

Abstract
Background:
Bisphenol A (BPA) is a synthetic plasticizer that is commonly used in the pro-duction of polycarbonate plastics and epoxy resins. Human exposure occurs when BPA migrates from food and beverage containers into the contents when heated or even under normal conditions of use. BPA exerts endocrine disruptor action due to its weak binding affinity for the estrogen receptors ERα and ERβ. BPA exerts other effects by activating the membrane receptor GPER (GPR30) and/or other receptors such as the estrogen-related receptors (ERRs).
Objective:
This review summarizes emerging data on BPA and cancer. These include data linking expo-sure to BPA with an increased risk of hormone-related cancers such as those of the ovary, breast, pros-tate, and even colon cancer. BPA can also induce resistance to various chemotherapeutics such as doxoru-bicin, cisplatin, and vinblastine in vitro. The development of chemoresistance to available therapeutics is an emerging significant aspect of BPA toxicity because it worsens the prognosis of many tumors.
Conclusion:
Recent findings support a causal role of BPA at low levels in the development of cancers and in dictating their response to cytotoxic therapy. Accurate knowledge and consideration of these issues would be highly beneficial to cancer prevention and management.
Keywords: Bisphenol A (BPA), Breast cancer, Chemotherapy, Colorectal cancer, Ovarian cancer, Prostate cancer
1. Introduction
Bisphenol A (BPA) is widely used in many consumer products, epoxy resins, and polycarbonate plastics such as baby bottles and the lining of beverage cans. However, BPA can leach from the polymers into food and water upon exposure to elevated temperature and hence accumulates in the human body. Harsh detergents can also cause BPA to leach into the food and beverages due to the changes in pH, which can hydrolyze the ester bond linking BPA monomers [1-3]. Biomonitoring studies have demonstrated the presence of low doses of BPA in human urine, blood, saliva, and milk [4]. Moreover, epoxy resin BPA can pass through the placenta causing miscarriage and can alter the ovarian cycle in females. The Reference Dose (RfD) established by the U.S. Environmental Protection Agency (EPA) indicates that less than 1 µg kg–1 intake of BPA per day is considered to be safe for humans. However, recent studies demonstrated that as little as 0.2 µg kg–1 administration of BPA can cause serious consequences such as lower sperm production and reduced fertility in male animals; hence it could exert these and other adverse effects on human health as well [5-7].
BPA is classified as an Endocrine Disrupting Chemical (EDC) because it can act as a xenoestrogen. EDCs interfere and prevent the binding of natural hormones to their receptors and/or can act as a hormone mimics; consequently, they exaggerate the effects of endogenous hormones [2, 8]. Researchers have also found that exposure to BPA is associated with an increased risk of insulin resistance, type 2 diabetes, and alteration of normal immune response resulting in high levels of Immunoglobulin E (IgE) and interleukin-4 potentially leading to allergies such as hay fever. BPA can also alter cardiac function leading to thickening of the left ventricular wall. In addition, many of the estrogen-related pathways can be influenced by BPA because its structure is similar to estradiol; consequently, it can cause numerous hormonal diseases including sexual behavior problems and accelerated puberty [7, 9, 10].
BPA can increase the susceptibility to tumorigenesis perhaps because of its endocrine-disrupting effect and similarity to estrogen. For example, Pupo et al. (2012) demonstrated that BPA can induce expression of G-protein coupled receptor target genes to activate the G Protein-coupled Estrogen Receptor 1 (GPER; formerly known as, GPR30, G-protein coupled receptor 30)/Epidermal Growth Factor Receptor (EGFR)/Extracellular Signal-Regulated Kinase (ERK)1/2 signaling pathway in cancer cells [11]. It has also been reported that BPA is directly genotoxic in human lymphocytes through induction of DNA double-strand breaks [12, 13]. Moreover, BPA can damage DNA and interfere with many cancer-related signaling pathways [8, 14, 15]. Low doses of BPA can also induce resistance in many types of cancer cells to classical chemotherapeutics such as doxorubicin, cisplatin, carboplatin, tamoxifen (TAM), bevacizumab, PARP inhibitors, vinblastine and other drugs both in vitro and in vivo (Table 1). Conversely, BPA can induce cell cycle inhibition in prostate cancer cells by activating androgen receptor (AR) and pathways involving estrogen receptor beta (ERβ)/EGFR complexes, ERK, and p53 [16]. Thus, BPA is a major threat to human health and other species. This review will highlight the latest findings linking BPA to cancer with a focus on the molecular mechanisms of this action and the emerging role of BPA in response to cytotoxic therapy.
Table 1. The association of BPA with resistance to therapy and disease outcome in cancer.
| Drug Resistance or other Effects | Mechanisms | Study Model | Evidence* | References | |
|---|---|---|---|---|---|
| Breast Cancer | Tamoxifen (TAM); Rapamycin | Evasion of apoptosis via downregulation of p53, p21 and BAX; activation of mTOR pathway; increased ERα/ERβ ratio | in vitro | direct | Dairkee et al. (2013) [17] |
| Tamoxifen (TAM) | Activation of ERK/MAPK results in upregulation of ERRγ and its nuclear localization | in vitro | indirect | Hackler et al. (2014); Song et al. (2015) [18,19] | |
| Lapatinib (EGFR inhibitor) | Activation of EGFR/ERK1/2 pathway and increasing the level of anti-apoptotic proteins | in vitro | direct | Sauer et al. (2017) [20] | |
| Doxorubicin; Vinblastine | Increased expression of antiapoptotic proteins is a potential mechanism, independent of classical ERs | in vitro | direct | LaPensee et al. (2009) [21] | |
| Cisplatin | Increased expression of antiapoptotic protein BCL2, independent of classical ERs | in vitro | direct | LaPensee et al. (2009; 2010) [21,22] | |
| Poor prognosis | BPA up-regulates HOXB9; and HOXB9 is associated with poor disease-free survival and overall survival in breast cancer patients. | Clinical; in vitro |
indirect | Deb et al. (2016); Seki et al. (2012) [23,24] | |
| Colorectal Cancer | Doxorubicin | Overexpression of fascin; activation of PI3k/Akt pathway helping cancer cells to evade apoptosis through suppression of pro-apoptotic caspase 9 and caspase 3 | in vivo; in vitro | indirect | Qualtrough et al. (2009); Chan et al. (2010); Chen et al. (2015) [25-27] |
| Prostate Cancer | Androgen deprivation therapy (ADT) | BPA activates mutant AR (AR-T877A) which results in dimerization of mutant AR and its dissociation from heat shock protein; localization to the nucleus and upregulation of target genes (PSA) and possibly downregulation of ERβ | in vitro | direct | Hess-Wilson (2009); Wetherill et al. (2005) [28,29] |
Footnote: *direct evidence is obtained from direct induction of resistance by BPA in the used model while indirect evidence refers to the combined evidence of activation of a molecular pathway by BPA in one publication and that this pathway is associated with chemoresistance in another setting. Other evidence derived indirectly from cells or tissues of different lineages are discussed in the main text.
2. Search Strategy
This review was mainly concerned with studies that tested the molecular effects of BPA on chemotherapy. Starting from August 15th 2018, the review team searched through “PubMed” and “Scopus” and collected references from 2005 to 2018 using the keywords “Bisphenol A” or “BPA” AND “Chemotherapy” or “Cancer Therapy”. The literature search was continued until the revision stage of this article on January 26th, 2019. The inclusion criteria were: 1. observational studies on the impact of BPA on chemotherapy and cancer treatment, 2. BPA measurement in blood and body fluids, 3. BPA exposure in patients suffering from breast, colon, ovarian, or prostate cancers and under drug treatment. The data excluded from this review were those that did not include or were not relevant to chemotherapy such as demographics of the population including location, age, and sex. The review team exported the literature into a reference library/database established in EndNote to manage references and citations. This review details the molecular effects of BPA and its role in chemoresistance to common chemotherapeutics such as cisplatin, tamoxifen, doxorubicin, lapatinib, and vinblastine.
3. BPA and Breast Cancer
Breast cancer continues to be the most common invasive malignancy and a major cause of cancer-related death among women worldwide. Intriguingly, in spite of the major advances in diagnostics and treatment of cancer, the incidence of breast cancer has failed to decline or is even increasing in some areas [30, 31]. Several factors contribute to the etiology of breast cancer in females. The most significant factors are reproductive, environmental, and lifestyle.
Molecular biology and genetics play a key role in breast cancer [32, 33]. Many observational epidemiological studies have suggested that constant exposure to estrogen signals is a major risk factor for the development of breast cancer. Late menopause, late pregnancy, early menarche, and lack of breastfeeding to the newborn baby are also reproductive factors that reflect increased or unbalanced estrogen exposure and therefore may account for the development of breast cancer [34]. Estrogen exerts its effects essentially via nuclear estrogen receptor alpha (ERα) and beta (ERβ); both of these act as DNA-binding transcription factors. Estrogen also affects the membrane estrogen receptors such as GPER/GPR30. Environmental exposure to chemical toxicants is a major concern because it might explain the currently increasing incidence of breast cancer worldwide. In particular, an accumulating body of evidence has led many scientists to believe that exposure to xenoestrogens during critical windows of mammary gland development is associated with an increased risk of breast cancer later in life [14, 35-38]. The following discussion will highlight the role of BPA in the development of chemoresistance in breast cancer as summarized in (Fig. 1).
Fig. (1).
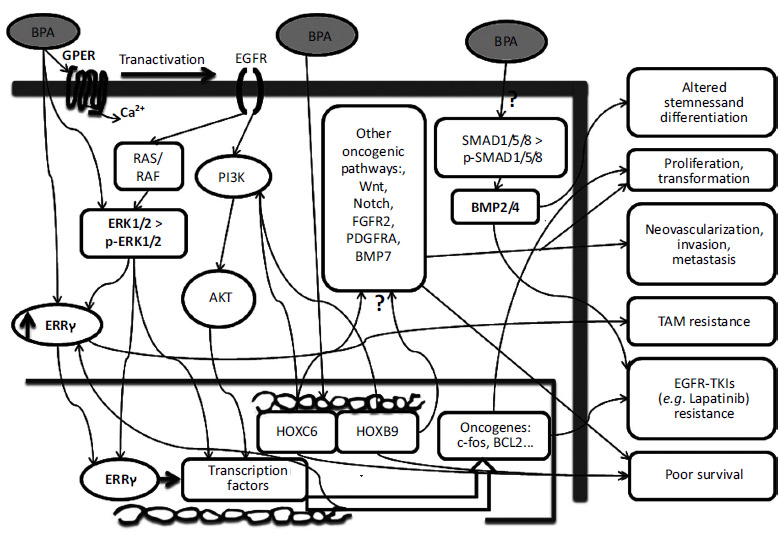
Major signaling pathways that link Bisphenol A (BPA) to breast cancer development and chemo-resistance. Exposure to BPA can act through ERK1/2/ERRγ signals to trigger the proliferation of breast cancer cells and TAM resistance. Binding of BPA to ERRγ results in its nuclear translocation and activation of transcription factors such as AP1 which mediates the above action. BPA can activate ERK1/2 through GPER/EGFR pathway which consequently activates transcription and upregulation of oncogenes such as c-fos and BCL2. This action also results in cell proliferation and resistance to EGFR-TKIs drugs such as Lapatinib. BPA can induce expression of the HOXC6 and HOXB9 genes through putative estrogen response element in their promoters. Both of these genes are associated with multiple oncogenic pathways and with poor disease-free survival in breast and/or other cancers. BPA may alter BMP2/4 signaling via the phosphorylation of SMAD1/5/8. Altered BMP signaling affects stem cell differentiation and induces resistance to chemotherapy. BPA, Bisphenol A; ERK1/2, extracellular signal-regulated kinase 1/2; ERRγ, estrogen-related-receptors-gamma; TAM, tamoxifen; AP1, Activator Protein 1; GPER, G protein-coupled estrogen receptor 1; HOXC6/HOXB9, Homeobox containing genes C6 and B9; BMP2/4, Bone morphogenetic proteins 2 and 4; SMAD1/5/8, small mothers against decapentaplegic1/5/8; Question mark indicates lack of details; upward arrow indicates upregulation.
3.1. BPA is Associated with Chemo-Resistance in Breast Cancer
Studies have demonstrated that BPA can accelerate mammary tumorigenesis and metastasis via multiple mechanisms. For example, low nanomolar doses of BPA, comparable to those that have been widely detected in human tissues, can significantly stimulate the proliferation of both Estrogen Receptor-negative (ER-) and Estrogen Receptor-positive (ER+) breast cancer cells [19]. Breast cancer patients with metastatic disease develop chemo-resistance to many drugs [21, 39]. TAM is a selective ER modulator to treat the ER+ breast cancers that account for most cases in both women and men. Alterations in the survival pathways in ER+ breast tumor cells can lead to the development of TAM resistance without the loss or mutation of ER itself. Alternatively, the high levels of oncogenic histone methyltransferase Enhancer of Zeste Homolog 2 (EZH2) conferred TAM resistance via regulation of ERα activity in response to the antagonist in ER+ breast cancer cells [40]. Dairekee et al. (2013) demonstrated that BPA induced evasion of TAM-induced apoptosis; BPA could also block the pro-apoptotic effects of rapamycin. BPA exerts this effect by suppressing signaling mediated by p53 and BAX pro-apoptotic proteins [17]. Furthermore, BPA can oppose the pro-apoptotic effects of the highly effective anti-cancer drugs cisplatin, vinblastine, and doxorubicin by increasing the expression of anti-apoptotic proteins including B-Cell Lymphoma 2 (BCL2) [21, 22].
3.2. Molecular Targets/Pathways of BPA Action
3.2.1. Estrogen-Related Receptors (ERRs)
The development and progression of breast cancer can occur via uncontrolled estrogenic signals as discussed above. A nuclear receptor superfamily called Estrogen-Related-Receptors (ERRs) has been characterized at the protein and RNA levels in breast and other types of cancer. They are called ERRs because of their sequence homology with estrogen receptors, but they do not appear to bind estrogens or other steroid hormones. Actually, they are often considered orphan receptors because no endogenous ligands have been identified [41].
Several studies have shown that ERRs can induce growth and proliferation in breast cancer cells and other cancers by altering the oncogene expression in these cells [18]. The ERR family contains three members: ERRα, ERRβ and ERRγ. The ERRα and ERRγ play significant roles as both transcriptional activators as well as cancer suppressors or promotors [41]. ERRγ strongly binds to BPA and can mediate BPA-stimulation effects. Successful silencing of ERRγ attenuated the BPA-induced proliferation of breast cancer cells [19]. The upregulation of ERRγ mRNA and protein expression and its nuclear localization by BPA was mediated by the phosphorylation of ERK1/2 [19]. In ER+ breast cancer cells, the ERRγ was preferentially expressed to promote TAM resistance [18]. Hence, BPA might have a role in TAM resistance because it can upregulate ERRγ mRNA and protein.
3.2.2. GPER/ERK1/2 Pathway
Previous studies have shown activation of ERK1/2 through GPER in human breast cancer cells after treatment with BPA. The phosphorylation of ERK1/2 by BPA was dependent upon the expression of GPER, but it was ER-independent. The ERK1/2 inhibitor PD98059 significantly blocked both the proliferation of ER+ breast cancer cells and upregulation of ERRγ by BPA [11, 42]. Interestingly, the expression of the proto-oncogene c-fos (a significant gene in the early estrogen response) was upregulated by BPA through the GPER/EGFR/ERK1/2 pathway [42]. Moreover, treatment of breast cancer cells with BPA upregulated the expression of pS2 protein, which contains an activator protein 1 (AP1) element in its promoter region and enhances the oncogenicity of mammary carcinoma. BPA induced the expression of AP1- target genes via the GPER/EGFR/ERK1/2 cascade as well [42].
Constant exposure to BPA and other EDCs induced resistance to an EGFR-targeted anti-cancer drug (Lapatinib) through activation of the EGFR/ ERK1/2 pathway and increasing the level of anti-apoptotic (BCL2) and anti-oxidant (superoxide dismutase 1, SOD1) proteins in breast cancer cells [20, 43].
3.2.3. Homeobox Containing Genes (HOX Genes)
Of the 39 homeobox-containing genes (HOX genes) that are associated with embryogenesis and postnatal development, some showed disrupted expression in a variety of cancers including breast cancer. The HOXC6 gene is involved in the development of mammary glands and production of milk; it is transcriptionally regulated by E2 via ERs [44]. HOXC6 is associated with signaling pathways such as PI3K/AKT, Wnt, and Notch as well as proteins that are key players in tumor proliferation and metastasis such as fibroblast growth factor receptor 2 (FGFR2), platelet-derived growth factor receptor alpha (PDGFRA), and bone morphogenetic protein 7 (BMP7). Hussain et al. (2015) showed that HOXC6 was overexpressed in breast cancer tissue and in ER+ breast cancer cell lines. The HOXC6 promoter is responsive to BPA and E2 treatment via two putative estrogen response element; consequently, BPA could induce the expression of the HOXC6 even in the absence of E2 both in vitro and in vivo [45].
Another HOX gene, HOXB9, was induced in breast cancer cells upon exposure to BPA [23]. The HOXB9 gene is responsible for cell cycle progression, embryonic segmentation, limb pattering, and cell proliferation as well as mammary gland and skeletal development. Studies have demonstrated that HOXB9 expression is altered in various cancer types including those of the breast [23]. The overexpression of HOXB9 in breast tumor cells stimulates tumor neovascularization, tumor invasion, and disease progression [24]. In the presence of BPA, the level of HOXB9 mRNA expression was upregulated in breast tumor cells both in vitro and in vivo through recruitment of transcription factors and chromatin-modification factors. Similar to HOXC6, the HOXB9 gene promoter also contains putative estrogen response element (ERE4) that mediates its response to both E2 and BPA [23].
Some reports have pointed to an association between HOX genes and response to chemotherapy of tumors. For example, overexpression of HOXB9 was associated with poor disease-free survival and poor overall survival in breast cancer patients [24]. HOXC6 overexpression in pretreatment biopsies of Esophageal Squamous Cell Carcinoma (ESCC) specimens was associated with low tumor regression. Downregulation of HOXC6 decreased the sensitivity of ESCC cell lines to cisplatin and paclitaxel [46].
3.2.4. BPA and Bone Morphogenetic Proteins
Soto and coworkers (2013) showed that exposure to BPA during gestation can increase the sensitivity to mammotropic hormones and thus increases the incidence of breast cancer later on during life [35]. Since the differentiation and fate of mammary epithelial stem cells is under the control of BMP2 and BMP4, it is likely that BPA exerts these effects by changing the mammary differentiation program mediated by BMPs. This hypothesis is supported by Clement et al. (2017) who showed that BPA can alter the differentiation of stem cells by changing their sensitivity to BMPs signaling, deregulating the production of BMPs by mammary fibroblast and changing the localization and expression of BMP type-1 receptors. BPA altered BMP signaling via the phosphorylation of “small mothers against decapentaplegic” 1/5/8 (SMAD1/5/8) [47]. The spectrum of this action could be extended to other cancer types-it is well established that BMPs are involved in the development of many cancers [48].
Activation of the BMP-BMPR-SMAD1/5 signaling pathway can lead to anti-cancer drug resistance in many cancer cells including lung squamous cell carcinoma [48, 49]. For example, resistance to EGFR tyrosine kinase inhibitors (EGFR-TKI) in lung squamous cell carcinoma was due to the activation of the BMP-BMPR-p70S6K pathway [49].
3.2.5. Other Signaling Pathways
Previous studies have revealed that BPA can trigger the proliferation and migration of breast cancer cells SKBR3 via stimulation of GPER to induce functional crosstalk between cancer-associated fibroblasts (CAFs) and cancer cells [11]. BPA can activate a GPER/caveolin 1 (Cav-1)/heat shock protein 90 (HSP90) signaling cascade under hypoxic conditions to upregulate the expression of hypoxia-inducible factor-1 alpha (HIF-1α) and vascular endothelial growth factor (VEGF) resulting in the proliferation of breast cancer cells [50].
BPA can also up-regulate the oncogenic protein c-Myc that predisposes breast cancer cells to accumulate DNA double-strand breaks and produce reactive oxygen species (ROS), which leads in turn to increased proliferation of these cells [51]. ROS can also promote and modulate other metabolic events leading to chemoresistance [43].
We emphasize that the list of cellular pathways and molecules affected by BPA are continuously expanding. Hence, the pathways mentioned here are by no mean comprehensive because we focused on the putative pathways associated with chemoresistance. The reader is referred to additional publications [52-56] and the emerging literature on this topic.
4. Ovarian Cancer and BPA
Ovarian cancer is relatively uncommon but very deadly: The 5-year survival rate is only 10-20%. In vitro studies have indicated that BPA stimulates VEGF production and may interfere with granulosa cell steroidogenesis and hence interfere with reproductive activity [57]. The association between BPA and ovarian cancer as well as the mechanisms of ovarian cancer development by BPA remain unclear. Biomonitoring studies have demonstrated that BPA is detectable at nanomolar concentrations in human blood, urine, and milk [4]. Such low doses of BPA can increase the proliferation and migration of several ovarian cancer cell lines. This oncogenic action was associated with the deregulation of several cellular pathways and molecular targets. BPA induced phosphorylation of “signal transducer and activator of transcription 3” (Stat3) and ERK1/2 [58], and upregulation of vascular endothelial growth factor-A (VEGF-A) and its VEGF-R2 receptor in both cancerous and non-cancerous cells. BPA triggered the epithelial-to-mesenchymal transition (EMT) through ER-mediated upregulation of N-cadherin, MMP-9, and MMP-2 [59, 60]. Conversely, BPA could suppress the expression of proapoptotic factors including caspase-3 and caspase-7 through ERK1/2 signaling pathway [61]. Sub-toxic doses of BPA could also enhance the glycolysis in an ERα-dependent manner resulting in higher levels of intracellular ATP production in ovarian cancer cells [62]. Most of these alterations change the cellular response to therapy and predispose the cells to chemoresistance.
The homeobox transcript antisense intergenic RNA (HOTAIR) is a long, noncoding RNA with a role in carcinogenesis. HOTAIR is associated with larger tumor size, metastasis, drug resistance, and poor prognosis in many tumors. Overexpression of HOTAIR resulted in cisplatin resistance by regulating the expression of p21 and downregulation of microRNA-130a by activating NF-κB, PIK3R3, and MAPK1 [63, 64]. BPA upregulated the expression of HOTAIR in breast cancer cells through epigenetic modification of its promoter [65].
BPA altered the production of different types of collagen around different cells [66]. The overexpression of collagen is associated with drug resistance in both breast and ovarian cancer due to the interaction of collagen with these cancer cells. The ovarian cancer cell line A2780 developed resistance to cisplatin when it was cultured in the presence of collagen. This drug resistance can be due to altered apoptosis sensitivity of cells and/or limited drug diffusion into cancer tissues [67].
5. Prostate cancer and BPA
Prostate cancer incidence has been steadily increasing. It is currently second most common cancer among men [30], which might reflect increased environmental exposure to potential prostatic carcinogens. Many studies have found a link between BPA and increased risk of hormone-related cancers. The most likely explanation for this association is that BPA can activate steroid receptors such as ERα, ERβ, ERRs, and AR. Data from animal models have demonstrated that overexpression of AR is sufficient for the growth of prostate cancer [68]. The most common treatment modality for prostatic cancer is Androgen Deprivation Therapy (ADT), which blocks AR activity. Unfortunately, many patients develop ADT-resistance and cannot ablate AR function in these tumors. ADT-resistance is usually a consequence of acquiring mutant ARs (the most common AR mutant form is AR-T877A), which arise due to selective pressure under ADT.
BPA activates androgen receptor mutant version AR-T877A leading to the proliferation of prostatic adenocarcinoma cells LNCaP [29]. BPA can act as an alternative ligand for mutant AR (AR-T877A) and facilitate its dissociation from heat shock protein resulting in dimerization of the mutant AR receptor and its localization to the nucleus. These dimers (BPA/AR-T877A), with assistance of co-activator molecules, bind to gene-regulatory regions (androgen response elements) such as those of prostate-specific antigens (PSA) to trigger cell proliferation. Additional work has suggested that BPA/AR-T877A dimers downregulate ERβ, which is known to inhibit prostatic cancer cells and consequently facilitate enhanced proliferation and therapy bypass [28]. Interestingly, BPA can also stimulate and increase the expression of normal AR mRNA. The administration of low doses of BPA to pregnant mice can permanently increase the prostate size in male offspring [69].
Conversely, BPA at high doses can exert anti-proliferative activity accompanied by cell cycle arrest in prostate cancer by triggering cross talk with AR and ERβ to rapidly activate EGFR. This can phosphorylates ERK and upregulate p53 and induce its phosphorylation at Ser15. This increases the transcription of p27 and p21 and induce cell cycle arrest [16].
It has been reported that approximately 30,000 patients die yearly due to the resistance of advanced prostate cancer to available drugs. These intrinsic drug-resistance mechanisms include increased drug efflux from cancer cells by overexpressed membrane bound efflux protein Adenosine Triphosphate (ATP)-Binding Cassette (ABC), overexpression of transporter P-glycoprotein (P-gp, or ATP-binding cassette sub-family B member 1 (ABCB1)), and overexpression of Multidrug Resistance-associated Protein 1 (MRP1) encoded by ATP-binding cassette subfamily C member 1 (ABCC1) gene. This is in addition to the acquired continued signaling from mutant AR and overexpression of normal AR that activate other pro-survival signaling pathways [68]. Other models have shown that increasing the concentration of conjugated BPA significantly correlated with high expression of ABCC1 mRNA-encoding MRP1 [70, 71].
6. BPA and Colorectal Cancer
Colorectal cancer is the third most common cancer worldwide and is a major cause of cancer deaths particularly in developing countries [72]. The development of colon cancer is associated with multiple molecular genetic and epigenetic alterations with a high degree of genetic instability [73]. In particular, the patterns of epigenetic alterations in colorectal cancers suggests a link to environmental exposure in some groups [72, 74]. Since orally ingested BPA will be in direct contact with the gastrointestinal mucosal cells, it is likely to exert its toxic effects on the gut and possibly enhance transformation of the gastrointestinal cells. However, relatively few reports have addressed the effect of BPA on the gastrointestinal tract versus breast and classical endocrine-responsive organs.
Chen and coworker (2015) presented a comprehensive proteomic study of BPA effects in colon cancer cell lines. Upon exposure to BPA, the expressions of more than 50 proteins were significantly modulated in colorectal cancer cell lines. These proteins were related to cell structure and motility, cell proliferation, apoptosis, energy metabolism, and oxidative stress. The authors focused on one of the most dominant features in colon cancer and confirmed that BPA can promote the motility and invasion of colorectal cancer cells through induction of EMT. The authors also showed that this effect may be attributed to phosphorylation of AKT and GSK3β that mediates the upregulation of mesenchymal markers Snail, N-cadherin, and vimentin as well as downregulation of E-cadherin [27]. Other studies showed that EMT markers such as AXL and Moesin are associated with resistance to standard therapy in colon and breast cancer cells [32, 75].
The most significant hits detected by proteomic analysis in colon cancer cells after BPA exposure were upregulation of fascin and eukaryotic initiation factor 4A-1 (TIF4A) and downregulation of cytokeratin 8 (KRT8) [27]. Interestingly, the expression of Fascin in colorectal cancer cells was shown to be associated with poor prognosis [25, 26]. Moreover, the overexpression of fascin in xenografted breast cancer cells induced resistance to doxorubicin via activation of PI3K/Akt pathway, which led cancer cells to evade apoptosis through suppression of pro-apoptotic caspase 9 and caspase 3 [76].
As discussed above, the induction of HOX genes such as HOXB9 by BPA can play a role in the development and formation of some types of cancers such as breast cancer. HOXB9 can enhance chemoresistance to anti-VEGF agent bevacizumab in colorectal cancer in mouse xenograft models by regulating and controlling the expression of many angiogenic factors such as angiopoietin-like 2 (Angptl2), CXCL1,TGF-β1, and IL8 [77, 78].
CONCLUSION
BPA is a common EDC and is detectable in a wide range of body fluids from vulnerable groups such as the urine of children and healthy infants, serum of pregnant women, follicular and amniotic fluid, fetal serum, cord blood, placenta tissue, breast milk, and saliva of patients exposed to some dental treatments. This review addressed an important aspect of BPA’s adverse effects in humans: its association with cancer and particularly resistance to chemotherapy and poor prognosis in cancer patients. Due to the ubiquitous nature of BPA and the potential for continuous exposure particularly through hospital equipment and instruments, the impact of this effect might be dramatic because it worsens the prognosis of already suffering cancer patients. More generally, the association between BPA and cancer may be one of the environmental risks that predispose people to increased incidence and poor outcomes of common cancers. This argument should not be surprising considering that the most common cancers in 2018 in men and women (those of the prostate, breast, and colon) are known to be associated with increased exposure to environmental toxins. We explored the molecular mechanisms of this action and showed that low concentrations of BPA below the “safe level” approved by the regulatory authorities can disrupt many cellular pathways and trigger molecular mechanisms that result in increased cellular proliferation and chemoresistance (Abdel-Rahman et al., unpublished data).
In spite of the mounting evidence supporting such deleterious effects of low dose BPA on human health, there is still a big debate and denial from the regulatory authorities including the Food and Drug Administration (FDA or USFDA) as highlighted by results of the major collaborative project, that was conducted by 14 investigators funded by the National Institute of Environmental Health Sciences. The report is called the Consortium Linking Academic and Regulatory Insights on Toxicity of BPA (CLARITY-BPA) and is available at https://ntp.niehs.nih.gov/results/areas/bpa/index.html. The report triggered a debate in the scientific community and public media reports. A recent review summarized the flaws in the design and execution of CLARITY-BPA that biased the experiment toward not finding significant results. Examples of these flaws are: 1. The NCTR CD-SD rat is an estrogen-insensitive strain, 2. The administration procedure of BPA via a daily gavage throughout life was stressful, 3. Non-gavaged negative controls were not included. Furthermore, the FDA ignored non-monotonic low dose-response relationships [79, 80]. The data discussed here highlight a major aspect of BPA toxicity: chemoresistance in cancer. That aspect of BPA toxicity was not fully addressed in the CLARITY-BPA project.
ACKNOWLEDGEMENTS
Declared none.
CONSENT FOR PUBLICATION
Not applicable.
FUNDING
This work was supported by: University of Sharjah, Research Board, grant # 15010501010-P to Wael M. Abdel-Rahman.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
REFERENCES
- 1.Gao H., Yang B.J., Li N., Feng L.M., Shi X.Y., Zhao W.H., Liu S.J. Bisphenol A and hormone-associated cancers: Current progress and perspectives. Medicine (Baltimore) 2015;94:e211. doi: 10.1097/MD.0000000000000211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fenichel P., Chevalier N., Brucker-Davis F. Bisphenol A: an endocrine and metabolic disruptor. Ann. Endocrinol. (Paris) 2013;74:211–220. doi: 10.1016/j.ando.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 3.Staples C.A., Dorn P.B., Klecka G.M., O’Block S.T., Harris L.R. A review of the environmental fate, effects, and exposures of bisphenol A. Chemosphere. 1998;36:2149–2173. doi: 10.1016/s0045-6535(97)10133-3. [DOI] [PubMed] [Google Scholar]
- 4.Vandenberg L.N., Chahoud I., Heindel J.J., Padmanabhan V., Paumgartten F.J., Schoenfelder G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010;118:1055–1070. doi: 10.1289/ehp.0901716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lorber M., Schecter A., Paepke O., Shropshire W., Christensen K., Birnbaum L. Exposure assessment of adult intake of bisphenol A (BPA) with emphasis on canned food dietary exposures. Environ. Int. 2015;77:55–62. doi: 10.1016/j.envint.2015.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hines E.P., Mendola P., von Ehrenstein O.S., Ye X., Calafat A.M., Fenton S.E. Concentrations of environmental phenols and parabens in milk, urine and serum of lactating North Carolina women. Reprod. Toxicol. 2015;54:120–128. doi: 10.1016/j.reprotox.2014.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Metz C., Bisphenol A. Understanding the Controversy. Workplace Health Saf. 2016;64:28–36. doi: 10.1177/2165079915623790. [DOI] [PubMed] [Google Scholar]
- 8.Abdel-Rahman W.M., Moustafa Y.M., Ahmed B.O., Mostafa R.M. Endocrine disruptors and breast cancer risk – time to consider the environment. Asian Pac. J. Cancer Prev. 2012;13:5937–5946. [Google Scholar]
- 9.Zamkowska D., Karwacka A., Jurewicz J., Radwan M. Environmental exposure to non-persistent endocrine disrupting chemicals and semen quality: An overview of the current epidemiological evidence. Int. J. Occup. Med. Environ. Health. 2018;31:377–414. doi: 10.13075/ijomeh.1896.01195. [DOI] [PubMed] [Google Scholar]
- 10.Rahmani S., Pour Khalili N., Khan F., Hassani S., Ghafour-Boroujerdi E., Abdollahi M., Bisphenol A. What lies beneath its induced diabetes and the epigenetic modulation? Life Sci. 2016;214:136–144. doi: 10.1016/j.lfs.2018.10.044. [DOI] [PubMed] [Google Scholar]
- 11.Pupo M., Pisano A., Lappano R., Santolla M.F., De Francesco E.M., Abonante S., Rosano C., Maggiolini M. Bisphenol A induces gene expression changes and proliferative effects through GPER in breast cancer cells and cancer-associated fibroblasts. Environ. Health Perspect. 2012;120:1177–1182. doi: 10.1289/ehp.1104526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Drozdz K., Wysokinski D., Krupa R., Wozniak K. Bisphenol A-glycidyl methacrylate induces a broad spectrum of DNA damage in human lymphocytes. Arch. Toxicol. 2011;85:1453–1461. doi: 10.1007/s00204-010-0593-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Audebert M., Dolo L., Perdu E., Cravedi J.P., Zalko D. Use of the gammaH2AX assay for assessing the genotoxicity of bisphenol A and bisphenol F in human cell lines. Arch. Toxicol. 2011;85:1463–1473. doi: 10.1007/s00204-011-0721-2. [DOI] [PubMed] [Google Scholar]
- 14.Wang Z., Liu H., Liu S. Low-Dose Bisphenol A Exposure: A Seemingly Instigating Carcinogenic Effect on Breast Cancer. Adv. Sci. (Weinh.) 2016;4:1600248. doi: 10.1002/advs.201600248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jalal N., Surendranath A.R., Pathak J.L., Yu S., Chung C.Y. Bisphenol A (BPA) the mighty and the mutagenic. Toxicol. Rep. •••;1016(5):76–84. doi: 10.1016/j.toxrep.2017.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bilancio A., Bontempo P., Di Donato M., Conte M., Giovannelli P., Altucci L., Migliaccio A., Castoria G. Bisphenol A induces cell cycle arrest in primary and prostate cancer cells through EGFR/ERK/p53 signaling pathway activation. Oncotarget. 2017;8:115620–115631. doi: 10.18632/oncotarget.23360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dairkee S.H., Luciani-Torres M.G., Moore D.H., Goodson W.H. 3rd Bisphenol-A-induced inactivation of the p53 axis underlying deregulation of proliferation kinetics, and cell death in non-malignant human breast epithelial cells. Carcinogenesis. 2013;34:703–712. doi: 10.1093/carcin/bgs379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Heckler M.M., Thakor H., Schafer C.C., Riggins R.B. ERK/MAPK regulates ERRgamma expression, transcriptional activity and receptor-mediated tamoxifen resistance in ER+ breast cancer. FEBS J. 2014;281:2431–2442. doi: 10.1111/febs.12797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Song H., Zhang T., Yang P., Li M., Yang Y., Wang Y., Du J., Pan K., Zhang K. Low doses of bisphenol A stimulate the proliferation of breast cancer cells via ERK1/2/ERRgamma signals. Toxicol. In Vitro. 2015;30:521–528. doi: 10.1016/j.tiv.2015.09.009. [DOI] [PubMed] [Google Scholar]
- 20.Sauer S.J., Tarpley M., Shah I., Save A.V., Lyerly H.K., Patierno S.R., Williams K.P., Devi G.R. Bisphenol A activates EGFR and ERK promoting proliferation, tumor spheroid formation and resistance to EGFR pathway inhibition in estrogen receptor-negative inflammatory breast cancer cells. Carcinogenesis. 2017;38:252–260. doi: 10.1093/carcin/bgx003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lapensee E.W., Tuttle T.R., Fox S.R., Ben-Jonathan N. Bisphenol A at low nanomolar doses confers chemoresistance in estrogen receptor-alpha-positive and -negative breast cancer cells. Environ. Health Perspect. 2009;117:175–180. doi: 10.1289/ehp.11788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.LaPensee E.W., LaPensee C.R., Fox S., Schwemberger S., Afton S., Ben-Jonathan N. Bisphenol A and estradiol are equipotent in antagonizing cisplatin-induced cytotoxicity in breast cancer cells. Cancer Lett. 2010;290:167–173. doi: 10.1016/j.canlet.2009.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Deb P., Bhan A., Hussain I., Ansari K.I., Bobzean S.A., Pandita T.K., Perrotti L.I., Mandal S.S. Endocrine disrupting chemical, bisphenol-A, induces breast cancer associated gene HOXB9 expression in vitro and in vivo. Gene. 2016;590:234–243. doi: 10.1016/j.gene.2016.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seki H., Hayashida T., Jinno H., Hirose S., Sakata M., Takahashi M., Maheswaran S., Mukai M., Kitagawa Y. HOXB9 expression promoting tumor cell proliferation and angiogenesis is associated with clinical outcomes in breast cancer patients. Ann. Surg. Oncol. 2012;19:1831–1840. doi: 10.1245/s10434-012-2295-5. [DOI] [PubMed] [Google Scholar]
- 25.Qualtrough D., Singh K., Banu N., Paraskeva C., Pignatelli M. The actin-bundling protein fascin is overexpressed in colorectal adenomas and promotes motility in adenoma cells in vitro. Br. J. Cancer. 2009;101:1124–1129. doi: 10.1038/sj.bjc.6605286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chan C., Jankova L., Fung C.L., Clarke C., Robertson G., Chapuis P.H., Bokey L., Lin B.P., Dent O.F., Clarke S. Fascin expression predicts survival after potentially curative resection of node-positive colon cancer. Am. J. Surg. Pathol. 2010;34:656–666. doi: 10.1097/PAS.0b013e3181db36c0. [DOI] [PubMed] [Google Scholar]
- 27.Chen Z.J., Yang X.L., Liu H., Wei W., Zhang K.S., Huang H.B., Giesy J.P., Liu H.L., Du J., Wang H.S. Bisphenol A modulates colorectal cancer protein profile and promotes the metastasis via induction of epithelial to mesenchymal transitions. Arch. Toxicol. 2015;89:1371–1381. doi: 10.1007/s00204-014-1301-z. [DOI] [PubMed] [Google Scholar]
- 28.Hess-Wilson J.K. Bisphenol A may reduce the efficacy of androgen deprivation therapy in prostate cancer. Cancer Causes Control. 2009;20:1029–1037. doi: 10.1007/s10552-009-9337-8. [DOI] [PubMed] [Google Scholar]
- 29.Wetherill Y.B., Fisher N.L., Staubach A., Danielsen M., de Vere White R.W., Knudsen K.E. Xenoestrogen action in prostate cancer: pleiotropic effects dependent on androgen receptor status. Cancer Res. 2005;65:54–65. [PubMed] [Google Scholar]
- 30.Bray F., Ferlay J., Soerjomataram I., Siegel R.L., Torre L.A., Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 31.Azubuike S.O., Muirhead C., Hayes L., McNally R. Rising global burden of breast cancer: the case of sub-Saharan Africa (with emphasis on Nigeria) and implications for regional development: a review. World J. Surg. Oncol. 2018;16:63. doi: 10.1186/s12957-018-1345-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Alam F., Mezhal F., El Hasasna H., Nair V.A., Aravind S.R., Saber Ayad M., El-Serafi A., Abdel-Rahman W.M. The role of p53-microRNA 200-Moesin axis in invasion and drug resistance of breast cancer cells. Tumour Biol. 2017;39:1010428317714634. doi: 10.1177/1010428317714634. [DOI] [PubMed] [Google Scholar]
- 33.Lotsari J.E., Gylling A., Abdel-Rahman W.M., Nieminen T.T., Aittomaki K., Friman M., Pitkanen R., Aarnio M., Jarvinen H.J., Mecklin J.P., Kuopio T., Peltomaki P. Breast carcinoma and Lynch syndrome: molecular analysis of tumors arising in mutation carriers, non-carriers, and sporadic cases. Breast Cancer Res. 2012;14:R90. doi: 10.1186/bcr3205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Feng Y., Spezia M., Huang S., Yuan C., Zeng Z., Zhang L., Ji X., Liu W., Huang B., Luo W., Liu B., Lei Y., Du S., Vuppalapati A., Luu H.H., Haydon R.C., He T.C., Ren G. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018;5:77–106. doi: 10.1016/j.gendis.2018.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Soto A.M., Brisken C., Schaeberle C., Sonnenschein C. Does cancer start in the womb? altered mammary gland development and predisposition to breast cancer due to in utero exposure to endocrine disruptors. J. Mammary Gland Biol. Neoplasia. 2013;18:199–208. doi: 10.1007/s10911-013-9293-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Siddique S., Kubwabo C., Harris S.A. A review of the role of emerging environmental contaminants in the development of breast cancer in women. Emerg. Contam. 2016;2:204–219. [Google Scholar]
- 37.Rochefort H. Endocrine disruptors (EDs) and hormone-dependent cancers: Correlation or causal relationship? C. R. Biol. 2017;340:439–445. doi: 10.1016/j.crvi.2017.07.007. [DOI] [PubMed] [Google Scholar]
- 38.Delgado-López F., Pilar Zamora-León S. Breast cancer and environmental contamination: A real connection. Cogent Med. 2018;5:1–15. [Google Scholar]
- 39.Ju T., Jin H., Ying R., Xie Q., Zhou C., Gao D. Overexpression of NAC1 confers drug resistance via HOXA9 in colorectal carcinoma cells. Mol. Med. Rep. 2017;16:3194–3200. doi: 10.3892/mmr.2017.6986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wu Y., Zhang Z., Cenciarini M.E., Proietti C.J., Amasino M., Hong T., Yang M., Liao Y., Chiang H.C., Kaklamani V.G., Jeselsohn R., Vadlamudi R.K., Huang T.H., Li R., De Angelis C., Fu X., Elizalde P.V., Schiff R., Brown M., Xu K. Tamoxifen Resistance in Breast Cancer Is Regulated by the EZH2-ERalpha-GREB1 Transcriptional Axis. Cancer Res. 2018;78:671–684. doi: 10.1158/0008-5472.CAN-17-1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ranhotra H.S. The estrogen-related receptors in metabolism and cancer: newer insights. J. Recept. Signal Transduct. Res. 2018;38:95–100. doi: 10.1080/10799893.2018.1456552. [DOI] [PubMed] [Google Scholar]
- 42.Dong S., Terasaka S., Kiyama R. Bisphenol A induces a rapid activation of Erk1/2 through GPR30 in human breast cancer cells. Environ. Pollut. 2011;159:212–218. doi: 10.1016/j.envpol.2010.09.004. [DOI] [PubMed] [Google Scholar]
- 43.Okon I.S., Zou M.H. Mitochondrial ROS and cancer drug resistance: Implications for therapy. Pharmacol. Res. 2015;100:170–174. doi: 10.1016/j.phrs.2015.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ansari K.I., Hussain I., Shrestha B., Kasiri S., Mandal S.S. HOXC6 Is transcriptionally regulated via coordination of MLL histone methylase and estrogen receptor in an estrogen environment. J. Mol. Biol. 2011;411:334–349. doi: 10.1016/j.jmb.2011.05.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hussain I., Bhan A., Ansari K.I., Deb P., Bobzean S.A., Perrotti L.I., Mandal S.S. Bisphenol-A induces expression of HOXC6, an estrogen-regulated homeobox-containing gene associated with breast cancer. Biochim. Biophys. Acta. 2015;1849:697–708. doi: 10.1016/j.bbagrm.2015.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shen L.Y., Fan M.Y., Dong B., Yan W.P., Chen K.N. Increased HOXC6 expression predicts chemotherapy sensitivity in patients with esophageal squamous cell carcinoma. Oncol. Lett. 2017;14:4835–4840. doi: 10.3892/ol.2017.6772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Clement F., Xu X., Donini C.F., Clement A., Omarjee S., Delay E., Treilleux I., Fervers B., Le Romancer M., Cohen P.A., Maguer-Satta V. Long-term exposure to bisphenol A or benzo(a)pyrene alters the fate of human mammary epithelial stem cells in response to BMP2 and BMP4, by pre-activating BMP signaling. Cell Death Differ. 2017;24:155–166. doi: 10.1038/cdd.2016.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bach D.H., Park H.J., Lee S.K. The dual role of bone morphogenetic proteins in cancer. Mol. Ther. Oncolytics. 2018;8:1–13. doi: 10.1016/j.omto.2017.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang Z., Shen Z., Li Z., Duan J., Fu S., Liu Z., Bai H., Zhang Z., Zhao J., Wang X., Wang J. Activation of the BMP-BMPR pathway conferred resistance to EGFR-TKIs in lung squamous cell carcinoma patients with EGFR mutations. Proc. Natl. Acad. Sci. USA. 2015;112:9990–9995. doi: 10.1073/pnas.1510837112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.De Francesco E.M., Maggiolini M., Musti A.M. Crosstalk between Notch, HIF-1alpha and GPER in Breast Cancer EMT. Int. J. Mol. Sci. 2018;•••:19. doi: 10.3390/ijms19072011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pfeifer D., Chung Y.M., Hu M.C. Effects of Low-Dose Bisphenol A on DNA Damage and Proliferation of Breast Cells: The Role of c-Myc. Environ. Health Perspect. 2015;123:1271–1279. doi: 10.1289/ehp.1409199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Williams K.E., Lemieux G.A., Hassis M.E., Olshen A.B., Fisher S.J., Werb Z. Quantitative proteomic analyses of mammary organoids reveals distinct signatures after exposure to environmental chemicals. Proc. Natl. Acad. Sci. USA. 2016;113:E1343–E1351. doi: 10.1073/pnas.1600645113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Liu J., Jin X., Zhao N., Ye X., Ying C. Bisphenol A promotes X-linked inhibitor of apoptosis protein-dependent angiogenesis via G protein-coupled estrogen receptor pathway. J. Appl. Toxicol. 2015;35:1309–1317. doi: 10.1002/jat.3112. [DOI] [PubMed] [Google Scholar]
- 54.Mlynarcikova A., Macho L., Fickova M. Bisphenol A alone or in combination with estradiol modulates cell cycle- and apoptosis-related proteins and genes in MCF7 cells. Endocr. Regul. 2013;47:189–199. doi: 10.4149/endo_2013_04_189. [DOI] [PubMed] [Google Scholar]
- 55.Goodson W.H., III, Luciani M.G., Sayeed S.A., Jaffee I.M., Moore D.H., II, Dairkee S.H. Activation of the mTOR pathway by low levels of xenoestrogens in breast epithelial cells from high-risk women. Carcinogenesis. 2011;32:1724–1733. doi: 10.1093/carcin/bgr196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dairkee S.H., Seok J., Champion S., Sayeed A., Mindrinos M., Xiao W., Davis R.W., Goodson W.H. Bisphenol A induces a profile of tumor aggressiveness in high-risk cells from breast cancer patients. Cancer Res. 2008;68:2076–2080. doi: 10.1158/0008-5472.CAN-07-6526. [DOI] [PubMed] [Google Scholar]
- 57.Grasselli F., Baratta L., Baioni L., Bussolati S., Ramoni R., Grolli S., Basini G. Bisphenol A disrupts granulosa cell function. Domest. Anim. Endocrinol. 2010;39:34–39. doi: 10.1016/j.domaniend.2010.01.004. [DOI] [PubMed] [Google Scholar]
- 58.Ptak A., Gregoraszczuk E.L. Bisphenol A induces leptin receptor expression, creating more binding sites for leptin, and activates the JAK/Stat, MAPK/ERK and PI3K/Akt signalling pathways in human ovarian cancer cell. Toxicol. Lett. 2012;210:332–337. doi: 10.1016/j.toxlet.2012.02.003. [DOI] [PubMed] [Google Scholar]
- 59.Ptak A., Gregoraszczuk E.L. Effects of bisphenol A and 17beta-estradiol on vascular endothelial growth factor A and its receptor expression in the non-cancer and cancer ovarian cell lines. Cell Biol. Toxicol. 2015;31:187–197. doi: 10.1007/s10565-015-9303-z. [DOI] [PubMed] [Google Scholar]
- 60.Kim Y.S., Hwang K.A., Hyun S.H., Nam K.H., Lee C.K., Choi K.C. Bisphenol A and nonylphenol have the potential to stimulate the migration of ovarian cancer cells by inducing epithelial-mesenchymal transition via an estrogen receptor dependent pathway. Chem. Res. Toxicol. 2015;28:662–671. doi: 10.1021/tx500443p. [DOI] [PubMed] [Google Scholar]
- 61.Ptak A., Wrobel A., Gregoraszczuk E.L. Effect of bisphenol-A on the expression of selected genes involved in cell cycle and apoptosis in the OVCAR-3 cell line. Toxicol. Lett. 2011;202:30–35. doi: 10.1016/j.toxlet.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 62.Shi X.Y., Wang Z., Liu L., Feng L.M., Li N., Liu S., Gao H. Low concentrations of bisphenol A promote human ovarian cancer cell proliferation and glycolysis-based metabolism through the estrogen receptor-alpha pathway. Chemosphere. 2017;185:361–367. doi: 10.1016/j.chemosphere.2017.07.027. [DOI] [PubMed] [Google Scholar]
- 63.Li J., Wang J., Zhong Y., Guo R., Chu D., Qiu H., Yuan Z. HOTAIR: A key regulator in gynecologic cancers. Cancer Cell Int. 2017;17:65. doi: 10.1186/s12935-017-0434-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Zhou X., Chen J., Tang W. The molecular mechanism of HOTAIR in tumorigenesis, metastasis, and drug resistance. Acta Biochim. Biophys. Sin. (Shanghai) 2014;46:1011–1015. doi: 10.1093/abbs/gmu104. [DOI] [PubMed] [Google Scholar]
- 65.Bhan A., Hussain I., Ansari K.I., Bobzean S.A., Perrotti L.I., Mandal S.S. Bisphenol-A and diethylstilbestrol exposure induces the expression of breast cancer associated long noncoding RNA HOTAIR in vitro and in vivo. J. Steroid Biochem. Mol. Biol. 2014;141:160–170. doi: 10.1016/j.jsbmb.2014.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chin K.Y., Pang K.L., Mark-Lee W.F. A Review on the Effects of Bisphenol A and Its Derivatives on Skeletal Health. Int. J. Med. Sci. 2018;15:1043–1050. doi: 10.7150/ijms.25634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Januchowski R., Swierczewska M., Sterzynska K., Wojtowicz K., Nowicki M., Zabel M. Increased Expression of Several Collagen Genes is Associated with Drug Resistance in Ovarian Cancer Cell Lines. J. Cancer. 2016;7:1295–1310. doi: 10.7150/jca.15371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Seruga B., Ocana A., Tannock I.F. Drug resistance in metastatic castration-resistant prostate cancer. Nat. Rev. Clin. Oncol. 2011;8:12–23. doi: 10.1038/nrclinonc.2010.136. [DOI] [PubMed] [Google Scholar]
- 69.Richter C.A., Taylor J.A., Ruhlen R.L., Welshons W.V., Vom Saal F.S. Estradiol and Bisphenol A stimulate androgen receptor and estrogen receptor gene expression in fetal mouse prostate mesenchyme cells. Environ. Health Perspect. 2007;115:902–908. doi: 10.1289/ehp.9804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Moscovitz J.E., Nahar M.S., Shalat S.L., Slitt A.L., Dolinoy D.C., Aleksunes L.M. Correlation between Conjugated Bisphenol A Concentrations and Efflux Transporter Expression in Human Fetal Livers. Drug Metab. Dispos. 2016;44:1061–1065. doi: 10.1124/dmd.115.068668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Nickel S., Bernd A., Miller D.S., Fricker G., Mahringer A. Bisphenol - A modulates function of ABC transporters in killifish. MDIBL. Bull. 2013;52:30. [Google Scholar]
- 72.Abdel-Rahman W.M., Faris M.E., Peltomaki P. Molecular Determinants of Colon Cancer Susceptibility in the East and West. Curr. Mol. Med. 2017;17:34–45. doi: 10.2174/1566524017666170220094705. [DOI] [PubMed] [Google Scholar]
- 73.Abdel-Rahman W.M. Genomic instability and carcinogenesis: An update. Curr. Genomics. 2008;9:535–541. doi: 10.2174/138920208786847926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nieminen T.T., Shoman S., Eissa S., Peltomaki P., Abdel-Rahman W.M. Distinct genetic and epigenetic signatures of colorectal cancers according to ethnic origin. Cancer Epidemiol. Biomarkers Prev. 2012;21:202–211. doi: 10.1158/1055-9965.EPI-11-0662. [DOI] [PubMed] [Google Scholar]
- 75.Abdel-Rahman W.M., Al-Khayyal N.A., Nair V.A., Aravind S.R., Saber-Ayad M. Role of AXL in invasion and drug resistance of colon and breast cancer cells and its association with p53 alterations. World J. Gastroenterol. 2017;23:3440–3448. doi: 10.3748/wjg.v23.i19.3440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ghebeh H., Al-Khaldi S., Olabi S., Al-Dhfyan A., Al-Mohanna F., Barnawi R., Tulbah A., Al-Tweigeri T., Ajarim D., Al-Alwan M. Fascin is involved in the chemotherapeutic resistance of breast cancer cells predominantly via the PI3K/Akt pathway. Br. J. Cancer. 2014;111:1552–1561. doi: 10.1038/bjc.2014.453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Itatani Y., Kawada K., Yamamoto T., Sakai Y. Resistance to Anti-Angiogenic Therapy in Cancer-Alterations to Anti-VEGF Pathway. Int. J. Mol. Sci. 2018;•••:19. doi: 10.3390/ijms19041232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Carbone C., Piro G., Simionato F., Ligorio F., Cremolini C., Loupakis F., Ali G., Rossini D., Merz V., Santoro R., Zecchetto C., Zanotto M., Di Nicolantonio F., Bardelli A., Fontanini G., Tortora G., Melisi D. Homeobox B9 Mediates Resistance to Anti-VEGF Therapy in Colorectal Cancer Patients. Clin. Cancer Res. 2017;23:4312–4322. doi: 10.1158/1078-0432.CCR-16-3153. [DOI] [PubMed] [Google Scholar]
- 79.Vom Saal F.S. Flaws in design, execution and interpretation limit CLARITY-BPA’s value for risk assessments of bisphenol A. Basic Clin. Pharmacol. Toxicol. 2018;••• doi: 10.1111/bcpt.13195. [DOI] [PubMed] [Google Scholar]
- 80.Hill C.E., Myers J.P., Vandenberg L.N. Nonmonotonic Dose-Response Curves Occur in Dose Ranges That Are Relevant to Regulatory Decision-Making. Dose Response. 2018;16(3):1559325818798282. doi: 10.1177/1559325818798282. [DOI] [PMC free article] [PubMed] [Google Scholar]