

Abstract
Bisphenol A (BPA), a known endocrine disrupting chemical (EDC), was administered by diet to gilthead sea bream (Sparus aurata) in order to study its effects on the endocannabinoid system (ECS) and gonadal steroidogenesis. 2-year-old male gilthead sea bream were fed with two different concentrations of BPA (LOW at 4 and HIGH at 4000 µg/kg body weight for 21 days during the reproductive season. Exposure to 4000 µg BPA/kg bw/day (BPA HIGH) reduced sperm motility and altered the straight-line velocity (VSL) and linearity (LIN). Effects on steroidogenesis were evident, with testosterone (T) being up-regulated by both treatments and 11-ketotestosterone (11-KT) down-regulated by BPA HIGH. Plasma levels of 17β-estradiol (E2) were not affected. The Gonadosomatic Index (GSI) increased in the BPA HIGH group. Interestingly, the levels of endocannabinoids and endocannabinoid-like compounds were significantly reduced after both treatments. Unpredictably, a few changes were noticed in the expression of genes coding for ECS enzymes, while the receptors were up-regulated depending on the BPA dose. Reproductive markers in testis (leptin receptor (lepr), estrogen receptors (era, erb), progesterone receptors (pr) and the gonadotropin releasing hormone receptor (gnrhr)) were up-regulated. BPA induced the up-regulation of the hepatic genes involved in oogenesis (vitellogenin (vtg) and zona pellucida 1 (zp1)).
Keywords: BPA, endocannabinoids, gonads, gilthead sea bream
1. Introduction
Since 1950, Bisphenol A (BPA, 2,2-bis-(4-hydroxyphenyl)-propane; CAS No. 80-05-7) has been one of the key materials used for the manufacture of polycarbonate plastics and epoxy resins [1]. In a recently published report, the global BPA market is expected to reach around 7348 thousand tons by the end of 2023 [2], making this compound one of the top manufactured and used chemicals. Consequently, BPA has become ubiquitous, being identified in multiple environmental matrices [3,4]. Indeed, BPA has the capacity to migrate from products such as metallic food cans, polycarbonate baby bottles or reusable water bottles, to water or canned food [5]. Although laboratory-based studies have shown evidence of the low ability of BPA to concentrate from the water to the biota and a rapid elimination of this component in fish occurs [3,6,7], it has been detected in tissues of different wildlife species, suggesting that wild organisms are chronically exposed to this contaminant [3,8,9].
The endocrine disrupting activities induced by BPA in teleosts can be summarized in: (1) alterations in the estrogen / androgen ratio [10,11]; (2) alterations in gonadal development and gamete quality [1,10,11,12,13,14]; (3) induction of hepatic vitellogenin production [1,15,16] and (4) epigenetic effects [17,18]. Even though BPA has been reported to exhibit lower estrogenic activity than 17β-estradiol [19], its disrupting potency has also been demonstrated to occur at environmental concentrations in fish [1,11,12,17,20].
On the other hand, the implication of the Endocannabinoid System (ECS) in both mammalian and non-mammalian species reproductive events is also well documented, being a lipid signaling system pivotal for the success of reproductive performance and closely related to the levels of sex-steroids [21]. The ECS is present in the gonads of teleosts, among other tissues, as it has been previously reported in zebrafish Danio rerio [22], goldfish Carassius auratus [23] and gilthead sea bream Sparus aurata [24], with a functional relevance in teleost reproduction [23,25]. Local mediators, namely the endocannabinoids (Anandamide (AEA) and 2-arachidonoylglycerol (2-AG)), their membrane receptors (endocannabinoid receptor type I (CNR1) and type II (CNR2), vanilloid receptor (TRPV1)) and the enzymatic machinery that regulates the levels of the endocannabinoids, compose the ECS, as thoroughly described in several review articles [26,27,28]. Thus, the main goal of the present study was to elucidate whether BPA consumed with food might alter the gonadal ECS and steroidogenesis and, in turn, sperm characteristics during the first reproductive cycle of male gilthead sea bream.
2. Results
2.1. BPA Increased GSI and Altered Sperm Quality
Only the highest concentration of BPA increased the GSI (Figure 1a). No effects were found on the spermiation index (Figure 1b), sperm density (Figure 1c) and survival (Figure 1e). However, sperm motility duration was significantly decreased in the BPA HIGH group with respect to the POST-CTRL group (Figure 1d). Whilst the percentage of motile cells (Figure 2a) and the curvilinear velocity (Figure 2b) were not affected by the treatments with respect to the POST-CTRL group, the straight-line velocity (Figure 2c) and the linearity (Figure 2d) were significantly altered by the BPA HIGH treatment regarding the POST-CTRL animals.
Figure 1.

Mean ± SEM of gilthead sea bream (a) Gonadosomatic Index (GSI); (b) Spermiation index; (c) Sperm density (szoa/mL); (d) Sperm motility duration (sec); (e) Sperm survival (days). Letter superscripts above the means indicate significant differences among treatments (one-way ANOVA, Tukey’s post hoc test, p < 0.05). Absence of letters means no significant differences among groups. Spermiation index was evaluated as S0 = no milt released, S1 = only a drop of milt released after multiple stripping attempts, S2 = milt easily released after the first stripping attempt, S3 = copious amounts of milt flowing with the slightest pressure.
Figure 2.

Computer-Assisted Sperm Analysis (CASA) of gilthead sea bream milt. Mean ± SEM of (a) percentage of motile cells; (b) VCL as µm/sec; (c) VAP as µm/sec; (d) VSL as µm/sec and (e) linearity in percentage (%). Statistically significant differences (two-way ANOVA, Tukey’s post hoc test, p < 0.05) are indicated by different letter superscripts among treatments (in the legend, next to the experimental groups) and sample times (above the columns).
2.2. Gonadal Morphology
The histology of the testes was performed in order to assess whether BPA induced morpho-pathological alterations in the gonads. No differences with respect to the control group (Figure 3a) were observed for each experimental groups (Figure 3b,c). All individuals were in the peak of the spawning season and all cell stages, from spermatogonia to spermatozoa, were present. Regarding the small ovarian part of the testes (Figure 3d), all groups exhibited previtellogenic oocytes.
Figure 3.

Histological images of gilthead sea bream gonads. Testicular tissue from (a) POST-CTRL (b) BPA LOW; (c) BPA HIGH and (d) ovotestis containing an ovarian part with primary oocytes (PO). Testicular part with spermatozoa (sp); sd: spermatids, sg: spermatogonia, sc: spermatocytes. Scale bar: 100 µm.
2.3. Altered Endocannabinoid and Endocannabinoid-Like Mediator Levels in the Testis
Levels of AEA (Figure 4a), and endocannabinoid-like mediators PEA and OEA (Figure 4c,d) were significantly reduced in both treated groups, while the other endocannabinoid, 2-AG, was decreased only in the BPA HIGH individuals (Figure 4b). In agreement with the observed levels of AEA, PEA and OEA, the activity of the enzyme in charge of the hydrolysis of these biochemically related mediators, i.e., fatty acid amide hydrolase (FAAH), was significantly increased in the BPA-exposed fish (Figure 4e).
Figure 4.

(a–d) Testicular Endocannabinoid levels. Data reported as mean ± SEM. Asterisks above each column denote significant differences between POST-CTRL and BPA groups (one-way ANOVA, Dunnett’s multiple comparisons test, * (p < 0.05), ** (p < 0.01), *** (p < 0.001)). AEA expressed as pmol/gr tissue, while 2-AG, PEA and OEA as pmol/mg tissue. (e) FAAH enzymatic activity in testes. Data reported as mean ± SEM of pmol/minutes incubation/mg protein. Asterisks above each column indicate significant differences between POST-CTRL and BPA group (one-way ANOVA, Dunnett’s multiple comparisons test, * (p < 0.05)).
2.4. Sex Steroid Levels
Treatment with BPA did not alter the plasma levels of E2 (Figure 5a), however, the 11-KT ones were significantly reduced in the BPA HIGH group (Figure 5b). Both concentrations of BPA triggered an increase in T and 17, 20β-P plasma levels (Figure 5c,d).
Figure 5.
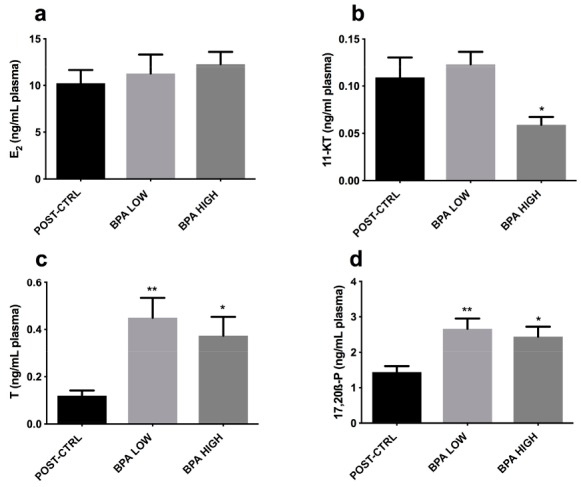
Mean ± SEM plasma levels of (a) Estradiol (E2), (b) 11-Ketosterone (11-KT), (c) Testosterone (T) and (d) 17,20β-P. Asterisks above each column denote significant differences between POST-CTRL and BPA groups (one-way ANOVA, Dunnett’s multiple comparisons test, * (p < 0.05), ** (p < 0.01)).
2.5. Modifications at The Transcriptomic Level in The Male Sea Bream Liver and Testis
qPCR-arrays were performed for genes coding for ECS components and reproductive signals in samples form gilthead sea bream liver and testis. The results obtained are reported in Table 1 for the testis and Table 2 for the liver. Fold-changes of the target genes are represented in heatmaps (Figure 6A,B). Basically, in the testis, for the BPA LOW group, genes coding for the endocannabinoid receptor type I (cnr1), vanilloid receptor (trpv1), leptin receptor (lepr) and estrogen receptor (era) were up-regulated and only the α,β-hydrolase-4 (abdh4), an alternative biosynthetic enzyme for AEA, PEA and OEA, was down-regulated. Regarding the effects of BPA HIGH, the mRNA levels of the endocannabinoid receptor II (crn2), estrogen receptor (erb), progesterone receptor (pr) and gonadotropin-releasing hormone receptor (gnrhr) were increased. In the liver, BPA LOW treatment induced an up-regulation of the genes coding for vitelogenin (vtg) and zona pellucida protein (zp1).
Table 1.
Transcriptional effects of BPA on gilthead sea bream testis. Data are reported as mean ± SEM. All data are normalized to the expression level of cyt-pla2 POST-CTRL fish with an arbitrarily assigned value of 1. Asterisk superscript (*) indicates significant differences between the POST-CTRL group and the treatment (one-way ANOVA, Dunnett’s multiple comparisons test, * (p < 0.05), ** (p < 0.01), **** (p < 0.0001)). Abbreviations: cnr1: endocannabinoid receptor type i, cnr2: endocannabinoid receptor type ii, trpv1: transient receptor potential cation channel subfamily v member 1, ppar: peroxisome proliferator-activated receptor, nape-pld: n-acyl phosphatidylethanolamine phospholipase d, faah: fatty acid amide hydrolase, daglα: diacylglycerol lipase alpha, cox-2: cyclooxigenase-2, abdh: α/β-hydrolase, cyt-pla2: cytosolic phospholipase a2, lepr: leptin receptor, lepa: leptin a, er: estrogen receptor, pr: progesterone receptor, ar: androgen receptor, lhr: luteinizing hormone receptor, fshr: follicle-stimulating hormone receptor, gnrhr: gonadotropin-releasing hormone receptor, 17β-hsd: 17β-hydroxysteroid dehydrogenase, 3β-hsd: 3β-hydroxysteroid dehydrogenase.
| Gene | POST-CTRL | BPA LOW | BPA HIGH |
|---|---|---|---|
| cnr1 | 0.38 ± 0.07 | 0.64 ± 0.10 * | 0.60 ± 0.06 |
| cnr2 | 0.35 ± 0.04 | 0.24 ± 0.04 | 0.63 ± 0.08 ** |
| ppar α | 3.33 ± 0.34 | 2.43 ± 0.22 | 2.49 ± 0.24 |
| ppar β | 6.41 ± 0.63 | 6.82 ± 0.80 | 6.86 ± 0.72 |
| ppar γ | 12.68 ± 1.69 | 16.14 ± 2.13 | 12.63 ± 1.52 |
| trpv1 | 0.24 ± 0.03 | 0.37 ± 0.03 * | 0.30 ± 0.03 |
| faah | 4.87 ± 0.49 | 6.81 ± 0.67 | 4.81 ± 0.62 |
| nape-pld | 22.71 ± 1.55 | 25.50 ± 2.50 | 19.29 ± 1.94 |
| abdh4 | 3.69 ± 0.35 | 2.52 ± 0.35* | 3.00 ± 0.19 |
| cyt-pla2 | 1.00 ± 0.12 | 0.94 ± 0.12 | 1.22 ± 0.15 |
| cox2 | 1.43 ± 0.18 | 1.13 ± 0.12 | 0.98 ± 0.12 |
| daglα | 4.39 ± 0.46 | 5.96 ± 0.64 | 4,17 ± 0.49 |
| abdh 6a | 4.42 ± 0.34 | 3.56 ± 0.35 | 4.00 ± 0.36 |
| abdh 12b | 2.56 ± 0.29 | 2.65 ± 0.33 | 2.54 ± 0.34 |
| lepr | 15.88 ± 2.80 | 51.49 ± 5.16 **** | 24.75 ± 4.78 |
| lepa | 0.30 ± 0.03 | 0.25 ± 0.03 | 0.37 ± 0.04 |
| era | 3.04 ± 0.30 | 4.23 ± 0.34 * | 4.07 ± 0.33 |
| erb | 11.89 ± 0.67 | 11.68 ± 1.13 | 15.33 ± 1.02 * |
| pr | 4.36 ± 0.60 | 5.67 ± 0.66 | 7.57 ± 0.65 ** |
| ar | 8.59 ± 0.78 | 7.28 ± 0.89 | 7.72 ± 0.67 |
| lhr | 0.56 ± 0.06 | 0.50 ± 0.09 | 0.61 ± 0.04 |
| fshr | 0.46 ± 0.10 | 0.63 ± 0.07 | 0.56 ± 0.07 |
| gnrhr | 5.36 ± 0.81 | 5.11 ± 0.49 | 8.44 ± 0.96 * |
| 17b-hsd | 5.20 ± 0.50 | 4.75 ± 0.53 | 4.85 ± 0.45 |
| 3b-hsd | 5.91 ± 0.59 | 6.80 ± 0.62 | 5.63 ± 0.76 |
Table 2.
Transcriptional effects of BPA on the liver of gilthead sea bream. Data are reported as mean ± SEM. All data are normalized to the expression level of zp1 of POST-CTRL fish with an arbitrarily assigned value of 1. Asterisk superscript (*) indicates significant differences between the POST-CTRL group and the treatment (one-way ANOVA, Dunnett’s multiple comparisons test, * (p < 0.05), ** (p < 0.01)). Abbreviations: vtga: vitellogenin a; zp1: zona pellucida—like domain—containing protein 1; zp3: zona pellucida sperm-binding protein 3.
| Gene | POST-CTRL | BPA LOW | BPA HIGH |
|---|---|---|---|
| vtga | 0.76 ± 0.43 | 1.71 ± 0.43 ** | 0.96 ± 0.33 |
| zp1 | 1.00 ± 0.35 | 1.91 ± 0.48 * | 1.00 ± 0.26 |
| zp3 | 116.32 ± 39.82 | 58.51 ± 37.54 | 73.11 ± 35.97 |
Figure 6.

Gene expression heat maps in the testes (A) and the liver (B). Scales of colors represent fold-change values in relation to the POST-CTRL group (green, down-regulation; red, up-regulation).
3. Discussion
The ubiquitous presence of BPA in a large number of items, including canned food, poses a question around the safety of this compound, which is reported to behave as an estrogen-like chemical. In the present study, BPA was administered by food to adult male gilthead sea bream. Gilthead sea bream represents an interesting model for the study of EDCs due to its sex reversion [29]. For this reason, in the present study, gilthead sea bream were sampled during their first reproductive cycle, when all individuals still function as males. However, unraveling the activity induced by BPA is generally puzzling, due to the tissue-specific disruptive activities, species-specific sensibilities, and dose-dependent effects of this compound [11,30]. Indeed, BPA can differentially affect teleost male GSI depending on the dose administered and duration of treatment, making it difficult to draw conclusions regarding its effects on GSI, which in turn, would consequently not be considered a good indicator for BPA effects [11,31]. However, increased GSI was reported with different BPA concentrations administered for 43 days to male fathead minnows Pimephales promelas [14], and with 5 µg BPA/L to male rare minnows Gobiocypris rarus [32]. Controversially, chronic treatments with environmental relevant concentrations of BPA did not alter GSI in male goldfish [11,12], in brown trout Salmo trutta male at 50 µg/L [15], in male medaka Oryzias latipes at higher concentrations [33] and in rare minnows treated with 15 and 225 µg/L [34]. Nevertheless, as some previtellogenic oocytes were observed, we hypothesized that the GSI rise may have been caused by the increasing size of the ovarian part, particularly since all groups showed the same spermiation index and similar histological results. Indeed, as previously reported [14], EDC effects on adult teleosts not only depend on the type of EDC, but also on the duration and timing of the exposure. Furthermore, we need to take into account that when BPA is orally administered and not diluted in water, the kinetics could be different due to low BPA absorption or fast metabolism [35].
BPA did not induce any meaningful change in E2 levels in our experiments. This finding was in agreement with the unaltered levels of this hormone found in male goldfish treated with BPA [12]. Additionally, decreased levels of E2 have been reported after treatment with environmental concentrations of BPA in male carps [10] and a reduction of E2 metabolism was described in immature lake trout Salvelinus namaycush [36]. Though E2 was unaffected, mRNA levels of vtg and era were increased in the BPA LOW group; indeed, in male goldfish, 0.2 µg BPA /L increased the expression of erb1 mRNA in the liver but not vtg mRNA; however, a higher dose of BPA stimulated both era and erb, up-regulating vtg [11]. On the other hand, erb was only augmented by the highest dose of BPA tested, underlying a dose-related action of the compound on ERα and ERβ expression [37]. It was unexpected to find such levels of T, which was probably due to the modulation of aromatase activity by BPA [11,38]. In this sense, higher levels of T (although not statistically significant) have been found in male carps treated with 1 µg BPA/L [10], and increased levels of T in male rare minnows treated with 225 µg/L [38]. Instead, the levels of 11-KT, the active androgen of teleost, were reduced in the present study, similarly to male goldfish exposed to 4.5 and 11 µg BPA/L for 20 days [12].
In spite of these hormonal changes, no morphological alteration was observed in the testis, even if these steroids were supposed to decrease during spermiation [39], suggesting time-dependent levels, as previously reported in gilthead sea bream treated with 5 µg EE2/g food [40]. The study of [40] evidenced an increase in plasma E2 and T levels after seven days, followed by a decrease of T and 11-KT levels after 28 days. Furthermore, taking into account that injections of exogenous E2 switch functional testis to the post-spawning stage, [41] suggested that higher levels of this sex hormone were needed to orchestrate testicular regression.
In the case of 17,20β-P, previous studies evidenced that this hormone stimulated milt production and sperm motility in male trout [42] and triggers oocyte maturation in female ovaries. However, no morphological difference was observed in the BPA groups. In spite of the clearly changed levels of sex steroids, it seems that that alteration and/or the exposure time was likely to be insufficient to induce morphological changes in the testis, in agreement with studies showing pathological testicular alterations after longer BPA exposure times [14,43].
Comparing the steroidogenic effects of BPA with that induced by DiNP in our previous ones [44], similar results were found regarding the lower levels of 11-KT associated with higher levels of T. Unlike DiNP, BPA did not modify the plasma E2 levels. However, while DiNP reduced the levels of 17,20β-P, BPA increased them. The differences found between these two contaminants showed that both contaminants were able to alter steroidogenesis but in different ways, suggesting a different way of controlling steroidogenesis.
Focusing on sperm, previous publications have indicated that different dosages of BPA decrease sperm velocity in teleosts [12,13,34,45], as in the BPA HIGH group. On the other hand, different trends have been observed for sperm concentration: while in the brown trout it was reduced [13]; [34] reported unaltered sperm concentrations for rare minnows exposed to 15 and 225 µg BPA/L, which was in agreement with our results. As it was previously reported [44], differences in sperm parameters were found between PRE-CTRL and POST-CTRL groups, likely associated with the different sampling time. PRE-CTRL group was sampled during January, and POST-CTRL in late February, during the peak of the spawning season. This could explain the significant differences found in sperm quality between these two groups [46].
The importance of the ECS in male reproduction was first documented in sea urchins [47]. Thereafter, several studies evidenced the role of this system in both male and females of several species reproduction [25,48,49]. Indeed, the ECS interferes with the brain-pituitary-gonadal axis, affecting Sertoli and Leydig cells, germ cell differentiation, sperm motility and the acrosome reaction [27]. Additionally, a cross-talk among the ECS and sex steroids exists [21,50]. In mouse Sertoli cells, E2 via ER binds the ERE (estrogen response element) sites of the FAAH promoter, thereby stimulating faah expression and FAAH activity, and subsequently decreasing AEA levels and protecting the testis from AEA-induced apoptosis [50,51]. In the present work, treatment with BPA did not increase E2 levels or faah expression, but enhanced FAAH enzymatic activity. Consequently, reduced levels of AEA and AEA-related compounds (PEA and OEA) were found. The levels of 2-AG were also reduced, in agreement with previous data indicating that this endocannabinoid may act as an alternative FAAH substrate [52]. In comparison to the results of Grimaldi and Rossi mentioned above, who reported increased FAAH activity concomitantly with faah mRNA expression in Sertoli cells after E2 treatment; BPA seemed to execute an estrogen-like effect on FAAH activity rather than faah expression in gilthead sea bream. Therefore, two hypotheses were formulated: 1) BPA may exert a different effect at the transcriptional and translational/post-translational levels, or 2) transcription and translation can be uncoupled in testes [53,54]. The effects of BPA on ECS were similar to that observed with DiNP [44] suggesting a similar ECS control by these two contaminants.
Finally, a correct endocannabinoid “tone” is essential for the successful progression of spermatogenesis. In fact, lower levels of AEA and 2-AG have been found in the sperm of infertile men [55], suggesting that further fertilizing effects may be observed in gilthead sea bream treated with BPA, jointly with the changed sperm parameters mentioned before. The mRNA expression levels of some endocannabinoid targets and related mediators (cnr1, cnr2 and trpv1) were slightly increased by one of the two doses of BPA, possibly as a compensatory response to the reduction of AEA, 2-AG, PEA and OEA. These receptors have all been implicated in various aspects of the physiopathology of sperm cells and oocytes [55,56,57,58,59]. The role of these receptors in the potentially ECS-mediated effects of BPA constitutes an interesting line of research that merits further consideration in future studies.
In summary (Figure 7), the effects of BPA—one of the most worldwide manufactured EDCs—on the reproductive physiology of different species has been largely studied. However, the effects of BPA are still difficult to accurately predict due to their variability depending on the dose used, the duration of exposure and the developmental stage of the model used. The present study has attempted to shed some more light on the alterations induced by the temporary tolerable daily intake set by the European Food Safety Authority (EFSA) in 2015 for BPA (4 µg BPA/kg bw/day). Even if lack of a clear effect was observed concerning histological changes, spermiation index and sperm quality, significant changes were observed at the hormonal and endocannabinoid levels.
Figure 7.

Summarized results. Abbreviations: BPAH (BPA HIGH group), FAAH (Fatty acid amide hydrolase), cnr (endocannabinoid receptors), T (testosterone), 11-KT (11-Ketotestosterone). Red arrows: increase, green arrows: decrease.
4. Materials and Methods
4.1. Fish Maintenance, Food Preparation and Animal Treatment
Two-year-old gilthead sea bream (461.09 ± 16.93 gr; 301.7 ± 10.7 mm) were maintained at the AQUALABS facilities of the Institute of Marine Biology, Biotechnology and Aquaculture of the Hellenic Centre for Marine Research (HCMR), Iraklion, Crete, Greece, following the conditions reported previously [44,60]. Gilthead sea bream are protandrous hermaphrodites, and at two years of age all individuals function as males with developed testis and previtellogenic ovaries.
After 3 months of acclimation and before the beginning of the experimentation, and following previous publications [44,60], the sperm quality parameters were assessed in January to obtain the pre-treatment data (PRE-CTRL), as explained below. Then, the fish were divided into three groups of 10 specimens (in duplicates) and left to acclimatize in the experimental tanks (2 m3). After the acclimation, the fish were fed with due consideration for the total amount of pellet, which was administered equal to 0.7% of the body weight (bw). The treatment was conducted for 21 days following the Organization for Economic Co-operation and Development (OECD) guidelines # 60, 78, 109 [31,61,62] via food intake, being the BPA (Sigma-Aldrich, Milano, Italy) introduced into commercial pellets (IRIDA S.A. Greece) as explained [35,44,60]. The doses administered were adjusted to give the nominal doses (4 and 4000 µg BPA/kg bw/day) according to the weight of fish shared in several meals along the day to ensure the fish ingested the proportional BPA amount.
The tanks (in duplicate) were organized as: Control group (POST-CTRL) fed with the prepared food with the vehicle (ethanol, EtOH); BPA LOW food enriched with 4 µg BPA/kg bw/day); BPA HIGH with 4000 µg BPA/kg bw/day. The lowest concentration of BPA was based on the Tolerable Daily Intake (TDI) for humans ruled by the European Food Safety Authority (EFSA), which was set from January 2015 to 4 µg of BPA/kg bw/day. 4000 µg BPA/kg bw/day was chosen as an effective concentration (EC).
The PRE-CTRL and POST-CTRL individuals were the same of those previously reported [44] since both experiments were running in parallel at the same time.
The experimentation was performed under the same conditions (tanks, temperature and photoperiod) already described elsewhere [44,60]. The experimental protocol was approved by the Greek National Veterinary Agency with the Protocol Number #255361 (29 November 2017) of the experimental facility EL91-BIOexp-04.
After 21 days (February), five fish were randomly chosen per tank and were anesthetized by clove oil (30 mg/L) following the indications of [63] and sampled. Blood was collected from the ventral vasculature behind the anal fin and stored at –20 °C for ELISA assay. Sperm was collected by gentle abdominal massage and stored at 4 °C until its evaluation. Gonads were weighed for Gonadosomatic Index (GSI) calculation ((Gonads weight (g)/Fish weight (g)) × 100). Pieces of the testes were collected and stored at –80 °C for endocannabinoid measurements and FAAH enzymatic assay. Since gilthead sea bream is a hermaphrodite species, pieces of ovo-testis were embedded in a solution of Formaldehyde:glutaraldehyde (4:1) for histological studies. For gene expression analysis, small pieces of liver and testes were stored within RNAlater (Ambion Inc., Austin, TX, USA) at 4 °C until processing.
4.2. Sperm Quality Evaluation
The evaluation of sperm quality was performed in the PRE-CTRL, POST-CTRL and BPA groups as previously described [44,64]. The parameters studied were the sperm density (szoa·mL−1), sperm survival (days), sperm motility duration (s), percentage of motile cells, Curvilinear Velocity (VCL, μm·s−1), Straight Line Velocity (VSL, μm·s−1) and Linearity (LIN, %). The analyzed spermatozoa (63–228) were within the reported range. All sperm parameters were evaluated before (PRE-CTRL) and after treatments (POST-CTRL and BPA groups).
4.3. Testes Histology
Histological procedures were performed as previously described [33] in the POST-CTRL and BPA groups. Briefly, pieces of ovo-testes were embedded in methacrylate resin (Technovit 7100®, Heraeus Kulzer, Germany). Then, they were sectioned with a microtome (Leica RM2245) and stained with Methylene Blue (Sigma-Aldrich, Munich, Germany)/Azure II (Sigma-Aldrich)/Basic fuchsine (Polysciences, Warrington, PA, USA) [65]. Being a protandrous hermaphrodite species, gilthead sea bream testes always contain a small quiescent ovarian section, which was also examined.
4.4. Enzyme-Linked ImmunoSorbent Assay (ELISA)
Plasma (200 µL) from POST-CTRL and BPA groups were extracted with diethyl ether following [44] procedures for quantification of testosterone (Τ), 11-Ketotestosterone (11-ΚΤ), 17β-estradiol (E2) and 17,20β -dihydroxy-4prengnen-3-one (17,20β-P or maturation inducing steroid, MIS), by readily established protocols [66,67].
4.5. RNA Extraction, cDNA Synthesis and Real-Time PCR
RNA extraction and cDNA synthesis from the liver and testis from POST-CTRL and BPA groups were conducted as previously described [60] with RNAzol solution (Sigma-Aldrich, Italy). For cDNA synthesis, the High-Capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA, USA) was used with 500-ng total RNA in a final volume of 100 µL.
Real-time quantitative PCR was carried out as previously described [60] with an Eppendorf Mastercycler Ep Realplex real-time PCR system (Eppendorf, Wesseling-Berzdorf, Germany), using two 96-well PCR-array layouts designed for the simultaneous profiling of a panel of 25 genes for gonad samples and 3 genes for liver samples, selected as: i) endocannabinoid system markers: cb1, cb2, pparα, pparβ, pparγ, faah, nape-pld, abdh4, cyt-pla2, cox2, dalgα, abdh6a, abdh12a, trpv1 ii) reproductive markers: era, erb, pr, ar, lhr, fshr, gnrhr, 17β-hsd, 3β-hsd, lepr, lep, vtg, zp1, zp3. PCR-array primers (Supplementary Table S1) were designed to obtain amplicons of 50–150 bp in length. A housekeeping gene (β-actin) and controls for general PCR performance were included on each array. The genes selected as well as the PCR results from POST-CTRL were the same from the previous published study from our group [44] to possibly compare the effects of the di-isononyl phthalate (DiNP) and BPA.
4.6. Concentration of Endogenous Cannabinoids (Anandamide (AEA), 2-Arachidonoylglycerol (2-AG)), Endocannabinoid-Like Mediators (Palmitoylethanolamide (PEA), Oleoylethanolamide (OEA)) and Fatty Acid Amide Hydrolase (FAAH) Enzymatic Activity in the Testis
The testes were processed and analyzed for levels of endocannabinoids and FAAH activity as previously described [60].
4.7. Statistical Analysis
Statistical analysis of gene expression, FAAH activity, endocannabinoid levels, ELISA and GSI were performed using one-way analysis of variance (ANOVA) followed by a Tukey’s or Dunnett’s multiple comparison test, while the CASA data was analyzed by two-way ANOVA. Results are reported as mean ± SEM (Standard Error of the Mean). The data fulfilled the condition for applying a parametric test; given the log-normalization to homogenize the variance when needed. Data in percentages were transformed to arc sinus to apply the statistical test. All statistical procedures were run using GraphPad Prism 6. Superscript letters specified statistical differences among treatments (p < 0.05). A superscript asterisk (*) indicated statistical significance with respect to the POST-CTRL group (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001).
Acknowledgments
This work received funding from Progetti di Rilevante Interesse Nazionale (PRIN) 2010–2011 (prot 2010W87LBJ) to OC and from the European Union’s Horizon 2020 research and innovation program under the TNA programme (project ID AE030014) within AQUAEXCEL2020 project (652831) to IFP and OC for accessing to IATS-CSIC facilities. This output reflects only the author’s view and the European Union cannot be held responsible for any use that may be made of the information contained herein. The authors would like to thank the assistance of Marco Allarà for the enzymatic activity assay, Roberta Verde for assistance with the LC-MS functioning and M. Ángeles González for technical assistance in the PCR assays.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/20/20/5003/s1.
Author Contributions
Conceptualization, I.F.-P. and O.C.; methodology, I.F.-P., I.F., C.CM., F.P., J.C.-G., J.P.-S. and O.C.; formal analysis, I.F.-P., I.F., C.CM., F.P., J.C.-G., O.C.; investigation, I.F.P., I.F., C.CM., F.P., J.C.-G. and O.C.; resources, C.CM., V.D.M., J.C.-G., J.P.-S. and O.C.; data curation, I.F.P., I.F., C.CM., F.P., J.C.-G. and O.C.; writing—original draft preparation, I.F.-P., I.F., J.C.-G.; writing—review and editing, C.CM., F.P., V.D.M., F.M., J.P.-S. and O.C.; visualization, I.F.-P.; supervision, C.CM., V.D.M., J.C.-G., J.P.-S. and O.C.; project administration, J.P.-S. and O.C.; funding acquisition, I.F.-P., J.C.-G., J.P.-S. and O.C.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1.Crain D.A., Eriksen M., Iguchi T., Jobling S., Laufer H., LeBlanc G.A., Guillette L.J. An ecological assessment of bisphenol-A: Evidence from comparative biology. Reprod. Toxicol. 2007;24:225–239. doi: 10.1016/j.reprotox.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 2.Williams & Marshall Strategy The Global Bisphenol A Market. [(accessed on 12 March 2019)];2019 Available online: https://www.researchandmarkets.com/research/hl86rz/global_bisphenol?w=5.
- 3.Corrales J., Kristofco L.A., Steele W.B., Yates B.S., Breed C.S., Williams E.S., Brooks B.W. Global Assessment of Bisphenol A in the Environment. Dose-Response. 2015;13:155932581559830. doi: 10.1177/1559325815598308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fromme H., Küchler T., Otto T., Pilz K., Müller J., Wenzel A. Occurrence of phthalates and bisphenol A and F in the environment. Water Res. 2002;36:1429–1438. doi: 10.1016/S0043-1354(01)00367-0. [DOI] [PubMed] [Google Scholar]
- 5.Vandenberg L.N., Maffini M.V., Sonnenschein C., Rubin B.S., Soto A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009;30:75–95. doi: 10.1210/er.2008-0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Staples C.A., Dome P.B., Klecka G.M., Oblock S.T., Harris L.R. A review of the environmental fate, effects, and exposures of bisphenol A. Chemosphere. 1998;36:2149–2173. doi: 10.1016/S0045-6535(97)10133-3. [DOI] [PubMed] [Google Scholar]
- 7.Lindholst C., Pedersen S.N., Bjerregaard P. Uptake, metabolism and excretion of bisphenol A in the rainbow trout (Oncorhynchus mykiss) Aquat. Toxicol. 2001;55:75–84. doi: 10.1016/S0166-445X(01)00157-6. [DOI] [PubMed] [Google Scholar]
- 8.Mita L., Bianco M., Viggiano E., Zollo F., Bencivenga U., Sica V., Monaco G., Portaccio M., Diano N., Colonna A., et al. Bisphenol A content in fish caught in two different sites of the Tyrrhenian Sea (Italy) Chemosphere. 2011;82:405–410. doi: 10.1016/j.chemosphere.2010.09.071. [DOI] [PubMed] [Google Scholar]
- 9.Lee C.-C., Jiang L.-Y., Kuo Y.-L., Chen C.-Y., Hsieh C.-Y., Hung C.-F., Tien C.-J. Characteristics of nonylphenol and bisphenol A accumulation by fish and implications for ecological and human health. Sci. Total Environ. 2015;502:417–425. doi: 10.1016/j.scitotenv.2014.09.042. [DOI] [PubMed] [Google Scholar]
- 10.Mandich A., Bottero S., Benfenati E., Cevasco A., Erratico C., Maggioni S., Massari A., Pedemonte F., Viganò L. In vivo exposure of carp to graded concentrations of bisphenol A. Gen. Comp. Endocrinol. 2007;153:15–24. doi: 10.1016/j.ygcen.2007.01.004. [DOI] [PubMed] [Google Scholar]
- 11.Hatef A., Zare A., Alavi S.M.H., Habibi H.R., Linhart O. Modulations in androgen and estrogen mediating genes and testicular response in male goldfish exposed to bisphenol A. Environ. Toxicol. Chem. 2012;31:2069–2077. doi: 10.1002/etc.1919. [DOI] [PubMed] [Google Scholar]
- 12.Hatef A., Alavi S.M.H., Abdulfatah A., Fontaine P., Rodina M., Linhart O. Adverse effects of bisphenol A on reproductive physiology in male goldfish at environmentally relevant concentrations. Ecotoxicol. Environ. Saf. 2012;76:56–62. doi: 10.1016/j.ecoenv.2011.09.021. [DOI] [PubMed] [Google Scholar]
- 13.Lahnsteiner F., Berger B., Kletzl M., Weismann T. Effect of bisphenol A on maturation and quality of semen and eggs in the brown trout, Salmo trutta f. fario. Aquat. Toxicol. 2005;75:213–224. doi: 10.1016/j.aquatox.2005.08.004. [DOI] [PubMed] [Google Scholar]
- 14.Sohoni P., Tyler C.R., Hurd K., Caunter J., Hetheridge M., Williams T., Woods C., Evans M., Toy R., Gargas M., et al. Reproductive Effects of Long-Term Exposure to Bisphenol A in the Fathead Minnow (Pimephales promelas) Environ. Sci. Technol. 2001;35:2917–2925. doi: 10.1021/es000198n. [DOI] [PubMed] [Google Scholar]
- 15.Bjerregaard L.B., Madsen A.H., Korsgaard B., Bjerregaard P. Gonad histology and vitellogenin concentrations in brown trout (Salmo trutta) from Danish streams impacted by sewage effluent. Ecotoxicology. 2006;15:315–327. doi: 10.1007/s10646-006-0061-9. [DOI] [PubMed] [Google Scholar]
- 16.Maradonna F., Nozzi V., Dalla Valle L., Traversi I., Gioacchini G., Benato F., Colletti E., Gallo P., Di Marco Pisciottano I., Mita D.G., et al. A developmental hepatotoxicity study of dietary bisphenol A in Sparus aurata juveniles. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014;166:1–13. doi: 10.1016/j.cbpc.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 17.Santangeli S., Maradonna F., Gioacchini G., Cobellis G., Piccinetti C.C., Dalla Valle L., Carnevali O. BPA-Induced deregulation of epigenetic patterns: Effects on female zebrafish reproduction. Sci. Rep. 2016;6 doi: 10.1038/srep21982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Santangeli S., Maradonna F., Olivotto I., Piccinetti C.C., Gioacchini G., Carnevali O. Effects of BPA on female reproductive function: The involvement of epigenetic mechanism. Gen. Comp. Endocrinol. 2017;245 doi: 10.1016/j.ygcen.2016.08.010. [DOI] [PubMed] [Google Scholar]
- 19.Witorsch R.J. Endocrine Disruptors: Can Biological Effects and Environmental Risks Be Predicted? Regul. Toxicol. Pharmacol. 2002;36:118–130. doi: 10.1006/rtph.2002.1564. [DOI] [PubMed] [Google Scholar]
- 20.Oehlmann J., Schulte-Oehlmann U., Kloas W., Jagnytsch O., Lutz I., Kusk K.O., Wollenberger L., Santos E.M., Paull G.C., Van Look K.J.W., et al. A critical analysis of the biological impacts of plasticizers on wildlife. Philos. Trans. R. Soc. B Biol. Sci. 2009;364:2047–2062. doi: 10.1098/rstb.2008.0242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gorzalka B.B., Dang S.S. Minireview: Endocannabinoids and Gonadal Hormones: Bidirectional Interactions in Physiology and Behavior. Endocrinology. 2012;153:1016–1024. doi: 10.1210/en.2011-1643. [DOI] [PubMed] [Google Scholar]
- 22.Forner-Piquer I., Santangeli S., Maradonna F., Rabbito A., Piscitelli F., Habibi H.R., Di V. Disruption of the gonadal endocannabinoid system in zebra fish exposed to diisononyl phthalate. Environ. Pollut. 2018;241:1–8. doi: 10.1016/j.envpol.2018.05.007. [DOI] [PubMed] [Google Scholar]
- 23.Cottone E., Pomatto V., Bovolin P. Role of the endocannabinoid system in the central regulation of nonmammalian vertebrate reproduction. Int. J. Endocrinol. 2013;2013:941237. doi: 10.1155/2013/941237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ruggeri B., Soverchia L., Mosconi G., Franzoni M.F., Cottone E., Polzonetti-Magni A.M. Changes of gonadal CB1 cannabinoid receptor mRNA in the gilthead seabream, Sparus aurata, during sex reversal. Gen. Comp. Endocrinol. 2007;150:263–269. doi: 10.1016/j.ygcen.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 25.Cottone E., Guastalla A., Mackie K., Franzoni M.F. Endocannabinoids affect the reproductive functions in teleosts and amphibians. Mol. Cell. Endocrinol. 2008;286:S41–S45. doi: 10.1016/j.mce.2008.01.025. [DOI] [PubMed] [Google Scholar]
- 26.Battista N., Meccariello R., Cobellis G., Fasano S., Di Tommaso M., Pirazzi V., Konje J.C., Pierantoni R., Maccarrone M. The role of endocannabinoids in gonadal function and fertility along the evolutionary axis. Mol. Cell. Endocrinol. 2012;355:1–14. doi: 10.1016/j.mce.2012.01.014. [DOI] [PubMed] [Google Scholar]
- 27.Grimaldi P., Di Giacomo D., Geremia R. The endocannabinoid system and spermatogenesis. Front. Endocrinol. (Lausanne). 2013;4:192. doi: 10.3389/fendo.2013.00192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Maccarrone M., Bab I., Bíró T., Cabral G.A., Dey S.K., Di Marzo V., Konje J.C., Kunos G., Mechoulam R., Pacher P., et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015;36:277–296. doi: 10.1016/j.tips.2015.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zohar Y., Abraham M., Gordin H. The gonadal cycle of the captivity-reared hermaphroditic teleost Sparus aurata (L.) during the first two years of life. Ann. Biol. Anim. Bioch. Biophys. 1978;18:877–882. doi: 10.1051/rnd:19780519. [DOI] [Google Scholar]
- 30.Welshons W.V., Nagel S.C., vom Saal F.S. Large effects from small exposures. III. Endocrine mechanisms mediating effects of bisphenol A at levels of human exposure. Endocrinology. 2006;147:s56–s69. doi: 10.1210/en.2005-1159. [DOI] [PubMed] [Google Scholar]
- 31.Organisation for Economic Co-Operation and Development Number 60. Report of the initial work towards the validation of the 21-day fish screening assay for the detection of endocrine active substances (Phase 1A) [(accessed on 5 June 2019)];2006 Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2006)27&doclanguage=en.
- 32.Zhang Y., Yuan C., Hu G., Li M., Zheng Y., Gao J., Yang Y., Zhou Y., Wang Z. Characterization of four nr5a genes and gene expression profiling for testicular steroidogenesis-related genes and their regulatory factors in response to bisphenol A in rare minnow Gobiocypris rarus. Gen. Comp. Endocrinol. 2013;194:31–44. doi: 10.1016/j.ygcen.2013.08.014. [DOI] [PubMed] [Google Scholar]
- 33.Kang I.J., Yokota H., Oshima Y., Tsuruda Y., Oe T., Imada N., Tadokoro H., Honjo T. Effects of bisphenol a on the reproduction of Japanese medaka (Oryzias latipes) Environ. Toxicol. Chem. 2002;21:2394–2400. doi: 10.1002/etc.5620211119. [DOI] [PubMed] [Google Scholar]
- 34.Zhang Y., Guan Y., Zhang T., Yuan C., Liu Y., Wang Z. Adult exposure to bisphenol A in rare minnow Gobiocypris rarus reduces sperm quality with disruption of testicular aquaporins. Chemosphere. 2018;193:365–375. doi: 10.1016/j.chemosphere.2017.11.034. [DOI] [PubMed] [Google Scholar]
- 35.Bjerregaard P., Andersen S.B., Pedersen K.L., Pedersen S.N., Korsgaard B. Orally administered bisphenol a in rainbow trout (Oncorhynchus mykiss): EStrogenicity, metabolism, and retention. Environ. Toxicol. Chem. 2007;26:1910. doi: 10.1897/06-645R.1. [DOI] [PubMed] [Google Scholar]
- 36.Jurgella G.F., Marwah A., Malison J.A., Peterson R., Barry T.P. Effects of xenobiotics and steroids on renal and hepatic estrogen metabolism in lake trout. Gen. Comp. Endocrinol. 2006;148:273–281. doi: 10.1016/j.ygcen.2006.03.011. [DOI] [PubMed] [Google Scholar]
- 37.Hiroi H., Tsutsumi O., Momoeda M., Takai Y., Osuga Y., Taketani Y. Differential interactions of bisphenol A and 17 beta-estradiol with estrogen receptor alpha (ERalpha) and ERbeta. Endocr. J. 1999;46:773–778. doi: 10.1507/endocrj.46.773. [DOI] [PubMed] [Google Scholar]
- 38.Zhang Y., Yuan C., Gao J., Liu Y., Wang Z. Testicular transcript responses in rare minnow Gobiocypris rarus following different concentrations bisphenol A exposure. Chemosphere. 2016;156:357–366. doi: 10.1016/j.chemosphere.2016.05.006. [DOI] [PubMed] [Google Scholar]
- 39.Schulz R.W., de França L.R., Lareyre J.-J., LeGac F., Chiarini-Garcia H., Nobrega R.H., Miura T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010;165:390–411. doi: 10.1016/j.ygcen.2009.02.013. [DOI] [PubMed] [Google Scholar]
- 40.Cabas I., Chaves-Pozo E., García-Alcázar A., Meseguer J., Mulero V., García-Ayala A. The effect of 17α-ethynylestradiol on steroidogenesis and gonadal cytokine gene expression is related to the reproductive stage in marine hermaphrodite fish. Mar. Drugs. 2013;11:4973–4992. doi: 10.3390/md11124973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mendez-Sanchez N., Zamora-Valdes D., Pichardo-Bahena R., Barredo-Prieto B., Ponciano-Rodriguez G., Bermejo-Martínez L., Chavez-Tapia N.C., Baptista-González H.A., Uribe M. Endocannabinoid receptor CB2 in nonalcoholic fatty liver disease. Liver Int. 2007;27:215–219. doi: 10.1111/j.1478-3231.2006.01401.x. [DOI] [PubMed] [Google Scholar]
- 42.Scott A.P., Sumpter J.P., Stacey N. The role of the maturation-inducing steroid, 17,20β-dihydroxypregn-4-en-3-one, in male fishes: A review. J. Fish Biol. 2010;76:183–224. doi: 10.1111/j.1095-8649.2009.02483.x. [DOI] [PubMed] [Google Scholar]
- 43.Mihaich E., Rhodes J., Wolf J., van der Hoeven N., Dietrich D., Hall A.T., Caspers N., Ortego L., Staples C., Dimond S., et al. Adult fathead minnow, Pimephales promelas, partial life-cycle reproductive and gonadal histopathology study with bisphenol A. Environ. Toxicol. Chem. 2012;31:2525–2535. doi: 10.1002/etc.1976. [DOI] [PubMed] [Google Scholar]
- 44.Forner-Piquer I., Mylonas C.C., Fakriadis I., Papadaki M., Piscitelli F., Di Marzo V., Calduch-Giner J., Pérez-Sánchez J., Carnevali O. Effects of diisononyl phthalate (DiNP) on the endocannabinoid and reproductive systems of male gilthead sea bream (Sparus aurata) during the spawning season. Arch. Toxicol. 2019 doi: 10.1007/s00204-018-2378-6. [DOI] [PubMed] [Google Scholar]
- 45.Hatef A., Alavi S.M.H., Linhartova Z., Rodina M., Policar T., Linhart O. In vitro effects of Bisphenol A on sperm motility characteristics in Perca fluviatilis L. (Percidae; Teleostei) J. Appl. Ichthyol. 2010;26:696–701. doi: 10.1111/j.1439-0426.2010.01543.x. [DOI] [Google Scholar]
- 46.Rouxel C., Suquet M., Cosson J., Severe A., Quemener L., Fauvel C. Changes in Atlantic cod (Gadus morhua L.) sperm quality during the spawning season. Aquac. Res. 2008;39:434–440. doi: 10.1111/j.1365-2109.2007.01852.x. [DOI] [Google Scholar]
- 47.Schuel H., Schuel R., Zimmerman A.M., Zimmerman S. Cannabinoids reduce fertility of sea urchin sperm. Biochem. Cell Biol. 1987;65:130–136. doi: 10.1139/o87-018. [DOI] [PubMed] [Google Scholar]
- 48.Rapino C., Battista N., Bari M., Maccarrone M. Endocannabinoids as biomarkers of human reproduction. Hum. Reprod. Update. 2014;20:501–516. doi: 10.1093/humupd/dmu004. [DOI] [PubMed] [Google Scholar]
- 49.Bovolin P., Cottone E., Pomatto V., Fasano S., Pierantoni R., Cobellis G., Meccariello R. Endocannabinoids are involved in male vertebrate reproduction: Regulatory mechanisms at central and gonadal level. Front. Endocrinol. (Lausanne) 2014;5:54. doi: 10.3389/fendo.2014.00054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Grimaldi P., Pucci M., Di Siena S., Di Giacomo D., Pirazzi V., Geremia R., Maccarrone M. The faah gene is the first direct target of estrogen in the testis: Role of histone demethylase LSD1. Cell. Mol. Life Sci. 2012;69:4177–4190. doi: 10.1007/s00018-012-1074-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rossi G., Gasperi V., Paro R., Barsacchi D., Cecconi S., Maccarrone M. Follicle-stimulating hormone activates fatty acid amide hydrolase by protein kinase A and aromatase-dependent pathways in mouse primary sertoli cells. Endocrinology. 2007;148:1431–1439. doi: 10.1210/en.2006-0969. [DOI] [PubMed] [Google Scholar]
- 52.Di Marzo V., Bisogno T., Sugiura T., Melck D., De Petrocellis L. The novel endogenous cannabinoid 2-arachidonoylglycerol is inactivated by neuronal- and basophil-like cells: Connections with anandamide. Pt 1Biochem. J. 1998;331:15–19. doi: 10.1042/bj3310015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chalmel F., Rolland A.D. Linking transcriptomics and proteomics in spermatogenesis. Reproduction. 2015;150:R149–R157. doi: 10.1530/REP-15-0073. [DOI] [PubMed] [Google Scholar]
- 54.Geremia R., Boitani C., Conti M., Monesi V. RNA synthesis in spermatocytes and spermatids and preservation of meiotic RNA during spermiogenesis in the mouse. Cell Differ. 1977;5:343–355. doi: 10.1016/0045-6039(77)90072-0. [DOI] [PubMed] [Google Scholar]
- 55.Lewis S.E.M., Rapino C., Di Tommaso M., Pucci M., Battista N., Paro R., Simon L., Lutton D., Maccarrone M. Differences in the endocannabinoid system of sperm from fertile and infertile men. PLoS ONE. 2012;7:e47704. doi: 10.1371/journal.pone.0047704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Amoako A.A., Marczylo T.H., Marczylo E.L., Elson J., Willets J.M., Taylor A.H., Konje J.C. Anandamide modulates human sperm motility: Implications for men with asthenozoospermia and oligoasthenoteratozoospermia. Hum. Reprod. 2013;28:2058–2066. doi: 10.1093/humrep/det232. [DOI] [PubMed] [Google Scholar]
- 57.Barbonetti A., Vassallo M.R.C., Fortunato D., Francavilla S., Maccarrone M., Francavilla F. Energetic metabolism and human sperm motility: Impact of CB₁ receptor activation. Endocrinology. 2010;151:5882–5892. doi: 10.1210/en.2010-0484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chioccarelli T., Cacciola G., Altucci L., Lewis S.E.M., Simon L., Ricci G., Ledent C., Meccariello R., Fasano S., Pierantoni R., et al. Cannabinoid receptor 1 influences chromatin remodeling in mouse spermatids by affecting content of transition protein 2 mRNA and histone displacement. Endocrinology. 2010;151:5017–5029. doi: 10.1210/en.2010-0133. [DOI] [PubMed] [Google Scholar]
- 59.Bari M., Battista N., Pirazzi V., Maccarrone M. The manifold actions of endocannabinoids on female and male reproductive events. Front. Biosci. (Landmark Ed.) 2011;16:498–516. doi: 10.2741/3701. [DOI] [PubMed] [Google Scholar]
- 60.Forner-Piquer I., Mylonas C.C., Calduch-Giner J., Maradonna F., Gioacchini G., Allarà M., Piscitelli F., Di Marzo V., Pérez-Sánchez J., Carnevali O. Endocrine disruptors in the diet of male Sparus aurata: Modulation of the endocannabinoid system at the hepatic and central level by Di-isononyl phthalate and Bisphenol A. Environ. Int. 2018;119:54–65. doi: 10.1016/j.envint.2018.06.011. [DOI] [PubMed] [Google Scholar]
- 61.Organisation for Economic Co-Operation and Development Number 78. Final Report of the Validation of the 21-day Fish Screening Assay for the Detection of Endocrine Active Substances. Phase 2: Testing Negative Substances. [(accessed on 5 June 2019)];2007 Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2006)27&doclanguage=en.
- 62.Organisation for Economic Co-Operation and Development Number 109. Literature Review on the 21-day Fish Assay and the Fish Short-term Reproduction Assay. [(accessed on 5 June 2019)];2009 Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2009)14&doclanguage=en.
- 63.Mylonas C.C., Cardinaletti G., Sigelaki I., Polzonetti-Magni A. Comparative efficacy of clove oil and 2-phenoxyethanol as anesthetics in the aquaculture of European sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) at different temperatures. Aquaculture. 2005;246:467–481. doi: 10.1016/j.aquaculture.2005.02.046. [DOI] [Google Scholar]
- 64.Papadaki M., Mazzella D., Santinelli V., Fakriadis I., Sigelaki I., Mylonas C.C. Hermaphroditism and reproductive function of hatchery-produced sharpsnout seabream (Diplodus puntazzo) under attenuated annual thermal cycles. Aquaculture. 2018;482:231–240. doi: 10.1016/j.aquaculture.2017.09.028. [DOI] [Google Scholar]
- 65.Bennett H.S., Wyrick A.D., Lee S.W., McNeil J.H. Science and art in preparing tissues embedded in plastic for light microscopy, with special reference to glycol methacrylate, glass knives and simple stains. Stain Technol. 1976;51:71–97. doi: 10.3109/10520297609116677. [DOI] [PubMed] [Google Scholar]
- 66.Nash J.P., Cuisset B.D., Bhattacharyya S., Suter H.C., Le Menn F., Kime D.E. An enzyme linked immunosorbant assay (ELISA) for testosterone, estradiol, and 17,20β-dihydroxy-4-pregenen-3-one using acetylcholinesterase as tracer: Application to measurement of diel patterns in rainbow trout (Oncorhynchus mykiss) Fish Physiol. Biochem. 2000;22:355–363. doi: 10.1023/A:1007850014021. [DOI] [Google Scholar]
- 67.Rodríguez L., Begtashi I., Zanuy S., Carrillo M. Development and validation of an enzyme immunoassay for testosterone: Effects of photoperiod on plasma testosterone levels and gonadal development in male sea bass (Dicentrarchus labrax, L.) at puberty. Fish Physiol. Biochem. 2000;23:141–150. doi: 10.1023/A:1007871604795. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.