

ABSTRACT
A milder climate has during the last decade contributed to an increased density and spreading of ixodid ticks, thus enhancing their role as emerging vectors for pathogenic microorganisms in northern Europe. It remains unclear if they contribute to the occurrence of infections caused by the bacteria Bartonella spp., Francisella tularensis subspecies holarctica and the parasite Toxoplasma gondii in Sweden and on the Åland islands, Finland. In this study, we want to improve understanding of the tick-borne transmission of these pathogens. Volunteers were recruited at primary healthcare centers. Ticks and blood samples were acquired from participants recruited in 2008 and 2009. Health questionnaires were completed, and medical records were acquired where applicable. Feeding time was estimated and screening of pathogens in the ticks was performed through real-time PCR. Ticks (n = 1849) were of mixed developmental stages: 76 larvae, 1295 nymphs, 426 adults and 52 undetermined. All analyzed ticks were considered negative for these pathogens since the CT-values were all below the detection limit for Bartonella spp. (1663 ticks), Francisella spp. (1849 ticks) and Toxoplasma gondii (1813 ticks). We assume that infections with these pathogens are caused by other transmission pathways within these regions of Sweden and the Åland islands, Finland.
KEYWORDS: Ixodes ricinus, Bartonella spp., Francisella tularensis, Toxoplasma gondii, real-time PCR
Introduction
The ixodid ticks are emerging vectors of potential pathogenic microorganisms in northern Europe. Following a milder climate, density and spreading of the ixodid ticks have increased. The prospective Tick-Borne Diseases STING-study (TBD STING) aimed to investigate the risk and factors affecting the onset of tick-borne diseases following a tick bite [1]. Within the TBD STING material the following pathogens have been detected: Borrelia spp. [1], Tick-borne encephalitis virus [2], Anaplasma phagocytophilum [3], Rickettsia helvetica [4], Candidatus Neoehrlichia mikurensis [5]. Other assumed tick-borne pathogens that are yet to be detected within the TBD STING study include the bacteria Bartonella spp., Francisella tularensis and the parasite Toxoplasma gondii. These pathogens have been detected within Ixodes ricinus in neighboring countries (Table 1) such as Germany (F. tularensis) [6], Denmark (Bartonella spp.) [7] and Poland (T. gondii) [8, 9]. Since infection with F. tularensis subspecies (ssp.) holarctica is notifiable there is an incidence reported by the Swedish Agency of Public Health [10]; it has varied from 0.9 and 8.7 per 100 000 person-years [11, 12]. Neither Bartonella spp. nor T. gondii is notifiable and thus the best data available in Sweden are blood donor serology: 16.1% for Bartonella spp. [[13]] and 23% for T. gondii [14].
Table 1.
Epidemiology of Bartonella spp., F. tularensis ssp. holarctica and T. gondii.
| Pathogen | Occurrence in Sweden | Prevalence in Ixodes ricinus | Other vectors and reservoirs |
|---|---|---|---|
| Bartonella spp. | 16% blood donor seroprevalence [13] | 4/661 Denmark [7] Ticks were collected from Danish domestic dogs. Detection through PCR and sequencing. | Cats [15] |
| F. tularensis ssp. holarctica | Reported incidence [10] 2015: 8.7/100 000 2017: 0.9/100 000 |
4-16/1556 Germany [6] Field collected ticks. Detection through MALDI-TOF mass spectrometry or 16S rRNA gene sequencing. | Mosquitos, Rodents, Environment [16, 15] |
| T. gondii | 23% blood donor seroprevalence [14] | 74/114 Poland [9] Pet and field collected ticks. 33/259 Poland [8] Field collected ticks. Detection through nested PCR. |
Warm blooded vertebrates [17] |
These emerging pathogens may present with an initial partly similar clinical picture. It is thus of clinical importance to be aware of these unusual tick-borne diseases as antibiotic treatment differs, and untreated or maltreated disease may have severe consequences. Prolonged bacteremia of virulent strains of Bartonella may result in endocarditis [15], arthropod transmission of F. tularensis holarctica may give rise to typhoidal tularemia (which is a severe systemic disease) [16] and toxoplasmosis in pregnant women may lead to abortion [17]. There are no published data on the prevalence of these pathogens in neither questing nor blood-feeding ticks collected in Sweden except for a minor study [18] that found 167 questing ticks from the Stockholm/Uppsala area in Sweden negative for Bartonella spp..
This study aims to investigate the risk of infection by Bartonella spp., F. tularensis or T. gondii after exposure to a tick-bite in different regions of Sweden and the Åland Islands, Finland by determining the prevalence of these pathogens in blood-feeding ticks collected from humans.
Materials and methods
Study design of the TBD STING study and collection of samples and preparation
Healthy, adult tick-bitten volunteers were recruited at primary healthcare centers through advertisements (Figure 1). Written consent, ticks, blood samples and health questionnaires were collected at inclusion at the first visit to the primary health-care center within 3 days after removal of the tick by the participant or by personnel at the primary health-care center. At the 3 months follow-up visit to the primary healthcare center, blood samples were collected, and a new questionnaire was completed. Medical records were available if the participant had visited the primary healthcare center during the 3-months study period. Samples were collected between 2007 and 2015 and were transported to Linköping University Hospital. Collected ticks were stored at −70°C until analysis and were morphologically determined to be I. ricinus and feeding time was estimated [19]. Nucleic acids were extracted, and the complementary (c) DNA was used in real-time PCR analysis of tick-borne pathogens as previously described by Wilhelmsson et al. [1]. In the event of positive real-time PCR screening, confirmation by conventional PCR analysis and sequencing. Successful sequencing would incur examination of the participant’s health questionnaires, serology of the blood samples and analysis of medical records [1].
Figure 1.
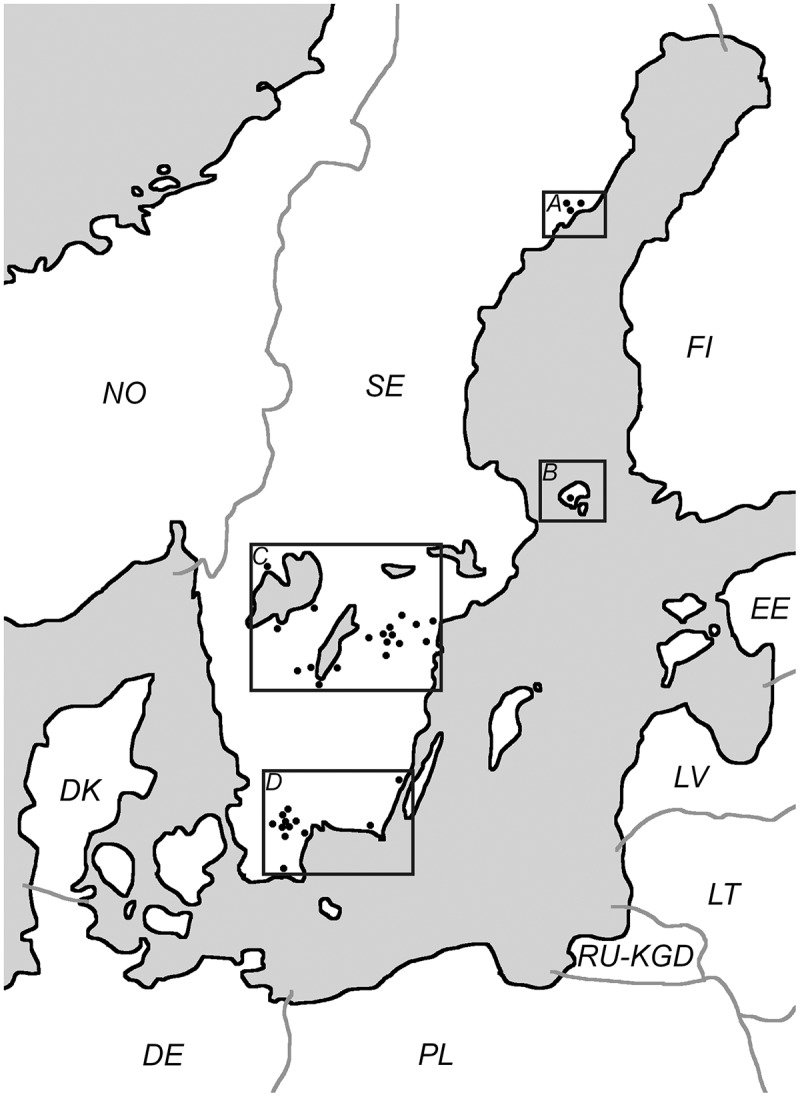
Distribution of the participating primary healthcare centers (n = 34), each black dot represents a primary healthcare center. Regions shown are A Northern Sweden (15 ticks), B Åland islands, Finland (633 ticks), C South Central Sweden (800 ticks) and D Southernmost Sweden (401 ticks). Countries coded in accordance with ISO 3166.
Ethics
The TBD STING study was approved by the Regional Ethical review board at Linköping University (M132-06) and by the local Ethics Committee of the Åland Health Care, 2008-05-23.
Real-time PCR assay for analyses of tick-borne pathogens
Bartonella spp.
The individually synthesized cDNA was analyzed in pools of four with 0.5 μl of cDNA from each tick. The forward and reverse primers (Invitrogen Corporation) (Table 2) were used in an SYBR Green assay. The primers target the ssrA gene, which is a prokaryotic gene involved in translatory regulation [20]. The positive control used was Bartonella grahamii DNA [30 ng/µl], kindly provided by Dr. Martin Andersson, Department of Biology, Lund University, Sweden. Detection limit was determined at [3 pg/µl].
Table 2.
Real-time PCR assays used to detect Bartonella spp., Francisella spp. and T. gondii in I. ricinus.
| Sequence (5ʹ-3ʹ) | Target gene | Size of amplicon (bp) | Reference | |
|---|---|---|---|---|
| Bartonella spp. | ssrA1 | 301 | Diaz et al. [20] | |
| Forward | GCT ATG GTA ATA AAT GGA CAA TGA AAT AA | |||
| Reverse | GCT TCT GTT GCC AGG TG | |||
| Francisella spp. | sucC2 | 125 | This study | |
| Forward | AAC TGG CTG ACC TTC AGC AT | |||
| Reverse | GTG GTC GTG GTA AAG CTG GT | |||
| Probe | FAM3-CCG ATT AGG CTT TCT GCT ACT TCA CGA-BHQ14 | |||
| Forward (PHV5) | GGG CGA ATC ACA GAT TGA ATC | |||
| Reverse (PHV) | GCG GTT CCA AAC GTA CCA A | |||
| Probe (PHV) | TR6-TTT TTA TGT GTC CGC CAC CAT CTG GAT C-BHQ1 | |||
| T. gondii | B17 | 98 | Lin et al. [22] | |
| Forward | TCC CCT CTG GCG AAA AGT | |||
| Reverse | AGC GTT CGT GGT CAA CTA TCG ATT G | |||
| Probe | FAM-TCT GTG CAA CTT TGG TGT ATT CGC AG-TAMRA8 |
One reaction contained 16 µl master mix and 4 µl sample cDNA. The master mix contained 7.2 µl of RNase/DNase free water, 10 µl of Maxima SYBR Green qPCR Master Mix (Thermo Fisher Scientific), 0.4 µl of Bartonella forward primer [10 µM] and 0.4 µl of Bartonella reverse primer [10 µM].
The PCR assay consisted of one initializing denaturation step at 95°C for 2 min and then 45 cycles of 95°C denaturation for 15 s, 60°C for 60 s and 72°C for 15 s ending with 72°C for 5 min. Immediately after real-time PCR analysis, melting curve analyses were performed by heating to 95°C for 15 s, followed by cooling to 60°C for 1 min, and subsequent heating to 95°C at 0.8°C min-1 with continuous fluorescence recording. A C1000 thermal cycler (Bio-Rad Laboratories Inc, Hercules, CA) was used for the real-time PCR.
Francisella spp.
The individually synthesized cDNA was analyzed in pools of four with 1 μl of cDNA from each tick. The primers and the probe used were developed and provided by the Swedish Defense Research Agency (FOI) and target a 125 bp long fragment of the gene coding for the Succinyl-CoA-synthetase beta chain of the Francisella genus (Table 2). An internal control consisting of Phocine Herpes Virions [21], kindly provided by the Public Health Agency of Sweden was used to make sure that the PCR reaction was working despite the pool being negative for Francisella. The positive control used was Francisella tularensis subsp. holarctica live vaccine strain (LVS) extracted DNA from culture by FOI [38.3 ng/μl]. Detection limit was determined at [38.3 fg/µl].
One reaction contained 21 µl of master mix and 4 µl sample cDNA. The master mix contained 4.5 µl of RNase/DNase free water, 12.5 µl of Maxima Probe qPCR Master Mix [2X] (Thermo Fisher Scientific), 0.625 µl of Francisella spp. forward primer [20 µM], 0.625 µl of Francisella spp. reverse primer [20 µM], 0.5 µl of Francisella spp. probe [5 µM], 0.625 µl of internal control forward primer [20 µM], 0.625 µl of internal control reverse primer [20 µM], 0.5 µl of Internal control probe [5 µM] and 0.5 µl DNA for the internal control.
The PCR assay consisted of one initializing denaturation at 95°C for 2 min and then 45 cycles of 95°C denaturation for 30 s and 60°C Annealing/Elongation for 60 s. A C1000 thermal cycler (Bio-Rad) was used for the real-time PCR.
Toxoplasma gondii
The individually synthesized cDNA was analyzed in pools of four with 0.5 μl of cDNA from each tick. The primers and probe (Invitrogen Corporation) target the B1 gene of T. gondii [22], its function is unknown, but it is present in all investigated genotypes of T. gondii. The positive control used was T. gondii DNA [445 ng/µl]. It was kindly provided by professor Krzysztof Solarz through Dr Olga Pawelczyk, both the Department of Parasitology, Medical University of Silesia, Poland. Detection limit was determined at [445 pg/µl].
One reaction contained 18 µl of master mix and 2 µl sample cDNA. The master mix contained 6.3 of µl RNase/DNase free water, 10.5 µl of Maxima Universal Master Mix (Thermo Fisher Scientific), 0.4 µl of T. gondii forward primer [5 µM], 0.4 µl of T. gondii reverse primer [5 µM], 0.4 µl of TaqMan probe [10 µM]. A period of 30 s at 50°C was added to every cycle before the 15 s of denaturation. A C1000 thermal cycler (Bio-Rad) was used for the real-time PCR.
The PCR assay consisted of one initializing denaturation at 95°C for 10 min and then 40 cycles of 95°C denaturation for 15 s and 60°C Annealing/Elongation for 60 s. A C1000 thermal cycler (Bio-Rad) was used for the real-time PCR.
Sequencing of positive samples
Any positive pools were repeated individually in the real-time PCR assay and then conventional PCR and sequencing.
Any real-time PCR products matching the positive control in size were sequenced by Macrogen (Amsterdam, Netherlands).
Results
In total, cDNA from 1 849 ticks (76 larvae, 1295 nymphs, 426 adults, and 52 were unable to be morphologically staged) collected in 2008 and 2009, were available for analysis. The distribution of primary healthcare centers can be seen in Figure 1. All these ticks were analyzed for Francisella spp., 1 813 ticks for T. gondii and 1 663 ticks for Bartonella spp. The variance in number of analyzed ticks is due to limited amounts of cDNA. All samples were negative i.e. all CT-values obtained were below the detection limit.
Discussion
In Sweden, vector-borne tularemia is mainly associated with mosquitos [23, 24, 25] and it is unknown whether ticks contribute to the presence of the disease in Sweden. However, there are cases of explicitly suspected tick-borne tularemia in Sweden [26, 27]. Despite the high number of ticks analyzed in this study, Francisella spp. remains to be detected in ticks collected in Sweden and the Åland islands, Finland. F. tularensis has been found in I. ricinus collected in Germany within a smaller sample size by Tomaso et al. [6], it is thus likely that the prevalence is lower in the southern regions of Sweden and on the Åland islands, Finland. It is still uncertain how F. tularensis is transmitted between ticks; transovarian transmission was shown in early experiments [28, 29] but has not been reproduced with modern methods [30]. The mode of vector-mediated transmission matters since it could affect the geographic distribution of the bacteria; transovarian transmission would likely result in highly concentrated spots of related ticks colonized with F. tularensis, while a vector-reservoir transmission could dilute the infested ticks over a greater area. In this study, the participating primary healthcare centers (Figure 1) do not co-localize with the geographic areas known to be endemic for tularemia and there are only 15 ticks collected from the Northern region of Sweden. Since tularemia in Sweden is known to be a focal disease in the middle and northern parts of Sweden [11, 12] it would be a topic for further studies to include additional primary healthcare centers from the areas endemic for the disease.
Bartonella spp. and T. gondii also remain to be detected in ticks collected in Sweden even though there is seroprevalence among healthy Swedish blood donors (Bartonella spp.: 16.1% T. gondii: 23%) [14, 13]. Interestingly, these pathogens have been found in I. ricinus collected in Poland [9] and Denmark [7]. Due to the larger sample size in this study, it is likely that the prevalence of these pathogens in I. ricinus from South Central and Southernmost Sweden and the Åland islands, Finland is considerably lower. Regarding these pathogens, it is possible that the seroprevalence is due to proximity to household cats (cat scratch disease or T. gondii oocysts in cat feces) instead of ticks.
A possible explanation for that we were unable to detect Bartonella and T. gondii in I. ricinus collected in Sweden may be the climate change affecting the distribution and prevalence of tick-borne pathogens. The lack of Bartonella in Norway [31] and Finland [32] supports that tick-borne Bartonella spp. might not tolerate the harsher climate in most of the Nordic countries compared to Denmark [7]. There are no previously published data on the prevalence of T. gondii in neither blood-feeding nor questing ticks from the Nordic countries. Up to 4 days may have elapsed between the removal of the tick and freezing at −70°C. It is unlikely that lysis of the bacterial cells within the tick may have affected the quality of the extracted nucleic acids. We have previously detected several pathogens using the above-mentioned approach [1, 3, 4].
In conclusion, the results obtained from this large material indicate that due to the absence of detection, within South Central and Southernmost Sweden and the Åland islands, there is very low risk of tick-borne infection caused by Bartonella spp., F. tularensis or T. gondii. This knowledge may be useful when deciding on antibiotic treatment in conjunction with a tick bite. It is likely that ticks do not significantly contribute to the presence of these pathogens in South Central and Southernmost Sweden and the Åland islands, Finland. It is also possible that the lack of detection is explained by very focal occurrence of the pathogens.
Biographies
Samuel Cronhjort is a junior researcher at the Division of Medical Microbiology, Department of Clinical and Experimental Medicine at Linköping University with a research interest in tick-borne disease.
Peter Wilhelmsson is a specialist in microbiology and molecular diagnostics with a position as a researcher in Region Jönköping County and as adjunct senior lecturer at Linköping University. He has pursued research in the field of tick-borne diseases for more than 10 years.
Linda Karlsson is a research engineer at Swedish defence research agency, the division of CBRN defence and security. Her focus is on molecular biological methods of high-risk agents, including next generation sequencing.
Johanna Thelaus defended her PhD in Microbial Ecology at Umeå University in 2008. Since then, she has been associated to the Swedish Defence Research Agency, FOI, the division of CBRN Defence and Security with focus on Francisella tularensis ecology as well as work related to Swedish civil defence and bio preparedness.
Andreas Sjödin is Deputy Research Director at Swedish Defence Research Agency (FOI). His work focus on metagenomics and genome evolution with expertise in biological select agents.
Pia Forsberg is professor emerita in infectious medicine at Linköping university and a senior consultant in infectious diseases. Research interest in tick borne diseases.
Per-Eric Lindgren received his PhD at Dept. Microbiology, SLU and since 2010 he is professor and chair of Medical Microbiology, Department of Clinical and Experimental Medicine, Linköping University, Linköping, Sweden. Project leader of the EU Interreg North Sea Region project NorthTick.
Funding Statement
This work was supported by the Medical Research Council of South-East Sweden under Grant FORSS-657881; EU-interreg programme ScandTick Innovation under Project ID [20200422], reference no 2015-000167; the Swedish Civil Contingencies Agency under Grant [B4010]; Swedish Research Council Branch of Medicine under Grant no. K2008-58X-14631-06-3 Swedish Ministry of Defence under Grant [A4042];
Notes
Involved in translator regulation.
Succinyl-CoA reductase.
6-carboxyfluorescein.
Black Hole Quencher.
Phocine Herpes Virion.
Texas Red fluorescein.
Gene of unknown significance.
5-Carboxytetramethylrhodamine.
Acknowledgments
We would like to thank all participants of the TBD STING study as well as the staff at the PHCs involved in the TBD STING-study. We would also like to thank the TBD STING-study group, consisting of Clas Ahlm, Johan Berglund, Sten-Anders Carlsson, Christina Ekerfelt, Mats Haglund, Anna J Henningsson, Peter Nolskog, Dag Nyman, Marika Nordberg and Katarina Ornstein for all the valuable work and advice on the study design. A special thanks to Liselott Lindvall, Susanne Olausson, and Mari-Anne Åkeson for their continuous work with the study collection logistics. We would also like to thank Karoline Eklund for performing the PCR assay for Bartonella spp. and T. gondii on material collected in 2008.
Disclosure statement
No potential conflict of interest was reported by the authors.
Data availability statement
The data supporting the conclusions of this article are included within the article. Raw data can be shared with researchers upon a specific request.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
References
- [1].WILHELMSSON P, FRYLAND L, LINDBLOM P, et al. A prospective study on the incidence of Borrelia burgdorferi sensu lato infection after a tick bite in Sweden and on the Aland Islands, Finland (2008–2009). Ticks Tick Borne Dis. 2016;7:71–6. [DOI] [PubMed] [Google Scholar]
- [2].LINDBLOM P, WILHELMSSON P, FRYLAND L, et al. Tick-borne encephalitis virus in ticks detached from humans and follow-up of serological and clinical response. Ticks Tick Borne Dis. 2014;5:21–28. [DOI] [PubMed] [Google Scholar]
- [3].HENNINGSSON AJ, WILHELMSSON P, GYLLEMARK P, et al. Low risk of seroconversion or clinical disease in humans after a bite by an Anaplasma phagocytophilum-infected tick. Ticks Tick Borne Dis. 2015;6:787–792. [DOI] [PubMed] [Google Scholar]
- [4].LINDBLOM A, WALLMENIUS K, SJOWALL J, et al. Prevalence of Rickettsia spp. in ticks and serological and clinical outcomes in Tick-Bitten individuals in Sweden and on the Aland Islands. PLoS One. 2016;11:e0166653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].GRANKVIST A, SANDELIN LL, ANDERSSON J, et al. Infections with candidatus neoehrlichia mikurensis and cytokine responses in 2 persons bitten by Ticks, Sweden. Emerg Infect Dis. 2015;21:1462–1465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].TOMASO H, OTTO P, PETERS M, et al. Francisella tularensis and other bacteria in hares and ticks in North Rhine-Westphalia (Germany). Ticks Tick Borne Dis. 2018;9:325–329. [DOI] [PubMed] [Google Scholar]
- [7].STENSVOLD CR, MARAI AL, ANDERSEN LO, et al. Babesia spp. and other pathogens in ticks recovered from domestic dogs in Denmark. Parasit Vectors. 2015;8:262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].SROKA J, WÓJCIK-FATLA A, ZWOLIŃSKI J, et al. Preliminary study on the occurrence of Toxoplasma gondii in Ixodes ricinus ticks from north-western Poland with the use of PCR. Ann Agric Environ Med. 2008;15:333–338. [PubMed] [Google Scholar]
- [9].ASMAN M, SOLARZ K, CUBER P, et al. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: ixodida) collected in Tarnogorski district (Upper Silesia, Poland). Ann Agric Environ Med. 2015;22:80–83. [DOI] [PubMed] [Google Scholar]
- [10].FOLKHÄLSOMYNDIGHETEN Epidemiologisk årsrapport, tabellsamling. FOLKHÄLSOMYNDIGH-ETEN (ed.) 2019. [Google Scholar]
- [11].DESVARS-LARRIVE A, LIU X, HJERTQVIST M, et al. High-risk regions and outbreak modelling of tularemia in humans. Epidemiol Infect. 2017;145:482–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].DESVARS A, FURBERG M, HJERTQVIST M, et al. Epidemiology and ecology of tularemia in Sweden, 1984–2012. Emerg Infect Dis. 2015;21:32–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].MCGILL S, WESSLEN L, HJELM E, et al. Bartonella spp. seroprevalence in healthy Swedish blood donors. Scand J Infect Dis. 2005;37:723–730. [DOI] [PubMed] [Google Scholar]
- [14].BIRGISDOTTIR A, ASBJORNSDOTTIR H, COOK E, et al. Seroprevalence of toxoplasma gondii in Sweden, Estonia and Iceland. Scand J Infect Dis. 2006;38:625–631. [DOI] [PubMed] [Google Scholar]
- [15].ANGELAKIS E, RAOULT D.. Pathogenicity and treatment of Bartonella infections. Int J Antimicrob Agents. 2014;44:16–25. [DOI] [PubMed] [Google Scholar]
- [16].MAURIN M, GYURANECZ M. Tularaemia: clinical aspects in Europe. Lancet Infect Dis. 2016;16:113–124. [DOI] [PubMed] [Google Scholar]
- [17].SKOTARCZAK BI. The role of ticks in transmission cycle of Toxoplasma gondii. Ann Parasitol. 2016;62:185–191. [DOI] [PubMed] [Google Scholar]
- [18].SCOLA BL, HOLMBERG M, RAOULT D. Lack of bartonella sp. in 167 ixodes ricinus ticks collected in Central Sweden. Scand J Infect Dis. 2009;36:305–306. [DOI] [PubMed] [Google Scholar]
- [19].GRAY J, STANEK G, KUNDI M, et al. Dimensions of engorging Ixodes ricinus as a measure of feeding duration. Int J Med Microbiol. 2005;295:567–572. [DOI] [PubMed] [Google Scholar]
- [20].DIAZ MH, BAI Y, MALANIA L, et al. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J Clin Microbiol. 2012;50:1645–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].VAN DOORNUM GJ, GULDEMEESTER J, OSTERHAUS AD, et al. Diagnosing herpesvirus infections by real-time amplification and rapid culture. J Clin Microbiol. 2003;41:576–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].LIN M-H, CHEN T-C, KUO -T-T, et al. Real-time PCR for quantitative detection of toxoplasma gondii. J Clin Microbiol. 2000;38:4121–4125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].ELIASSON HLJ, NUORTI JP, ARNEBORN M, et al. The 2000 Tularemia outbreak: a case-control study of risk factors in disease-endemic and emergent areas, Sweden. Emerg Infect Dis. 2002;8:956–960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].RYDEN P, BJORK R, SCHAFER ML, et al. Outbreaks of tularemia in a boreal forest region depends on mosquito prevalence. J Infect Dis. 2012;205:297–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].THELAUS J, ANDERSSON A, BROMAN T, et al. Francisella tularensis subspecies holarctica occurs in Swedish mosquitoes, persists through the developmental stages of laboratory-infected mosquitoes and is transmissible during blood feeding. Microb Ecol. 2014;67:96–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].ÖHRMALM L, TYNELL E. Misstänk harpest vid typiska symtom även utan typisk anamnes. Läkartidningen. 2012;109:1172–1173. [PubMed] [Google Scholar]
- [27].LUNDMAN T. Se upp för tularemi även i södra Sverige. Läkartidningen. 2005;102:1986–1987. [PubMed] [Google Scholar]
- [28].PHILIP CB, JELLISON WL. The American dog tick, dermacentor variabilis, as a host of bacterium tularense. Public Health Rep. 1934;49:386–392. [Google Scholar]
- [29].PARKER RR, SPENCER RR. Hereditary Transmission of Tularæmia Infection by the Wood Tick, Dermacentor andersoni Stiles. Public Health Rep. 1926;41:1403–1407.19315042 [Google Scholar]
- [30].GENCHI M, PRATI P, VICARI N, et al. Francisella tularensis: no Evidence for Transovarial Transmission in the Tularemia Tick Vectors Dermacentor reticulatus and Ixodes ricinus. PLoS One. 2015;10:e0133593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].QUARSTEN H, SKARPAAS T, FAJS L, et al. Tick-borne bacteria in Ixodes ricinus collected in southern Norway evaluated by a commercial kit and established real-time PCR protocols. Ticks Tick Borne Dis. 2015;6:538–544. [DOI] [PubMed] [Google Scholar]
- [32].SORMUNEN JJ, PENTTINEN R, KLEMOLA T, et al. Tick-borne bacterial pathogens in southwestern Finland. Parasit Vectors. 2016;9:168. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data supporting the conclusions of this article are included within the article. Raw data can be shared with researchers upon a specific request.