

Abstract
Kinesin-3 and kinesin-1 molecular motors are two families of the kinesin superfamily. It has been experimentally revealed that in monomeric state kinesin-3 is inactive in motility and cargo-mediated dimerization results in superprocessive motion, with an average run length being more than 10-fold longer than that of kinesin-1. In contrast to kinesin-1 showing normally single-exponential distribution of run lengths, dimerized kinesin-3 shows puzzlingly Gaussian distribution of run lengths. Here, based on our proposed model, we studied computationally the dynamics of kinesin-3 and compared with that of kinesin-1, explaining quantitatively the available experimental data and revealing the origin of superprocessivity and Gaussian run length distribution of kinesin-3. Moreover, predicted results are provided on ATP-concentration dependence of run length distribution and force dependence of mean run length and dissociation rate of kinesin-3.
Subject terms: Biophysics, Computational biophysics
Introduction
Kinesins are a large superfamily of motor proteins capable of moving on microtubule (MT) filaments by hydrolyzing ATP1–3. They are responsible for a variety of biological functions in cell such as intracellular cargo transport, chromosome segregation, spindle assembly, cytoskeletal organization, etc.3. Among the kinesin superfamily, kinesin-3 motors constitute one of the largest families4,5, which are involved in functions of vesicle transport6–12, signaling13,14, mitosis15,16, interkinetic nuclear migration in neural stem cells17, transport of alphaherpesvirus particles in neurons18,19, early embryonic development20, and so on.
While a lot of researches have focused on studies of dynamics of kinesin-1 motors, the dynamics of kinesin-3 motors has also been studied insightfully5,21,22. Recently, using Forster resonance energy transfer microscopy in live cells, Soppina et al.23 showed that while in monomeric state kinesin-3 is inactive in motility, the dimerization results in superprocessive motion, with an average run length of ~ 10 μm, which is more than 10-fold longer than that of kinesin-1. Considering that under no load the kinesin dimer moves on average with a constant velocity and during the processive movement there is a constant rate of dissociation from MT, a single-exponential or nearly single-exponential distribution of run lengths would be expected24,25, as the available experimental data showed for kinesin-123,26,27, kinesin-4 KIF21B28, kinesin-7 CENP-E29, kinesin-12 Kif1530, and so on. However, it was found surprisingly that the dimerized kinesin-3 has approximately a Gaussian-form distribution of run lengths, which is deviated significantly from the single-exponential form23. Furthermore, Scarabelli et al.31 found that substitutions of prominent kinesin-3 residues that contribute to interaction energy with MT decrease kinesin-3 processivity by about 10-fold, thus approaching kinesin-1 levels. However, while the family-specific K-loop provides kinesin-3 motors with a high MT affinity in ADP state of the motor, the mutation of the K-loop has little effect on the superprocessive motion of the dimeric motors32.
Despite the extensive studies, the mechanism of how kinesin-3 dimers show superprocessivity is unclear. Is the higher MT affinity sufficient to give the superprocessivity of kinesin-3 dimers? While the K-loop enhances the MT affinity, why do mutations of the K-loop have little effect on the superprocessivity of kinesin-3 dimers? More puzzlingly, why do kinesin-3 dimers have approximately a Gaussian form of run length distributions rather than a nearly single-exponential form? The purpose of this work is to address these unclear issues, which is critical to the molecular mechanism of the mechanochemical coupling of kinesin-3 dimers.
Methods
The model of processive movement of kinesin-1 and kinesin-3 dimers and simulation methods
The model for kinesin-1 has been proposed before33,34, based on which the available single-molecule experimental data on the force dependence of the mean velocity and run length of the wild-type kinesin-1 and the mutant one with extension of its two neck linkers27,35 were reproduced quantitatively. In particular, the puzzlingly dramatic asymmetry of the mean run length of kinesin-1 dimers with respect to the direction of external load acting on the coiled-coil stalk was explained well33,34. Here, we consider that kinesin-3 and kinesin-1 dimers share the same mechanism of processive movement on MT. The model for the pathway of a typical forward stepping and dissociation from MT of kinesin-3 and kinesin-1 dimers is schematically shown in Fig. 1, which is built up based mainly on the following pieces of experimental and computational evidence and/or arguments.
Figure 1.

Schematic of a typical forward stepping of dimeric kinesin-1 and kinesin-3. (a) The trailing head in ATP state binds strongly to binding site I on MT while the leading head in nucleotide-free (ϕ) state binds strongly to site II. The rate of ATP hydrolysis and Pi release of the trailing head with the forward orientation of the NL is much higher than that of the leading head without the forward orientation of the NL. (b) After ATP hydrolysis and Pi release in the trailing head, due to the very weak affinity (Ew1) between the ADP-head and the local site I, the ADP-head detaches easily from site I and diffuses rapidly to the INT position relative to the MT-bound head, where the two heads have a high affinity. (c) ATP binds to the MT-bound ϕ-head. (d) The NL docking of MT-bound head takes place, weakening the interaction between the two heads, and the tethered ADP-head then diffuses rapidly to the forward site III on MT. Note that the NL docking provides an energy barrier ENL to prevent the tethered ADP-head from moving backward. (e) Stimulated by MT, ADP is released from the leading head. State in (e) is the same as that in (a) except a forward step was made. (f) From (c), ATP hydrolysis and Pi release can also occur occasionally in the MT-bound head before its NL docking. Before the affinity of the MT-bound ADP-head for the local site II changes from Ew1 to Ew2 (called Period I), the dimer can detach easily from MT by overcoming the very weak affinity Ew1. (g) From (d), ATP hydrolysis and Pi release can also occur occasionally in the trailing head before ADP is released from the leading head. The dimer also has a large probability to detach from MT before ADP is released from the MT-bound head (called Period II) by overcoming weak affinity Ew2. (h) From (g), the dimer has not detached from MT until ADP is released from the MT-bound head. State in (h) is the same as that in (b) except a forward step was made. Inset (a’ – e’) is the schematic illustration of the intermolecular interaction between two NC segments and intramolecular interaction of N-residues with the head during cargo-mediated dimerization of kinesin-3. (a’) Before cargo-mediated dimerization, the intramolecular interaction of the N-residues with head inhibits its ATPase activity. (b’,c’) After cargo binding, firstly the C-terminal residue (adjacent to the cargo) in one NC helix forms an intermolecular interaction with the corresponding residue in another NC helix. Then, the intermolecular interactions between residues in the two helixes form successively. Before formation of the intermolecular interaction between N-residues in the two helixes, in the state with two heads bound to MT, the stretching of the two NLs causes the N-residues to be away from their heads, with no intramolecular interaction between the N-residues and head. The two heads thus have the ATPase activity (b’). In INT state, the N-residues are close to the head and the intramolecular interaction between them is present, inhibiting the ATPase activity of the MT-bound head (c’). (d’, e’) After complete formation of the coiled-coil between two NC helixes, the formation of the intermolecular interaction between N-residues in the two helixes prevents the intramolecular interaction between the N-residues and MT-bound head in both the state with the two heads bound to MT (d’) and INT state (e’). Thus, in both states the MT-bound head has the ATPase activity.
(i) A kinesin head in nucleotide-free (ϕ), ATP or ADP.Pi state has a high binding affinity to MT, whereas in ADP state has a low affinity (denoted by Ew2), as available experimental data showed36–40. Moreover, it is argued that upon Pi release the binding energy of the ADP-head with the local binding site on MT (denoted by Ew1) is temporarily weaker than that with other unperturbed binding sites (i.e., Ew1 < Ew2)41–43, which is supported by recent all-atom molecular dynamics (MD) simulations44. This can be understood as follows. The large conformational changes in the local MT-tubulin heterodimer induced by the strong interaction with ϕ-, ATP- or ADP.Pi-head45 result in the local tubulin having a further weaker interaction with the ADP-head than other unperturbed tubulins. In a time of tr the local tubulin relaxes to its normally unperturbed conformation, with the interaction energy of the local tubulin with the ADP-head becoming the same as other tubulins. Thus, the interaction potential between the kinesin head and MT along an MT protofilament in an ATPase cycle can be approximately shown in Fig. S1 (see Supplementary Information) and the mathematical description of the interaction potential is presented in Section S1 (see Supplementary Information).
(ii) When an MT-bound head is in ATP or ADP.Pi state there is a small free energy (denoted by ENL) to facilitate its neck linker (NL) docking into its motor domain46. The NL docking involves NL strand β9 and strand β0 of the motor domain forming a cover-neck bundle47 and only when β0 and β9 are in proximity can the cover-neck bundle formation take place. When an MT-bound kinesin head is in ϕ or ADP state, its NL is unable to dock46. In Section S2 (see Supplementary Information) we give mathematical description of the potential characterizing the effect of NL docking of the MT-bound head on the movement of the other tethered ADP-head.
(iii) An interaction is present between the two heads. When NL of the MT-bound head is undocked, a high binding energy (denoted by EI1) is present between the MT-bound head and detached ADP-head. When NL of the MT-bound head is docked, the binding energy (denoted by EI2) is reduced greatly, as argued before48,49 and supported by recent all-atom molecular dynamics simulations50. In Section S3 (see Supplementary Information) we give mathematical description of the potential of the interaction between the two heads. Note that with this argument and argument (ii), the biochemical data showing that upon the kinesin-1 dimer with both heads bound by ADP mixing with MT only half fraction of ADP molecules are released and addition of ATP molecules leads to release of other half fraction of ADP molecules51 can be explained well49.
As done before33,34, for simplicity of analysis, we do not consider the dissociation of the ϕ- or ATP- or ADP.Pi-head from MT under no or a small load, because in these states the kinesin head binds to MT strongly. Based on the model, the movement of tethered ADP-head relative to the other ϕ-, ATP- or ADP.Pi-head bound fixedly to MT can be described by Eqs. (S6) – (S11) (see Section S5 in Supplementary Information). When both ADP-heads are bound simultaneously to MT, the movement of one head relative to the other MT-bound head can still be described by Eqs. (S6) – (S11). When the two ADP-heads are bound together strongly, the movement of the MT-bound ADP-head relative to MT can be described by Eqs. (S12) – (S17) (see Section S6 in Supplementary Information).
Using Eqs. (S6) – (S17) we can simulate the mechanical motion of a kinesin head following Pi release relative to the other MT-bound kinesin head and the dissociation of the dimer from MT by using stochastic Runge–Kutta algorithm, as done before33,34. Then, we can simulate processive movement of the dimer by also considering continuous ATPase activities, which can be simulated using Monte-Carlo algorithm (see Section S7 in Supplementary Information).
As mentioned above, we consider that when one head is in ϕ, ATP or ADP.Pi state, the binding energy of the head to MT is so large that the dissociation of the head from the MT is negligibly small under no or a small load. This implies that only when both heads are in ADP state the dissociation of the motor is considered. In the calculations, when both heads move to positions that are away from the MT filament by more than 10 nm, the motor is considered to dissociate from MT33,34.
From a simulated trace of the displacement of the center of mass of the dimer versus time, the total displacement (or run length) of the trace can be obtained and the velocity of the trace can be calculated by dividing the total displacement by the total time before dissociation33,34. The mean run length and mean velocity are computed statistically using about 500 simulated traces.
Intermolecular interaction between two neck coil segments and intramolecular interaction of residues near the N-terminus of neck coil segment with the head during cargo-mediated dimerization of kinesin-3
Before cargo binding, we make following argument for kinesin-3. The intramolecular interaction of residues near the N-terminus of neck coil (NC) segment (called N-residues) with the head inhibits its ATPase activity (concretely, ATP hydrolysis and Pi release), with the ATPase rate equal to 0 (Fig. 1a’). In other words, in the monomeric form kinesin-3 is inactive in the ATPase activity, which is consistent with the experimental data showing that kinesin-3 monomer, although can bind to MT, is inactive in motility23.
After cargo binding, we make following argument. Firstly, the C-terminal residue (adjacent to the cargo) in one NC helix forms an intermolecular interaction with the corresponding residue in another NC helix (Fig. 1b’ and c’). Then, the intermolecular interactions between residues in the two helixes form successively. Thus, the coiled-coil formation between two NC helixes is composed of multiple (n > 1) rate-limiting steps, with each rate-limiting step corresponding to the formation of the intermolecular interaction between a pair of residues in the two helixes. This implies that the cargo-mediated dimerization process, which begins with the formation of the intermolecular interaction between two C-terminal residues in two NC helixes and ends with the formation of the intermolecular interaction between N-residues in the two helixes, is composed of multiple rate-limiting steps. Moreover, we have the following expectation of the intramolecular interaction between the N-residues and head during the period after formation of the intermolecular interaction between two C-terminal residues in two NC helixes and before formation of the intermolecular interaction between N-residues in the two helixes. In the state with two heads bound to an MT filament, the stretching of the two NLs causes the N-residues to be away from their heads (Fig. 1b’). Thus, there is no intramolecular interaction between the N-residues and head, the two heads having the ATPase activity (Fig. 1b’). In the intermediate (INT) state, the N-residues are close to the head (Fig. 1c’). Thus, the intramolecular interaction between them is present, inhibiting the ATPase activity of the MT-bound head (Fig. 1c’). After formation of the stable coiled-coil between two NC helixes (Fig. 1d’ and e’), the formation of the intermolecular interaction between N-residues in the two helixes prevents the intramolecular interaction between the N-residues and head. Thus, the MT-bound head has the ATPase activity in both INT state (Fig. 1e’) and state with two heads bound to MT (Fig. 1d’).
The choice of parameter values
In this work, we focus on two types of kinesin: kinesin-1 and kinesin-3. Their parameter values are chosen as follows.
Parameter values of kinesin-1
First, we consider kinesin-1. We take similar parameter values for kinesin-1 to those taken in our previous works33,34. We take tr = 10 μs and Ew2 = 39kBT. We take Ew1 ≤ 18kBT (varying value of Ew1 has little effect on our results provided Ew1 ≤ 18kBT). Provided EI1 ≥ 40kBT and EI2 ≤ 20kBT, varying values of EI1 and EI2 has little effect on our results. We take NL-docking energy ENL ≥ 4kBT, a small value that is consistent with the available experimental data46. In INT state, we take NL-docking rate kNL = 800 s−1 when the MT-bound head is in ATP or ADP.Pi state and kNL = 0 in ϕ or ADP state. This value of kNL = 800 s−1 is consistent with the biochemical data52. We take the rate of ATP hydrolysis and Pi release of the trailing head with the forward NL orientation, = kc = 140 , which is consistent with the biochemical data53. The rate of ATP hydrolysis and Pi release of the leading head without the forward NL orientation is taken to be = , with = 40. In INT state, the rate of ATP hydrolysis and Pi release of the MT-bound head without the forward NL orientation is also = . The fact that the head with the forward NL orientation has a much larger rate of ATP hydrolysis and Pi release than the head without the forward NL orientation can be understood as follows. With the forward orientation the NL can interact with the head and the interaction enhances the rate. This is consistent with the experimental data showing that by deleting NL (implying that the interaction of the NL with the head is removed) the ATPase rate is reduced greatly while the ADP-release rate is unaffected54. When ADP-head is bound to MT, the ADP-release rate kD = 350 , which is consistent with the biochemical data53. When ADP-head is detached from MT, without MT stimulation the ADP-release rate kD = 0. We take the second-order rate constant of ATP binding, kb = 2 , which is consistent with the biochemical data53. The above parameter values for kinesin-1 are listed in Table 1 (noting that some parameters whose values are not required to have definite values are not included).
Table 1.
Values of parameters for kinesin-1 and kinesin-3.
| Parameter | Value | |
|---|---|---|
| Kinesin-1 | Kinesin-3 | |
| tr (μs) | 10 | 10 |
| Ew1 | ≤18kBT | ≤18kBT |
| Ew2 | 39kBT | ≥45kBT |
| kNL (s−1) | 800 | 3400 |
| kc (s−1) (NL pointing forward) | 140 | 270 |
| kc(lead) (s−1) (NL not pointing forward) | kc/40 | kc/20 |
| kD (s−1) | 350 | 350 |
| kb (μM−1 s−1) | 2 | 2 |
| kcc (s−1) | — | 0.17 |
Symbol “–” denotes that the value is not required.
As mentioned above, values of parameters kNL, kc, kD and kb are consistent with the available biochemical data52,53, while values of other parameters tr, Ew1, Ew2 and are taken as done in our previous works33,34, with which the available single-molecule data on the force dependence of mean velocity and run length (in particular the dramatic asymmetry of mean run length with respect to the force direction) for kinesin-127,35 were reproduced quantitatively33,34.
Parameter values of kinesin-3
Then, we consider kinesin-3. Here, we focus mainly on mammalian kinesin-3 from subfamilies KIF1, KIF13, and KIF16, as used in the experiments by Verhey and his coworkers23,31,32. Since kinesin-3 head has a larger binding energy to MT than kiensin-1 head31,32, we take Ew2 ≥ 45kBT for kinesin-3, which is larger than that for kinesin-1.
Denoting by t = 0 the moment when the intermolecular interaction forms between two C-terminal residues in the two NC helixes and t = τ the moment when the intermolecular interaction forms between N-residues in the two helixes, the distribution of time τ has the Gamma form55
| 1 |
where n is the number of rate-limiting steps of the coiled-coil formation between two NC helixes and k0 is the rate constant of each rate-limiting step. Note here that for simplicity of treatment but without loss of generality, we take each rate-limiting step having a constant rate. Before τ, in the state with two heads bound to MT the rates of ATP hydrolysis and Pi release of the trailing and leading heads are kc and , respectively, and in INT state the rate of ATP hydrolysis and Pi release of the MT-bound head is 0. After τ, in the state with two heads bound to MT the rates of ATP hydrolysis and Pi release of the trailing and leading heads are kc and , respectively, and in INT state the rate of ATP hydrolysis and Pi release of the MT-bound head is . In the calculation, we take kc = 270 and = 20. The NL-docking rate is taken to be kNL = 3400 when the MT-bound head is in ATP or ADP.Pi state and kNL = 0 in or ADP state. We take n as a variable parameter, with k0 = nkcc and kcc = 0.17 , where 1/kcc corresponds to the mean time of complete formation of the coiled-coil between the two NC helixes.
Except for the above parameters, we take all other parameters for kinesin-3 having the same values as those for kinesin-1. For clarity, the parameter values for kinesin-3 are also listed in Table 1.
Results
Velocity and run length distributions of dimeric kinesin-1
First, we consider kinesin-1 dimer at saturating ATP (2 mM). In Fig. 2a,b (left panels) we show the calculated results for distribution of velocities and that of run lengths, respectively. For comparisons, in Fig. 2 (right panels) we show the corresponding experimental data of Soppina et al.23. The calculated and experimental data are in quantitative agreement with each other. The data show that the velocity distribution has approximately a Gaussian form and the run length distribution has a single-exponential form. The Gaussian distribution of velocities is understandable easily, because under no load the backward stepping rate is negligibly small compared with the forward stepping rate24. The single-exponential distribution of run lengths can be understood as follows.
Figure 2.

Results for dimeric kinesin-1 at saturating ATP. Left panels are data calculated with parameter values given in Table 1 for kinesin-1, while right panels are experimental data taken from Soppina et al.23. The mean values of the data are indicated. (a) Velocity distribution. Lines are Gaussian fits. (b) Run length distribution. Lines are single-exponential fits.
As noted from Fig. 1 and discussed before33,34, in a mechanochemical coupling cycle the dissociation of the motor from MT occurs mainly in two periods when the MT-bound head in INT state binds weakly to MT. One period (called Period I) is that when the MT-bound head in ADP state has an affinity Ew1 for MT, namely, the period from the moment when Pi is released from the MT-bound head through the moment when the affinity of the ADP-head for the local MT tubulin is changed from Ew1 to Ew2 (Fig. 1f, state shaded in green). The other period (called Period II) is that when the MT-bound head in ADP state has an affinity Ew2 for MT (Fig. 1g, state shaded in light red). The occurrence of Period I is determined by the rate of ATP hydrolysis and Pi release of the MT-bound head relative to that of its NL docking in INT state, while the occurrence of Period II is determined by the rate of ADP release from the leading head relative to the rate of ATP hydrolysis and Pi release of the trailing head. In Period I, due to the small affinity Ew1 for MT, the head has a very large (~ 100%) probability to dissociate within time tr. In Period II, with the weak affinity Ew2 for MT, the head also has a large (<100%) probability to dissociate before ADP release within time 1/kD. For the case of kinesin-1 with parameter values given in Table 1, both the dissociation occurring during Period I and that occurring during Period II have contributions to the overall dissociation, with the latter having a larger contribution than the former. Moreover, the dissociation probabilities in both periods have constant values in each step. Thus, the run length distribution is expected to have a single-exponential form.
Velocity and run length distributions of dimerized kinesin-3
Then, we consider kinesin-3 dimer at saturating ATP (2 mM). In Fig. 3 we show the calculated results of velocity distribution (Fig. 3a–e, upper panels) and run length distribution (Fig. 3f–j, lower panels) for different values of n. It is seen that as for the case of kinesin-1, the velocity distribution for kinesin-3 also has approximately a Gaussian form. Both the mean value of velocity and width of velocity distribution are independent of n. Interestingly, the run length distribution for kinesin-3 has approximately a Gaussian form rather than a single-exponential form. Although the mean value of run lengths is nearly independent of n, the width of run length distribution decreases with the increase of n. In particular, it is noted that when n = 10, the width of run length distribution is in good agreement with the experimental data23 (Fig. 4). This implies that there are about 10 rate-limiting steps for the coiled-coil formation between two NC helixes. Thus, in following calculations we fix n = 10.
Figure 3.

Results for dimerized kinesin-3 at saturating ATP. The data are calculated with parameter values given in Table 1 for kinesin-3 and different values of n. The mean values of the data are indicated. (a–e) Velocity distribution. Lines are Gaussian fits. (f–j) Run length distribution. Lines are Gaussian fits.
Figure 4.

Results for dimerized kinesin-3 at saturating ATP. Left panels are data calculated with parameter values given in Table 1 for kinesin-3 and n = 10, while right panels are experimental data for KIF13B(1–412ΔP) taken from Soppina et al.23. The mean values of the data are indicated. (a) Velocity distribution. Lines are Gaussian fits. (b) Run length distribution. Lines are Gaussian fits.
The approximate Gaussian distribution of run lengths can be understood as follows. In our model, the value of Ew2 determines only the probability of dissociation occurring during Period II. Our calculations show that the large value of Ew2 ≥ 45kBT gives the probability of dissociation occurring during Period II to be negligibly small. Thus, the run length for dimerized kinesin-3 is determined only by the dissociation occurring during Period I. Before the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state changes from 0 to , Period I cannot occur and thus nearly no dissociation can occur. After the rate of ATP hydrolysis and Pi release of the MT-bound head changes to at t = τ, Period I can occur. Once Period I occurring, the MT-bound head, which is bound strongly with the detached ADP-head, has nearly 100% probability to dissociate from the potential well of depth Ew1 (≤ 18kBT) within time tr. Thus, from the Gamma distribution of τ [Eq. (1)] an approximate Gaussian distribution of run lengths would be expected at large n. However, it is noted that since = 20 is much larger than 1, implying that the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state is much smaller than that of the trailing head, the change of the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state from 0 to has nearly no effect on velocity.
As mentioned above, our calculations show that provided that Ew2 ≥45kBT, changing Ew2 has little effect on our results of velocity and run length. This provides an explanation of the experimental data showing that although the K-loop enhances the binding affinity to MT, the mutation of the K-loop has little effect on the velocity and run length of kinesin-3 dimers32. This is understandable easily. For Ew2 ≥45kBT, the dissociation during Period II is negligibly small and thus changing value of Ew2 caused by the mutation of the K-loop has nearly no effect on the overall dissociation. On the other hand, provided that Ew1 ≤18kBT, once Period I occurring the MT-bound head has nearly 100% probability to dissociate from the potential well of depth Ew1 within time tr. Thus, the change of Ew1 caused by the mutation of the K-loop also has no effect on the dissociation.
To examine whether compared to kinesin-1 the enhancement of Ew2 is sufficient to give the superprocessivity of kinesin-3, we calculate the distribution of velocities and that of run lengths with parameter values given in Table 1 for kinesin-1 except with Ew2 ≥45kBT (Fig. 5). Comparing Fig. 5b with Fig. 2b and with Fig. 4b, it is seen that although the enhancement of Ew2 increases the mean run length by about 2.3-fold the mean run length is still about 4-fold smaller than that of kinesin-3. In addition, the run length distribution (Fig. 5b) still has a single-exponential form. This is because with Ew2 ≥45kBT although the dissociation occurring during Period II is negligibly small, the large dissociation probability during Period I, which can now occur at t = 0, prevents the dimers from becoming the supperprocessive motors. Thus, we conclude that compared to kinesin-1 the enhancement of Ew2 is insufficient to give the superprocessive motion of kinesin-3. The superprocessivity of kinesin-3 is due to both the slow transition of the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state from 0 to and the large value of Ew2.
Figure 5.
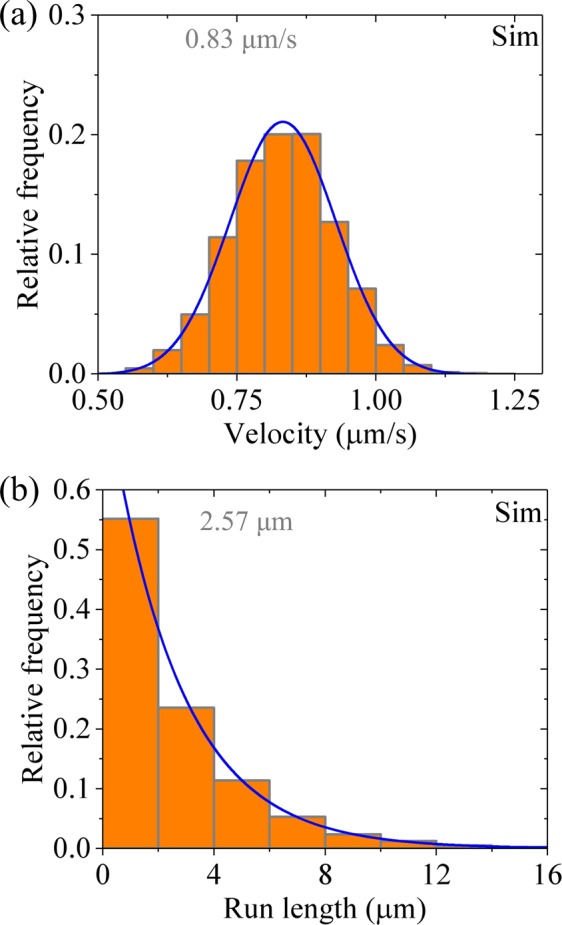
Results for dimeric kinesin-1 at saturating ATP. The data are calculated with parameter values given in Table 1 for kinesin-1 except for Ew2 ≥ 45kBT. The mean values of the data are indicated. (a) Velocity distribution. Line is the Gaussian fit. (b) Run length distribution. Line is the single-exponential fit.
To explain the experimental data on the effect of reducing Ew2 on run length of kinesin-3 by mutating residues that have large effects on binding affinity to MT31, we calculate the distribution of run lengths with Ew2 = 39kBT while with other parameter values being given in Table 1 for kinesin-3. The calculated results are shown in Fig. 6 (left panels). For comparisons, the experimental data for L8 mutant of KIF13B (ΔP391) in Scarabelli et al.31 are also shown in Fig. 6 (right panels). It is seen that the calculated data reproduce well the experimental data. That the run length distribution has a single-exponential form can be understood as follows. The value of Ew2 = 39kBT gives a large dissociation probability occurring during Period II, giving the dimer to dissociate from MT before the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state changes from 0 to . On the other hand, with the rate of ATP hydrolysis and Pi release equal to zero, no dissociation can occur during Period I and thus the dissociation arises solely from that occurring during Period II. Since in each step the probability of dissociation occurring during Period II is constant, it is expected that the run length distribution has a single-exponential form.
Figure 6.

Results for for dimerized kinesin-3 at saturating ATP. Left panels are data calculated with parameter values given in Table 1 for kinesin-3 and n = 10 except for Ew2 = 39kBT, while right panels are experimental data for L8 mutant of KIF13B (ΔP391) taken from Scarabelli et al.31. The mean values of the data are indicated. (a) Velocity distribution. Lines are Gaussian fits. (b) Run length distribution. Lines are single-exponential fits.
Now, we give an explanation of the experimental data on KIF1A(1–393) dimer showing only moderately processive with an average run length of ∼2.5 μm and a single-exponential distribution of run lengths. In KIF1A(1–393), since the NC segment could be unstable, no interaction between the NC and head would be present. Thus, we consider that when the cargo-mediated dimerization begins at t = 0, the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state has the non-zero value of . With this consideration and parameter values for kinesin-3 given in Table 1, we calculate the velocity and run length distributions, with the results being shown in Fig. 7 (left panels). For comparison, the corresponding experimental data for KIF1A(1–393)23 are shown in Fig. 7 (right panels). It is seen that the calculated data are in good agreement with the experimental data.
Figure 7.

Results for dimerized KIF1A(1–393) at saturating ATP. It is considered that at t = 0 the rate of ATP hydrolysis and Pi release of the MT-bound head in INT state has the non-zero value of . Left panels are data calculated with parameter values given in Table 1 for kinesin-3 and n = 10, while right panels are experimental data for KIF1A(1–393) taken from Soppina et al.23. The mean values of the data are indicated. (a) Velocity distribution. Lines are Gaussian fits. (b) Run length distribution. Lines are single-exponential fits.
Velocity and run length of dimeric kinesin-1 and kinesin-3 at low ATP concentrations
In the above, we made studies at saturating ATP, explaining quantitatively the available experimental data. In this section, we make studies at non-saturating ATP, providing some predicted results.
First, we consider kinesin-1 dimer. In Fig. 8 we show the calculated results of velocity distribution (Fig. 8a–e, upper panels) and run length distribution (Fig. 8f–j, lower panels) at different ATP concentrations. It is seen that at any ATP concentration, the velocity distribution has the Gaussian form and the run length distribution has the single-exponential form. The mean velocity increases with the increase of ATP concentration, and the mean run length is independent of ATP concentration, which is consistent with the available single-molecule experimental data27.
Figure 8.

Results for dimeric kinesin-1 at non-saturating ATP concentrations. The data are calculated with parameter values given in Table 1 for kinesin-1. The mean values of the data are indicated. (a–e) Velocity distribution. Lines are Gaussian fits. (f–j) Run length distribution. Lines are single-exponential fits.
Second, we consider kinesin-3 dimer. In Fig. 9 we show the calculated results of velocity distribution (Fig. 9a–e, upper panels) and run length distribution (Fig. 9f–j, lower panels) at different ATP concentrations. It is seen that at any ATP concentration, the velocity distribution has the Gaussian form. However, as ATP concentration decreases, the run length distribution deviates from the Gaussian form. Moreover, both the mean velocity and mean run length decrease with the decrease of ATP concentration.
Figure 9.

Results for dimerized kinesin-3 at non-saturating ATP concentrations. The data are calculated with parameter values given in Table 1 for kinesin-3 and n = 10. The mean values of the data are indicated. (a–e) Velocity distribution. Lines are Gaussian fits. (f–j) Run length distribution. Line in (f) for [ATP] = 2 μM is the single-exponential fit, while lines in (g–j) for other values of [ATP] are the Gaussian fits.
Force dependence of run length for dimerized kinesin-3
Up to now, we have made studies under no load. The dependencies of the mean velocity and run length on load for kinesin-1 have been calculated before33,34. Thus, in this section we make only studies for dimerized kinesin-3 under a backward load, F, on the cargo, providing some predicted results and giving an explanation of the available single-molecule data for another species of dimerized kinesin-3, U356-Kstalk-GFP, which was created by joining the kinesin-1 neck coiled-coil and stalk to Unc104 motor domain22. In this section, we consider saturating ATP and F in the range smaller than the stall force so that the dissociation can be neglected when at least one head binds strongly to MT56.
Under the backward load on the cargo, we make following considerations. (i) The load on the NC helixes would disrupt the intramolecular interaction between the N-residues and kinesin-3 head in INT state before the formation of the intermolecular interaction between N-residues in the two helixes. This implies that at beginning of the processive movement of the dimerized kinesin-3, i.e., at t = 0, the ATPase activity of the MT-bound head in INT state is not inhibited, with the rate of ATP hydrolysis and Pi release being all the time. (ii) In INT state, when the NL of the MT-bound head is stretched to a length lNL > 2.8 nm under the backward load, the rate constant of NL docking (λ ≥ 1), as done before34. Here, we take two values of λ, with λ = 2 and 5, for the calculations. The equations for the calculations under the load are the same as those described previously33,34.
With the above two considerations, taking ENL = 5kBT and other parameters as given in Table 1 for kinesin-3, we calculate the run length and velocity for different values of the backward load F. Note here that the rate constants of ATPase activity are independent of F, as done before48,49,57,58. In Fig. 10a we show the calculated results of the run length distribution under no load (same as the left panel of Fig. 4b). In Fig. 10b and c we show the calculated results of the run length distribution under F = 2 pN for λ = 2 and 5, respectively. It is seen that while under no load the distribution of run lengths has evidently a non-single-exponential form (Fig. 10a), under the load the distribution becomes single exponential (Fig. 10b,c). In Fig. 10d and e we show the calculated results of the mean velocity and mean run length versus load F. From Fig. 10e we see interestingly that even under a small backward load the run length decreases significantly relative to that under no load, and as the backward load increases the run length decreases slowly. This feature for kinesin-3 is in sharp contrast to that for kinesin-1, whose run length under a small backward load is reduced only slightly relative to that under no load27,33–35. In Fig. 10f we show the force dependence of dissociation rate kd for kinesin-3, which is calculated from the mean detachment time (noting that kd can also be obtained by dividing mean velocity by mean run length). From Fig. 10f it is seen that a small load can enhance the dissociation rate significantly, especially with the large value of λ. As the load increases, the dissociation rate changes only slightly. This feature is consistent with the available single-molecule data for U356-Kstalk-GFP (Fig. 10g)22. Moreover, it is noted that the calculated curve of velocity versus F (Fig. 10d) also resembles well the available experimental curve for U356-Kstalk-GFP (Fig. 10h)22. The measured run length distribution under no load for U356-Kstalk-GFP (Fig. 10i)22 also evidently has a non-single-exponential form, similar to the calculated one (Fig. 10a).
Figure 10.

Results of the force dependencies of velocity and run length for dimerized kinesin-3 at saturating ATP. The theoretical data are calculated with ENL = 5kBT and other parameter values given in Table 1 for kinesin-3 and n = 10. The experimental data are taken from Tomishige et al.22. (a) Calculated run length distribution under no load. (b) Calculated run length distribution under F = 2 pN for λ = 2. (c) Calculated run length distribution under F = 2 pN for λ = 5. (d) Calculated mean velocity versus load. (e) Calculated mean run length versus load. (f) Calculated dissociation rate versus load. (g) Experimentally measured dissociation rate versus load for U356-Kstalk-GFP. (h) Experimentally measured velocity versus load for U356-Kstalk-GFP. (i) Experimentally measured run length distribution under no load for U356-Kstalk-GFP.
Based on our model, the great increase of the dissociation rate under a backward load relative to that under no load arises from two effects. One effect is that the load disrupts the intramolecular interaction between the N-residues and head in INT state before the formation of the intermolecular interaction between N-residues in the two helixes. This brings forward the time when Period I occurs, increasing the dissociation rate by μ-fold (μ > 1), as noted by comparing Fig. 4b (left panel) with Fig. 7b (left panel). The other effect is that under the backward load the rate constant of NL docking in the MT-bound head in INT state is reduced by λ-fold, increasing the occurrence probability of Period I by approximately λ-fold. Both effects are involved only in the dissociations during Period I. Since the dissociation during Period II is negligible, the two effects thus result in the total dissociation rate to increase by approximately μλ-fold.
In addition, we consider that the dissociation during Period II can also occur. We take Ew2 = 40.5kBT, which is smaller than Ew2 ≥45kBT, while with other parameter values being given as just above in Fig. 10 for kinesin-3. Under no load, the distribution of run lengths with Ew2 = 40.5kBT becomes single exponential and the mean run length is reduced greatly (Fig. 11a). This is consistent with the available experimental data for U356-Kstalk-GFP-K0 (Fig. 11b), where the K-loop in U356-Kstalk-GFP was swapped with that from kinesin-122, implying that Ew2 for U356-Kstalk-GFP-K0 is reduced relative to that for U356-Kstalk-GFP. The calculated results of dissociation rate versus F with Ew2 = 40.5kBT are shown in Fig. 11c (unfilled circles, λ = 5), where for comparison the corresponding calculated data with Ew2 ≥45kBT are re-shown (filled circles, λ = 5). From Fig. 11c, it is seen that while under no load the dissociation rate with Ew2 = 40.5kBT is much larger than that with Ew2 ≥45kBT, under loads the dissociation rates with Ew2 = 40.5kBT become closer to those with Ew2 ≥45kBT. These features are also consistent with the available single-molecule data for U356-Kstalk-GFP-K0 (Fig. 11d, unfilled circles) relative to those for U356-Kstalk-GFP (Fig. 11d, filled circles)22.
Figure 11.

Results of the force dependencies of velocity and run length for dimerized kinesin-3 with reduced affinity Ew2 at saturating ATP. The theoretical data are calculated with ENL = 5kBT and other parameter values given in Table 1 for kinesin-3 and n = 10 except for Ew2 = 40.5kBT. The experimental data are taken from Tomishige et al.22. (a) Calculated run length distribution under no load. (b) Experimentally measured run length distribution under no load for U356-Kstalk-GFP-K0. (c) Calculated dissociation rate versus load (unfilled circles). λ = 5. (d) Experimentally measured dissociation rate versus load for U356-Kstalk-GFP-K0 (unfilled circles). The filled circles in (c) are calculated data with Ew2 ≥ 45kBT and λ = 5. The filled circles in (d) are experimental data for U356-Kstalk-GFP.
Discussion
Soppina et al23. showed that in the monomeric state the full-length kinesin-3 is inactive in motility and only after the dimerization can the kinesin-3 move processively. By contrast, Okada et al59,60. and Oriola et al61,62. showed clearly that the truncated monomeric kinesin-3 KIF1A can move processively. Based on our proposed model, these contradictory results can be explained easily. In our model, in the monomeric state the formation of the intramolecular interaction between the N-residues in NC segment and the head inhibits the ATPase activity of the head, and the dimerization prevents the formation of the intramolecular interaction between the N-residues and the head, without inhibiting the ATPase activity of the head. In the monomeric state of the full-length kinesin-3 used by Soppina et al.23, the intramolecular interaction can form between the N-residues of NC segment and the head, inhibiting the ATPase activity of the head. By contrast, in the monomeric state of truncated kinesin-3 KIF1A (amino acids 1–382) used by Okada et al59,60. and Oriola et al.61,62, where the N-residues in NC segment are supposed to be deleted, the ATPase activity of the head is not inhibited. As a result, the single KIF1A monomer can move processively with the Brownian ratchet mechanism proposed before43,59–63.
As previous theoretical studies showed24,25, if a molecular motor has a constant rate to dissociate from its track during its processive movement or the motor has a constant probability to dissociate in each mechanochemical coupling cycle, a single-exponential distribution of run lengths is obtained. For kinesin-1 it is true that the motor has a constant dissociation rate, but for kinesin-3 this is not the case, as described as follows. As mentioned above, the dissociation of the kinesin dimer from MT can occur in two periods—Period I and Period II (see Fig. 1), and the motor has a constant probability to dissociate in each period. Thus, the dissociation rate of the kinesin dimer can be calculated by , where kI is the occurrence rate of Period I, PI the dissociation probability in Period I, kII the occurrence rate of Period II and PII the dissociation probability in Period II. Since PI and PII have constant values, it is thus seen that the non-exponential distribution of run lengths, which arises from a non-constant ε, requires kI and/or kII having non-constant values. For kinesin-1, both Period I and Period II occur with constant rates kI and kII, respectively, giving a constant ε and thus a single-exponential distribution of run lengths. By contrast, for kinesin-3, Period I occurs with a non-constant rate kI, which is dictated by the non-exponential distribution of time when the intermolecular interaction between the N-residues in the two helixes is formed [see, Eq. (1)]. This gives a non-constant ε and thus a non-exponential distribution of run lengths. Note that for kinesin-3, since PII 0, kII has no effect on ε. Thus, it is only required that kI has a non-constant value to give a non-constant ε and a non-exponential distribution of run lengths.
In summary, we provided quantitative explanations of the experimental data on dynamics of dimerized kinesin-3. The peculiar features of approximate Gaussian distribution of run lengths and superprocessivity for kinesin-3 are due to the reason that after cargo-mediated dimerization the ATPase activity of the MT-bound head in INT state is still inhibited for a time period after the heads become active in the two-heads-bound state. By contrast, for kinesin dimers such as kinesin-1, kinesin-2, kinesin-4, kinesin-7, etc., which have intrinsically stable coiled-coil structures and the ATPase activities of the MT-bound head in INT state are not inhibited, their run length distributions have the normally single-exponential form. Moreover, the force dependence of run length or dissociation rate for kinesin-3 was also studied, explaining the available experimental data showing contrast features of the force dependence of dissociation rate between kinesin-3 and kinesin-1.
Supplementary information
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 11775301) and National Key R&D Program of China (2016YFA0301500).
Author contributions
P.X. designed and organized the study. S.-K.G. performed simulations. S.-K.G. and P.X. analyzed the data. X.-X.S. and P.-Y.W. assisted in research and discussed results. P.X. wrote the manuscript. All authors reviewed the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-019-53550-2.
References
- 1.Vale RD. The molecular motor toolbox for intracellular transport. Cell. 2003;112:467–480. doi: 10.1016/S0092-8674(03)00111-9. [DOI] [PubMed] [Google Scholar]
- 2.Lawrence CJ, et al. A standardized kinesin nomenclature. The Journal of cell biology. 2004;167:19–22. doi: 10.1083/jcb.200408113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hirokawa N, Noda Y, Tanaka Y, Niwa S. Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Bio. 2009;10:682. doi: 10.1038/nrm2774. [DOI] [PubMed] [Google Scholar]
- 4.Miki H, Okada Y, Hirokawa N. Analysis of the kinesin superfamily: insights into structure and function. Trends Cell Biol. 2005;15:467–476. doi: 10.1016/j.tcb.2005.07.006. [DOI] [PubMed] [Google Scholar]
- 5.Siddiqui N, Straube A. Intracellular cargo transport by kinesin-3 motors. Biochemistry (Moscow). 2017;82:803–815. doi: 10.1134/S0006297917070057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hall DH, Hedgecock EM. Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans. Cell. 1991;65:837–847. doi: 10.1016/0092-8674(91)90391-B. [DOI] [PubMed] [Google Scholar]
- 7.Otsuka AJ, et al. The C. elegans unc-104 4 gene encodes a putative kinesin heavy chain-like protein. Neuron. 1991;6:113–122. doi: 10.1016/0896-6273(91)90126-K. [DOI] [PubMed] [Google Scholar]
- 8.Horiguchi K, Hanada T, Fukui Y, Chishti AH. Transport of PIP3 by GAKIN, a kinesin-3 family protein, regulates neuronal cell polarity. The Journal of cell biology. 2006;174:425–436. doi: 10.1083/jcb.200604031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jenkins B, Decker H, Bentley M, Luisi J, Banker G. A novel split kinesin assay identifies motor proteins that interact with distinct vesicle populations. J Cell Biol. 2012;198:749–761. doi: 10.1083/jcb.201205070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lo K, Kuzmin A, Unger S, Petersen J, Silverman M. KIF1A is the primary anterograde motor protein required for the axonal transport of dense-core vesicles in cultured hippocampal neurons. Neurosci Lett. 2011;491:168–173. doi: 10.1016/j.neulet.2011.01.018. [DOI] [PubMed] [Google Scholar]
- 11.Okada Y, Yamazaki H, Sekine-Aizawa Y, Hirokawa N. The neuron-specific kinesin superfamily protein KIF1A is a uniqye monomeric motor for anterograde axonal transport of synaptic vesicle precursors. Cell. 1995;81:769–780. doi: 10.1016/0092-8674(95)90538-3. [DOI] [PubMed] [Google Scholar]
- 12.Nangaku M, et al. KIF1B, a novel microtubule plus end-directed monomeric motor protein for transport of mitochondria. Cell. 1994;79:1209–1220. doi: 10.1016/0092-8674(94)90012-4. [DOI] [PubMed] [Google Scholar]
- 13.Lamason RL, Kupfer A, Pomerantz JL. The dynamic distribution of CARD11 at the immunological synapse is regulated by the inhibitory kinesin GAKIN. Mol Cell. 2010;40:798–809. doi: 10.1016/j.molcel.2010.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ahmed SM, et al. KIF14 negatively regulates Rap1a-Radil signaling during breast cancer progression. J Cell Biol. 2012;199:951–967. doi: 10.1083/jcb.201206051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Torres JZ, et al. The STARD9/Kif16a kinesin associates with mitotic microtubules and regulates spindle pole assembly. Cell. 2011;147:1309–1323. doi: 10.1016/j.cell.2011.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Samwer M, et al. The nuclear F‐actin interactome of Xenopus oocytes reveals an actin‐bundling kinesin that is essential for meiotic cytokinesis. The EMBO journal. 2013;32:1886–1902. doi: 10.1038/emboj.2013.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tsai J-W, Lian W-N, Kemal S, Kriegstein AR, Vallee RB. Kinesin 3 and cytoplasmic dynein mediate interkinetic nuclear migration in neural stem cells. Nat Neurosci. 2010;13:1463. doi: 10.1038/nn.2665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kratchmarov R, et al. Glycoproteins gE and gI are required for efficient KIF1A-dependent anterograde axonal transport of alphaherpesvirus particles in neurons. J Virol. 2013;87:9431–9440. doi: 10.1128/JVI.01317-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kramer T, et al. Kinesin-3 mediates axonal sorting and directional transport of alphaherpesvirus particles in neurons. Cell host & microbe. 2012;12:806–814. doi: 10.1016/j.chom.2012.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ueno H, Huang X, Tanaka Y, Hirokawa N. KIF16B/Rab14 molecular motor complex is critical for early embryonic development by transporting FGF receptor. Developmental cell. 2011;20:60–71. doi: 10.1016/j.devcel.2010.11.008. [DOI] [PubMed] [Google Scholar]
- 21.Verhey KJ, Kaul N, Soppina V. Kinesin assembly and movement in cells. Annual review of biophysics. 2011;40:267–288. doi: 10.1146/annurev-biophys-042910-155310. [DOI] [PubMed] [Google Scholar]
- 22.Tomishige M, Klopfenstein DR, Vale RD. Conversion of Unc104/KIF1A kinesin into a processive motor after dimerization. Science. 2002;297:2263–2267. doi: 10.1126/science.1073386. [DOI] [PubMed] [Google Scholar]
- 23.Soppina V, et al. Dimerization of mammalian kinesin-3 motors results in superprocessive motion. Proceedings of the National Academy of Sciences. 2014;111:5562–5567. doi: 10.1073/pnas.1400759111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vu HT, Chakrabarti S, Hinczewski M, Thirumalai D. Discrete step sizes of molecular motors lead to bimodal non-Gaussian velocity distributions under force. Phys Rev Lett. 2016;117:078101. doi: 10.1103/PhysRevLett.117.078101. [DOI] [PubMed] [Google Scholar]
- 25.Zhang Y, Kolomeisky AB. Theoretical Investigation of Distributions of Run Lengths for Biological Molecular Motors. J. Phys. Chem. B. 2018;122:3272–3279. doi: 10.1021/acs.jpcb.7b09352. [DOI] [PubMed] [Google Scholar]
- 26.Walter WJ, Beránek V, Fischermeier E, Diez S. Tubulin acetylation alone does not affect kinesin-1 velocity and run length in vitro. PLoS One. 2012;7:e42218. doi: 10.1371/journal.pone.0042218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Milic B, Andreasson JO, Hancock WO, Block SM. Kinesin processivity is gated by phosphate release. Proceedings of the National Academy of Sciences. 2014;111:14136–14140. doi: 10.1073/pnas.1410943111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.van Riel WE, et al. Kinesin-4 KIF21B is a potent microtubule pausing factor. Elife. 2017;6:e24746. doi: 10.7554/eLife.24746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yardimci H, Van Duffelen M, Mao Y, Rosenfeld SS, Selvin PR. The mitotic kinesin CENP-E is a processive transport motor. Proceedings of the National Academy of Sciences. 2008;105:6016–6021. doi: 10.1073/pnas.0711314105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sturgill EG, et al. Kinesin-12 Kif15 targets kinetochore fibers through an intrinsic two-step mechanism. Curr Biol. 2014;24:2307–2313. doi: 10.1016/j.cub.2014.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Scarabelli G, et al. Mapping the processivity determinants of the kinesin-3 motor domain. Biophys J. 2015;109:1537–1540. doi: 10.1016/j.bpj.2015.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Soppina V, Verhey KJ. The family-specific K-loop influences the microtubule on-rate but not the superprocessivity of kinesin-3 motors. Mol Biol Cell. 2014;25:2161–2170. doi: 10.1091/mbc.e14-01-0696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guo S, Shi X, Wang P, Xie P. Processivity of dimeric kinesin-1 molecular motors. FEBS Open Bio. 2018;8:1332–1351. doi: 10.1002/2211-5463.12486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Guo S-K, Wang W-C, Wang P-Y, Xie P. Force dependence of velocity and run length of kinesin-1, kinesin-2 and kinesin-5 family molecular motors. Molecules. 2019;24:287. doi: 10.3390/molecules24020287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Andreasson JO, et al. Examining kinesin processivity within a general gating framework. Elife. 2015;4:e07403. doi: 10.7554/eLife.07403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Isabelle M-TC, Lockhart A, Cross RA. Weak and strong states of kinesin and ncd. J Mol Biol. 1996;257:66–76. doi: 10.1006/jmbi.1996.0147. [DOI] [PubMed] [Google Scholar]
- 37.Hancock WO, Howard J. Kinesin’s processivity results from mechanical and chemical coordination between the ATP hydrolysis cycles of the two motor domains. Proceedings of the National Academy of Sciences. 1999;96:13147–13152. doi: 10.1073/pnas.96.23.13147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sosa H, Peterman EJ, Moerner W, Goldstein LS. ADP-induced rocking of the kinesin motor domain revealed by single-molecule fluorescence polarization microscopy. Nature Structural and Molecular Biology. 2001;8:540. doi: 10.1038/88611. [DOI] [PubMed] [Google Scholar]
- 39.Uemura, S., Kawaguchi et al. nucleotide state and loading direction, Proceedings of the National Academy of Sciences. 99, 5977–5981. [DOI] [PMC free article] [PubMed]
- 40.Cross R. Mechanochemistry of the kinesin‐1 ATPase. Biopolymers. 2016;105:476–482. doi: 10.1002/bip.22862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xie P. Stepping behavior of two-headed kinesin motors. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 2008;1777:1195–1202. doi: 10.1016/j.bbabio.2008.04.040. [DOI] [PubMed] [Google Scholar]
- 42.Xie P. Mechanism of processive movement of monomeric and dimeric kinesin molecules. International Journal of Biological Sciences. 2010;6:665. doi: 10.7150/ijbs.6.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xie P, Dou S-X, Wang P-Y. Processivity of single-headed kinesin motors. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 2007;1767:1418–1427. doi: 10.1016/j.bbabio.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 44.Shi X-X, et al. Investigating role of conformational changes of microtubule in regulating its binding affinity to kinesin by all-atom molecular dynamics simulation. Proteins. 2018;86:1127–1139. doi: 10.1002/prot.25592. [DOI] [PubMed] [Google Scholar]
- 45.Morikawa, M. et al. X-ray and Cryo-EM structures reveal mutual conformational changes of Kinesin and GTP-state microtubules upon binding, The EMBO journal, e201490588 (2015). [DOI] [PMC free article] [PubMed]
- 46.Rice S, et al. Thermodynamic properties of the kinesin neck-region docking to the catalytic core. Biophys J. 2003;84:1844–1854. doi: 10.1016/S0006-3495(03)74992-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Khalil AS, et al. Kinesin’s cover-neck bundle folds forward to generate force. Proceedings of the National Academy of Sciences. 2008;105:19247–19252. doi: 10.1073/pnas.0805147105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Guo S-K, Wang P-Y, Xie P. A model of processive movement of dimeric kinesin. J Theor Biol. 2017;414:62–75. doi: 10.1016/j.jtbi.2016.11.023. [DOI] [PubMed] [Google Scholar]
- 49.Guo S-K, Wang P-Y, Xie P. Dynamics of dimeric kinesins: Limping, effect of longitudinal force, effects of neck linker extension and mutation, and comparison between kinesin-1 and kinesin-2. Int J Biol Macromol. 2017;105:1126–1137. doi: 10.1016/j.ijbiomac.2017.07.147. [DOI] [PubMed] [Google Scholar]
- 50.Shi X.-X., Guo S.-K., Wang P.-Y., Chen H., Xie P. All-atom molecular dynamics simulations reveal how kinesin transits from one-head-bound to two-heads-bound state. Proteins, 10.1002/prot.25833 (2019). [DOI] [PubMed]
- 51.Hackney DD. Evidence for alternating head catalysis by kinesin during microtubule-stimulated ATP hydrolysis. Proceedings of the National Academy of Sciences. 1994;91:6865–6869. doi: 10.1073/pnas.91.15.6865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rosenfeld SS, Jefferson GM, King PH. ATP reorients the neck linker of kinesin in two sequential steps. J Biol Chem. 2001;276:40167–40174. doi: 10.1074/jbc.M103899200. [DOI] [PubMed] [Google Scholar]
- 53.Cross RA. The kinetic mechanism of kinesin. Trends Biochem Sci. 2004;29:301–309. doi: 10.1016/j.tibs.2004.04.010. [DOI] [PubMed] [Google Scholar]
- 54.Cao L, et al. The structure of apo-kinesin bound to tubulin links the nucleotide cycle to movement. Nature communications. 2014;5:5364. doi: 10.1038/ncomms6364. [DOI] [PubMed] [Google Scholar]
- 55.Xie S. Single-Molecule Approach to Enzymology. Single Molecules. 2001;2:229–236. doi: 10.1002/1438-5171(200112)2:4<229::AID-SIMO229>3.0.CO;2-9. [DOI] [Google Scholar]
- 56.Guo S, Shi X, Wang P, Xie P. Force dependence of unbinding rate of kinesin motor during its processive movement on microtubule. Biophys. Chem. 2019;253:106216. doi: 10.1016/j.bpc.2019.106216. [DOI] [PubMed] [Google Scholar]
- 57.Xie P, Chen H. A non-tight chemomechanical coupling model for force-dependence of movement dynamics of molecular motors. Phys. Chem. Chem. Phys. 2018;20:4752–4759. doi: 10.1039/C7CP05557A. [DOI] [PubMed] [Google Scholar]
- 58.Xie P, Guo S-K, Chen H. ATP-concentration- and force-dependent chemomechanical coupling of kinesin molecular motors. J. Chem. Inf. Model. 2019;59:360–372. doi: 10.1021/acs.jcim.8b00577. [DOI] [PubMed] [Google Scholar]
- 59.Okada Y, Hirokawa N. A processive single-headed motor: kinesin superfamily protein KIF1A. Science. 1999;283:1152–1157. doi: 10.1126/science.283.5405.1152. [DOI] [PubMed] [Google Scholar]
- 60.Okada Y, Hirokawa N. Mechanism of the single-headed processivity: diffusional anchoring between the K-loop of kinesin and the C terminus of tubulin. Proc. Natl. Acad. Sci. USA. 2000;97:640–645. doi: 10.1073/pnas.97.2.640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Oriola D, Roth S, Dogterom M, Casademunt J. Formation of helical membrane tubes around microtubules by single-headed kinesin KIF1A. Nat. Commun. 2015;6:8025. doi: 10.1038/ncomms9025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mitra A, et al. A Brownian Ratchet Model Explains the Biased Sidestepping of Single-Headed Kinesin-3 KIF1A. Biophys. J. 2019;116:2266–2274. doi: 10.1016/j.bpj.2019.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Oriola D, Casademunt J. Cooperative force generation of KIF1A Brownian motors. Phys. Rev. Lett. 2013;111:048103. doi: 10.1103/PhysRevLett.111.048103. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.