endothelial-expressed JAGGED1. Circ Res, 2009. 104(4): p. 466–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Domenga V, et al. , Notch3 is required for arterial identity and maturation of vascular smooth muscle cells. Genes Dev, 2004. 18(22): p. 2730–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jin S, et al. , Notch signaling regulates platelet-derived growth factor receptor-beta expression in vascular smooth muscle cells. Circ Res, 2008. 102(12): p. 1483–91. [DOI] [PubMed] [Google Scholar]
- 62.Henshall TL, et al. , Notch3 is necessary for blood vessel integrity in the central nervous system. Arterioscler Thromb Vasc Biol, 2015. 35(2): p. 409–20. [DOI] [PubMed] [Google Scholar]
- 63.Cuervo H, et al. , PDGFRbeta-P2A-CreER(T2) mice: a genetic tool to target pericytes in angiogenesis. Angiogenesis, 2017. 20(4): p. 655–662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Darden J, et al. , Excess vascular endothelial growth factor-A disrupts pericyte recruitment during blood vessel formation. Angiogenesis, 2019. 22(1): p. 167–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wang Y, et al. , Notch3 establishes brain vascular integrity by regulating pericyte number. Development, 2014. 141(2): p. 307–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hellstrom M, et al. , Role of PDGF-B and PDGFR-beta in recruitment of vascular smooth muscle cells and pericytes during embryonic blood vessel formation in the mouse. Development, 1999. 126(14): p. 3047–55. [DOI] [PubMed] [Google Scholar]
- 67.Trotter J, Karram K, and Nishiyama A, NG2 cells: Properties, progeny and origin. Brain Res Rev, 2010. 63(1–2): p. 72–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhu X, Bergles DE, and Nishiyama A, NG2 cells generate both oligodendrocytes and gray matter astrocytes. Development, 2008. 135(1): p. 145–57. [DOI] [PubMed] [Google Scholar]
- 69.Murfee WL, et al. , Perivascular cells along venules upregulate NG2 expression during microvascular remodeling. Microcirculation, 2006. 13(3): p. 261–73. [DOI] [PubMed] [Google Scholar]
- 70.French WJ, Creemers EE, and Tallquist MD, Platelet-derived growth factor receptors direct vascular development independent of vascular smooth muscle cell function. Mol Cell Biol, 2008. 28(18): p. 5646–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Jung B, et al. , Visualization of vascular mural cells in developing brain using genetically labeled transgenic reporter mice. J Cereb Blood Flow Metab, 2018. 38(3): p. 456–468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bergers G and Song S, The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol, 2005. 7(4): p. 452–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Marmé D. and Fusenig NE, Tumor angiogenesis : basic mechanisms and cancer therapy 2008, New York: Springer; xviii, 845 p. [Google Scholar]
- 74.Ando K, et al. , Clarification of mural cell coverage of vascular endothelial cells by live imaging of zebrafish. Development, 2016. 143(8): p. 1328–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zhao H, Darden J, and Chappell JC, Establishment and characterization of an embryonic pericyte cell line. Microcirculation, 2018. 25(5): p. e12461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Jakobsson L, Kreuger J, and Claesson-Welsh L, Building blood vessels--stem cell models in vascular biology. J Cell Biol, 2007. 177(5): p. 751–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jakobsson L, et al. , Heparan sulfate in trans potentiates VEGFR-mediated angiogenesis. Dev Cell, 2006. 10(5): p. 625–34. [DOI] [PubMed] [Google Scholar]
- 78.Tillet E, et al. , N-cadherin deficiency impairs pericyte recruitment, and not endothelial differentiation or sprouting, in embryonic stem cell-derived angiogenesis. Exp Cell Res, 2005. 310(2): p. 392–400. [DOI] [PubMed] [Google Scholar]
- 79.Winkler EA, et al. , Reductions in brain pericytes are associated with arteriovenous malformation vascular instability. J Neurosurg, 2018: p. 1–11. [DOI] [PMC free article] [PubMed]
- 80.Gaengel K, et al. , Endothelial-mural cell signaling in vascular development and angiogenesis. Arterioscler Thromb Vasc Biol, 2009. 29(5): p. 630–8. [DOI] [PubMed] [Google Scholar]
- 81.Enge M, et al. , Endothelium-specific platelet-derived growth factor-B ablation mimics diabetic retinopathy. EMBO J, 2002. 21(16): p. 4307–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Hirschi KK, et al. , Endothelial cells modulate the proliferation of mural cell precursors via platelet-derived growth factor-BB and heterotypic cell contact. Circ Res, 1999. 84(3): p. 298–305. [DOI] [PubMed] [Google Scholar]
- 83.Abramsson A, et al. , Defective N-sulfation of heparan sulfate proteoglycans limits PDGF-BB binding and pericyte recruitment in vascular development. Genes Dev, 2007. 21(3): p. 316–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lindblom P, et al. , Endothelial PDGF-B retention is required for proper investment of pericytes in the microvessel wall. Genes Dev, 2003. 17(15): p. 1835–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Abramsson A, Lindblom P, and Betsholtz C, Endothelial and nonendothelial sources of PDGF-B regulate pericyte recruitment and influence vascular pattern formation in tumors. J Clin Invest, 2003. 112(8): p. 1142–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lindahl P, et al. , Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science, 1997. 277(5323): p. 242–5. [DOI] [PubMed] [Google Scholar]
- 87.Kisler K, et al. , Pericyte degeneration leads to neurovascular uncoupling and limits oxygen supply to brain. Nat Neurosci, 2017. [DOI] [PMC free article] [PubMed]
- 88.Stratman AN, et al. , Endothelial-derived PDGF-BB and HB-EGF coordinately regulate pericyte recruitment during vasculogenic tube assembly and stabilization. Blood, 2010. 116(22): p. 4720–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Stratman AN, Davis MJ, and Davis GE, VEGF and FGF prime vascular tube morphogenesis and sprouting directed by hematopoietic stem cell cytokines. Blood, 2011. 117(14): p. 3709–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Song N, et al. , Overexpression of platelet-derived growth factor-BB increases tumor pericyte content via stromal-derived factor-1alpha/CXCR4 axis. Cancer Res, 2009. 69(15): p. 6057–64. [DOI] [PubMed] [Google Scholar]
- 91.Davis GE and Senger DR, Endothelial extracellular matrix: biosynthesis, remodeling, and functions during vascular morphogenesis and neovessel stabilization. Circ Res, 2005. 97(11): p. 1093–107. [DOI] [PubMed] [Google Scholar]
- 92.Kalluri R, Basement membranes: structure, assembly and role in tumour angiogenesis. Nat Rev Cancer, 2003. 3(6): p. 422–33. [DOI] [PubMed] [Google Scholar]
- 93.Sweeney MD, Ayyadurai S, and Zlokovic BV, Pericytes of the neurovascular unit: key functions and signaling pathways. Nat Neurosci, 2016. 19(6): p. 771–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sava P, et al. , Human microvascular pericyte basement membrane remodeling regulates neutrophil recruitment. Microcirculation, 2015. 22(1): p. 54–67. [DOI] [PubMed] [Google Scholar]
- 95.Stratman AN, et al. , Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood, 2009. 114(24): p. 5091–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Egginton S, et al. , In vivo pericyte-endothelial cell interaction during angiogenesis in adult cardiac and skeletal muscle. Microvasc Res, 1996. 51(2): p. 213–28. [DOI] [PubMed] [Google Scholar]
- 97.Hansen-Smith FM, Hudlicka O, and Egginton S, In vivo angiogenesis in adult rat skeletal muscle: early changes in capillary network architecture and ultrastructure. Cell Tissue Res, 1996. 286(1): p. 123–36. [DOI] [PubMed] [Google Scholar]
- 98.Dave JM, et al. , Pericyte ALK5/TIMP3 Axis Contributes to Endothelial Morphogenesis in the Developing Brain. Dev Cell, 2018. [DOI] [PMC free article] [PubMed]
- 99.Alavi MV, et al. , Col4a1 mutations cause progressive retinal neovascular defects and retinopathy. Sci Rep, 2016. 6: p. 18602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Baluk P, et al. , Abnormalities of basement membrane on blood vessels and endothelial sprouts in tumors. Am J Pathol, 2003. 163(5): p. 1801–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Capone C, et al. , Reducing Timp3 or vitronectin ameliorates disease manifestations in CADASIL mice. Ann Neurol, 2016. 79(3): p. 387–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Yao Y, et al. , Astrocytic laminin regulates pericyte differentiation and maintains blood brain barrier integrity. Nat Commun, 2014. 5: p. 3413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Walpole J, et al. , Agent-based Computational Model of Retinal Angiogenesis Simulates Microvascular Network Morphology as a Function of Pericyte Coverage. Microcirculation, 2017. [DOI] [PMC free article] [PubMed]
- 104.Seynhaeve ALB, et al. , Spatiotemporal endothelial cell-pericyte association in tumors as shown by high resolution 4D intravital imaging. Sci Rep, 2018. 8(1): p. 9596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Eglinger J, Karsjens H, and Lammert E, Quantitative assessment of angiogenesis and pericyte coverage in human cell-derived vascular sprouts. Inflammation and Regeneration, 2017. 37(1): p. 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Chang WG, et al. , Pericytes modulate endothelial sprouting. Cardiovasc Res, 2013. 100(3): p. 492–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Hellstrom M, et al. , Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature, 2007. 445(7129): p. 776–80. [DOI] [PubMed] [Google Scholar]
- 108.Jakobsson L, et al. , Endothelial cells dynamically compete for the tip cell position during angiogenic sprouting. Nat Cell Biol, 2010. 12(10): p. 943–53. [DOI] [PubMed] [Google Scholar]
- 109.Ubezio B, et al. , Synchronization of endothelial Dll4-Notch dynamics switch blood vessels from branching to expansion. Elife, 2016. 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Sagare AP, et al. , Shedding of soluble platelet-derived growth factor receptor-beta from human brain pericytes. Neurosci Lett, 2015. 607: p. 97–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Hutter-Schmid B and Humpel C, Platelet-derived Growth Factor Receptor-beta is Differentially Regulated in Primary Mouse Pericytes and Brain Slices. Curr Neurovasc Res, 2016. 13(2): p. 127–34. [DOI] [PubMed] [Google Scholar]
- 112.Hosaka K, et al. , Pericyte-fibroblast transition promotes tumor growth and metastasis. Proc Natl Acad Sci U S A, 2016. 113(38): p. E5618–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Mendelson K, et al. , Stimulation of platelet-derived growth factor receptor beta (PDGFRbeta) activates ADAM17 and promotes metalloproteinase-dependent cross-talk between the PDGFRbeta and epidermal growth factor receptor (EGFR) signaling pathways. J Biol Chem, 2010. 285(32): p. 25024–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Duan DS, et al. , A functional soluble extracellular region of the platelet-derived growth factor (PDGF) beta-receptor antagonizes PDGF-stimulated responses. J Biol Chem, 1991. 266(1): p. 413–8. [PubMed] [Google Scholar]
- 115.Pfister F, et al. , Pericyte migration: a novel mechanism of pericyte loss in experimental diabetic retinopathy. Diabetes, 2008. 57(9): p. 2495–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Barron L, Gharib SA, and Duffield JS, Lung Pericytes and Resident Fibroblasts: Busy Multitaskers. Am J Pathol, 2016. 186(10): p. 2519–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Stefanska A, et al. , Interstitial pericytes decrease in aged mouse kidneys. Aging (Albany NY), 2015. 7(6): p. 370–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Schrimpf C, et al. , The role of pericyte detachment in vascular rarefaction. J Vasc Res, 2014. 51(4): p. 247–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Lin SL, et al. , Pericytes and perivascular fibroblasts are the primary source of collagen-producing cells in obstructive fibrosis of the kidney. Am J Pathol, 2008. 173(6): p. 1617–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Sava P, et al. , Human pericytes adopt myofibroblast properties in the microenvironment of the IPF lung. JCI Insight, 2017. 2(24). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Li Y, et al. , Pericytes impair capillary blood flow and motor function after chronic spinal cord injury. Nat Med, 2017. 23(6): p. 733–741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Fernandez-Klett F, et al. , Early loss of pericytes and perivascular stromal cell-induced scar formation after stroke. J Cereb Blood Flow Metab, 2013. 33(3): p. 428–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Mendel TA, et al. , Pericytes derived from adipose-derived stem cells protect against retinal vasculopathy. PLoS One, 2013. 8(5): p. e65691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Berthiaume AA, et al. , Dynamic Remodeling of Pericytes In Vivo Maintains Capillary Coverage in the Adult Mouse Brain. Cell Rep, 2018. 22(1): p. 8–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Cuevas P, et al. , Pericyte endothelial gap junctions in human cerebral capillaries. Anat Embryol (Berl), 1984. 170(2): p. 155–9. [DOI] [PubMed] [Google Scholar]
- 126.Tilton RG, Kilo C, and Williamson JR, Pericyte-endothelial relationships in cardiac and skeletal muscle capillaries. Microvasc Res, 1979. 18(3): p. 325–35. [DOI] [PubMed] [Google Scholar]
- 127.Gerhardt H, Wolburg H, and Redies C, N-cadherin mediates pericytic-endothelial interaction during brain angiogenesis in the chicken. Dev Dyn, 2000. 218(3): p. 472–9. [DOI] [PubMed] [Google Scholar]
- 128.Fox MA, et al. , Distinct target-derived signals organize formation, maturation, and maintenance of motor nerve terminals. Cell, 2007. 129(1): p. 179–93. [DOI] [PubMed] [Google Scholar]
- 129.Fox MA, et al. , Growth conelike sensorimotor structures are characteristic features of postmigratory, premyelinating oligodendrocytes. Glia, 2006. 53(5): p. 563–6. [DOI] [PubMed] [Google Scholar]
- 130.Timpl R and Brown JC, Supramolecular assembly of basement membranes. Bioessays, 1996. 18(2): p. 123–32. [DOI] [PubMed] [Google Scholar]
- 131.Charonis AS, et al. , Binding of laminin to type IV collagen: a morphological study. J Cell Biol, 1985. 100(6): p. 1848–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Jakobsson L, et al. , Laminin deposition is dispensable for vasculogenesis but regulates blood vessel diameter independent of flow. FASEB J, 2007. [DOI] [PubMed] [Google Scholar]
- 133.Tigges U, et al. , TNF-alpha promotes cerebral pericyte remodeling in vitro, via a switch from alpha1 to alpha2 integrins. J Neuroinflammation, 2013. 10: p. 33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Hurtado-Alvarado G, Cabanas-Morales AM, and Gomez-Gonzalez B, Pericytes: brain-immune interface modulators. Front Integr Neurosci, 2014. 7: p. 80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Hinz B, It has to be the alphav: myofibroblast integrins activate latent TGF-beta1. Nat Med, 2013. 19(12): p. 1567–8. [DOI] [PubMed] [Google Scholar]
- 136.Rock JR, et al. , Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc Natl Acad Sci U S A, 2011. 108(52): p. E1475–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Falcon BL, et al. , Contrasting actions of selective inhibitors of angiopoietin-1 and angiopoietin-2 on the normalization of tumor blood vessels. Am J Pathol, 2009. 175(5): p. 2159–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Thurston G, et al. , Leakage-resistant blood vessels in mice transgenically overexpressing angiopoietin-1. Science, 1999. 286(5449): p. 2511–4. [DOI] [PubMed] [Google Scholar]
- 139.Dumont DJ, et al. , tek, a novel tyrosine kinase gene located on mouse chromosome 4, is expressed in endothelial cells and their presumptive precursors. Oncogene, 1992. 7(8): p. 1471–80. [PubMed] [Google Scholar]
- 140.Wakui S, et al. , Localization of Ang-1, −2, Tie-2, and VEGF expression at endothelial-pericyte interdigitation in rat angiogenesis. Laboratory Investigation, 2006. 86(11): p. 1172–1184. [DOI] [PubMed] [Google Scholar]
- 141.Benedito R, et al. , The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell, 2009. 137(6): p. 1124–35. [DOI] [PubMed] [Google Scholar]
- 142.Eilken HM, et al. , Pericytes regulate VEGF-induced endothelial sprouting through VEGFR1. Nat Commun, 2017. 8(1): p. 1574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Cao R, et al. , VEGFR1-mediated pericyte ablation links VEGF and PlGF to cancer-associated retinopathy. Proc Natl Acad Sci U S A, 2010. 107(2): p. 856–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Fruttiger M, Development of the mouse retinal vasculature: angiogenesis versus vasculogenesis. Invest Ophthalmol Vis Sci, 2002. 43(2): p. 522–7. [PubMed] [Google Scholar]
- 145.Shih SC, et al. , Selective stimulation of VEGFR-1 prevents oxygen-induced retinal vascular degeneration in retinopathy of prematurity. J Clin Invest, 2003. 112(1): p. 50–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Matsumoto K, et al. , Study of normal and pathological blood vessel morphogenesis in Flt1-tdsRed BAC Tg mice. Genesis, 2012. 50(7): p. 561–71. [DOI] [PubMed] [Google Scholar]
- 147.Luo L, et al. , Photoreceptor avascular privilege is shielded by soluble VEGF receptor-1. Elife, 2013. 2: p. e00324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Potente M, Gerhardt H, and Carmeliet P, Basic and therapeutic aspects of angiogenesis. Cell, 2011. 146(6): p. 873–87. [DOI] [PubMed] [Google Scholar]
- 149.Zozulya A, Weidenfeller C, and Galla HJ, Pericyte-endothelial cell interaction increases MMP-9 secretion at the blood-brain barrier in vitro. Brain Res, 2008. 1189: p. 1–11. [DOI] [PubMed] [Google Scholar]
- 150.Machida T, et al. , Brain pericytes are the most thrombin-sensitive matrix metalloproteinase-9-releasing cell type constituting the blood-brain barrier in vitro. Neurosci Lett, 2015. 599: p. 109–14. [DOI] [PubMed] [Google Scholar]
- 151.Underly RG, et al. , Pericytes as Inducers of Rapid, Matrix Metalloproteinase-9-Dependent Capillary Damage during Ischemia. J Neurosci, 2017. 37(1): p. 129–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Lopez-Luppo M, et al. , Blood Vessel Basement Membrane Alterations in Human Retinal Microaneurysms During Aging. Invest Ophthalmol Vis Sci, 2017. 58(2): p. 1116–1131. [DOI] [PubMed] [Google Scholar]
- 153.Yang D, et al. , Overexpression of vascular endothelial growth factor in the germinal matrix induces neurovascular proteases and intraventricular hemorrhage. Sci Transl Med, 2013. 5(193): p. 193ra90. [DOI] [PubMed] [Google Scholar]
- 154.Chappell JC, et al. , Flt-1 (VEGFR-1) Coordinates Discrete Stages of Blood Vessel Formation. Cardiovasc Res, 2016. 111(1): p. 84–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Nesmith JE, et al. , Blood vessel anastomosis is spatially regulated by Flt1 during angiogenesis. Development, 2017. 144(5): p. 889–896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Siekmann AF, Affolter M, and Belting HG, The tip cell concept 10 years after: new players tune in for a common theme. Exp Cell Res, 2013. 319(9): p. 1255–63. [DOI] [PubMed] [Google Scholar]
- 157.Lenard A, et al. , In vivo analysis reveals a highly stereotypic morphogenetic pathway of vascular anastomosis. Dev Cell, 2013. 25(5): p. 492–506. [DOI] [PubMed] [Google Scholar]
- 158.Fantin A, et al. , Tissue macrophages act as cellular chaperones for vascular anastomosis downstream of VEGF-mediated endothelial tip cell induction. Blood, 2010. 116(5): p. 829–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Bentley K, et al. , Tipping the balance: robustness of tip cell selection, migration and fusion in angiogenesis. PLoS Comput Biol, 2009. 5(10): p. e1000549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Zygmunt T, et al. , Semaphorin-PlexinD1 signaling limits angiogenic potential via the VEGF decoy receptor sFlt1. Dev Cell, 2011. 21(2): p. 301–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Blum Y, et al. , Complex cell rearrangements during intersegmental vessel sprouting and vessel fusion in the zebrafish embryo. Dev Biol, 2008. 316(2): p. 312–22. [DOI] [PubMed] [Google Scholar]
- 162.Ribeiro C, Ebner A, and Affolter M, In vivo imaging reveals different cellular functions for FGF and Dpp signaling in tracheal branching morphogenesis. Dev Cell, 2002. 2(5): p. 677–83. [DOI] [PubMed] [Google Scholar]
- 163.Benjamin LE, Hemo I, and Keshet E, A plasticity window for blood vessel remodelling is defined by pericyte coverage of the preformed endothelial network and is regulated by PDGF-B and VEGF. Development, 1998. 125(9): p. 1591–8. [DOI] [PubMed] [Google Scholar]
- 164.Simonavicius N, et al. , Pericytes promote selective vessel regression to regulate vascular patterning. Blood, 2012. 120(7): p. 1516–27. [DOI] [PubMed] [Google Scholar]
- 165.Fang JS, et al. , Connexin45 regulates endothelial-induced mesenchymal cell differentiation toward a mural cell phenotype. Arterioscler Thromb Vasc Biol, 2013. 33(2): p. 362–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hirschi KK, et al. , Gap junction communication mediates transforming growth factor-beta activation and endothelial-induced mural cell differentiation. Circ Res, 2003. 93(5): p. 429–37. [DOI] [PubMed] [Google Scholar]
- 167.Paik JH, et al. , Sphingosine 1-phosphate receptor regulation of N-cadherin mediates vascular stabilization. Genes Dev, 2004. 18(19): p. 2392–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Durham JT, et al. , Pericyte chemomechanics and the angiogenic switch: insights into the pathogenesis of proliferative diabetic retinopathy? Invest Ophthalmol Vis Sci, 2015. 56(6): p. 3441–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.O’Farrell FM and Attwell D, A role for pericytes in coronary no-reflow. Nat Rev Cardiol, 2014. 11(7): p. 427–32. [DOI] [PubMed] [Google Scholar]
- 170.Hamilton NB, Attwell D, and Hall CN, Pericyte-mediated regulation of capillary diameter: a component of neurovascular coupling in health and disease. Front Neuroenergetics, 2010. 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Sieczkiewicz GJ and Herman IM, TGF-beta 1 signaling controls retinal pericyte contractile protein expression. Microvasc Res, 2003. 66(3): p. 190–6. [DOI] [PubMed] [Google Scholar]
- 172.Shih AY, et al. , Robust and fragile aspects of cortical blood flow in relation to the underlying angioarchitecture. Microcirculation, 2015. 22(3): p. 204–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Nehls V and Drenckhahn D, Heterogeneity of microvascular pericytes for smooth muscle type alpha-actin. J Cell Biol, 1991. 113(1): p. 147–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.O’Farrell FM, et al. , Capillary pericytes mediate coronary no-reflow after myocardial ischaemia. Elife, 2017. 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Arreola A, et al. , Von Hippel-Lindau mutations disrupt vascular patterning and maturation via Notch. JCI Insight, 2018. 3(4). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Attwell D, et al. , What is a pericyte? J Cereb Blood Flow Metab, 2016. 36(2): p. 451–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Ratelade J, et al. , Severity of arterial defects in the retina correlates with the burden of intracerebral haemorrhage in COL4A1-related stroke. J Pathol, 2018. 244(4): p. 408–420. [DOI] [PubMed] [Google Scholar]
- 178.Fortuna V, et al. , Vascular Mural Cells Promote Noradrenergic Differentiation of Embryonic Sympathetic Neurons. Cell Rep, 2015. 11(11): p. 1786–96. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.