

Abstract
Alzheimer's disease (AD) subtypes have been described according to genetics, neuropsychology, neuropathology, and neuroimaging. Thirty‐one patients with clinically probable AD were selected based on perisylvian metabolic decrease on FDG‐PET. They were compared to 25 patients with a typical pattern of decreased posterior metabolism. Tree‐based machine learning was used on those 56 images to create a classifier that was subsequently applied to 207 Alzheimer's Disease Neuroimaging Initiative (ADNI) patients with AD. Machine learning was also used to discriminate between the two ADNI groups based on neuropsychological scores. Compared to AD patients with a typical precuneus metabolic decrease, the new subtype showed stronger hypometabolism in the temporoparietal junction. The classifier was able to distinguish the two groups in the ADNI population. Both groups could only be distinguished cognitively by Trail Making Test‐A scores. This study further confirms that there is more than a typical metabolic pattern in probable AD with amnestic presentation.
Keywords: Alzheimer, FDG‐PET, machine learning, neuroimaging, subtypes
1. INTRODUCTION
Alzheimer's disease (AD) is known as the primary cause of dementia worldwide. With the growing knowledge of AD biomarkers and physiopathology of the disease (Jack Jr. et al., 2016), diagnostic criteria have evolved to obtain a common and reliable disease definition that can be used in both research and clinical practice (McKhann et al., 2011). However, it has also long been known that AD is a composite disease with heterogeneous subtypes in terms of genetics, with more than 20 loci described as part of the pathophysiology (Karch & Goate, 2015), with early and late age at onset, characterized by difference in distribution of cortical hypometabolism (Frisoni et al., 2005; Kim et al., 2005), and different clinical presentations, with amnestic, posterior cortical atrophy, progressive primary aphasia, and frontal variants (Alladi et al., 2007; Galton, Patterson, Xuereb, & Hodges, 2000). Even within the typical amnestic presentation, there is heterogeneity in cognitive profiles (Fisher, Rourke, & Bieliauskas, 1999; Lambon, Patterson, Graham, Dawson, & Hodges, 2003; Stopford, Snowden, Thompson, & Neary, 2008; Strite, Massman, Cooke, & Doody, 1997; Vardy et al., 2013) and patterns of cortical hypometabolism of patients (Salmon et al., 2009).
Among the many tools used in the diagnosis and classification of AD, Positron Emission Tomography with [18F]fluorodeoxyglucose (FDG‐PET) is one of the most common neuroimaging technique both in clinical practice and in the research field. In AD, cortical metabolism is typically decreased in the temporoparietal cortex, the precuneus, and the posterior cingulate cortex (Herholz et al., 2002; Minoshima et al., 1997). These regions belong to the default mode network (DMN), involved in self‐directed cognition. The DMN has been divided into three subsystems: the medial‐temporal lobe (MTL) subsystem (including the hippocampal and parahippocampal regions), the Dorsomedial subsystem (including the dorsomedial prefrontal cortex and the temporoparietal junction) and the Midline Core subsystem (including the precuneus, the posterior cingulate cortex, and the anteromedial prefrontal cortex; Andrews‐Hanna, Reidler, Sepulcre, Poulin, & Buckner, 2010). In AD, the MTL subsystem is the most affected in terms of cortical atrophy, with typical hippocampal volume reduction, whereas hypometabolism is particularly important in the core subsystem (Grothe, Teipel,, & Alzheimer's Disease Neuroimaging, 2016).
Beside the classical distribution of cortical hypometabolism, heterogeneity was frequently reported in FDG‐PET among AD patients (Bokde et al., 2001; Foster et al., 1983; Kim et al., 2005; Salmon et al., 2009). With this heterogeneity in mind, the goal of the current study was to characterize a peculiar pattern of FDG‐PET hypometabolism where decreased activity is prominent in the perisylvian area on visual analysis by comparison to a more typical posterior medial cortical involvement (typical pattern) in AD patients with an amnestic clinical profile.
The first step in the current study was to study a small sample of patients from the Memory Clinic, CHU Liege, to statistically confirm the visual analysis of predominant perisylvian hypometabolic pattern. The second step was to analyze clinical profiles related to the metabolic subtype in a larger group of AD patients, using the ADNI database.
2. MATERIALS AND METHODS
2.1. Population 1: GIGA CRC group
2.1.1. Participants
Subjects referred to our nuclear medicine department for differential diagnosis of AD type dementia were selected based on FDG‐PET visual inspection.
A first group of 31 patients was gathered because FDG‐PET was characterized by a predominant posterior perisylvian hypometabolism at visual inspection. Brain CT showed predominant atrophy in the same regions. A second group (named typical), gathered over 1 year, included 25 subjects with a classical AD‐like cortical hypometabolism on FDG‐PET, comprising involvement of the temporoparietal and posteromedial cortices (Herholz et al., 2002). The later participants were selected to have similar age and gender compared to the patients with major perisylvian involvement (the new temporoparietal junction or TPJ subtype). The demographic data are represented in Table 1, and t‐tests show no significant difference between the typical and TPJ subtype groups in terms of age at PET time, age at first symptoms, age at diagnosis, modified Hachinski ischemic scale (MIHS) score (Hachinski, Oveisgharan, Romney, & Shankle, 2012) and mini‐mental state examination (MMSE) score (Folstein, Folstein, & McHugh, 1975).
Table 1.
T test results studying the homogeneity of the “TPJ subtype” group and the typical group in the CRC population
| TPJ subtype (CRC) | Typical (CRC) | N TPJ | N typical | t value | Degree of freedom | p–value | |
|---|---|---|---|---|---|---|---|
| Age at PET (years) | 78.9 ± 6.7 | 78.3 ± 8.0 | 30 | 25 | 0.29 | 53 | .77 |
| Age at first symptom (years) | 76.4 ± 6.9 | 75.3 ± 8.4 | 25 | 25 | 0.49 | 48 | .62 |
| Age at diagnosis (years) | 78.7 ± 6.4 | 77.4 ± 8.2 | 30 | 25 | 0.62 | 53 | .54 |
| MHIS | 1.33 ± 1.75 | 1.58 ± 1.56 | 30 | 24 | −0.58 | 52 | .59 |
| MMSE | 23.04 ± 4.18 | 20.90 ± 4.55 | 26 | 21 | 1.67 | 45 | .10 |
Note: Values expressed as mean ± SD; MIHS, modified Hachinski ischemic score; MMSE, mini‐mental state exam; TPJ, temporoparietal.
All subjects met National Institute on Aging‐Alzheimer's Association clinical criteria for possible AD dementia (McKhann et al., 2011). Due to the retrospective design of this part of the study, biomarker information was limited to neurodegeneration (variable hippocampal atrophy) and patients did not have consistent neuropsychological data to be compared between groups.
For group comparison in SPM12, a control group of 12 healthy older adults without cognitive complaints consisted in participants who underwent a cerebral FDG‐PET on the same scanner as the patients. The control subjects were significantly younger than the AD subjects and age was introduced as a covariate of no interest in the statistical analysis of FDG‐PET.
2.1.2. PET‐scan acquisition and processing
FDG‐PET scan was performed 30 min after intravenous injection of ~150 MBq of FDG using a Gemini TF or Gemini TF Big‐Bore PET‐CT scanner (Philips Medical Systems, Amsterdam, the Netherlands) with a 18 cm axial field of view and a 4.8 mm resolution in air. Patients had their eyes closed. A low‐dose CT was acquired for attenuation correction, followed by a 12‐min emission scan. The images were reconstructed using a LOR‐RAMLA algorithm and corrections for attenuation, dead‐time, random events, and scatter were applied. Reconstructed images had 2 mm isotropic spatial resolution and a 128 × 128 × 90 matrix size.
Preprocessing and statistical analyses of FDG‐PET images were conducted with SPM12 (Wellcome Centre for Human Neuroimaging, UCL, London, UK). Spatial normalization was first obtained by matching each subject's PET image to the MNI PET template provided in SPM. Those normalized images were then averaged to create an FDG‐PET group‐specific template smoothed with a Gaussian kernel of 8 mm full width at half maximum. This group‐specific template was then used to spatially normalize patients and controls' PET images. For this classical statistical analysis only, normalized images underwent a smoothing with a Gaussian kernel of 12 mm full width at half maximum.
For statistical comparison, processed FDG‐PET images of the three groups were entered in a General Linear Model in SPM12 using a factorial design to perform statistical comparison between the two patient groups (TPJ and typical subtypes), and healthy controls using proportional scaling by cerebral global mean values to control for FDG uptake variability. Age, mean‐centered on the overall mean, was entered as a nuisance variable. The threshold for statistical significance was set at p < .05 with correction for multiple comparisons (FWER, family‐wise error rate) at the voxel level over the whole brain volume.
2.2. Population 2: ADNI database
2.2.1. Selection of subjects
Variability in clinical profile associated with the two FDG‐PET (classical and TPJ) subtypes observed in our initial sample was evaluated in a larger population well‐characterized at the neuropsychological level, using ADNI (Alzheimer's disease neuroimaging initiative) database of AD patients (for up‐to‐date information, see http://www.adni-info.org).
All included subjects underwent the same neuropsychological tests, as well as cerebral FDG‐PET. We selected all 241 mild AD patients for which a FDG‐PET image was available at baseline in this database. On visual inspection, thirty‐four subjects were discarded because FDG‐PET images were of suboptimal quality. The FDG‐PET data (n = 207) corresponded to the raw images recorded at their entrance in the study (ADNI1 and ADNI2 baseline). Beside neuropsychological variables, we also collected data regarding age at PET scan, gender, level of education and amyloid burden (estimated with [18F]Florbetapir PET).
Furthermore, similarly to the CRC data, the ADNI were also visually inspected and manually labeled as showing the classical AD‐like cortical or the new TPJ hypometabolic profile.
2.2.2. FDG‐PET analysis
Raw FDG‐PET images from ADNI database were preprocessed in SPM12 following a similar procedure as for Population 1, without smoothing.
The visually labeled data in the CRC dataset were used to construct a machine learning‐based classifier to predict FDG‐PET labels in the 207AD subjects from ADNI. A group‐selection procedure (Wehenkel, Sutera, Bastin, Geurts, & Phillips, 2018) with 500 Extra‐trees (Geurts, Ernst, & Wehenkel, 2006) based on the automated anatomical labelling (AAL) atlas (Tzourio‐Mazoyer et al., 2002) was used to reduce the feature set before learning. Extra‐trees are a variant of Breiman's Random forests (Breiman, 2001), which replaces bootstrap sampling in this latter method by a randomization of the discretization thresholds. This often results in improved performance (Geurts et al., 2006). The method depends on two parameters, the number T of trees in the ensemble and the number K of features drawn at each decision tree node. In all experiments in this article, T was fixed to 500 and K was set to its default value, which is the square root of the total number of input features. Our group selection procedure (Wehenkel et al., 2018) exploits variable importance scores derived from Random forests models and random permutations to rank groups of features, corresponding in our case to brain regions from the AAL atlas (Tzourio‐Mazoyer et al., 2002), according to their relevance for predicting the output class. We used the CER r variant of this procedure with a significance threshold set to 0.05 and, for increased robustness, we retained the regions that were selected more than half of the time in 10 repetitions of 10‐fold cross‐validation.
Performance of the classifier was assessed by cross‐validation and reverse learning. The accuracy of the CRC data based classifier was estimated through a 10 repeated 10‐fold cross‐validation scheme. Afterward, ADNI labels inferred from CRC data were compared to those obtained by visual inspection. Likewise, a new Extra‐trees model was fitted from ADNI with the same parameters and feature reduction, then this model was tested on the GIGA CRC population. These transfer and reverse learning procedures, combined with the manually obtained labels, allowed the evaluation of the performance of the classifiers, while controlling for the potential subjectivity of human raters.
As the classifier provides class probability estimates, the performance was measured by computing the area under receiver operating characteristic (ROC) curves (AUC).
2.2.3. Neuropsychological data
From all the information contained in the ADNI database, we selected the following tests or scales of interest that were available for all participants: Alzheimer's Disease Assessment Scale (ADAS 13), Neuropsychiatric Inventory (NPI), Clinical Dementia Rating Scale (CDR), Mini Mental State Exam (MMSE), Rey Auditory‐Verbal Learning Test (RAVLT), animal fluency, Trail Making Test (TMT A and B), Logical Memory immediate and delayed recall, and the Boston Naming Test (BNT). Data from Digit Span and vegetable fluency were not selected because of too many missing values.
Using the labels predicted from the classifier (trained on the first population to predict the label in the second one), machine learning was used to construct a new model discriminating between the two AD groups, based on neuropsychological scores. In order to have good interpretability of the problem, the model chosen was a Random forests ensemble (Extra‐trees). The analysis of variable importance scores derived from this model (Louppe, Wehenkel, & Geurts, 2013) enabled the identification of the most relevant neuropsychological scores to differentiate the two groups. Importance scores were averaged over 10 repeats to obtain this ranking. Posthoc Student t‐test was used to confirm between‐group differences that had clinical meaning.
2.2.4. Interregional correlations in FDG‐PET data in the ADNI population
In order to better characterize the cerebral networks affected in the two groups of subjects, SPM12 multiple regression was used to search for interregional metabolic correlations, between peak voxel values of the most discriminant regions obtained in our first population (left precuneus and left temporoparietal junction reported in Table 2, taken as seed regions) and FDG‐PET metabolism in the other regions of the brain, in the ADNI population. The threshold for statistical significance was set at p < .05 with correction for multiple comparisons (FWER, family‐wise error rate) at the voxel level for the whole brain.
Table 2.
MNI coordinates of the different regions showing hypometabolism in FDG‐PET
| Region | x | y | z | Cluster size | |
|---|---|---|---|---|---|
| 1.Typical (<control) | Left precuneus | −15 | −55 | 73 | 4,141 |
| 2. Atypical (<control) | Left precuneus | −15 | −55 | 76 | 1,601 |
| 3.Typical (<atypical) | Left precuneus | −3 | −46 | 46 | 104 |
| Right latero‐inferior temporal cortex | 57 | −52 | −11 | 30 | |
| 4. Atypical (<typical) | Left TPJ | 60 | −4 | 7 | 229 |
Note: p < .05 FWER corrected.
3. RESULTS
3.1. FDG‐PET statistical analysis in Population 1 (GIGA CRC group)
The results of the SPM12 group comparisons are summarized in Table 2. When each group was separately compared to the control group, both AD types showed a significant decrease of metabolic activity in the left precuneus (Figure 1). The direct comparison between the typical and the TPJ subtype group (with perisylvian involvement) revealed that the typical subjects showed significant hypometabolism in the left precuneus and the right inferior temporal cortex compared to the TPJ subtype, whereas the TPJ subtype subjects showed a significant decrease of metabolic activity in the temporoparietal junction compared to typical subjects (Figure 2). This analysis thereby statistically confirmed and specified the visual inspection.
Figure 1.
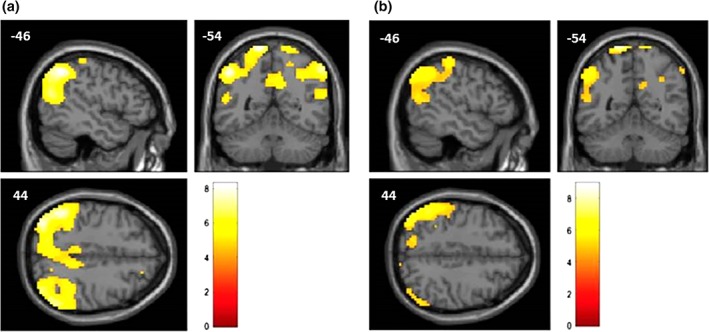
Representation, on a standard structural magnetic resonance (MR) image, of the cortical regions showing a significant metabolic decrease (measured with FDG‐PET) in the typical group (a) and the TPJ subtype group (b) compared to the control group, using the age as a nuisance variable. The regions, represented in the MNI space, are mostly posterior associative cortices. Color scale represents t‐value ((a, degree of freedom = 36; b, degree of freedom = 42)
Figure 2.
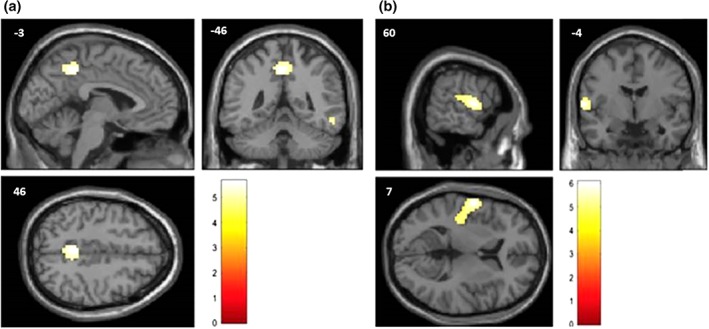
(a) Representation, on a standard structural MRI, of the cortical regions showing a significant metabolic decrease in the typical group compared to the TPJ subtype group, using the age as a nuisance variable. The regions, represented in the MNI space, are mostly the precuneus and the right latero‐inferior temporal cortex. (Table 2). (b) Representation, on a standard structural MRI, of the cortical regions showing a significant metabolic decrease in the TPJ subtype group compared to the typical group, using the age as a nuisance variable. The region, represented in the MNI space, is mostly the parieto‐temporal junction. (Table 2). Color scale represents t‐value (degree of freedom = 55)
3.2. Population 2: Classification of ADNI FDG‐PET data and analysis of clinical profiles
3.2.1. Classification of subjects
The feature selection procedure highlighted 11 relevant regions of interest for subsequent classification (the left rolandic operculum, the superior parietal gyri, the right angular gyrus, the bilateral precuneus, the Heschl gyri, the left superior temporal gyrus, the right middle temporal gyrus, and the right inferior temporal gyri), based on the AAL (Tzourio‐Mazoyer et al., 2002). We thus reduced our feature set to only these regions and fitted an ensemble of 500 Extra‐trees on the learning set. The cross‐validation assessment of this selection and classification on the GIGA‐CRC data lead to an area under the curve of 71.73% (±2.87%).
Using only these regions of interest, the classifier was able to distinguish two groups in the ADNI population similar to the initial two subtypes observed in the GIGA CRC data. Out of 207 subjects, the percentage of TPJ subtype subjects was 38% (78 participants, while 129 subjects FDG‐PET were labeled as typical). The comparison between this prediction and the manually labeled ADNI data showed an area under the curve value of 82.52% (±0.48%). This shows a good level of consistency between the two approaches, transfer learning and manual labelling, even though there is no “ground truth” available.
Using the reverse learning method with the CRC population, the area under the curves showed 86.11% (± 0.36%) accuracy of our classifier.
Considering the labels defined by the classifier, the TPJ subtype subjects from the ADNI population were significantly older than typical subjects (p < .001) but did not differ in terms of level of education or MMSE (Table 3). The proportion of amyloid positive PET was similar in both groups (62% in TPJ, 77% in typical patients, chi‐square 2.344, p = .126), with the limitation that only half of the subjects underwent a [18F]Florbetapir PET.
Table 3.
T test results studying the homogeneity of the “TPJ subtype” group and the “typical” group in the ADNI population
| TPJ subtype (ADNI) | Typical (ADNI) | N TPJ | N typical | t value | Degree of freedom | p–value | |
|---|---|---|---|---|---|---|---|
| Age at PET | 77.47 ± 7.73 | 72.84 ± 7.59 | 78 | 129 | −4.27 | 202 | < .001 |
| Level of education | 15.26 ± 3.12 | 15.47 ± 2.64 | 0.52 | .09 |
Note: Values expressed as mean ± SD.
3.2.2. Neuropsychological results
Machine learning was used to construct a model discriminating one group from the other, based on neuropsychological scores. In this model, the features are the neuropsychological scores and the corresponding output is the group of the AD patient previously predicted by the classifier model. The only test that consistently occupied the first position throughout the 10 repeats from the Extra‐trees model was the TMT‐A (time of completion). The importance of this test to discriminate between the subjects was further confirmed with a t‐test, and TMT‐A response times was significantly faster in the TPJ subtype group (M = 57.97 s, SD = 29.2) than in the typical group (M = 73.23 s, SD = 40.17; t [203] = 2.99, p < .005). This was confirmed when performing correction for multiple comparisons of the scores selected in Section 2.2.3.
3.2.3. Interregional correlations in FDG‐PET
In the typical group, activity in the left precuneus (seed: x = −3, y = −46, z = 46) mostly significantly correlated with metabolism in parietal and premotor regions (Figure 3a). In the TPJ subtype, activity in the left temporoparietal junction (seed: x = −60, y = −4, z = 7) essentially correlated with metabolism in perisylvian regions (Figure 3b).
Figure 3.
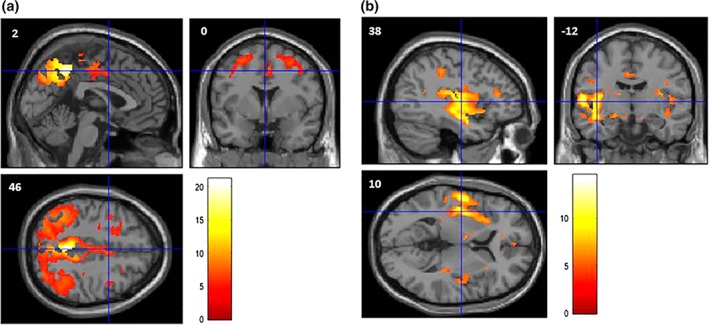
Interregional metabolic correlations, between peak voxel values of the most discriminant regions taken as “seed regions” (obtained in the GIGA‐CRC population) and metabolism in the other regions of the brain, in the ADNI population, with p‐value <.05 (FWER corrected). MNI space. Color scale represents t‐value (degree of freedom = 205). (a) In the “Typical” group, activity in the left precuneus correlates with metabolism in parietal and premotor regions. (b) In the “TPJ subtype” group, activity in the left temporoparietal junction correlates with metabolism in perisylvian regions
4. DISCUSSION
In this study, we report a new metabolic subtype with decreased glucose metabolism (and atrophy) predominant in the perisylvian region, that could be referred to as temporoparietal junction (TPJ) subtype. Initially detected by visual inspection in patients from a clinical sample, significant differences in metabolic activity between this subtype (TPJ hypometabolism) and a more typical group with predominant hypometabolism of the posteromedial cortex were confirmed using SPM12. A Extra‐trees classifier was then constructed based on this first analysis and was applied to an ADNI population. With this classifier, we were able to label 78 ADNI AD patients as TPJ subtypes out of 207 subjects with a probable mild AD diagnosis, with high accuracy level (85.84% of correct labelling with a reverse learning method). Accordingly, the TPJ subtype observed in about 38% of ADNI amnestic AD patients appears as a consistent variant of the typical FDG‐PET profile.
Heterogeneity of regional brain pathology is well‐documented in AD. Subtypes have been described from a neuropathological viewpoint, with the observation of typical, limbic, and cortical subtypes (Murray et al., 2011) and with the description of Alzheimer's pathology in atypical clinical presentations (Alladi et al., 2007; Galton et al., 2000). These neuropathological presentations correlate with the heterogeneity observed in neuroimaging studies with MRI (Ridgway et al., 2012; Whitwell et al., 2012) or FDG‐PET (Bokde et al., 2001; Foster et al., 1983; Kim et al., 2005). Based on these findings of heterogeneity, AD is currently seen as a wide spectrum with different clinical and neuropsychological presentations (amnestic, posterior cortical atrophy, progressive primary aphasia, and frontal variants) that are reflected in imaging and neuropathological studies (Galton et al., 2000; McKhann et al., 2011; Murray et al., 2011). These correlations have also been studied in the early onset presentation of the disease: typical amnestic early‐onset AD, logopenic progressive aphasia and posterior cortical atrophy were associated with disease‐specific reductions in cortical thickness (Ridgway et al., 2012).
Clinical and neuropsychological heterogeneity has also been associated with different patterns of cortical hypometabolism. Different networks are specifically involved depending on the neuropsychological presentation. Although the Default Mode Network (DMN) is involved in all AD subtypes, focal patterns of hypometabolism characterize each variant: the left temporoparietal cortex in the logopenic variant, the ventral DMN in the early‐onset or the bilateral prefrontal cortex in the executive/frontal variants (Lehmann et al., 2013; Vanhoutte et al., 2017).
Even if both regions belong to the DMN, predominant TPJ and posteromedial hypometabolism do not seem to reflect variants of a same network breakdown. Indeed, with seed‐based interregional correlation analysis previously used to reveal metabolic patterns in AD (Lehmann et al., 2013; Seeley, Crawford, Zhou, Miller, & Greicius, 2009), we found a predominant correlation of the metabolism of the left precuneus with parietal and premotor regions in the typical group, and a prominent correlation between the metabolism of the left TPJ and that of perisylvian regions in the TPJ subtype. The TPJ subtype was not characterized by severe medial temporal involvement or by relative sparing of the hippocampus compared to the typical subtype (Ferreira et al., 2017; Murray et al., 2011). It cannot be considered as a diffuse involvement of associative cortices (Noh et al., 2014; Park et al., 2017) or as a group with no atrophy (Ferreira et al., 2017).
ADNI data allowed examining whether these different metabolic profiles are associated with distinct clinical presentations. Actually, the two groups (typical and TPJ subtype) could only be distinguished by age (i.e., TPJ subtype patient being older) and by very few, nonmemory neuropsychological tests such as TMT‐A, with faster response time in the TPJ subtype group compared to the typical subtype group. As TMT‐A test mainly involves visuomotor and attention abilities, we could assume that these abilities are more affected in the typical group. Interestingly, in an fMRI study, brain activity was observed in motor, premotor, and visual areas in healthy subjects performing a TMT‐A task (Karimpoor et al., 2017). Therefore, the metabolic connectivity we described between premotor regions and metabolically impaired left precuneus in the typical group could participate to their lower performance at this test. Nevertheless, apart from this difference, patients from the two types could not be clearly distinguished in terms of cognitive or neuropsychiatric profile. Clinically, they were all found to fulfill the clinical diagnosis of amnestic AD (McKhann et al., 2011). Moreover, compared to healthy older participants, they all demonstrated reduced metabolism in the precuneus. Also, most of the patients had significant amyloid burden on PET in both groups. Yet, the prominent cortical site of neurodegeneration varied between them.
5. CONCLUSION
The current findings suggest that a relatively comparable clinical presentation in AD can be accompanied by a typical FDG‐PET pattern or by a clearly distinct metabolic TPJ subtype, although one cannot exclude that current clinical evaluation was not sensitive enough to differentiate the two groups.
ACKNOWLEDGMENT, CONFLICTS, AND FUNDING SOURCES
The authors have no conflict of interest. F. M. is a Candidate PhD Specialist, M. W. is a Research Fellow, C. P. is a Senior Research Associate, and C. B. is a Research Associate at the FRS‐FNRS (Belgian Fund for Scientific Research).
The authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or nonfinancial‐interest in the subject matter or materials discussed in this manuscript.
Data collection and sharing for this project was funded by the Alzheimer's Disease Neuroimaging Initiative (ADNI; National Institutes of Health Grant U01 AG024904) and DOD ADNI (Department of Defense award number W81XWH‐12‐2‐0012). ADNI is funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering, and through generous contributions from the following: AbbVie, Alzheimer's Association; Alzheimer's Drug Discovery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen; Bristol‐Myers Squibb Company; CereSpir, Inc.; Cogstate; Eisai Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly and Company; EuroImmun; F. Hoffmann‐La Roche Ltd and its affiliated company Genentech, Inc.; Fujirebio; GE Healthcare; IXICO Ltd.; Janssen Alzheimer Immunotherapy Research & Development, LLC.; Johnson & Johnson Pharmaceutical Research & Development LLC.; Lumosity; Lundbeck; Merck & Co., Inc.; Meso Scale Diagnostics, LLC.; NeuroRx Research; Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer Inc.; Piramal Imaging; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Private sector contributions are facilitated by the Foundation for the National Institutes of Health (http://www.fnih.org). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Therapeutic Research Institute at the University of Southern California. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California.
Meyer F, Wehenkel M, Phillips C, et al. Characterization of a temporoparietal junction subtype of Alzheimer's disease. Hum Brain Mapp. 2019;40:4279–4286. 10.1002/hbm.24701
Funding information Department of Defense, Grant/Award Number: W81XWH‐12‐2‐0012; National Institutes of Health, Grant/Award Number: U01 AG024904; Alzheimer's Disease Neuroimaging Initiative
REFERENCES
- Alladi, S. , Xuereb, J. , Bak, T. , Nestor, P. , Knibb, J. , Patterson, K. , & Hodges, J. R. (2007). Focal cortical presentations of Alzheimer's disease. Brain, 130(Pt 10), 2636–2645. 10.1093/brain/awm213 [DOI] [PubMed] [Google Scholar]
- Andrews‐Hanna, J. R. , Reidler, J. S. , Sepulcre, J. , Poulin, R. , & Buckner, R. L. (2010). Functional‐anatomic fractionation of the brain's default network. Neuron, 65(4), 550–562. 10.1016/j.neuron.2010.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokde, A. L. , Pietrini, P. , Ibanez, V. , Furey, M. L. , Alexander, G. E. , Graff‐Radford, N. R. , … Horwitz, B. (2001). The effect of brain atrophy on cerebral hypometabolism in the visual variant of Alzheimer disease. Archives of Neurology, 58(3), 480–486. [DOI] [PubMed] [Google Scholar]
- Breiman, L. (2001). Random forests. Machine Learning, 45(1), 5–32. [Google Scholar]
- Ferreira, D. , Verhagen, C. , Hernandez‐Cabrera, J. A. , Cavallin, L. , Guo, C. J. , Ekman, U. , … Westman, E. (2017). Distinct subtypes of Alzheimer's disease based on patterns of brain atrophy: Longitudinal trajectories and clinical applications. Scientific Reports, 7, 46263 10.1038/srep46263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, N. J. , Rourke, B. P. , & Bieliauskas, L. A. (1999). Neuropsychological subgroups of patients with Alzheimer's disease: An examination of the first 10 years of CERAD data. Journal of Clinical and Experimental Neuropsychology, 21(4), 488–518. 10.1076/jcen.21.4.488.887 [DOI] [PubMed] [Google Scholar]
- Folstein, M. F. , Folstein, S. E. , & McHugh, P. R. (1975). "mini‐mental state". A practical method for grading the cognitive state of patients for the clinician. Journal of Psychiatric Research, 12(3), 189–198. [DOI] [PubMed] [Google Scholar]
- Foster, N. L. , Chase, T. N. , Fedio, P. , Patronas, N. J. , Brooks, R. A. , & Di Chiro, G. (1983). Alzheimer's disease: Focal cortical changes shown by positron emission tomography. Neurology, 33(8), 961–965. [DOI] [PubMed] [Google Scholar]
- Frisoni, G. B. , Testa, C. , Sabattoli, F. , Beltramello, A. , Soininen, H. , & Laakso, M. P. (2005). Structural correlates of early and late onset Alzheimer's disease: Voxel based morphometric study. Journal of Neurology, Neurosurgery, and Psychiatry, 76(1), 112–114. 10.1136/jnnp.2003.029876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galton, C. J. , Patterson, K. , Xuereb, J. H. , & Hodges, J. R. (2000). Atypical and typical presentations of Alzheimer's disease: A clinical, neuropsychological, neuroimaging and pathological study of 13 cases. Brain, 123(Pt 3), 484–498. [DOI] [PubMed] [Google Scholar]
- Geurts, P. , Ernst, D. , & Wehenkel, L. (2006). Extremely randomized trees. Machine Learning, 63, 3–42. [Google Scholar]
- Grothe, M. J. , Teipel, S. J. , & Alzheimer's Disease Neuroimaging Initiative . (2016). Spatial patterns of atrophy, hypometabolism, and amyloid deposition in Alzheimer's disease correspond to dissociable functional brain networks. Human Brain Mapping, 37(1), 35–53. 10.1002/hbm.23018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hachinski, V. , Oveisgharan, S. , Romney, A. K. , & Shankle, W. R. (2012). Optimizing the Hachinski ischemic scale. Archives of Neurology, 69(2), 169–175. 10.1001/archneurol.2011.1698 [DOI] [PubMed] [Google Scholar]
- Herholz, K. , Salmon, E. , Perani, D. , Baron, J. C. , Holthoff, V. , Frolich, L. , … Heiss, W. D. (2002). Discrimination between Alzheimer dementia and controls by automated analysis of multicenter FDG PET. NeuroImage, 17(1), 302–316. [DOI] [PubMed] [Google Scholar]
- Jack, C. R., Jr. , Bennett, D. A. , Blennow, K. , Carrillo, M. C. , Feldman, H. H. , Frisoni, G. B. , … Dubois, B. (2016). A/T/N: An unbiased descriptive classification scheme for Alzheimer disease biomarkers. Neurology, 87(5), 539–547. 10.1212/WNL.0000000000002923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karch, C. M. , & Goate, A. M. (2015). Alzheimer's disease risk genes and mechanisms of disease pathogenesis. Biological Psychiatry, 77(1), 43–51. 10.1016/j.biopsych.2014.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karimpoor, M. , Churchill, N. W. , Tam, F. , Fischer, C. E. , Schweizer, T. A. , & Graham, S. J. (2017). Tablet‐based functional MRI of the trail making test: Effect of tablet interaction mode. Frontiers in Human Neuroscience, 11, 496 10.3389/fnhum.2017.00496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, E. J. , Cho, S. S. , Jeong, Y. , Park, K. C. , Kang, S. J. , Kang, E. , … Na, D. L. (2005). Glucose metabolism in early onset versus late onset Alzheimer's disease: An SPM analysis of 120 patients. Brain, 128(Pt 8), 1790–1801. 10.1093/brain/awh539 [DOI] [PubMed] [Google Scholar]
- Lambon, R.,. M. A. , Patterson, K. , Graham, N. , Dawson, K. , & Hodges, J. R. (2003). Homogeneity and heterogeneity in mild cognitive impairment and Alzheimer's disease: A cross‐sectional and longitudinal study of 55 cases. Brain, 126(Pt 11), 2350–2362. 10.1093/brain/awg236 [DOI] [PubMed] [Google Scholar]
- Lehmann, M. , Ghosh, P. M. , Madison, C. , Laforce, R., Jr. , Corbetta‐Rastelli, C. , Weiner, M. W. , … Rabinovici, G. D. (2013). Diverging patterns of amyloid deposition and hypometabolism in clinical variants of probable Alzheimer's disease. Brain, 136(Pt 3), 844–858. 10.1093/brain/aws327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Louppe, G. , Wehenkel, L.,. A.,. S. , & Geurts, P. (2013). Understanding variable importances in forests of randomized trees In Burges C. J. C., Bottou L., Welling M., Ghahramani Z., & Weinberger K. Q. (Eds.), Advances in neural information processing systems 26 (NIPS 2013) (Vol. 26, pp. 431–439). NY: Curran Associates, Inc. [Google Scholar]
- McKhann, G. M. , Knopman, D. S. , Chertkow, H. , Hyman, B. T. , Jack, C. R., Jr. , Kawas, C. H. , … Phelps, C. H. (2011). The diagnosis of dementia due to Alzheimer's disease: Recommendations from the National Institute on Aging‐Alzheimer's Association workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement, 7(3), 263–269. 10.1016/j.jalz.2011.03.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minoshima, S. , Giordani, B. , Berent, S. , Frey, K. A. , Foster, N. L. , & Kuhl, D. E. (1997). Metabolic reduction in the posterior cingulate cortex in very early Alzheimer's disease. Annals of Neurology, 42(1), 85–94. 10.1002/ana.410420114 [DOI] [PubMed] [Google Scholar]
- Murray, M. E. , Graff‐Radford, N. R. , Ross, O. A. , Petersen, R. C. , Duara, R. , & Dickson, D. W. (2011). Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: A retrospective study. Lancet Neurology, 10(9), 785–796. 10.1016/S1474-4422(11)70156-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noh, Y. , Jeon, S. , Lee, J. M. , Seo, S. W. , Kim, G. H. , Cho, H. , … Na, D. L. (2014). Anatomical heterogeneity of Alzheimer disease: Based on cortical thickness on MRIs. Neurology, 83(21), 1936–1944. 10.1212/WNL.0000000000001003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, J. Y. , Na, H. K. , Kim, S. , Kim, H. , Kim, H. J. , Seo, S. W. , … Alzheimer's Disease Neuroimaging Initiative . (2017). Robust identification of Alzheimer's disease subtypes based on cortical atrophy patterns. Scientific Reports, 7, 43270 10.1038/srep43270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridgway, G. R. , Lehmann, M. , Barnes, J. , Rohrer, J. D. , Warren, J. D. , Crutch, S. J. , & Fox, N. C. (2012). Early‐onset Alzheimer disease clinical variants: Multivariate analyses of cortical thickness. Neurology, 79(1), 80–84. 10.1212/WNL.0b013e31825dce28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salmon, E. , Kerrouche, N. , Perani, D. , Lekeu, F. , Holthoff, V. , Beuthien‐Baumann, B. , … Herholz, K. (2009). On the multivariate nature of brain metabolic impairment in Alzheimer's disease. Neurobiology of Aging, 30(2), 186–197. [DOI] [PubMed] [Google Scholar]
- Seeley, W. W. , Crawford, R. K. , Zhou, J. , Miller, B. L. , & Greicius, M. D. (2009). Neurodegenerative diseases target large‐scale human brain networks. Neuron, 62(1), 42–52. 10.1016/j.neuron.2009.03.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stopford, C. L. , Snowden, J. S. , Thompson, J. C. , & Neary, D. (2008). Variability in cognitive presentation of Alzheimer's disease. Cortex, 44(2), 185–195. 10.1016/j.cortex.2005.11.002 [DOI] [PubMed] [Google Scholar]
- Strite, D. , Massman, P. J. , Cooke, N. , & Doody, R. S. (1997). Neuropsychological asymmetry in Alzheimer's disease: Verbal versus visuoconstructional deficits across stages of dementia. Journal of the International Neuropsychological Society, 3(5), 420–427. [PubMed] [Google Scholar]
- Tzourio‐Mazoyer, N. , Landeau, B. , Papathanassiou, D. , Crivello, F. , Etard, O. , Delcroix, N. , … Joliot, M. (2002). Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single‐subject brain. NeuroImage, 15(1), 273–289. 10.1006/nimg.2001.0978 [DOI] [PubMed] [Google Scholar]
- Vanhoutte, M. , Semah, F. , Rollin Sillaire, A. , Jaillard, A. , Petyt, G. , Kuchcinski, G. , … Lopes, R. (2017). (18)F‐FDG PET hypometabolism patterns reflect clinical heterogeneity in sporadic forms of early‐onset Alzheimer's disease. Neurobiology of Aging, 59, 184–196. 10.1016/j.neurobiolaging.2017.08.009 [DOI] [PubMed] [Google Scholar]
- Vardy, E. R. , Ford, A. H. , Gallagher, P. , Watson, R. , McKeith, I. G. , Blamire, A. , & O'Brien, J. T. (2013). Distinct cognitive phenotypes in Alzheimer's disease in older people. International Psychogeriatrics, 25(10), 1659–1666. 10.1017/S1041610213000914 [DOI] [PubMed] [Google Scholar]
- Wehenkel, M. , Sutera, A. , Bastin, C. , Geurts, P. , & Phillips, C. (2018). Random forests based group importance scores and their statistical interpretation: Application for Alzheimer's disease. Frontiers in Neuroscience, 12, 411 10.3389/fnins.2018.00411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitwell, J. L. , Dickson, D. W. , Murray, M. E. , Weigand, S. D. , Tosakulwong, N. , Senjem, M. L. , … Josephs, K. A. (2012). Neuroimaging correlates of pathologically defined subtypes of Alzheimer's disease: A case‐control study. Lancet Neurology, 11(10), 868–877. 10.1016/S1474-4422(12)70200-4 [DOI] [PMC free article] [PubMed] [Google Scholar]