

Abstract
Nicotinic acid has been used for decades for its antiatherogenic properties in humans. Its actions on lipid metabolism intersect with multiple sleep regulatory mechanisms, but its effects on sleep have never been documented. For the first time, we investigated the effects of acute systemic administration of nicotinic acid on sleep in mice. Intraperitoneal and oral gavage administration of nicotinic acid elicited robust increases in non-rapid-eye movement sleep (NREMS) and decreases in body temperature, energy expenditure and food intake. Preventing hypothermia did not affect its sleep-inducing actions suggesting that altered sleep is not secondary to decreased body temperature. Systemic administration of nicotinamide, a conversion product of nicotinic acid, did not affect sleep amounts and body temperature, indicating that it is not nicotinamide that underlies these actions. Systemic administration of monomethyl fumarate, another agonist of the nicotinic acid receptor GPR109A, fully recapitulated the somnogenic and thermoregulatory effects of nicotinic acid suggesting that they are mediated by the GPR109A receptor. The cyclooxygenase inhibitor indomethacin completely abolished the effects of nicotinic acid indicating that prostaglandins play a key role in mediating the sleep and thermoregulatory responses of nicotinic acid.
Subject terms: Neurophysiology, Circadian rhythms and sleep
Introduction
Metabolic signals play an important role in sleep regulation. Positive energy balance, such as postprandial state, overfeeding and obesity, is accompanied by increased sleep1–4, while negative energy states, such as fasting, promote wakefulness5–8. Furthermore, hormones that are associated with satiety and/or obesity (e.g., cholecystokinin, tumor necrosis factor) induce sleep9,10, while orexigenic hormones that are associated with fasting and negative energy states (e.g., ghrelin, orexin), enhance wakefulness11–13. Sleep-inducing signals also arise from peripheral metabolic organs, such as the intestines, adipose tissues, liver and skeletal muscle14–18.
Sleep is particularly sensitive to shifts in lipid metabolism. The activation of the somatotropic axis, proinflammatory cytokines, bacterial cell wall components, norepinephrine and β3-adrenergic receptor (β3-AR) agonists all stimulate lipolysis, increase circulating levels of free fatty acids (FFAs)19–22 and also facilitate sleep16,23–28. In nocturnal rodents, sleep is more abundant during the light, lipolytic phase, and wakefulness dominates during the dark, lipogenic phase of the day. Reversal of the lipolytic and lipogenic phases by sequential administration of metabolic hormones25 or by restricting feeding to the light phase in rats or mice results in the reversal of the sleep-wake cycle29,30. Nicotinic acid, a member of the B3 vitamin group, has been used for over 60 years to improve plasma lipid profile in humans31,32. Given the long history of the clinical use of nicotinic acid and its complex actions on lipid metabolism, it is remarkable that its effects on sleep has never been documented.
The acute effects of nicotinic acid intersect with multiple pathways in sleep regulation. In addition to its effects on lipid metabolism, nicotinic acid also markedly stimulates prostaglandin synthesis33,34. Prostaglandin D2 (PGD2) is one of the most potent endogenous sleep-promoting substances35. Furthermore, nicotinic acid facilitates the alternative activation of macrophages36–38. Alternatively activated macrophages are permissive for maintaining sleep after sleep loss and in cold environment39. Nicotinic acid is a potent stimulus for growth hormone secretion40,41, and the somatotropic axis has long been implicated in sleep regulation27. Finally, butyrate, which binds to the nicotinic acid receptor GPR109A, as well as the short-chain fatty acid receptors FFAR2 and FFAR3, has pronounced sleep-promoting effects42.
Considering the reciprocal relationship between sleep and lipid metabolism, the complex actions of nicotinic acid on lipid metabolism and the existence of other multiple regulatory nodes where nicotinic acid may potentially modulate sleep, we investigated the effects of systemic administration of nicotinic acid on sleep. We report that systemic administration of nicotinic acid and another GPR109A receptor agonist, monomethyl fumarate, induces robust sleep and hypothermic responses in mice. Preventing the nicotinic acid-induced hypothermia with β3-AR agonist pretreatment did not attenuate the sleep responses, but the inhibition of prostaglandin synthesis abolished both the sleep and thermoregulatory actions of nicotinic acid.
Results
Systemic administration of nicotinic acid elicits robust non-rapid-eye movement sleep (NREMS) increases
Intraperitoneal (ip) injection of nicotinic acid brought about marked, dose-dependent increases in NREMS (Fig. 1, Table 1). Intraperitoneal administration of 50 mg/kg nicotinic acid did not affect sleep-wake activity, body temperature or motor activity. The effects of 100 mg/kg nicotinic acid were manifested after a latency of one hour; starting from h 2, NREMS increased 41% above baseline for 4 hours (NREMS amounts in h 2–5, baseline: 82.7 ± 7.3 min, treatment: 116.8 ± 9.6 min, p < 0.01). Administration of 250 mg/kg nicotinic acid elicited robust and long-lasting increases in NREMS; NREMS was elevated 120% above baseline in the 2–11 h time block (baseline: 179.0 ± 7.1 min, treatment: 394.6 ± 20.4 min, p < 0.001). The effects of oral administration of 1 g/kg nicotinic acid were similar to those of the highest ip dose. NREMS increased by 86% in the 2–11 h time block, then returned to baseline in the latter half of the recording period (baseline: 197.9 ± 8.1 min, treatment: 368.9 ± 25.0 min, p < 0.001). NREMS increases were accompanied by decreased electroencephalographic (EEG) slow-wave activity (SWA) and suppressed motor activity (Fig. 1, Table 1).
Figure 1.

Effects of intraperitoneal (ip) administration of 50, 100 and 250 mg/kg and intragastric delivery of 1 g/kg nicotinic acid on non-rapid-eye movement sleep (NREMS), rapid-eye movement sleep (REMS), electroencephalographic slow wave activity (EEG SWA), body temperature and motor activity in mice. Data are expressed in 1-h time blocks, except EEG SWA values which are shown in 2-h blocks. Nicotinic acid and saline were injected at time “0”. Grey shaded areas represent the dark period. Asterisks: significant difference between control and nicotinic acid treatments, Tukey’s HSD test; error bar: SE.
Table 1.
The effects of nicotinic acid on non-rapid-eye movement sleep (NREMS), rapid-eye movement sleep (REMS), body temperature, motor activity and electroencephalographic slow-wave activity (EEG SWA): statistical results.
| NREMS | REMS | Temperature | Activity | SWA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| 50 mg/kg, ip | |||||||||||||||
| Treatment | 1,6 | 0.6 | n.s. | 1,6 | 12.6 | p < 0.05 | 1,6 | 13.9 | p < 0.01 | 1,6 | 1.7 | n.s. | 1,6 | 0.2 | n.s. |
| Time | 23,138 | 6.3 | p < 0.001 | 23,138 | 7.7 | p < 0.001 | 23,138 | 28.5 | p < 0.001 | 23,138 | 7.4 | p < 0.001 | 11,66 | 1.5 | n.s. |
| Treatment × Time | 23,138 | 1.2 | n.s. | 23,138 | 0.9 | n.s. | 23,138 | 1.0 | n.s. | 23,138 | 1.3 | n.s. | 11,66 | 0.8 | n.s. |
| 100 mg/kg, ip | |||||||||||||||
| Treatment | 1,7 | 13.6 | p < 0.01 | 1,7 | 0.0 | n.s. | 1,7 | 0.0 | n.s. | 1,7 | 10.0 | p < 0.05 | 1,7 | 4.2 | n.s. |
| Time | 23,161 | 14.7 | p < 0.001 | 23,161 | 17.8 | p < 0.001 | 23,161 | 27.9 | p < 0.001 | 23,161 | 16.9 | p < 0.001 | 11,77 | 9.9 | p < 0.001 |
| Treatment × Time | 23,161 | 1.5 | n.s. | 23,161 | 0.9 | n.s. | 23,161 | 4.5 | p < 0.001 | 23,161 | 1.2 | n.s. | 11,77 | 2.0 | p < 0.05 |
| 250 mg/kg, ip | |||||||||||||||
| Treatment | 1,6 | 95.3 | p < 0.001 | 1,6 | 8.2 | p < 0.05 | 1,6 | 26.2 | p < 0.01 | 1,6 | 87.3 | p < 0.001 | 1,6 | 18.0 | p < 0.01 |
| Time | 23,138 | 8.9 | p < 0.001 | 23,138 | 15.2 | p < 0.001 | 23,138 | 15.7 | p < 0.001 | 23,138 | 10.1 | p < 0.001 | 11,66 | 14.3 | p < 0.001 |
| Treatment × Time | 23,138 | 9.3 | p < 0.001 | 23,138 | 1.4 | n.s. | 23,138 | 35.2 | p < 0.001 | 23,138 | 8.0 | p < 0.001 | 11,66 | 16.5 | p < 0.001 |
| 1,000 mg/kg, oral gavage | |||||||||||||||
| Treatment | 1,4 | 23.3 | p < 0.01 | 1,4 | 2.1 | n.s. | 1,4 | 4.2 | n.s. | 1,5 | 19.2 | p < 0.01 | 1,4 | 1.2 | n.s. |
| Time | 23,92 | 3.4 | p < 0.001 | 23,92 | 11.0 | p < 0.001 | 23,92 | 3.7 | p < 0.001 | 23,115 | 5.2 | p < 0.001 | 11,44 | 4.3 | p < 0.001 |
| Treatment × Time | 23,92 | 5.2 | p < 0.001 | 23,92 | 1.5 | n.s. | 23,92 | 5.9 | p < 0.001 | 23,115 | 3.2 | p < 0.001 | 11,44 | 5.1 | p < 0.001 |
Nicotinic acid had only minor effects on rapid-eye movement sleep (REMS) without any apparent dose-dependency. Oral gavage administration of 1 g/kg or ip injection of 100 mg/kg nicotinic acid did not affect REMS, while 50 and 250 mg/kg had a slight, but statistically significant effect (Table 1, Fig. 1). After 50 mg/kg nicotinic acid, REMS was elevated in h 21–23, while after 250 mg/kg, it was slightly below baseline in h 12 and 20.
On the recovery days following the ip administration of 250 mg/kg or the gavage administration of 1 g/kg nicotinic acid, sleep-wake activity returned to baseline with no significant rebound responses (for ip 250 mg/kg nicotinic acid, wakefulness baseline: 482.2 ± 9.1 and 208.6 ± 7.8 min/12 h during the dark and light periods, respectively, recovery day: 460.7 ± 17.7 and 201.1 ± 4.5 min/12 h during the dark and light periods, respectively; NREMS baseline: 224.5 ± 8.7 and 449.1 ± 7.8 min/12 h, recovery day: 240.9 ± 15.1 and 452.9 ± 4.6 min/12 h; REMS baseline: 13.3 ± 1.7 and 62.4 ± 5.5 min /12 h, recovery day: 18.4 ± 3.1 and 65.9 ± 1.8 min/12 h; for gavage 1 g/kg nicotinic acid, wakefulness baseline: 471.0 ± 15.6 and 217.8 ± 14.7 min/12 h during the dark and light periods, respectively, recovery day: 460.5 ± 20.5 and 214.4 ± 21.6 min/12 h during the dark and light periods, respectively; NREMS baseline: 235. 7 ± 11.3 and 443.2 ± 15.8 min/12 h, recovery day: 241.9 ± 14.8 and 447.6 ± 18.2 min/12 h; REMS baseline: 13.3 ± 4.9 and 59.1 ± 4.7 min /12 h, recovery day: 11.7 ± 3.5 and 57.9 ± 7.0 min/12 h).
The sleep responses were paralleled by significant suppressions in body temperature. Body temperature decreased by ~1 °C for 4 h after the 100 mg/kg dose. In response to 250 mg/kg nicotinic acid, the maximum of the hypothermic response reached 3 °C in the second hour after the treatment (Fig. 1, Table 1). The average body temperature in the 2–11 h time period, when NREMS increases were significant, was 1.34 ± 0.2 °C below baseline (p < 0.001). During the second half of the recording period temperatures increased slightly, but significantly, above baseline. The effects of oral administration of 1 g/kg nicotinic acid on body temperature were similar, but somewhat attenuated as compared to the effects of the highest ip dose.
Indomethacin blocks the effects of nicotinic acid on sleep and body temperature
To examine the role of endogenous prostaglandins in the sleep and thermoregulatory effects of nicotinic acid, mice were pretreated with 5 mg/kg indomethacin, a cyclooxygenase inhibitor, before the administration of 100 mg/kg nicotinic acid. Indomethacin pretreatment completely abolished the effects of nicotinic acid on sleep, EEG SWA, body temperature and motor activity (Fig. 2, Table 2). Indomethacin, when administered by itself, did not have any significant effect on sleep and body temperature (data not shown).
Figure 2.
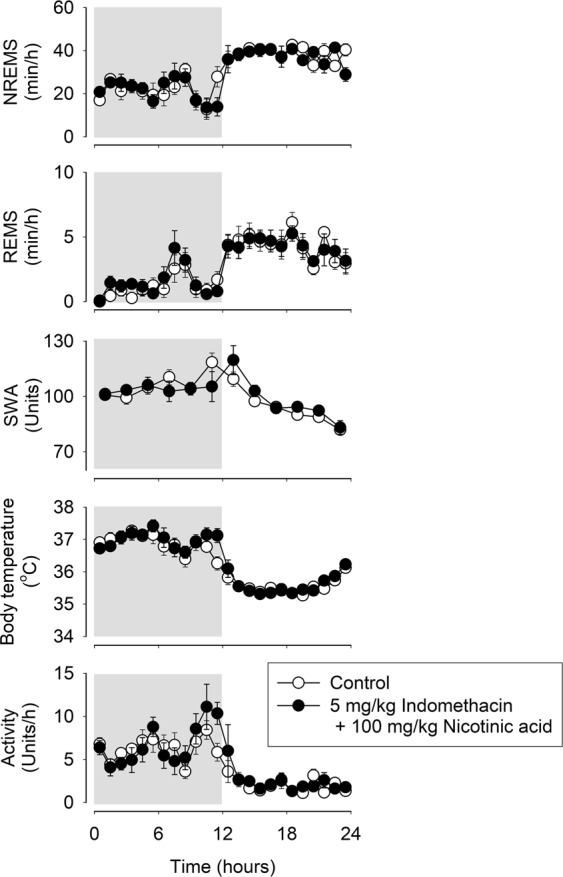
Effects of 5 mg/kg indomethacin pretreatment on the NREMS, REMS, EEG SWA, motor activity and body temperature-modulating actions of 100 mg/kg nicotinic acid in mice. See legend to Fig. 1 for details.
Table 2.
Indomethacin pretreatment followed by administration of nicotinic acid. NREMS, REMS, body temperature, motor activity and EEG SWA: statistical results.
| NREMS | REMS | Temperature | Activity | SWA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| Treatment | 1,6 | 0.4 | n.s. | 1,6 | 0.9 | n.s. | 1,6 | 1.2 | n.s. | 1,6 | 0.7 | n.s. | 1,6 | 0.2 | n.s. |
| Time | 23,138 | 14.4 | p < 0.001 | 23,138 | 12.3 | p < 0.001 | 23,138 | 44.7 | p < 0.001 | 23,138 | 10.8 | p < 0.001 | 11,66 | 10.7 | p < 0.001 |
| Treatment × Time | 23,138 | 1.2 | n.s. | 23,138 | 0.5 | n.s. | 23,138 | 1.3 | n.s. | 23,138 | 1.0 | n.s. | 11,66 | 1.3 | n.s. |
Decreased body temperature is not the cause of nicotinic acid-induced sleep
To investigate if decreased body temperature, per se, could account for increased sleep in response to nicotinic acid, mice were pretreated with CL-316,243, a β3-AR agonist, to stimulate thermogenesis thereby preventing hypothermia. CL-316,243 pretreatment completely abolished the hypothermic response to 250 mg/kg nicotinic acid but did not attenuate the late increases in body temperature in the second half of the recording period (Table 3, Fig. 3). The NREMS-promoting effects of nicotinic acid were not attenuated by the β3-AR agonist pretreatment, and sleep was increased by 134% in the 2–11 h time block (NREMS amounts in the 2–11 h time block; baseline: 179.0 ± 7.1 min, CL-316,243 + nicotinic acid treatment: 394.6 ± 20.4 min, p < 0.001). Likewise, the motor activity and EEG SWA suppressions after nicotinic acid administration were not affected by the CL-316,243 pretreatment.
Table 3.
CL-316,243 pretreatment followed by administration of nicotinic acid. NREMS, REMS, body temperature, motor activity EEG SWA: statistical results.
| NREMS | REMS | Temperature | Activity | SWA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| Treatment | 1,6 | 65.3 | p < 0.001 | 1,6 | 13.4 | p < 0.05 | 1,6 | 17.4 | p < 0.01 | 1,6 | 22.8 | p < 0.01 | 1,6 | 28.4 | p < 0.01 |
| Time | 23,138 | 4.2 | p < 0.001 | 23,138 | 21.5 | p < 0.001 | 23,138 | 25.7 | p < 0.001 | 23,138 | 9.2 | p < 0.001 | 11,66 | 3.2 | p < 0.01 |
| Treatment × Time | 23,138 | 9.2 | p < 0.001 | 23,138 | 0.8 | n.s. | 23,138 | 3.1 | p < 0.001 | 23,138 | 5.4 | p < 0.001 | 11,66 | 11.9 | p < 0.001 |
Figure 3.

Effects of the β3-adrenergic receptor agonist, CL-316,243, pretreatment on the NREMS, REMS, EEG SWA, motor activity and body temperature-modulating actions of 250 mg/kg nicotinic acid in mice. See legend to Fig. 1 for details.
Systemic administration of nicotinamide does not affect sleep amounts and body temperature
To test the possibility that its conversion into nicotinamide underlies the sleep-promoting effects of nicotinic acid, we investigated the effects of nicotinamide on sleep and body temperature. Systemic injection of 100 mg/kg (data not shown) or 250 mg/kg nicotinamide did not have any effect on sleep time and body temperature (Fig. 4, Table 4). The higher dose slightly decreased motor activity and suppressed EEG SWA.
Figure 4.
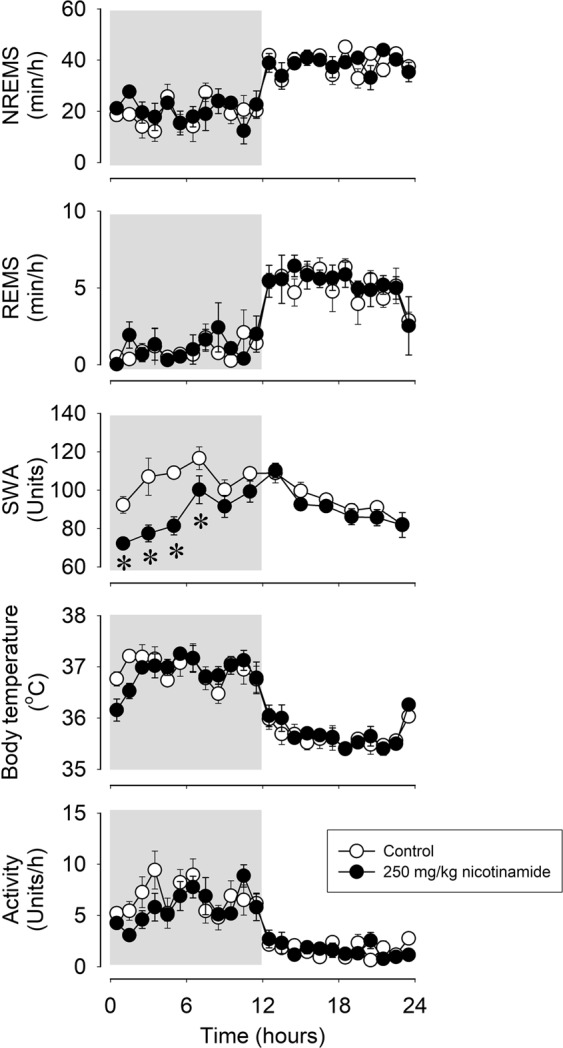
Effects of ip administration of 250 mg/kg nicotinamide on NREMS, REMS, EEG SWA, motor activity and body temperature in mice. See legend to Fig. 1 for details.
Table 4.
The effects of 250 mg/kg nicotinamide on NREMS, REMS, body temperature, motor activity and EEG SWA: statistical results.
| NREMS | REMS | Temperature | Activity | SWA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| Treatment | 1,4 | 0.1 | n.s. | 1,4 | 0.8 | n.s. | 1,4 | 0.0 | n.s. | 1,4 | 12.1 | p < 0.05 | 1,4 | 419.3 | p < 0.001 |
| Time | 23,92 | 15.6 | p < 0.001 | 23,92 | 18.7 | p < 0.001 | 23,92 | 32.9 | p < 0.001 | 23,92 | 12.2 | p < 0.001 | 11,44 | 4.3 | p < 0.001 |
| Treatment × Time | 23,92 | 1.2 | n.s. | 23,92 | 0.6 | n.s. | 23,92 | 1.1 | n.s. | 23,92 | 1.2 | n.s. | 11,44 | 4.8 | p < 0.001 |
Monomethylfumarate induces NREMS
To further investigate the involvement of the nicotinic acid receptor, GPR109A, in the sleep and thermoregulatory responses, we studied the effects of monomethyl fumarate (MMF), another GPR109A agonist. Administration of MMF resulted in sleep and body temperature responses similar to those of nicotinic acid. The lowest effective dose to induce sleep was 50 mg/kg, which elicited 84% increase in NREMS during the 4-h time block starting from h 2 (NREMS amounts in h 2–5, baseline: 75.4 ± 11.8 min, treatment: 138.4 ± 6.8 min, p < 0.001; Fig. 5, Table 5). The sleep effects were accompanied by a 0.6–0.9 °C drop in body temperature, suppressed motor activity and EEG SWA. A lower dose of MMF, 20 mg/kg, did not have significant effects on sleep and body temperature.
Figure 5.

Effects of ip administration of 20 and 50 mg/kg monomethyl fumarate on NREMS, REMS, EEG SWA, motor activity and body temperature in mice. See legend to Fig. 1 for details.
Table 5.
Intraperitoneal administration of monomethyl fumarate. NREMS, REMS, body temperature, motor activity and EEG SWA: statistical results.
| NREMS | REMS | Temperature | Activity | SWA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| 20 mg/kg | |||||||||||||||
| Treatment | 1,7 | 0.4 | n.s. | 1,7 | 3.1 | n.s. | 1,7 | 21.3 | p < 0.01 | 1,7 | 0.6 | n.s. | 1,7 | 0.9 | n.s. |
| Time | 23,161 | 7.0 | p < 0.001 | 23,161 | 17.5 | p < 0.001 | 23,161 | 23.3 | p < 0.001 | 23,161 | 8.1 | p < 0.001 | 11,77 | 3.8 | p < 0.001 |
| Treatment × Time | 23,161 | 1.0 | n.s. | 23,161 | 1.0 | n.s. | 23,161 | 1.1 | n.s. | 23,161 | 0.8 | n.s. | 11,77 | 1.1 | n.s. |
| 50 mg/kg | |||||||||||||||
| Treatment | 1,7 | 17.3 | p < 0.01 | 1,7 | 9.8 | p < 0.05 | 1,8 | 10.7 | p < 0.05 | 1,8 | 16.5 | p < 0.01 | 1,7 | 8.8 | p < 0.05 |
| Time | 23,161 | 8.3 | p < 0.001 | 23,161 | 13.0 | p < 0.001 | 23,184 | 23.2 | p < 0.001 | 23,184 | 12.9 | p < 0.001 | 11,77 | 6.6 | p < 0.001 |
| Treatment × Time | 23,161 | 3.1 | p < 0.001 | 23,161 | 1.6 | n.s. | 23,184 | 3.3 | p < 0.001 | 23,184 | 2.7 | p < 0.001 | 11,77 | 2.0 | p < 0.05 |
Nicotinic acid suppresses VO2, respiratory exchange ratio (RER) and feeding
To gain further insight in the action of somnogenic doses of nicotinic acid on metabolism, we investigated the effects of ip and oral administration of nicotinic acid on O2 consumption (VO2), RER and food intake. Intraperitoneal injection of 50 mg/kg nicotinic acid did not have any effect on these measures, while 250 mg/kg nicotinic acid induced marked and long-lasting suppression of VO2, RER and feeding. Similar robust effects were observed after gavage administration of 1 g/kg nicotinic acid (Fig. 6, Table 6).
Figure 6.

Effects of ip administration of 50 and 250 mg/kg and intragastric delivery of 1 g/kg nicotinic acid on oxygen consumption (VO2), respiratory exchange ratio (RER) and food intake in mice. Data are expressed in 1-h time blocks. See legend to Fig. 1 for details.
Table 6.
Effects of nicotinic acid on VO2, RER and food intake: statistical results.
| VO2 | RER | Food intake | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| Treatment | 1,3 | 3.2 | n.s. | 1,3 | 3.4 | n.s. | 1,3 | 6.9 | n.s. |
| Time | 23,69 | 17.7 | p < 0.001 | 23,69 | 8.9 | p < 0.001 | 23,69 | 9.1 | p < 0.001 |
| Treatment × Time | 23,69 | 1.1 | n.s. | 23,69 | 1.0 | n.s. | 23,69 | 1.4 | n.s. |
| 250 mg/kg, ip | |||||||||
| Treatment | 1,3 | 5.5 | n.s. | 1,3 | 39.7 | p < 0.01 | 1,3 | 10.4 | p < 0.05 |
| Time | 23,69 | 4.7 | p < 0.001 | 23,69 | 1.8 | p < 0.05 | 23,69 | 12.5 | p < 0.01 |
| Treatment × Time | 23,69 | 2.8 | p < 0.001 | 23,69 | 1.5 | n.s. | 23,69 | 3.8 | p < 0.001 |
| 1 g/kg, oral gavage | |||||||||
| Treatment | 1,3 | 6.4 | n.s. | 1,3 | 9.1 | 0.057 | 1,3 | 29.9 | p < 0.05 |
| Time | 23,69 | 7.3 | p < 0.001 | 23,69 | 1.6 | n.s. | 23,69 | 2.9 | p < 0.001 |
| Treatment × Time | 23,69 | 4.4 | p < 0.001 | 23,69 | 4.5 | p < 0.001 | 23,69 | 7.8 | p < 0.001 |
Discussion
Nicotinic acid was introduced into clinical therapy more than 60 years ago to improve plasma lipid profile, but its effects on sleep and body temperature have never been documented. Our main finding is that systemic injection or oral gavage administration of nicotinic acid induces robust increases in NREMS in mice. The effects were dose-dependent, ip administration of 100 and 250 mg/kg doses increased NREMS 41–120% above baseline for 4–11 h. Oral administration of 1 g/kg nicotinic acid had similar sleep-promoting potency as the 250 mg/kg injected ip. The effects were specific to NREMS, as REMS responses were minor and did not show dose-dependency.
Nicotinic acid is part of a group known as B3 complex, which also includes nicotinamide and nicotinamide riboside. After ip administration, 25% of the injected nicotinic acid is converted to nicotinamide in the liver in mice43. To determine if the acute sleep-promoting actions of nicotinic acid could be due to its conversion to nicotinamide, we also investigated the effects of bolus injection of nicotinamide. Intraperitoneal injection of 100 and 250 mg/kg nicotinamide did not have any significant effect on sleep amounts or body temperature, thus, it is unlikely that the sleep-promoting and thermoregulatory actions of nicotinic acid are mediated by its conversion to nicotinamide. Further, portions of circulating nicotinic acid and nicotinamide are converted to nicotinamide adenine dinucleotide (NAD) in the liver44. It has been reported that metabolic signals from the hepatoportal region modulate sleep17,42. Since the conversion of nicotinamide to NAD is significantly more efficient than that of nicotinic acid45, but nicotinamide lacks sleep-inducing activity, it is unlikely that nicotinic acid-induced increases in liver NAD content are responsible for the sleep-promoting effects.
Nicotinic acid induced significant decreases in body temperature. This can be explained, in part, by increased heat loss through cutaneous vasodilation. Nicotinic acid-induced vasodilation has been described in guinea pigs46, rats47 and mice34 and it is the cause of flushing, the well-known side effect of nicotinic acid megadoses in humans48. Vasodilation is a prompt and transient response to nicotinic acid48–50. In mice, vasodilation induced by subcutaneous injection of 100–300 mg/kg nicotinic acid reaches a peak within 2–3 min, and by 10 min the effect is reduced by 50%34. The sleep-inducing and the hypothermic actions of nicotinic acid, however, lasted for ~7–11 hours, thus it is unlikely that they are the direct consequences of the actions of nicotinic acid on cutaneous blood flow.
For thermoregulation, mice rely more on metabolic thermogenesis rather than vasomotor mechanisms51, and it is likely that the hypothermia is due to suppressed metabolic heat production. In fact, metabolic heat production (VO2) was suppressed in response to nicotinic acid in our study. Consistent with our finding, 200 mg/kg nicotinic acid attenuates cold-induced increases in thermogenesis in mice52 and norepinephrine-induced53 or spontaneous54 thermogenesis in rats. A major factor contributing to decreased thermogenesis is suppressed feeding in mice. Food intake was reduced by ~75% after 250 mg/kg ip and 1 g/kg oral doses of nicotinic acid in the first 8 hours, when VO2 and sleep responses were the most pronounced. Similar reductions in feeding were also reported after repeated nicotinic acid treatment in mice55. Since naturally occurring NREMS is associated by decreased energy expenditure and body temperature56, it is also possible that the hypothermic response is simply the thermic manifestation of enhanced NREMS after nicotinic acid treatment. Similar robust ~4 °C hypothermia was observed during enhanced sleep in response to the chemogenetic stimulation of the ventrolateral preoptic sleep center57.
There is a complex relationship between sleep and feeding. For example, sleep loss increases feeding58,59 and increased food intake induces postprandial sleep2,3. Since short-term food deprivation elicits robust increases in wakefulness and motor activity in mice5–7, it is unlikely that increased sleep and suppressed motor activity after nicotinic acid treatment are regulated energy-saving mechanisms in response to suppressed caloric intake. It is possible that the primary effect of nicotinic acid is increased sleep, and since sleep and feeding are mutually exclusive behaviors, decreased feeding may be the consequence of the general behavioral inactivity. Alternatively, the effects of nicotinic acid on sleep and feeding may be independent and mediated by different mechanisms.
It has been proposed that changes in body temperature may trigger somnogenic brain areas to initiate sleep60. To ascertain whether hypothermia per se could be a factor in the sleep-inducing effects of nicotinic acid, we also tested the effects in mice after the stimulation of thermogenesis by CL-316,243. CL-316,243 pretreatment completely abolished the hypothermic response, but the NREMS-promoting effects of nicotinic acid were not attenuated indicating that decreased body temperature, by itself, cannot account for the somnogenic effects of nicotinic acid.
Most, but not all, of the effects of nicotinic acid are mediated by GPR109A, the high-affinity nicotinic acid receptor61–63. To explore the role of the receptor in the sleep and thermoregulatory responses to nicotinic acid, we also used another receptor agonist, MMF64. MMF treatment fully recapitulated the effects of nicotinic acid on sleep, EEG SWA, body temperature and motor activity. Further, butyrate, another ligand for the GPR109A receptors as well as for the FFAR2, FFAR3 receptors, has similar marked NREMS-promoting and hypothermic effects as nicotinic acid42. These findings are consistent with the notion that the effects on sleep and temperature are mediated by the GPR109A receptor.
Nicotinic acid and MMF are potent stimuli for prostaglandin production via the activation of GPR109A receptors65–67. Ingestion of nicotinic acid results in 400 to 800-fold increases in PGD2 plasma levels in humans33, and systemic injection of 100–300 mg/kg leads to 1.5–4-fold increases in mice34. Since PGD2 has been identified as one of the most potent endogenous sleep-promoting molecules35, we sought to assess the role of prostaglandins in nicotinic acid-induced sleep by using the cyclooxygenase inhibitor indomethacin. Indomethacin pretreatment completely abolished the sleep-promoting as well as thermoregulatory actions of nicotinic acid indicating the involvement of endogenous prostaglandins both in the somnogenic and hypothermic effects. The only known significant sources of nicotinic acid-induced prostaglandin production are Langerhans cells and keratinocytes in the skin66,67. The ventral surface of the rostral basal forebrain has been identified as a PGD2-sensitive sleep-promoting site, and it is possible that peripherally produced prostaglandins reach this structure. Alternatively, prostaglandins may act on a peripheral target to induce sleep. For example, vagus afferents express EP3 and EP4 prostaglandin receptors68,69 and prostaglandins activate vagal sensory nerves70–72. It is known that vagal afferents carry sleep-inducing signals73–76.
Nicotinic acid, unlike nicotinamide, does not readily pass between blood and the cerebrospinal fluid77–79 and the relatively small amounts that enter the brain are promptly converted to nicotinamide77. Therefore, it is less likely that centrally-produced prostaglandins underly the sleep-inducing actions of nicotinic acid, although this possibility cannot be ruled out with certainty.
Nicotinic acid has complex actions on lipid metabolism. It is considered an antilipolytic agent because it suppresses FFA release from adipocytes in vitro and causes transient decreases in circulating FFA levels in vivo31. However, decreased FFA levels are followed by robust rebound increases 2–4 h after a single bolus treatment in humans80–82, dogs83 and rats83,84. Since the antilipolytic actions of nicotinic acid are independent of prostaglandins85, but the sleep-promoting actions require intact prostaglandin synthesis, it is unlikely that the antilipolytic actions of nicotinic acid directly contribute to its sleep effects. Independent of its lipolytic actions, nicotinic acid also promotes complex changes in plasma lipid profile, an action that is thought to underlie its antiatherogenic properties86. It cannot be ruled out that these changes may also contribute to metabolic signaling for sleep.
The extent to which our findings in mice are also applicable to humans is not known. It has been argued that the dose range used in our study corresponds well to the doses used in humans, if the 10-times faster metabolism of mice as compared to humans is considered87. The use of nicotinic acid supplements is widespread in the United States, and they are often advertised as sleep aids. In human trials, the most common reason for withdrawal is flushing followed by fatigue/sleepiness88–91. Our findings together with the above observational evidence point to the need of characterizing the effects of nicotinic acid on sleep in humans.
Methods
Animals
All procedures involving the use of animals followed the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All animal husbandry and experimental procedures were carried out in accordance with the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) and approved by the Institutional Animal Care and Use Committee (IACUC) of the Washington State University (protocol number 6031). Breeding pairs of C57BL/6J mice were purchased from The Jackson Laboratories, Inc. and were bred in-house at Washington State University. During the experiments, the animals were housed individually in temperature-controlled (29 ± 1 °C), sound-attenuated environmental chambers on a 12:12-hour light-dark cycle (lights on at 3 AM). Food (Harlan Teklad, Product no. 2016) and water were available unrestricted throughout all experiments.
Surgery
Male mice were surgically instrumented using ketamine-xylazine anesthesia (87 and 13 mg/kg, respectively). For sleep-wake activity recordings three cortical EEG electrodes, placed over the frontal and parietal cortices, and two nuchal electromyographic (EMG) electrodes were implanted. Leads from the EEG and EMG electrodes were anchored to the skull with dental cement. Telemetry transmitters were implanted ip for body temperature and motor activity recordings (Starr Life Sciences Corp., model G2 Emitter). The weight of a G2 Emitter is 1.1 g. The animals were allowed to recover from surgery for at least 10 days before any experimental manipulation started and handled daily to adapt them to the experimental procedures. The body weight of the mice at the time of the experiment was 28.9 ± 0.5 g.
Sleep-wake activity recordings and analyses
The animals were connected to the recording system through a lightweight, flexible tether plugged into a commutator, which was further routed to Grass Model 15 Neurodata amplifier system (Grass Instrument Division of Astro-Med, Inc., West Warwick, RI). The amplified EEG and EMG signals were digitized at 256 Hz and recorded by computer. The high-pass and low-pass filters for EEG signals were 0.5 and 30.0 Hz, respectively. The EMG signals were filtered with low and high cut-off frequencies at 100 and 10,000 Hz, respectively. The outputs from the 12A5 amplifiers were fed into an analog-to-digital converter and collected by computer using SleepWave software (Biosoft Studio, Hersey, PA). Sleep-wake states were scored visually off-line in 10-s segments. The vigilance states were defined as NREMS, REMS and wakefulness according to standard criteria as described previously92. Artifact-free EEG epochs were subjected to off-line spectral analysis by fast Fourier transformation. EEG power data in the range of 0.5 to 4.0 Hz during NREMS were used to compute EEG SWA. EEG SWA data were normalized for each animal by using the average EEG SWA across 24 h on the baseline day as 100.
Telemetry recordings
Core body temperature and locomotor activity were recorded by MiniMitter telemetry system (Starr Life Sciences Corp., model G2 Emitter and ER-4000 receiver) using VitalView software. Temperature and activity values were collected every 1 and 10 min, respectively, throughout the experiment and were averaged over 1-h time blocks.
Experimental procedures
Experiment 1: The effects of systemic administration of nicotinic acid on sleep-wake activity, body temperature and metabolic parameters in mice
Three groups of male mice were habituated to the injection procedure by administering 0.3 ml isotonic saline daily for 7 days 5–10 min before dark onset. On the baseline days, the animals were injected ip with 10 ml/kg isotonic NaCl. On the test day, the animals received 50, 100 or 250 mg/kg nicotinic acid (Millipore Sigma) in a volume of 10 ml/kg (n = 6, n = 8, n = 7, respectively). The treatments were performed 5–10 min before dark onset. Sleep and telemetry recordings started at the onset of the dark phase and continued for 24 h.
Another group of mice (n = 5) was habituated to the gavage procedure by administering 0.3 ml water daily for 7 days 5–15 min before dark onset. After the habituation period, a baseline day was recorded after the oral gavage of 0.3 ml vehicle. The following day, 1 g/kg nicotinic acid was administered in the volume of 0.3 ml. The treatments were performed 5–10 min before dark onset. Sleep and telemetry recordings started at the onset of the dark phase and continued for 24 h.
Experiment 2: The effects of indomethacin pretreatment on the sleep-wake activity and body temperature-modulating effects of nicotinic acid in mice
A group of mice (n = 7) was habituated to the injection procedure as described above. On the baseline day, the animals were injected ip with 10 ml/kg isotonic NaCl. On the test day, the animals were pretreated with 5 mg/kg indomethacin subcutaneously 30 min prior the beginning of the dark phase, followed by 100 mg/kg nicotinic acid ip injection 5 minutes before dark onset. Sleep and telemetry recordings started at onset of the dark phase and continued for 24 h.
Experiment 3: The effects of β3-adrenergic receptor agonist, CL-316,243, pretreatment on the sleep-wake activity and body temperature-modulating effects of nicotinic acid in mice
A group of mice (n = 7) was habituated to the injection procedure as described above. On the baseline days, the animals were injected with 10 ml/kg isotonic NaCl ip. On the test day, the animals were pretreated with subcutaneous 0.2 mg/kg CL-316,243 (Millipore Sigma) 30 min prior the beginning of the dark phase, followed by ip 250 mg/kg nicotinic acid injection 5 minutes before dark onset. Sleep and telemetry recordings started at onset of the dark phase and continued for 24 h.
Experiment 4: The effects of ip administration of nicotinamide in mice
Two doses of nicotinamide were tested in the same group of mice (n = 6). After the habituation period, the animals received 10 ml/kg isotonic NaCl ip to obtain baseline values. On the test day, the mice were injected with 100 mg/kg nicotinamide ip. Two days later, a new baseline day was recorded followed by the test day of 250 mg/kg nicotinamide. The treatments were performed 5–10 min before dark onset. Sleep and telemetry recordings started at onset of the dark phase and continued for 24 h.
Experiment 5: The effects of ip administration of monomethyl fumarate in mice
Ten male mice were habituated to the injection procedure by daily administration of isotonic saline 5–10 min before dark onset. On the baseline days, the animals were injected ip with 10 ml/kg isotonic NaCl. On the test day, the animals received 20 or 50 mg/kg monomethyl fumarate in a volume of 10 ml/kg (n = 8, for both; five mice were injected with both doses of MMF). The treatments were at least two days apart and performed 5–10 min before dark onset. Sleep and telemetry recordings started at onset of the dark phase and continued for 24 h.
Experiment 6: The effects of nicotinic acid on energy expenditure and food intake in mice
Oxygen consumption (VO2, ml/kg/h) and RER were measured via indirect calorimetry; food intake was measured simultaneously by an automated system (Oxymax System, Columbus Instruments, Columbus, OH). Animals were habituated to the calorimetry cages for at least three days before the recordings. On the baseline day, vehicle was administered, on the test days 50 mg/kg (n = 4) or 250 mg/kg (n = 4) nicotinic acid was injected ip, or 1 g/kg nicotinic acid was administered via oral gavage (n = 4). All treatments were performed 5–10 min before the onset of the dark period.
Statistics
Time spent in wakefulness, NREMS and REMS, as well as body temperature and motor activity, VO2 and RER were calculated in 1-h blocks, and EEG SWA was calculated in 2-h blocks. Two-way repeated measures ANOVA was performed across 24 h between test days and the corresponding baselines (factors: treatment and time, both repeated). When appropriate, Tukey’s HSD test was applied post hoc. An α-level of P < 0.05 was considered to be significant.
Acknowledgements
We thank the excellent technical help of Ashley Massie. This work was supported by the National Institute of Health, National Heart, Lung, and Blood Institute, grant number R01HL122390.
Author contributions
É.S. & L.K. conceived, designed and conducted the experiments, analyzed the data and wrote the manuscript. All authors reviewed the manuscript.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Danguir J, Nicolaidis S. Sleep and feeding patterns in the ventromedial hypothalamic lesioned rat. Physiol. Behav. 1978;21:769–777. doi: 10.1016/0031-9384(78)90017-3. [DOI] [PubMed] [Google Scholar]
- 2.Danguir J. Cafeteria diet promotes sleep in rats. Appetite. 1987;8:49–53. doi: 10.1016/S0195-6663(87)80026-0. [DOI] [PubMed] [Google Scholar]
- 3.Shemyakin A, Kapás L. L-364,718, a cholecystokinin-A receptor antagonist, suppresses feeding-induced sleep in rats. Am. J. Physiol Regul. Integr. Comp. Physiol. 2001;280:R1420–R1426. doi: 10.1152/ajpregu.2001.280.5.R1420. [DOI] [PubMed] [Google Scholar]
- 4.Guan Z, Vgontzas AN, Bixler EO, Fang J. Sleep is increased by weight gain and decreased by weight loss in mice. Sleep. 2008;31:627–633. doi: 10.1093/sleep/31.5.627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Esposito M, Pellinen J, Kapás L, Szentirmai É. Impaired wake-promoting mechanisms in ghrelin receptor-deficient mice. Eur. J. Neurosci. 2012;35:233–243. doi: 10.1111/j.1460-9568.2011.07946.x. [DOI] [PubMed] [Google Scholar]
- 6.Yamanaka A, et al. Hypothalamic orexin neurons regulate arousal according to energy balance in mice. Neuron. 2003;38:701–713. doi: 10.1016/S0896-6273(03)00331-3. [DOI] [PubMed] [Google Scholar]
- 7.Gelegen C, Collier DA, Campbell IC, Oppelaar H, Kas MJ. Behavioral, physiological, and molecular differences in response to dietary restriction in three inbred mouse strains. Am. J. Physiol. Endocrinol. Metab. 2006;291:E574–E581. doi: 10.1152/ajpendo.00068.2006. [DOI] [PubMed] [Google Scholar]
- 8.Nicolaidis S. Metabolic mechanism of wakefulness (and hunger) and sleep (and satiety): Role of adenosine triphosphate and hypocretin and other peptides. Metabolism. 2006;55:S24–S29. doi: 10.1016/j.metabol.2006.07.009. [DOI] [PubMed] [Google Scholar]
- 9.Kapás L, et al. Effects of nocturnal intraperitoneal administration of cholecystokinin in rats: simultaneous increase in sleep, increase in EEG slow-wave activity, reduction of motor activity, suppression of eating, and decrease in brain temperature. Brain Res. 1988;438:155–164. doi: 10.1016/0006-8993(88)91334-0. [DOI] [PubMed] [Google Scholar]
- 10.Kapás L, et al. Somnogenic, pyrogenic, and anorectic activities of tumor necrosis factor-α and TNF-α fragments. Am. J. Physiol. 1992;263:R708–R715. doi: 10.1152/ajpcell.1992.263.3.C708. [DOI] [PubMed] [Google Scholar]
- 11.Hagan JJ, et al. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc. Natl. Acad. Sci. USA. 1999;96:10911–10916. doi: 10.1073/pnas.96.19.10911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Szentirmai É, Hajdu I, Obal F, Jr., Krueger JM. Ghrelin-induced sleep responses in ad libitum fed and food-restricted rats. Brain Res. 2006;1088:131–140. doi: 10.1016/j.brainres.2006.02.072. [DOI] [PubMed] [Google Scholar]
- 13.Szentirmai É, Kapás L, Krueger JM. Ghrelin microinjection into forebrain sites induces wakefulness and feeding in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007;292:R575–R585. doi: 10.1152/ajpregu.00448.2006. [DOI] [PubMed] [Google Scholar]
- 14.Kukorelli T, Juhasz G. Electroencephalographic synchronization induced by stimulation of small intestine and splanchnic nerve in cats. Electroencephalogr. Clin. Neurophysiol. 1976;41:491–500. doi: 10.1016/0013-4694(76)90061-4. [DOI] [PubMed] [Google Scholar]
- 15.Szentirmai É, Kapás L. Intact brown adipose tissue thermogenesis is required for restorative sleep responses after sleep loss. Eur. J. Neurosci. 2014;39:984–998. doi: 10.1111/ejn.12463. [DOI] [PubMed] [Google Scholar]
- 16.Szentirmai É, Kapás L. The role of the brown adipose tissue in β3-adrenergic receptor activation-induced sleep, metabolic and feeding responses. Sci. Rep. 2017;7:958. doi: 10.1038/s41598-017-01047-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.El Hajjaji FZ, et al. Sleep structure and feeding pattern changes induced by the liver’s thermal status in the rat. J. Sleep Res. 2012;21:204–211. doi: 10.1111/j.1365-2869.2011.00973.x. [DOI] [PubMed] [Google Scholar]
- 18.Ehlen, J. C. et al. Bmal1 function in skeletal muscle regulates sleep. Elife. 6 (2017). [DOI] [PMC free article] [PubMed]
- 19.Zu L, et al. Bacterial endotoxin stimulates adipose lipolysis via toll-like receptor 4 and extracellular signal-regulated kinase pathway. J. Biol. Chem. 2009;284:5915–5926. doi: 10.1074/jbc.M807852200. [DOI] [PubMed] [Google Scholar]
- 20.Chaves VE, Junior FM, Bertolini GL. The metabolic effects of growth hormone in adipose tissue. Endocrine. 2013;44:293–302. doi: 10.1007/s12020-013-9904-3. [DOI] [PubMed] [Google Scholar]
- 21.Grunfeld C, Feingold KR. Regulation of lipid metabolism by cytokines during host defense. Nutrition. 1996;12:S24–S26. doi: 10.1016/0899-9007(96)90013-1. [DOI] [PubMed] [Google Scholar]
- 22.Carpene C, et al. Desensitization of beta-1 and beta-2, but not beta-3, adrenoceptor-mediated lipolytic responses of adipocytes after long-term norepinephrine infusion. J. Pharmacol. Exp. Ther. 1993;265:237–247. [PubMed] [Google Scholar]
- 23.Key BJ, Marley DE. The effect of the sympathomimetic amines on behaviour and electrocortical activity of the chicken. Electroencephalogr. Clin. Neurophysiol. 1962;14:90–105. doi: 10.1016/0013-4694(62)90011-1. [DOI] [PubMed] [Google Scholar]
- 24.Tikal K. The quantitative evaluation and some aspects of catecholamine-induced sleep. Act. Nerv. Super. (Praha) 1969;11:136–140. [PubMed] [Google Scholar]
- 25.Danguir J, Nicolaidis S. Circadian sleep and feeding patterns in the rat: possible dependence on lipogenesis and lipolysis. Am. J. Physiol. 1980;238:E223–E230. doi: 10.1152/ajpendo.1980.238.3.E223. [DOI] [PubMed] [Google Scholar]
- 26.Szentirmai É, Kapás L. Sleep and body temperature in TNFα knockout mice: The effects of sleep deprivation, β3-AR stimulation and exogenous TNFalpha. Brain Behav. Immun. 2019;81:260–271. doi: 10.1016/j.bbi.2019.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Obal F, Jr., Krueger JM. GHRH and sleep. Sleep Med. Rev. 2004;8:367–377. doi: 10.1016/j.smrv.2004.03.005. [DOI] [PubMed] [Google Scholar]
- 28.Krueger JM, Majde JA. Microbial products and cytokines in sleep and fever regulation. Crit. Rev. Immunol. 2017;37:291–315. doi: 10.1615/CritRevImmunol.v37.i2-6.70. [DOI] [PubMed] [Google Scholar]
- 29.Roky R, Kapás L, Taishi TP, Fang J, Krueger JM. Food restriction alters the diurnal distribution of sleep in rats. Physiol. Behav. 1999;67:697–703. doi: 10.1016/S0031-9384(99)00137-7. [DOI] [PubMed] [Google Scholar]
- 30.Szentirmai É, Kapás L, Sun Y, Smith RG, Krueger JM. Restricted feeding-induced sleep, activity, and body temperature changes in normal and preproghrelin-deficient mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010;298:R467–R477. doi: 10.1152/ajpregu.00557.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Carlson LA. Nicotinic acid: the broad-spectrum lipid drug. A 50th anniversary review. J. Intern. Med. 2005;258:94–114. doi: 10.1111/j.1365-2796.2005.01528.x. [DOI] [PubMed] [Google Scholar]
- 32.Ginsberg HN, Reyes-Soffer G. Niacin: a long history, but a questionable future. Curr. Opin. Lipidol. 2013;24:475–479. doi: 10.1097/MOL.0000000000000017. [DOI] [PubMed] [Google Scholar]
- 33.Morrow JD, Parsons WG, III, Roberts LJ. Release of markedly increased quantities of prostaglandin D in vivo in humans following the administration of nicotinic acid. Prostaglandins. 1989;38:263–274. doi: 10.1016/0090-6980(89)90088-9. [DOI] [PubMed] [Google Scholar]
- 34.Cheng K, et al. Antagonism of the prostaglandin D receptor 1 suppresses nicotinic acid-induced vasodilation in mice and humans. Proc. Natl. Acad. Sci. USA. 2006;103:6682–6687. doi: 10.1073/pnas.0601574103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Urade Y, Hayaishi O. Prostaglandin D and sleep/wake regulation. Sleep Med. Rev. 2011;15:411–418. doi: 10.1016/j.smrv.2011.08.003. [DOI] [PubMed] [Google Scholar]
- 36.Rahman M, et al. The beta-hydroxybutyrate receptor HCA2 activates a neuroprotective subset of macrophages. Nat. Commun. 2014;5:3944. doi: 10.1038/ncomms4944. [DOI] [PubMed] [Google Scholar]
- 37.Yang R, He J, Wang Y. Activation of the niacin receptor HCA2 reduces demyelination and neurofilament loss, and promotes functional recovery after spinal cord injury in mice. Eur. J. Pharmacol. 2016;791:124–136. doi: 10.1016/j.ejphar.2016.08.020. [DOI] [PubMed] [Google Scholar]
- 38.Wakade C, et al. Niacin modulates macrophage polarization in Parkinson’s disease. J. Neuroimmunol. 2018;320:76–79. doi: 10.1016/j.jneuroim.2018.05.002. [DOI] [PubMed] [Google Scholar]
- 39.Massie A, Boland E, Kapás L, Szentirmai É. Mice lacking alternatively activated (M2) macrophages show impairments in restorative sleep after sleep loss and in cold environment. Sci. Rep. 2018;8:8625. doi: 10.1038/s41598-018-26758-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Irie M, Sakuma M, Tsushima T, Shizume K, Nakao K. Effect of nicotinic acid administration on plasma growth hormone concentrations. Proceedings of the Society for Experimental Biology and Medicine. 1967;126:708–711. doi: 10.3181/00379727-126-32549. [DOI] [PubMed] [Google Scholar]
- 41.Quabbe HJ, Ramek W, Luyckx AS. Growth hormone, glucagon, and insulin response to depression of plasma free fatty acids and the effect of glucose infusion. J. Clin. Endocrinol. Metab. 1977;44:383–391. doi: 10.1210/jcem-44-2-383. [DOI] [PubMed] [Google Scholar]
- 42.Szentirmai É, Millican NS, Massie AR, Kapás L. Butyrate, a metabolite of intestinal bacteria, enhances sleep. Sci. Rep. 2019;9:7035. doi: 10.1038/s41598-019-43502-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Collins PB, Chaykin S. The management of nicotinamide and nicotinic acid in the mouse. J. Biol. Chem. 1972;247:778–783. [PubMed] [Google Scholar]
- 44.DiPalma JR, Thayer WS. Use of niacin as a drug. Annu. Rev. Nutr. 1991;11:169–187. doi: 10.1146/annurev.nu.11.070191.001125. [DOI] [PubMed] [Google Scholar]
- 45.Petrack B, Greengard P, Kalinsky H. On the relative efficacy of nicotinamide and nicotinic acid as precursors of nicotinamide adenine dinucleotide. J. Biol. Chem. 1966;241:2367–2372. [PubMed] [Google Scholar]
- 46.Chevillard L, Giono H, Laury MC. [Comparative effects of some vasodilators on the body temperature and the respiratory metabolism of guinea pigs] C. R. Seances Soc. Biol. Fil. 1958;152:1074–1077. [PubMed] [Google Scholar]
- 47.Turenne SD, Seeman M, Ross BM. An animal model of nicotinic-acid-induced vasodilation: effect of haloperidol, caffeine and nicotine upon nicotinic acid response. Schizophr. Res. 2001;50:191–197. doi: 10.1016/S0920-9964(00)00082-7. [DOI] [PubMed] [Google Scholar]
- 48.Carlson LA, Oro L, Ostman J. Effect of a single dose of nicotinic acid on plasma lipids in patients with hyperlipoproteinemia. Acta Med. Scand. 1968;183:457–465. doi: 10.1111/j.0954-6820.1968.tb10508.x. [DOI] [PubMed] [Google Scholar]
- 49.Eriksson HE, Norrman SR, Wistrand PJ. Detection of drugs with peripheral vascular effects similar to those of nicotinic acid. J. Pharm. Pharmacol. 1969;21:192–193. doi: 10.1111/j.2042-7158.1969.tb08226.x. [DOI] [PubMed] [Google Scholar]
- 50.Walters RW, et al. β-Arrestin1 mediates nicotinic acid-induced flushing, but not its antilipolytic effect, in mice. J. Clin. Invest. 2009;119:1312–1321. doi: 10.1172/JCI36806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gordon CJ. The mouse thermoregulatory system: Its impact on translating biomedical data to humans. Physiol. Behav. 2017;179:55–66. doi: 10.1016/j.physbeh.2017.05.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Estler CJ, Ammon HP, Lang B. Ineffectiveness of nicotinic acid to impair thermoregulation in cold-exposed mice. Eur. J. Pharmacol. 1970;9:257–260. doi: 10.1016/0014-2999(70)90309-2. [DOI] [PubMed] [Google Scholar]
- 53.Lafrance L, Routhier D, Tetu B, Tetu C. Effects of noradrenaline and nicotinic acid on plasma free fatty acids and oxygen consumption in cold-adapted rats. Can. J. Physiol. Pharmacol. 1979;57:725–730. doi: 10.1139/y79-109. [DOI] [PubMed] [Google Scholar]
- 54.Even P, Coulaud H, Nicolaidis S. Integrated metabolic control of food intake after 2-deoxy-D-glucose and nicotinic acid injection. Am. J. Physiol. 1988;255:R82–R89. doi: 10.1152/ajpregu.1988.255.1.R82. [DOI] [PubMed] [Google Scholar]
- 55.Hepler C, Foy CE, Higgins MR, Renquist BJ. The hypophagic response to heat stress is not mediated by GPR109A or peripheral β-OH butyrate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016;310:R992–R998. doi: 10.1152/ajpregu.00513.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Parmeggiani PL. Thermoregulation and sleep. Front. Biosci. 2003;8:s557–s567. doi: 10.2741/1054. [DOI] [PubMed] [Google Scholar]
- 57.Kroeger D, et al. Galanin neurons in the ventrolateral preoptic area promote sleep and heat loss in mice. Nat. Commun. 2018;9:4129. doi: 10.1038/s41467-018-06590-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Everson CA, Szabo A. Repeated exposure to severely limited sleep results in distinctive and persistent physiological imbalances in rats. PLoS. One. 2011;6:e22987. doi: 10.1371/journal.pone.0022987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Martins PJ, Fernandes L, de Oliveira AC, Tufik S, D’Almeida V. Type of diet modulates the metabolic response to sleep deprivation in rats. Nutr. Metab. (Lond) 2011;8:86. doi: 10.1186/1743-7075-8-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gilbert SS, van den Heuvel CJ, Ferguson SA, Dawson D. Thermoregulation as a sleep signalling system. Sleep Med. Rev. 2004;8:81–93. doi: 10.1016/S1087-0792(03)00023-6. [DOI] [PubMed] [Google Scholar]
- 61.Soga T, et al. Molecular identification of nicotinic acid receptor. Biochem. Biophys. Res. Commun. 2003;303:364–369. doi: 10.1016/S0006-291X(03)00342-5. [DOI] [PubMed] [Google Scholar]
- 62.Tunaru S, et al. PUMA-G and HM74 are receptors for nicotinic acid and mediate its anti-lipolytic effect. Nat. Med. 2003;9:352–355. doi: 10.1038/nm824. [DOI] [PubMed] [Google Scholar]
- 63.Wise A, et al. Molecular identification of high and low affinity receptors for nicotinic acid. J. Biol. Chem. 2003;278:9869–9874. doi: 10.1074/jbc.M210695200. [DOI] [PubMed] [Google Scholar]
- 64.Tang H, Lu JY, Zheng X, Yang Y, Reagan JD. The psoriasis drug monomethylfumarate is a potent nicotinic acid receptor agonist. Biochem. Biophys. Res. Commun. 2008;375:562–565. doi: 10.1016/j.bbrc.2008.08.041. [DOI] [PubMed] [Google Scholar]
- 65.Benyo Z, et al. GPR109A (PUMA-G/HM74A) mediates nicotinic acid-induced flushing. J. Clin. Invest. 2005;115:3634–3640. doi: 10.1172/JCI23626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Maciejewski-Lenoir D, et al. Langerhans cells release prostaglandin D in response to nicotinic acid. J. Invest. Dermatol. 2006;126:2637–2646. doi: 10.1038/sj.jid.5700586. [DOI] [PubMed] [Google Scholar]
- 67.Hanson J, et al. Nicotinic acid- and monomethyl fumarate-induced flushing involves GPR109A expressed by keratinocytes and COX-2-dependent prostanoid formation in mice. J. Clin. Invest. 2010;120:2910–2919. doi: 10.1172/JCI42273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ek M, Arias C, Sawchenko P, Ericsson-Dahlstrand A. Distribution of the EP3 prostaglandin E(2) receptor subtype in the rat brain: relationship to sites of interleukin-1-induced cellular responsiveness. J. Comp. Neurol. 2000;428:5–20. doi: 10.1002/1096-9861(20001204)428:1<5::AID-CNE2>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 69.Zhang J, Rivest S. Distribution, regulation and colocalization of the genes encoding the EP2- and EP4-PGE receptors in the rat brain and neuronal responses to systemic inflammation. Eur. J. Neurosci. 1999;11:2651–2668. doi: 10.1046/j.1460-9568.1999.00682.x. [DOI] [PubMed] [Google Scholar]
- 70.Maher SA, et al. Prostaglandin D and the role of the DP1, DP2 and TP receptors in the control of airway reflex events. Eur. Respir. J. 2015;45:1108–1118. doi: 10.1183/09031936.00061614. [DOI] [PubMed] [Google Scholar]
- 71.Matsumoto S, et al. Prostaglandin E -induced modification of tetrodotoxin-resistant Na+ currents involves activation of both EP2 and EP4 receptors in neonatal rat nodose ganglion neurones. Br. J. Pharmacol. 2005;145:503–513. doi: 10.1038/sj.bjp.0706212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Smith JA, Amagasu SM, Eglen RM, Hunter JC, Bley KR. Characterization of prostanoid receptor-evoked responses in rat sensory neurones. Br. J. Pharmacol. 1998;124:513–523. doi: 10.1038/sj.bjp.0701853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Dell P. Afferences baroreceptives, phases de synchronisation corticale et sommeil. Archives italiennes de biologie. 1973;111:553–563. [PubMed] [Google Scholar]
- 74.Chase MH, Nakamura Y, Clemente CD, Sterman MB. Afferent vagal stimulation: neurographic correlates of induced EEG synchronization and desynchronization. Brain Res. 1967;5:236–249. doi: 10.1016/0006-8993(67)90089-3. [DOI] [PubMed] [Google Scholar]
- 75.Kapás L, Hansen MK, Chang HY, Krueger JM. Vagotomy attenuates but does not prevent the somnogenic and febrile effects of lipopolysaccharide in rats. Am. J. Physiol. 1998;274:R406–R411. doi: 10.1152/ajpcell.1998.274.2.C406. [DOI] [PubMed] [Google Scholar]
- 76.Zielinski MR, Dunbrasky DL, Taishi P, Souza G, Krueger JM. Vagotomy attenuates brain cytokines and sleep induced by peripherally administered tumor necrosis factor-α and lipopolysaccharide in mice. Sleep. 2013;36:1227–1238. doi: 10.5665/sleep.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Spector R. Niacin and niacinamide transport in the central nervous system. In vivo studies. J. Neurochem. 1979;33:895–904. doi: 10.1111/j.1471-4159.1979.tb09919.x. [DOI] [PubMed] [Google Scholar]
- 78.Hankes LV, et al. Blood brain barrier blocks transport of carboxyl- C-nicotinic acid into human brain. Eur. J. Nucl. Med. 1988;14:53–54. doi: 10.1007/BF00252620. [DOI] [PubMed] [Google Scholar]
- 79.Hankes LV, et al. Effect of Huntington’s and Alzheimer’s diseases on the transport of nicotinic acid or nicotinamide across the human blood-brain barrier. Adv. Exp. Med. Biol. 1991;294:675–678. doi: 10.1007/978-1-4684-5952-4_91. [DOI] [PubMed] [Google Scholar]
- 80.Carlson LA, Oro L. The effect of nicotinic acid on the plasma free fatty acids. Acta Medica Scandinavica. 1962;172:641–645. doi: 10.1111/j.0954-6820.1962.tb07203.x. [DOI] [PubMed] [Google Scholar]
- 81.Pinter EJ, Pattee CJ. Biphasic nature of blood glucose and free fatty acid changes following intravenous nicotinic acid in man. J. Clin. Endocrinol. Metab. 1967;27:440–443. doi: 10.1210/jcem-27-3-440. [DOI] [PubMed] [Google Scholar]
- 82.Wang W, Basinger A, Neese RA, Christiansen M, Hellerstein MK. Effects of nicotinic acid on fatty acid kinetics, fuel selection, and pathways of glucose production in women. Am. J. Physiol. Endocrinol. Metab. 2000;279:E50–E59. doi: 10.1152/ajpendo.2000.279.1.E50. [DOI] [PubMed] [Google Scholar]
- 83.Pereira JN. The plasma free fatty acid rebound induced by nicotinic acid. J. Lipid Res. 1967;8:239–244. [PubMed] [Google Scholar]
- 84.Subissi A, Murmann W. Correlation between the plasma concentration of free nicotinic acid and some of its pharmacological effects in the fasted rat after an oral dose of sorbinicate and of nicotinic acid. Arzneimittelforschung. 1978;28:1143–1145. [PubMed] [Google Scholar]
- 85.Kaijser L, Eklund B, Olsson AG, Carlson LA. Dissociation of the effects of nicotinic acid on vasodilatation and lipolysis by a prostaglandin synthesis inhibitor, indomethacin, in man. Med. Biol. 1979;57:114–117. [PubMed] [Google Scholar]
- 86.Lukasova M, Hanson J, Tunaru S, Offermanns S. Nicotinic acid (niacin): new lipid-independent mechanisms of action and therapeutic potentials. Trends Pharmacol. Sci. 2011;32:700–707. doi: 10.1016/j.tips.2011.08.002. [DOI] [PubMed] [Google Scholar]
- 87.van der Hoorn JW, et al. Niacin increases HDL by reducing hepatic expression and plasma levels of cholesteryl ester transfer protein in APOE*3Leiden.CETP mice. Arterioscler. Thromb. Vasc. Biol. 2008;28:2016–2022. doi: 10.1161/ATVBAHA.108.171363. [DOI] [PubMed] [Google Scholar]
- 88.McKenney JM, Proctor JD, Harris S, Chinchili VM. A comparison of the efficacy and toxic effects of sustained- vs immediate-release niacin in hypercholesterolemic patients. JAMA. 1994;271:672–677. doi: 10.1001/jama.1994.03510330050033. [DOI] [PubMed] [Google Scholar]
- 89.Davignon J, et al. Comparative efficacy and safety of pravastatin, nicotinic acid and the two combined in patients with hypercholesterolemia. Am. J. Cardiol. 1994;73:339–345. doi: 10.1016/0002-9149(94)90005-1. [DOI] [PubMed] [Google Scholar]
- 90.Gibbons LW, Gonzalez V, Gordon N, Grundy S. The prevalence of side effects with regular and sustained-release nicotinic acid. Am. J. Med. 1995;99:378–385. doi: 10.1016/S0002-9343(99)80185-5. [DOI] [PubMed] [Google Scholar]
- 91.Plaisance EP, et al. Postprandial triglyceride responses to aerobic exercise and extended-release niacin. Am. J. Clin Nutr. 2008;88:30–37. doi: 10.1093/ajcn/88.1.30. [DOI] [PubMed] [Google Scholar]
- 92.Szentirmai É, Krueger JM. Sickness behaviour after lipopolysaccharide treatment in ghrelin deficient mice. Brain Behav. Immun. 2014;36:200–206. doi: 10.1016/j.bbi.2013.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.