

Abstract
Inspired by the roles of serotonin in an emotional aversion to harmful actions, we examined to what extent serotonin transporter gene (5-HTT)–linked polymorphic region (5-HTTLPR), a proxy for measuring serotonin function, underpinned the individual differences in moral judgment through cross-sectional analysis and two-wave comparison. The cross-sectional analysis with a larger cohort (N = 1197) showed that the SS carriers of the 5-HTTLPR polymorphism, corresponding to the low ratio of serotonin recycling from the synaptic cleft, rated impersonal harmful actions (e.g. flipping a switch to divert a train to hit one person instead of five people) as more permissible as compared with the L-allele carriers. The two-wave comparison with a subsample from the larger cohort (N = 563) indicated that the association between 5-HTTLPR polymorphism and moral permissibility of impersonal harmful actions was stable from wave 1 to wave 2. Thus, these findings highlight the importance of the 5-HTTLPR polymorphism to harmful moral behaviors.
Keywords: moral judgment, personal harm, impersonal harm, serotonin transporter gene, cross-sectional analysis, two-wave comparison
Introduction
Moral judgment refers to an evaluation or opinion regarding whether one’s behaviors are right or wrong according to ethical rules such as advocating for others’ well-being and prohibiting harming others (Heiphetz et al., 2018), and thereby, such judgment is related to one’s humanity identification (Kahane et al., 2015), prosocial behaviors (Ding et al., 2018; Li et al., 2018) and psychopathic status (Outhred et al., 2016). Permissibility of moral judgment, an acceptability for transgression on ethical rules, is restrained by an emotional aversion to harmful actions (Gleichgerrcht et al., 2013; McDonald et al., 2017), such that people are less likely to endorse emotional harms as permissible, even if doing so would achieve a greater good (Greene et al., 2001; Koenigs et al., 2012). In light of emotional aversion to social harms infusing moral judgment, the permissibility of harms in sacrificial moral dilemmas, where harmful actions promote the greater good (Greene et al., 2001), is greatly modulated by such emotion.
Serotonin is an ancient molecule that drives individuals’ avoidant responses to harmful social stimuli (Crockett et al., 2010; Siegel & Crockett, 2013). Violent social stimuli, harmful actions of sacrificial moral dilemmas (i.e. pushing one person in front of a runaway train to prevent it from hitting five other people), can trigger salient emotional aversion and harm-withdrawal responses (Greene et al., 2001; Koenigs et al., 2012). Studies indicated that serotonin levels in the brain shape permissibility of moral judgment through an up-regulation of emotional aversion to harmful actions (Crockett et al., 2010; Siegel & Crockett, 2013). Specifically, a serotonin administration study with 24 health participants indicates that taking citalopram (a selective serotonin reuptake inhibitor), which boosts serotonin levels by blocking serotonin reuptake (Sanchez et al., 2004), makes the subjects judge emotionally salient harms (i.e. personally killing an innocent person) as more impermissible compared to taking placebo (Crockett et al., 2010). Moreover, a serotonin depletion study indicates that a modest depletion of brain serotonin, relative to place, could abolish one’s withdrawal behavior in the face of aversive prediction. Therefore, basing on the previous findings, serotonin could modulate one’s permissibility rating of moral judgment by amplifying their emotional aversion and withdrawal behavior to harm.
Serotonin transporter (5-HTT) is a commonly investigated proxy for measuring serotonin functions. This transporter is largely recognized for its functions in transporting serotonin back into the pre-synaptic terminal after serotonin release (De Felice, 2016). The serotonin transporter gene linked polymorphic region (5-HTTLPR) is characterized by a short (S) allele and a long (L) allele, differing in the number of 20–23-base-pair repeated units. The S allele of this polymorphism is related to a decreased 5-HTT transcription, and thereby increases the synaptic serotonin levels by reducing the ratio of serotonin recycling from the synaptic cleft (Greenberg et al., 1999; Canli & Lesch, 2007). Studies have shown that the 5-HTTLPR polymorph is widely involved in emotion processing (Jonassen & Landro, 2014; Raab et al., 2016), such that the S allele of this polymorph is related to high sensitivity to emotional stimuli compared to the LL genotype (Jonassen & Landro, 2014). Inspired by the determination of serotonin to emotional aversion (Crockett et al., 2010; Siegel & Crockett, 2013) and the differential influences of the alleles of 5-HTTLPR polymorphism on serotonin levels via the serotonin transporter expression (Greenberg et al., 1999; Canli & Lesch, 2007), this polymorphism at the individual level may be involved in permissibility rating of moral behaviors. So far, a gene-association study with 63 healthy community registers indicated that the S allele of 5-HTTLPR is associated with lower permissibility rating of impersonal harms compared to the LL genotype (Marsh et al., 2011). The findings suggest that the 5-HTTLPR polymorphism may be associated with moral permissibility through the differential modulations of this polymorphism on emotion processing. Here, considering the differential effects of the 5-HTTLPR and citalopram on different types of harms (impersonal vs personal harms), and the small sample size of the previous gene-behavior study (Marsh et al., 2011), we re-examined the association with different experimental moral conditions in a larger sample.
Moral judgment is guided by one’s deontology (i.e. it is wrong to intervene in a way that causes a sacrifice of a few people) and utilitarianism (i.e. it is better to preserve a greater number of people’s well-being at the cost of a few persons’ interests; Greene et al., 2001; Koenigs et al., 2012). Given that such moral principles are referenced architectures of moral judgment, and a person who makes a particular judgment in one context will make a similar judgment in other contexts (Helzer et al., 2017; Hannikainen et al., 2018), one’s moral permissibility for a certain harmful action should be stable across time. However, previous gene-behavior studies up to date have only examined the associations between some genetic polymorphisms and moral judgment in a certain time window (Marsh et al., 2011; Bernhard et al., 2016; Gong et al., 2017; Ru et al., 2017), whether such associations remain stable across time is still unclear. Therefore, in this study, we investigated the stability of the association between 5-HTTLPR polymorphism and moral judgment through a two-wave longitudinal analysis.
Permissibility rating of moral judgment also depends on moral contexts such as dilemma type and frame of action (Crockett et al., 2010; Koenigs et al., 2012; Patil, 2015). The dilemma type (personal vs impersonal harms) indicates whether committing direct physical harm is involved in the moral scenarios (i.e. pushing one person off a bridge to stop a runaway train car from hitting five people); the frame of action (action vs inaction) refers to whether an action of the protagonist was taken or not to save five others by sacrificing one person. For the two binary factors, personal harms and performing harmful actions induce more salient emotional aversion as compared with impersonal harms and inactions (Greene et al., 2001). Thus, we conducted a 2 (dilemma type: personal vs impersonal) × 2 (frame of action: action vs inaction) × 3 (genotype: SS vs SL vs LL) study to examine whether the effects of 5-HTTLPR polymorphism on moral permissibility depend on emotional aversion to the harms of moral scenarios.
To overview, the study was conducted with the following ideas: (1) the moral permissibility rating of the harms in sacrificial dilemmas depends on the emotional aversion to harm; (2) serotonin is assumed to determine emotional aversion to harm; and (3) the alleles of the 5-HTTLPR differentially influence serotonin levels via the serotonin transporter expression. Based on this ideas, we investigated the association of 5-HTTLPR with individual differences in the permissibility and stability of gene-behavior association. In the cross-sectional analysis, we examined the association between the 5-HTTLPR polymorphism and moral judgment in a large college-student cohort (N = 1197). Given that high serotonin functions are related to low moral permissibility (Crockett et al., 2010; Marsh et al., 2011), we predicted that individuals with the SS genotype of 5-HTTLPR polymorphism, which is related to a low ratio of serotonin levels recycled from the synaptic cleft, were more likely to rate harmful actions as more impermissible than individuals with the SL and LL genotypes. In a two-wave comparison, we examined the association between 5-HTTLPR polymorphism and stability of permissibility rating in a subsample (N = 563) of the larger cohort. Given that a person who makes a particular judgment in one context tends to make a similar judgment in other contexts (Helzer et al., 2017; Hannikainen et al., 2018), we predicted that individuals with the SS genotype of 5-HTTLPR would rate harmful actions as more impermissible both at wave 1 and wave 2, with a 6-month interval.
A cross-sectional analysis of the association of 5-HTTLPR polymorphism with moral permissibility
Methods
Participants
One thousand and eight hundred college students of the Science and Technology University in China were invited to participate in this gene-behavior study. Within 1 week, 1197 students (823 females, mean age = 20.0 ± 1.4 years) voluntarily contacted us by telephone and signed the informed consents before the moral judgment assessment. These participants were ethnically Han Chinese, without any known ancestors of another ethnic origin. A priori power analysis with G*Power 3.1 (Faul et al., 2007) indicated that a sample of 787 participants is required (two-tailed α = 0.05, 1−β = 0.80) if the minimum regression coefficient reaches 0.01 (i.e. the coefficient of determination R2 = 0.01). However, given the high drop-out rate of participants in longitudinal studies, we were inclined to include all participants who signed up in the study. The study was carried out according to the principles of Declaration of Helsinki.
Moral judgment assessment
Moral judgment was assessed with 16 sacrificial dilemma scenarios (Online Supplementary Materials; Greene et al., 2001). For each scenario, participants indicated the extent to which the proposed action of the protagonist was permissible, with ‘1’ = totally impermissible and ‘7’ = totally permissible. To assess the permissibility rating of different harmful manners and actions, a 2 (dilemma type: personal vs impersonal) × 2 (frame of action: action vs inaction) design was adopted. For each experimental condition, we had four scenarios. The personal scenarios tap the direct physical harms, while the impersonal scenarios examine the indirect or remote harms (Greene et al., 2001). As for the frame of action (action vs inaction), the action scenarios involve actual actions of the protagonist to save five others by sacrificing one person as compared with the inaction scenarios (Greene et al., 2001). In this study, the test was administered in pencil-and-paper format in a group of 15–20 participants. The internal consistency of the moral judgment assessment, as measured with the Cronbach’s α, was 0.866 in this sample.
Genotyping
We collected three to five pieces of hair from each participant to extract genomic DNA with Chelex 100 method (de Lamballerie et al., 1994). For the 5-HTTLPR polymorphism, the upstream primer, 5′-ATCGCTCCTGCATCCCCCATTAT−3′, and the downstream primer, 5′-GAGGTGCAGGGGGATGCTGGAA—3′, were recruited (Perez-Edgar et al., 2010). The PCR reaction mixture contained 2.50 μl 2 × reaction mix, 0.20 μl DNA template, 1.90 μl ddH2O, 0.20 μl (25 pmol/μl) upstream primers and 0.20 μl (25 pmol/μl) downstream primers. The details of PCR procedures and polyacrylamide gel electrophoresis were described in our previous study (Yang et al., 2019). The 146 bp DNA fragment was assigned as the L allele, and the 103 bp DNA fragment was assigned as the S allele. In the larger sample, the distribution of genotypes deviated from Hardy–Weinberg equilibrium (SS = 909, SL = 249, LL = 39), χ2 = 16.67, P < 0.001. Moreover, the genotype frequencies in the sample significantly differed from the report of the previous study (SS = 13, SL = 30, LL = 20; Marsh et al., 2011), χ2 = 148.18, P < 0.001.
Results
A 2 (dilemma type: impersonal vs personal) × 2 (frame of action: action vs inaction) × 3 (genotype: SS vs SL vs LL) ANOVA analysis was conducted to reveal the impacts of the dilemma type, frame of action and genotypes on the permissibility rating of moral judgment. The results indicated that there were significant effects of the dilemma type, F(1, 1196) = 219.49, P < 0.001,  2 = 0.15; frame of action, F(1, 1196) = 172.01, P < 0.001, 2 = 0.13; and the interaction of dilemma type and frame of action, F(1, 1196) = 260.07, P < 0.001, 2 = 0.18. Specifically, the participants rated impersonal harms as more permissible than personal harms (mean ± SEM: impersonal vs personal = 4.15 ± 0.03 vs 3.35 ± 0.03), t(1196) = 33.07, P < 0.001, Cohen’s d = 0.78, and rated inaction harms as more permissible than action harms (mean ± SEM: inaction vs action = 4.06 ± 0.03 vs 3.44 ± 0.03), t(1196) = 24.33, P < 0.001, Cohen’s d = 0.58. Moreover, the above analysis indicated that the genotypes could significantly impact the permissibility rating for harms at the individual level, F(2, 1196) = 4.22, P = 0.015, 2 = 0.007. As described in the Figure 1, individuals with the SS carriers (mean ± SEM: 3.79 ± 0.03, N = 909) were more likely to rate the harms as permissible than individuals with the SL carriers (mean ± SEM: 3.61 ± 0.06, N = 249), P = 0.008, Cohen’s d = 0.15, while no significant difference between the SL group and the LL genotype was found (mean ± SEM: 3.58 ± 0.14, N = 39), P = 0.85, Cohen’s d = 0.02, and between the SS group and LL genotype group, P = 0.170, Cohen’s d = 0.17. Furthermore, a three-way interaction between the genotypes, dilemma type and frame of action on the permissibility rating of moral judgment were present, F(2, 1196) = 3.11, P = 0.045, 2 = 0.005. Given the interaction, we examined the impacts of the 5-HTTLPR polymorphism on the moral permissibility of the four harmful contexts, respectively. One-way ANOVA analysis indicated that the 5-HTTLPR polymorphism impacted the permissibility rating of impersonal action harms, F(2, 1196) = 6.50, P = 0.002, 2 = 0.011. The permissibility rating increased as a function of the number of S alleles, F(1, 1196) = 4.85, P = 0.028. However, this polymorphism was not associated with the permissibility ratings in the other experimental conditions (Figure 1).
2 = 0.15; frame of action, F(1, 1196) = 172.01, P < 0.001, 2 = 0.13; and the interaction of dilemma type and frame of action, F(1, 1196) = 260.07, P < 0.001, 2 = 0.18. Specifically, the participants rated impersonal harms as more permissible than personal harms (mean ± SEM: impersonal vs personal = 4.15 ± 0.03 vs 3.35 ± 0.03), t(1196) = 33.07, P < 0.001, Cohen’s d = 0.78, and rated inaction harms as more permissible than action harms (mean ± SEM: inaction vs action = 4.06 ± 0.03 vs 3.44 ± 0.03), t(1196) = 24.33, P < 0.001, Cohen’s d = 0.58. Moreover, the above analysis indicated that the genotypes could significantly impact the permissibility rating for harms at the individual level, F(2, 1196) = 4.22, P = 0.015, 2 = 0.007. As described in the Figure 1, individuals with the SS carriers (mean ± SEM: 3.79 ± 0.03, N = 909) were more likely to rate the harms as permissible than individuals with the SL carriers (mean ± SEM: 3.61 ± 0.06, N = 249), P = 0.008, Cohen’s d = 0.15, while no significant difference between the SL group and the LL genotype was found (mean ± SEM: 3.58 ± 0.14, N = 39), P = 0.85, Cohen’s d = 0.02, and between the SS group and LL genotype group, P = 0.170, Cohen’s d = 0.17. Furthermore, a three-way interaction between the genotypes, dilemma type and frame of action on the permissibility rating of moral judgment were present, F(2, 1196) = 3.11, P = 0.045, 2 = 0.005. Given the interaction, we examined the impacts of the 5-HTTLPR polymorphism on the moral permissibility of the four harmful contexts, respectively. One-way ANOVA analysis indicated that the 5-HTTLPR polymorphism impacted the permissibility rating of impersonal action harms, F(2, 1196) = 6.50, P = 0.002, 2 = 0.011. The permissibility rating increased as a function of the number of S alleles, F(1, 1196) = 4.85, P = 0.028. However, this polymorphism was not associated with the permissibility ratings in the other experimental conditions (Figure 1).
Fig. 1.
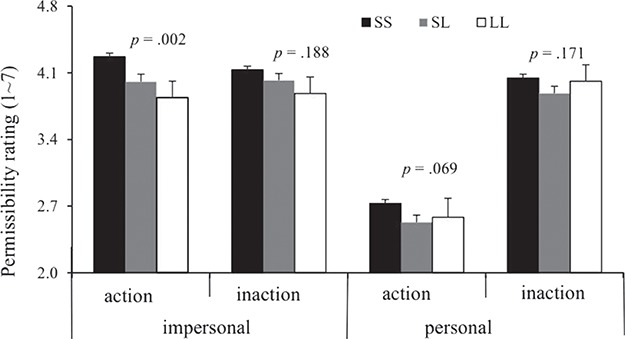
The effects of the 5-HTTLPR polymorphism on the permissibility rating (mean ± SEM) of moral dilemmas in the large cohort in wave 1 (SS = 909, SL = 249, LL = 39).
The two-wave comparison on the stability of association between 5-HTTLPR polymorphism and moral permissibility
Methods
Participants
In wave 2, 634 participants (451 females, mean age = 21.0 ± 1.3 years) from the original sample quit the study because they graduated (N = 180) or moved to other school districts (N = 454). The other 500 and 63 participants (372 females, mean age = 19.0 ± 1.6 years) who participated in the moral judgment assessment at wave 1 (October 2015 to June 2016) were invited to this study 6 months later (wave 2). As for the 5-HTTLPR polymorphism of this subsample, the genotype frequencies also deviated from Hardy–Weinberg equilibrium (SS = 441, SL = 102, LL = 20), χ2 = 17.84.67, P < 0.001. However, they had no significant deviation from those of the large sample in wave 1, χ2 = 1.729, P = 0.426.
Moral dilemma assessment
The same 16 moral dilemmas from wave 1 were used. To check whether the repeated assessment was confounded by the memory for how they previously responded to the harms, four non-moral dilemma scenarios (see in the Online Supplementary Materials) were organized in the scenarios. For the 16 moral scenarios, participants indicated the extent to which the proposed action of the protagonist was permissible (‘1’ = totally impermissible to ‘7’ = totally permissible). For the non-moral dilemma scenarios, participants indicated the extent to which they accepted the protagonist’s hypothetical option for the dilemma scenarios. After this, the participants received a post-test that consists of two non-moral dilemmas (new items) and two moral dilemmas (old items). The participants were asked to judge whether they have seen the four dilemma scenarios before 6 months. If the participants believed that they had seen it, they marked the scenario as an old item; otherwise, they marked it as a new item. To assess the accuracy of their memory of moral scenarios in wave 1, a comparison analysis with Wilcoxon signed-rank test indicated no significant difference in the ratios of reporting new (the ratio of marked non-moral dilemmas as new vs the ratio of marked moral dilemmas as new items) between the two types of scenarios (non-moral dilemmas vs moral dilemmas = 0.47 ± 0.31 vs 0.45 ± 0.30), Z = −0.929, P = 0.353. The results partially indicated that the participants did not have a good memory of moral scenarios from wave 1. The internal consistencies (Cronbach’s α) of moral scenarios were 0.864 and 0.735 at wave 1 and wave 2, respectively.
Result
In this subsample, the permissibility ratings of moral judgment at wave 1 and wave 2 were significantly correlated (impersonal action: r = 0.39, P < 0.001; impersonal inaction: r = 0.41, P < 0.001; personal action: r = 0.37, P < 0.001; impersonal action: r = 0.35, P < 0.001), suggesting that moral permissibility was stable across time. To compare the permissibility ratings between the two time windows and examine the impacts of dilemma types, frame of actions and genotypes, we conducted a 2 (time windows: wave 1 vs wave 2) × 2 (dilemma types: impersonal vs personal) × 2 (frame of actions: action vs inaction) × 3 (genotypes: SS vs SL vs LL) ANOVA analysis. The results indicated that significant impacts of time windows (mean ± SEM: wave 1 vs wave 2 = 3.46 ± 0.08 vs 3.90 ± 0.07), F(1, 563) = 13.049,
P < 0.001, 2 = 0.023; dilemma types, F(1, 563) = 159.937, P < 0.001, 2 = 0.22; frame of actions, F(1, 563) = 129.930, P < 0.001, 2 = 0.19; genotypes, F(2, 563) = 8.937, P < 0.001, 2 = 0.031; and a four-way interaction between the variables, F(2, 1196) = 3.07, P = 0.047, 2 = 0.005. With regard to the impact of the genotypes, we found that individuals with the SS genotype (mean ± SEM: 3.83 ± .04, N = 441) were more likely to rate harms as permissibility, compared with the ones with the SL carriers (mean ± SEM: 3.49 ± 0.08, N = 102), P < 0.001, Cohen’s d = 0.27. In contrast, no significant differences in the permissibility ratings between the SL and LL genotype groups were found (mean ± SEM: 3.49 ± 0.18, N = 20), P = 0.98, Cohen’s d < 0.001, and between the SS and LL genotype groups, P = 0.064, Cohen’s d = 0.27.
According to the significant four-way interaction between the above-mentioned variables, we further examined the impacts of the 5-HTTLPR polymorphism on different experimental conditions through a two-wave compared analysis. The analysis indicated that the association between this polymorphism and permissibility ratings of personal harms was not stable from wave 1 to wave 2 (Figure 2). In contrast, the associations between this polymorphism and permissibility ratings of impersonal actions and impersonal inactions were stable from wave 1 to wave 2 (Figure 2). The post hoc power analysis indicated that the sample of 563 participants had 97.12% probability of reaching significance (two-tailed α = 0.05) when the partial 2 reached 0.031(i.e. effect size f = 0.140).
Fig. 2.
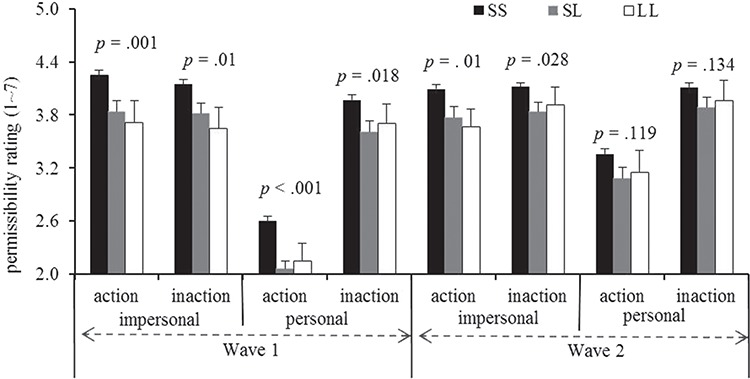
The effects of 5-HTTLPR polymorphism on moral permissibility (mean ± SEM) in the subsample (SS = 441, SL = 102, LL = 20) in wave 1 and wave 2.
Discussion
This study examined the association of 5-HTTLPR polymorphism with moral judgment in a relatively large cohort and compared the stability of this association in a subsample of the large cohort at two time windows. We found that the 5-HTTLPR polymorphism is associated with moral permissibility rating, e.g. the SS carriers rate impersonal harms as more permissible. Moreover, we found that such association is stable over time. Individuals with the SS genotype are likely to rate the impersonal harms as more permissible at two time windows. The findings provide robust evidence that the 5-HTTLPR polymorphism at individual level underpins moral judgment.
The previous study showed that serotonin administration with citalopram makes individuals judge personal harms as more impermissible (Crockett et al., 2010), through up-regulating an emotional aversion to prospective harms (Crockett et al., 2010; Tost & Meyer-Lindenberg, 2010). However, in the current study, we found that individuals with the SS genotype, corresponding to high serotonin levels in the synaptic cleft, rate impersonal harms as more permissible. Several differences between our study and Crockett’s study may explain the discrepancy. Firstly, the serotonin administration with citalopram and the genetic effect of the S allele of 5-HTTLPR may impact the permissibility rating of moral judgment in opposite directions. Crockett et al. found that increasing serotonin levels with citalopram is negatively related to the reduced permissibility rating. Our gene-association study indicated that an increasing number of the S allele, which is associated with high synaptic serotonin levels, is related to high permissibility rating. It is well-known that the 5-HTT is a key target of commonly prescribed selective serotonin reuptake inhibitors (Sanchez et al., 2004). Citalopram can boost synaptic serotonin levels by blocking the retaking function of 5-HTT, while the S allele of the 5-HTTLPR polymorphism is related to the low ratio of serotonin recycling by down-regulating the 5-HTT expression (Greenberg et al., 1999; Canli & Lesch, 2007). The opposite effect between citalopram and the S allele on moral permissibility rating may be attributed to their differential effects on the functional connectivity of amygdala during emotion processing (Outhred et al., 2016), such that the S allele is related to a decreased functional connectivity while escitalopram could enhance the functional connectivity (Outhred et al., 2016). Secondly, the increased serotonin levels induced by an acute citalopram administration can induce an interim prompting effect (about 1.5–2.0 h) on moral permissibility rating as the synaptic serotonin levels rise (Crockett et al., 2010), while our gene-association study reveals a long-term pre-disposition of the 5-HTTLPR polymorphism underlying moral permissibility rating. Thirdly, citalopram boosts moral permissibility rating only in cases with salient emotional aversion, while the 5-HTTLPR polymorphism is related to permissibility rating only for impersonal harms. Given that emotional aversion guides permissibility rating of personal harms while utilitarian influences the permissibility rating of impersonal action harms (Moll & de Oliveira-Souza, 2007; Gawronski et al., 2017), we infer that citalopram and the 5-HTTLPR polymorphism differentially impact moral permissibility ratings of personal harms and impersonal harms through different psychological regulations. Thus, these findings may suggest that escitalopram boosts permissibility rating by increasing emotional aversion and the S allele of the 5-HTTLPR regulates permissibility rating by reappraising the utilitarian.
The previous study indicated that the Caucasians with the SS genotype of 5-HTTLPR polymorphism rate impersonal harms as more impermissible (Marsh et al., 2011), while we found that the Asians with the SS genotype rate impersonal harms as more permissible. For Caucasians, the enhancement of the S allele on permissibility rating may attribute to the allele’s roles in heightening the amygdala activation (Dannlowski et al., 2010; Lonsdorf et al., 2014; Cheon et al., 2015; Kilpatrick et al., 2015; Christou et al., 2017). However, ethnic differences in the functions of S allele in amygdala activation should be noticed. For example, the S allele heightens the reactivity of the amygdala in Caucasians (Dannlowski et al., 2010; Lonsdorf et al., 2014; Cheon et al., 2015; Kilpatrick et al., 2015; Christou et al., 2017), while this allele decreases the reactivity of the amygdala in Asians (Lee & Ham, 2008; Li et al., 2012; Long et al., 2014). These findings may suggest that Asians with the S allele may show lower emotional aversion to harms than Caucasians with this allele. Therefore, the ethnic difference in the amygdala activation may contribute to the different effects of the S allele on moral judgment of Caucasians and Asians. Moreover, there are significant differences in allele frequency of 5-HTTLPR among different races, with the highest frequency in Asians, followed by Amerindian populations (Esau et al., 2008; Haberstick et al., 2015). The ethnic differences in the 5-HTTLPR also contribute to the divergent phenotypes of psychiatric disorders, personality traits and social behaviors of people from different races (Noskova et al., 2008). We found that the genotype frequencies in the larger Asian sample significantly differed from the Caucasian sample from the previous study (Marsh et al., 2011). According to a cross-cultural psychological study showing that cultural variation may be a result of gene-brain interplay (Minkov et al., 2015), the prevalence of the 5-HTTLPR among races may underpin the ethnic and cultural differences in moral behaviors (Baartman et al., 1999; Moon, 1986). Thus, the new findings extend our understanding of the genetic basis of moral judgment. Additionally, as compared with the previous study (Marsh et al., 2011), we examined the effect of frame of action as well as dilemma type in a larger population. The manipulation of dilemma type and frame of action allows us to investigate the association between 5-HTTLPR and permissibility rating in different moral contexts.
Permissibility rating of moral judgment is guided by one’s utilitarian and deontology (Moll & de Oliveira-Souza, 2007; Gawronski et al., 2017). The utilitarian principle addresses the consequences of harmful action, while the deontology principle emphasizes one’s duties and universal obligations to others. For the outcome-focused moral judgment, individuals tend to be concerned about the norm of saving more people by sacrificing one person and thus rate impersonal action harm as less norm violation (Greene et al., 2001). With regard to the roles of 5-HTTLPR in moral behaviors, the S allele of the 5-HTTLPR has been related to the high permissibility for unfair dictator assignments of the Dictator Game (Enge et al., 2017), which suggests that the S allele could promote permissibility of preserving one’s well-being by sacrificing other’s interests. Here, we found that individuals with the SS genotype are more likely to rate impersonal action harms as permissibility. Thus, given the link of the S allele with high permissibility for dictator assignments (Enge et al., 2017), the roles of the S allele 5-HTTLPR in increasing permissibility of impersonal harms may be attributed to its effect on decreasing injustice sense of utilitarian outcomes.
A central rationale for this study is that serotonin levels (and thus serotonin regulation via the 5-HTTLPR polymorphism) may shape moral judgment through emotional aversion to the idea of causing harm. This hypothesis is derived from the dual-process theory of moral judgment (Greene et al., 2001). The theory states that the rejection of harms is rooted in an emotional aversion to the idea of causing harm. Harms caused by personal actions are judged as more impermissible than equivalent harms from impersonal action (Greene et al., 2001). Here, we found that the 5-HTTLPR polymorphism is related to the moral permissibility of impersonal harms, but not personal harms. The selective genetic effect may be due to the differential emotional aversions of personal harms and impersonal harms. In cases of personal harms, the strong emotional aversion causes individuals to consider the harmful actions as impermissible (Cushman et al., 2006; Walter et al., 2012), regardless of the allele at the individual level (Gong et al., 2017). For the impersonal harms, however, the blunted emotional aversion to conflict between harmful outcome and innocent intention (Moll & de Oliveira-Souza, 2007; Gawronski et al., 2017) could be differentiated according to genotypes at the individual level. In this case, the effect of the S allele could be observed.
We found that the association between the 5-HTTLPR polymorphism and moral permissibility is stable over time. Individuals with the SS genotype are more likely to rate impersonal action harms as permissible from wave 1 to wave 2. Given that the association of 5-HTTLPR polymorphism with impersonal inaction harms in the subsample was not replicated in the larger population, we cautiously conclude that the 5-HTTLPR polymorphism is associated with moral permissibility rating only for impersonal action harms. Therefore, these findings suggest that the link between 5-HTTLPR polymorphism and moral judgment remains stable across time. Moreover, the patterns of permissibility ratings for different moral dilemma contexts were stable, although the permissibility ratings significantly fluctuated within the 6-month interval. However, the results of the subsample at wave 1 did not match the results from larger sample though the general patterns remained same, which is partly attributed to the non-random sampling of the subsample from the larger sample.
Several limitations should be noticed. Firstly, we adopted the same hypothetical moral dilemmas to assess the stability of moral judgment across time. Although the non-moral scenarios were added in the assessment of wave 2, retention of previous responses could not be completely ruled out. Secondly, the deviation from Hardy–Weinberg equilibrium of the 5-HTTLPR was present in the samples. Given that the comparison analyses were carried out with factors manipulated within participants, the deviation was balanced across experimental conditions. However, the deviation could potentially limit our findings to be extended to other populations. Finally, the theme of death for the moral scenarios could effectively induce one’s emotional aversion to harms. However, it has reduced the generalization of our findings to other moral scenarios with less serious harms.
Conclusion
The current study demonstrates that the 5-HTTLPR polymorphism is associated with individual differences in moral judgment over time. These findings highlight the importance of the 5-HTTLPR polymorphism to moral judgment.
Supplementary Material
Acknowledgments
This study was supported by the Ministry of Education of Humanities and Social Science Foundation (19YJAZH021) to P.G.
References
- Baartman H.E.M., Mey d.J.R.P.B., Schulze H.J. (1999). Ethnic variation and the development of moral judgment of youth in Dutch society. Youth & Society, 31(1), 54–75. doi: 10.1177/0044118X99031001003. [DOI] [Google Scholar]
- Bernhard R. M., Chaponis J., Siburian R., Gallagher P., Ransohoff K., Wikler D., . . . Greene J. D. (2016). Variation in the oxytocin receptor gene (OXTR) is associated with differences in moral judgment. Social Cognitive and Affective Neuroscience, 11(12), 1872–81. doi: 10.1093/scan/nsw103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canli T., Lesch K.P. (2007). Long story short: the serotonin transporter in emotion regulation and social cognition. Nature Neuroscience, 10(9), 1103–9. doi: 10.1038/nn1964. [DOI] [PubMed] [Google Scholar]
- Cheon B.K., Livingston R.W., Chiao J.Y., Hong Y.Y. (2015). Contribution of serotonin transporter polymorphism (5-HTTLPR) to automatic racial bias. Personality and Individual Differences, 79, 35–8. doi: 10.1016/j.paid.2015.01.019. [DOI] [Google Scholar]
- Christou A.I., Wallis Y., Bair H., Zeegers M., McCleery J.P. (2017). Serotonin 5-HTTLPR genotype modulates reactive visual scanning of social and non-social affective stimuli in young children. Frontiers in Behavioral Neuroscience, 11–2. doi: 10.3389/fnbeh.2017.00118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crockett M.J., Clark L., Hauser M.D., Robbins T.W. (2010). Serotonin selectively influences moral judgment and behavior through effects on harm aversion. Proceedings of the National Academy of Sciences of the United States of America, 107(40), 17433–8. doi: 10.1073/pnas.1009396107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cushman F., Young L., Hauser M. (2006). The role of conscious reasoning and intuition in moral judgment: testing three principles of harm. Psychological Science, 17(12), 1082–9. doi: 10.1111/j.1467-9280.2006.01834.x. [DOI] [PubMed] [Google Scholar]
- Dannlowski U., Konrad C., Kugel H., et al. (2010). Emotion specific modulation of automatic amygdala responses by 5-HTTLPR genotype. NeuroImage, 53(3), 893–8. doi: 10.1016/j.neuroimage.2009.11.073. [DOI] [PubMed] [Google Scholar]
- De Felice L J. (2016). A current view of serotonin transporters. F1000Research, 5. Doi: 10.12688/f1000research.8384.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding W., Shao Y.H., Sun B.G., Xie R.B., Li W.J., Wang X.Z. (2018). How can prosocial behavior be motivated? The different roles of moral judgment, moral elevation, and moral identity among the young Chinese. Frontiers in Psychology, 9, 10. doi: 10.3389/fpsyg.2018.00814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enge S., Mothes H., Fleischhauer M., Reif A., Strobel A. (2017). Genetic variation of dopamine and serotonin function modulates the feedback-related negativity during altruistic punishment. Scientific Reports, 7, 12. doi: 10.1038/s41598-017-02594-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esau L., Kaur M., Adonis L., Arieff Z. (2008). The 5-HTTLPR polymorphism in South African healthy populations: a global comparison. Journal of Neural Transmission (Vienna), 115(5), 755–60. doi: 10.1007/s00702-007-0012-5. [DOI] [PubMed] [Google Scholar]
- Faul F., Erdfelder E., Lang A.G., Buchner A. (2007). G*power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–91. doi: 10.3758/bf03193146. [DOI] [PubMed] [Google Scholar]
- Gawronski B., Armstrong J., Conway P., Friesdorf R., Huetter M. (2017). Consequences, norms, and generalized inaction in moral dilemmas: the CNI model of moral decision-making. Journal of Personality and Social Psychology, 113(3), 343–76. doi: 10.1037/pspa0000086. [DOI] [PubMed] [Google Scholar]
- Gleichgerrcht E., Torralva T., Rattazzi A., Marenco V., Roca M., Manes F. (2013). Selective impairment of cognitive empathy for moral judgment in adults with high functioning autism. Social Cognitive and Affective Neuroscience, 8(7), 780–8. doi: 10.1093/scan/nss067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong P., Fang P., Yang X., Ru W., Wang B., Gao X., Liu J. (2017). The CAG polymorphism in androgen receptor (AR) gene impacts the moral permissibility of harmful behavior in females. Psychoneuroendocrinology, 80, 74–9. doi: 10.1016/j.psyneuen.2017.03.008. [DOI] [PubMed] [Google Scholar]
- Greenberg B.D., Tolliver T.J., Huang S.J., Li Q., Bengel D., Murphy D.L. (1999). Genetic variation in the serotonin transporter promoter region affects serotonin uptake in human blood platelets. American Journal of Medical Genetics, 88(1), 83–7. doi: 10.1002/(sici)1096-8628(19990205)88:1<83::aid-ajmg15>3.0.co;2-0. [DOI] [PubMed] [Google Scholar]
- Greene J.D., Sommerville R.B., Nystrom L.E., Darley J.M., Cohen J.D. (2001). An fMRI investigation of emotional engagement in moral judgment. Science, 293(5537), 2105–8. doi: 10.1126/science.1062872. [DOI] [PubMed] [Google Scholar]
- Gutierrez B., Pintor L., Gasto C., Rosa A., Bertranpetit J., Vieta E., Fananas L. (1998). Variability in the serotonin transporter gene and increased risk for major depression with melancholia. Human Genetics, 103(3), 319–22. doi: 10.1007/s004390050823. [DOI] [PubMed] [Google Scholar]
- Haberstick B.C., Smolen A., Williams R.B., et al. (2015). Population frequencies of the triallelic 5HTTLPR in six ethnicially diverse samples from North America, Southeast Asia, and Africa. Behavior Genetics, 45, 255–61. doi: 10.1007/s10519-014-9703-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hannikainen I.R., Machery E., Cushman F.A. (2018). Is utilitarian sacrifice becoming more morally permissible? Cognition, 170, 95–101. doi: 10.1016/j.cognition.2017.09.013. [DOI] [PubMed] [Google Scholar]
- Heiphetz L., Strohminger N., Gelman S.A., Young L.L. (2018). Who am I? The role of moral beliefs in children’s and adults’ understanding of identity. Journal of Experimental Social Psychology, 78, 210–9. doi: 10.1016/j.jesp.2018.03.007. [DOI] [Google Scholar]
- Helzer E.G., Fleeson W., Furr R.M., Meindl P., Barranti M. (2017). Once a utilitarian, consistently a utilitarian? Examining principledness in moral judgment via the robustness of individual differences. Journal of Personality, 85, 505–17. doi: 10.1111/jopy.12256. [DOI] [PubMed] [Google Scholar]
- Jonassen R., Landro N.I. (2014). Serotonin transporter polymorphisms (5-HTTLPR) in emotion processing: implications from current neurobiology. Progress in Neurobiology, 117, 41–53. doi: 10.1016/j.pneurobio.2014.02.003. [DOI] [PubMed] [Google Scholar]
- Kahane G., Everett J.A., Earp B.D., Farias M., Savulescu J. (2015). ‘Utilitarian’ judgments in sacrificial moral dilemmas do not reflect impartial concern for the greater good. Cognition, 134, 193–209. doi: 10.1016/j.cognition.2014.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilpatrick L. A., Mayer E. A., Labus J. S., Gupta A., Hamaguchi T., Mizuno T., . . . Fukudo S. (2015). Serotonin transporter gene polymorphism modulates activity and connectivity within an emotional arousal network of healthy men during an aversive visceral stimulus. PLoS One, 10(4), 15. doi: 10.1371/journal.pone.0123183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenigs M., Kruepke M., Zeier J., Newman J.P. (2012). Utilitarian moral judgment in psychopathy. Social Cognitive and Affective Neuroscience, 7(6), 708–14. doi: 10.1093/scan/nsr048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohen R., Cain K. C., Mitchell P. H., Becker K., Buzaitis A., Millard S. P., . . . Veith R. (2008). Association of serotonin transporter gene polymorphisms with poststroke depression. Archives of General Psychiatry, 65(11), 1296–302. doi: 10.1001/archpsyc.65.11.1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Lamballerie X., Chapel F., Vignoli C., Zandotti C. (1994). Improved current methods for amplification of DNA from routinely processed liver tissue by PCR. Journal of Clinical Pathology, 47(5), 466–7. doi: 10.1136/jcp.47.5.466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee B.T., Ham B.J. (2008). Serotonergic genes and amygdala activity in response to negative affective facial stimuli in Korean women. Genes, Brain, and Behavior, 7(8), 899–905. doi: 10.1111/j.1601-183X.2008.00429.x. [DOI] [PubMed] [Google Scholar]
- Li S., Zou Q., Li J., Li J., Wang D., Yan C., . . . Zang Y. F. (2012). 5-HTTLPR polymorphism impacts task-evoked and resting-state activities of the amygdala in Han Chinese. PLoS One, 7(5), e36513. doi: 10.1371/journal.pone.0036513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J., Hao J., Shi B.G. (2018). From moral judgments to prosocial behavior: multiple pathways in adolescents and different pathways in boys and girls. Personality and Individual Differences, 134, 149–54. doi: 10.1016/j.paid.2018.06.017. [DOI] [Google Scholar]
- Long H.X., Liu B., Hou B., Wang C., Kendrick K.M., Yu C.S., Jiang T.Z. (2014). A potential ethnic difference in the association between 5-HTTLPR polymorphisms and the brain default mode network. Chinese Science Bulletin, 59(13), 1355–61. doi: 10.1007/s11434-014-0185-x. [DOI] [Google Scholar]
- Lonsdorf T.B., Juth P., Rohde C., Schalling M., Ohman A. (2014). Attention biases and habituation of attention biases are associated with 5-HTTLPR and COMTval158met. Cognitive, Affective, & Behavioral Neuroscience, 14(1), 354–63. doi: 10.3758/s13415-013-0200-8. [DOI] [PubMed] [Google Scholar]
- Marsh A.A., Crowe S.L., Yu H.H., Gorodetsky E.K., Goldman D., Blair R.J. (2011). Serotonin transporter genotype (5-HTTLPR) predicts utilitarian moral judgments. PLoS One, 6(10), e25148. doi: 10.1371/journal.pone.0025148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald M.M., Defever A.M., Navarrete C.D. (2017). Killing for the greater good: action aversion and the emotional inhibition of harm in moral dilemmas. Evolution and Human Behavior, 38(6), 770–8. doi: 10.1016/j.evolhumbehav.2017.06.001. [DOI] [Google Scholar]
- Minkov M., Blagoev V., Bond M.H. (2015). Improving research in the emerging field of cross-cultural sociogenetics: the case of serotonin. Journal of Cross-Cultural Psychology, 46(3), 336–54. doi: 10.1177/0022022114563612. [DOI] [Google Scholar]
- Moll J., Oliveira-Souza R. (2007). Moral judgments, emotions and the utilitarian brain. Trends in Cognitive Sciences, 11(8), 319–21. doi: 10.1016/j.tics.2007.06.001. [DOI] [PubMed] [Google Scholar]
- Moon Y.L. (1986). A review of cross-cultural studies on moral judgment development using the defining issues test. Cross-Cultural Research, 20(1–4), 147–77. doi: 10.1177/106939718602000107. [DOI] [Google Scholar]
- Noskova T., Pivac N., Nedic G., et al. (2008). Ethnic differences in the serotonin transporter polymorphism (5-HTTLPR) in several European populations. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 32(7), 1735–9. doi: 10.1016/j.pnpbp.2008.07.012. [DOI] [PubMed] [Google Scholar]
- Outhred T., Das P., Dobson-Stone C., et al. (2016). Impact of 5-HTTLPR on SSRI serotonin transporter blockade during emotion regulation: a preliminary fMRI study. Journal of Affective Disorders, 196, 11–9. doi: 10.1016/j.jad.2016.02.019. [DOI] [PubMed] [Google Scholar]
- Patil I. (2015). Trait psychopathy and utilitarian moral judgement: the mediating role of action aversion. Journal of Cognitive Psychology, 27(3), 349–66. doi: 10.1080/20445911.2015.1004334. [DOI] [Google Scholar]
- Perez-Edgar K., Bar-Haim Y., McDermott J. M., Gorodetsky E., Hodgkinson C. A., Goldman D., . . . Fox N. A. (2010). Variations in the serotonin-transporter gene are associated with attention bias patterns to positive and negative emotion faces. Biological Psychology, 83(3), 269–71. doi: 10.1016/j.biopsycho.2009.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raab K., Kirsch P., Mier D. (2016). Understanding the impact of 5-HTTLPR, antidepressants, and acute tryptophan depletion on brain activation during facial emotion processing: a review of the imaging literature. Neuroscience and Biobehavioral Reviews, 71, 176–97. doi: 10.1016/j.neubiorev.2016.08.031. [DOI] [PubMed] [Google Scholar]
- Ru W.Z., Fang P.P., Wang B., et al. (2017). The impacts of Val158Met in catechol-O-methyltransferase (COMT) gene on moral permissibility and empathic concern. Personality and Individual Differences, 106, 52–6. doi: 10.1016/j.paid.2016.10.041. [DOI] [Google Scholar]
- Sanchez C., Bogeso K.P., Ebert B., Reines E.H., Braestrup C. (2004). Escitalopram versus citalopram: the surprising role of the R-enantiomer. Psychopharmacology, 174(2), 163–76. doi: 10.1007/s00213-004-1865-z. [DOI] [PubMed] [Google Scholar]
- Siegel J.Z., Crockett M.J. (2013). How serotonin shapes moral judgment and behavior. Sociability, Responsibility, and Criminality: From Lab to Law, 1299, 42–51. doi: 10.1111/nyas.12229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talvenheimo J., Nelson P.J., Rudnick G. (1979). Mechanism of imipramine inhibition of platelet 5-hydroxytryptamine transport. The Journal of Biological Chemistry, 254(11), 4631–5. [PubMed] [Google Scholar]
- Tost H., Meyer-Lindenberg A. (2010). I fear for you: a role for serotonin in moral behavior. Proceedings of the National Academy of Sciences of the United States of America., 107(40), 17071–2. doi: 10.1073/pnas.1012545107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter N.T., Montag C., Markett S., Felten A., Voigt G., Reuter M. (2012). Ignorance is no excuse: moral judgments are influenced by a genetic variation on the oxytocin receptor gene. Brain and Cognition, 78(3), 268–73. doi: 10.1016/j.bandc.2012.01.003. [DOI] [PubMed] [Google Scholar]
- Yang Y., Li X., Zhao J., et al. (2019). 5-HTTLPR and COMT Val158Met are not associated with alexithymia: new evidence and meta-analyses. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 92, 263–70. doi: 10.1016/j.pnpbp.2019.01.015. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.