

Abstract
Previous studies that showed decreased brain activation in people with autism spectrum disorder (ASD) viewing expressive faces did not control that participants looked in the eyes. This is problematic because ASD is characterized by abnormal attention to the eyes. Here, we collected fMRI data from 48 participants (27 ASD) viewing pictures of neutral faces and faces expressing anger, happiness, and fear at low and high intensity, with a fixation cross between the eyes. Group differences in whole brain activity were examined for expressive faces at high and low intensity versus neutral faces. Group differences in neural activity were also investigated in regions of interest within the social brain, including the amygdala and the ventromedial prefrontal cortex (vmPFC). In response to low intensity fearful faces, ASD participants showed increased activation in the social brain regions, and decreased functional coupling between the amygdala and the vmPFC. This oversensitivity to low intensity fear coupled with a lack of emotional regulation capacity could indicate an excitatory/inhibitory imbalance in their socio‐affective processing system. This may result in social disengagement and avoidance of eye‐contact to handle feelings of strong emotional reaction. Our results also demonstrate the importance of careful control of gaze when investigating emotional processing in ASD. Hum Brain Mapp 38:5943–5957, 2017. © 2017 Wiley Periodicals, Inc.
Keywords: facial expressions, eye contact, autism, fMRI, amygdala, fear
INTRODUCTION
Others' emotional faces indicate their mental states. Our ability to read facial expressions accurately is therefore essential to understand our social partners. Previous studies have shown that facial emotion perception is a complex process supported by a network of interconnected brain regions that are activated during emotion perception, and referred to as the social brain network [see Brothers, 1990; Vuilleumier and Pourtois, 2007 for a review]. The regions that are part of the social brain network include the amygdala, the fusiform face area (FFA), the superior temporal gyrus/temporoparietal junction (STG/TPJ), the anterior cingulate cortex (ACC), and the ventromedial prefrontal cortex (vmPFC). Although the regions of the social brain network interact during facial emotion processing, they each carry out specific functions. Activity in the amygdala reflects emotional reactivity to signals of threat and is modulated by the intensity level of the facial expression. The FFA is a region that responds selectively to faces [e.g., Kanwisher et al., 1997] and shows enhanced activity for facial emotions signaling threat [e.g., Armony and Dolan, 2002; Vuilleumier et al., 2004] and higher intensity facial emotions [e.g., Surguladze et al., 2003]. The emotional effects on the FFA are thought to result from its direct projection from the amygdala, and to be modulated by visual attention [e.g., Vuilleumier et al., 2001]. The STS/TPJ is involved in processing biological motion [e.g., Puce et al., 1998] and theory of mind [e.g., Saxe and Kanwisher, 2003] and shows enhanced activity for emotional relative to neutral faces [e.g., Engell and Haxby, 2007]. The ACC and of the vmPFC have an important role in automatic emotion regulation [Etkin et al., 2015], and their activity increase during the presentation of threatening stimuli.
Autism spectrum disorder (ASD) is a neurodevelopmental condition associated with significant difficulties in decoding others' emotions [see Harms et al., 2010 for a review] and mental states [e.g., Baron‐Cohen et al., 1999; Castelli et al., 2002; Senju and Johnson, 2009]. As a result, people with ASD are profoundly impaired in the social domain [DSM‐5; American Psychiatric Association, 2013].
The underlying nature of emotion processing difficulties in people with ASD remains unclear. Traditional theoretical accounts have proposed that they have an “impoverished” sense of socio‐emotional engagement and lack interest in other people [e.g., Hobson, 1994; Kanner and Eisenberg, 1957], which could result in less visual interest and neural activity during emotional face perception. However, more recent theoretical accounts suggest that people with ASD might be overly responsive to others' feelings, and particularly to their distress [Hadjikhani et al., 2014; Smith, 2009]. According to this view, overwhelming negative feelings caused by perceiving others' distress might provoke the person with ASD to disengage from the situation—a behavior that could be mistaken for lack of socio‐affective concern. In addition, our group recently reported that eye‐contact may be stressful to those with ASD because of an over‐activation of the rapid threat‐perception subcortical system [Hadjikhani et al., 2017a]. A challenge for research lies in teasing apart these alternatives (lack of concern vs. oversensitivity) empirically.
Previous functional Magnetic Resonance Imaging (fMRI) studies investigated whether the social brain was hypoactivated or hyperactivated during facial emotion perception in ASD. Most previous studies reported a decreased activation [Ashwin et al., 2007; Critchley et al., 2000b; Dapretto et al., 2006; Grelotti et al., 2005; Pelphrey et al., 2002; Pinkham et al., 2008], but others reported an increased activation in ASD [Dalton et al., 2005]; see also [Dichter et al., 2012; Kleinhans et al., 2009; Monk et al., 2009]. Critically, these discrepant findings might potentially be explained by differences in where participants allocated their attention. Indeed, Dalton et al. [2005] monitored participants' gaze patterns and showed that the more they looked at the eye region of the face stimuli, the more their amygdala and fusiform gyrus were activated. Other neuroimaging studies investigating emotion perception have not monitored where participants were looking, and those with ASD might have avoided the eye region, explaining the observed hypoactivation. It is noteworthy that most eye‐tracking studies have shown that people with ASD spontaneously look less at the central parts of faces compared with controls [Klin et al., 2002; Pelphrey et al., 2002; Senju, 2013; Falck‐Ytter and von Hofsten, 2011], and it has also been shown that ASD individuals have a greater tendency to saccade away from the eyes when the information is present in this region, as is the case in fear [Spezio et al., 2006]. Hence, it is possible that hypoactivity in the social brain regions in studies of people with ASD is a simple reflection of lack of attention to the eyes of the face stimuli.
In this study, we showed participants with and without ASD pictures of faces expressing happiness, anger, or fear, and we constrained fixation to the eye region of those face stimuli by means of a fixation cross. Our first goal was to investigate the social brain activation of people with ASD during emotional face perception, when fixation was constrained in the eye region. We hypothesized that the presence of a fixation cross would reveal activation in the social brain for both groups.
In addition, in this experiment, facial emotions were expressed with two levels of intensity (40% or 100%). Indeed, a previous study using faces expressing fear at high and low intensity found that controls, but not those with ASD, modulated social brain regions with intensity [Ashwin et al., 2006]. Given that most of the information needed to identify a face as fearful is located in the eye region [Schyns et al., 2007] it is possible that this effect was due to ASD participants not looking at the eye region and consequently not assessing the intensity of a fearful expression accurately. Thus, we hypothesized that constraining gaze to the eye region would allow people with ASD to show a modulation of their social brain response with intensity when viewing fearful faces to the same extent as controls. No study to date has investigated whether people with and without ASD show differences in the modulation of their social brain responses with intensity for other emotions (i.e., happiness, anger), but we hypothesized that no such difference would be present with participants' fixation maintained to the eye region.
We also aimed at assessing emotion regulation capabilities in people with and without ASD by measuring the strength of the relationship between the key region for emotional regulation (i.e., vmPFC) and the key region for emotional reactivity (i.e., amygdala). In accordance with previous findings, we hypothesized that emotion regulation capabilities could be decreased in ASD [Mazefsky et al., 2013; Richey et al., 2015], especially for emotions signaling an environmental threat (i.e., fear).
Finally, trait anxiety is highly comorbid of ASD [van Steensel et al., 2011] and dramatically influences the processing of facial emotions [e.g., Rossignol et al., 2005]. In particular, trait anxiety was shown to correlate with amygdala activity in previous studies [Phan et al., 2006; Shin and Liberzon, 2010]. Thus, in this experiment, measured trait anxiety in a subsample of participants and assessed the difference in anxiety level between our two groups. We then investigated the relationship between participants' anxiety and their amygdala response, to ensure that group differences in emotion processing could not be attributed to a difference in anxiety level.
METHODS
Participants
Sixty‐eight male participants were recruited. Among those, 36 were clinically diagnosed with ASD using DSM IV‐TR criteria [APA, 2000] in combination with either the Diagnostic Instrument for Social and Communication Disorder [Leekam, et al., 2002] (8 participants) or the Autism Diagnostic Observation Schedule (ADOS) [Lord et al., 2000] (28 participants) and the Autism Diagnostic Interview‐Revised (ADI‐R) [Lord et al., 1994] (27 participants). None of the 32 controls (CON) had a history of psychiatric/neurological disorders. ASD participants were recruited from Lausanne, Brest, and Gothenburg while CON participants were recruited from Lausanne. Participants were scanned at the Centre Hospitalier Universitaire Vaudois in Lausanne. We excluded from the data analysis 20 subjects (9 ASD). Two subjects (1 ASD and 1 CON) were excluded for moving excessively (i.e., movement above a Framewise Displacement threshold of 0.9 mm [Siegel et al., 2014] >20% of time). Nine subjects (5 ASD and 4 CON) were excluded for performing the task too infrequently (<5/8), and nine subjects (3 ASD, 6 CON) because they did not pass the quality control of the data. A total of 48 male participants {27 ASD, see Table 1 for Calibrated Severity Score (CSS) [Gotham et al., 2007; Shumway et al., 2012] on the ADOS (1 module 2, 3 modules 3, 23 modules 4) and scores on ADI‐R (average score from the following subscales: social, communicative, repetitive behaviors, and age of onset)}1 were included in the final analysis. Their age ranged from 9 to 43 years old (mean = 21.91, SD = 9.14), and did not differ between the ASD and CON group (Table 1).
Table 1.
Demographic information
| ASD | CON | Comparison | ||||||
|---|---|---|---|---|---|---|---|---|
| N | Mean (SD) | Range | N | Mean (SD) | Range | t (DF) | P value | |
| Age (years) | 27 | 23.63 (9.86) | 9–43 | 21 | 19.70 (7.74) | 12–43 | 1.50 (46) | 0.14 |
| IQ | 27 | 113.15 (12.36) | 82–140 | 21 | 112 (13.73) | 85–141 | 0.30 (46) | 0.76 |
| STAI‐T | 11 | 47.82 (9.45) | 36–68 | 12 | 37.42 (5.82) | 28–47 | 3.21 (21) | <0.01 |
| ADOS (CSS) | 19 | 6.32 (1.57) | 3–9 | |||||
| ADI‐R (average overall score) | 18 | 41.61 (7.80) | 31–57 | |||||
Demographic information including age and test scores on questionnaires measuring intelligence (IQ), and anxiety (STAI‐T) for participants with ASD and controls (CON). For ASD participants, CSS on the ADOS and average scores on the ADI‐R are also provided (19 were diagnosed with AS, 6 with ASD, and 2 with Pervasive Developmental Disorder Not Otherwise Specified [PDD‐NOS]). SD: standard deviation, N: sample size. Group comparison on those variables using t‐tests, when applicable.
Intelligence Quotient (IQ) scores were obtained using the Wechsler Nonverbal Scale of Ability [Wechsler and Naglieri, 2006] or the Wechsler Abbreviated Scale of Intelligence [Wechsler, 1999]. All participants had an IQ score above 80, and there was no group difference in IQ scores (Table 1). Trait anxiety was evaluated with the State‐Trait Anxiety Inventory (STAI‐T) [Spielberger, et al., 1983] in 23 participants (11 ASD). STAI‐T scores were significantly higher in ASD than in CON participants, which was expected given the high clinical anxiety of ASD people [van Steensel et al., 2011] (see Table 1).
All procedures were in accordance with the Declaration of Helsinki and were approved by the Lausanne University Hospital ethics committee. Written informed consent was obtained from all adult participants and from all parents of participating children. All children participants also gave their oral assent.
Visual Stimuli
Pictures from eight face identities (4F, 4M) were selected from the MacBrain Face Stimulus set (ID 9, 7, 5, 2, 34, 20, 23, 33) [Tottenham et al., 2009]. For each identity, four pictures were selected expressing the fearful, angry, happy, and neutral state. Each emotional face was morphed from neutral to its full emotional expression using the Morph Age software (Creaceed). In addition to the full expression (Fearful100, Happy100, and Angry100), a snapshot was taken in the morphing sequence when the emotion was expressed at 40% of its intensity (Fearful40, Happy40, Angry40). This resulted in 56 different stimuli (seven pictures per each of the eight identities).
Experimental Design
The experiment consisted of two runs. In each of those runs, there were 16 blocks (2 Fearful40, 2 Fearful100, 2 Angry40, 2 Angry100, 2 Happy40, 2 Happy100, 2 Neutral) that were presented in pseudo‐random order. The blocks consisted of eight pictures from different identities that were overlaid by a red fixation cross and presented for 300 ms. The face stimuli were followed by a 1,200 ms fixation screen that consisted of a red fixation cross only, displayed at the center of the monitor. In half of the blocks, the red fixation cross turned blue in one trial. In between blocks, the fixation screen was presented for 3,000 ms. To monitor participants' attention, we asked them to press a button when the blue cross appeared.
Data Acquisition
fMRI data were collected with a 12‐channel radio frequency coil in a Siemens 3T scanner (Siemens Tim Trio, Erlangen, Germany) at the Centre d'Imagerie BioMédicale in Lausanne. The first scanning sequence consisted of Siemens's autoalign scout for the head allowing an automatic positioning and alignment of slices. Anatomical images were acquired using a multi‐echo magnetization prepared rapid gradient echo sequence: 176 slices; 256 × 256 matrix; echo time (TE): TE1: 1.64 ms, TE2: 3.5 ms, TE3: 5.36 ms, TE4: 7.22 ms; repetition time (TR): 2530 ms; flip angle 7°. The functional data were obtained using an echo planar imaging sequence (47 AC‐PC slices, 3 mm thick, 3.12 mm by 3.12 mm in plane resolution, 64 × 64 matrix; field of view: 216; TE: 30 ms; TR: 3000 ms; flip angle 90°) lasting 8 min and 54 s.
Data Analysis
Whole brain (WB) analysis
fMRI data processing, and preprocessing was carried out using FEAT Version 6.0, part of FSL (FMRIB's Software Library, http://www.fmrib.ox.ac.ul/fsl). Non‐brain tissues were removed from high‐resolution anatomical images using Christian Gaser's VBM8 toolbox for SPM8 fed into feat. Data were motion‐corrected using MCFLIRT and motion parameters were added as confound variables to the model. In addition, residual outlier time‐points were identified using FSL's motion outlier detection program and integrated as additional confound variables in the first‐level general linear model analysis. Preprocessing further included spatial smoothing using a Gaussian kernel of 8 mm, grand‐mean intensity normalization and high pass temporal filtering with sigma = 50.0 s.
The two runs (treated as fixed effect) from each participant were combined. Subject‐level statistical analysis was carried out for the following contrasts (Angry40 vs. Neutral, Angry100 vs. Neutral, Fearful40 vs. Neutral, Fearful100 vs. Neutral, Happy40 vs. Neutral, Happy100 vs. Neutral, Angry100 vs. Angry40, Fearful100 vs. Fearful40, Happy100 vs. Happy40) using FILM with local autocorrelation correction. Registration to high‐resolution structural images was carried out using FLIRT. Registration to Montreal Neurological Institute (MNI) standard space was then further refined using FNIRT (FMRIB's nonlinear registration tool) nonlinear registration.
We used age as a covariate given the wide age range of participants and the known effect of development on emotion processing. Group‐level analyses were performed using FLAME 1&2 (general linear mixed model analysis) with automatic outlier detection. In modeling subject variability, this kind of analysis allows inference about the population from which the subjects are drawn. Z‐statistic images were thresholded using clusters determined by Z > 2.3 and a (corrected) cluster significance threshold of P = 0.05 (two‐tails). Cluster‐corrected images were displayed on a standard brain (fsaverage for the surface and MNI template for the volume).
ROI analyses
The ROIs were chosen based on prior knowledge of the regions involved in emotion perception: the amygdala, the vmPFC, the ACC, the right STS/TPJ, and the FFA. All the aforementioned structures except the FFA were defined anatomically using the Harvard‐Oxford probabilistic atlases of FSL. The FFA mask was determined functionally from a previous study, looking at the difference between brain activation for faces and objects [Hadjikhani et al., 2004], because it is defined as the region of maximum sensitivity to faces [Kanwisher et al., 1997]. For each subject, the mean percentage blood‐oxygen‐level dependent signal change was extracted for the five structures and the six contrasts of interest, using the Featquery tool in FSL.
RESULTS
Behavioral Results
During the scanning session, both the ASD and the CON groups detected the change of color in the fixation cross with a high rate (mean = 99.07%, SD = 3.37% in ASD and mean = 98.21%, SD = 4.48% in CON), and there was no group difference for the detection rates of the cross (t(46) = 0.76, P = 0.45), even after age was controlled for (F = 0.84, P = 0.44).
Neuroimaging Results
WB analyses
We computed WB activity for the following contrasts: Fearful40 vs. Neutral, Fearful100 vs. Neutral, Angry40 vs. Neutral, Angry100 vs, Neutral, Happy40 vs. Neutral, Happy100 vs. Neutral, Fearful100 vs. Fearful 40, Angry 100 vs. Angry 40, and Happy100 vs. Happy 40. Within‐group activations and between‐group differences in activity were assessed using independent t‐tests available in FSL.
Within group
The contrasts Happy40 vs. Neutral, Happy100 vs. Neutral, and Happy100 vs. Happy40 yielded activation in CON participants only. The contrast Fearful100 vs. Fearful40 yielded activation in the ASD participants only. The contrasts Angry40 vs. Neutral, Angry100 vs. Neutral, Fearful40 vs. Neutral, Fearful100 vs. Neutral, Angry100 vs. Angry40 however yielded activations in both groups of participants (see Supporting Information Materials).
Between‐group
Participants with ASD showed more activation overall than CON participants for the contrast Fearful40 vs. Neutral (Table 2 and Fig. 1) and Angry100 vs. Angry40 (see Supporting Information Material). In addition, CON participants showed more activation than ASD participants for the contrast Happy100 vs. Neutral (Table 3 and Fig. 2), Fearful100 vs. Fearful40, Happy100 vs. Happy40 (see Supporting Information Materials). There was no group difference at the WB level for the other contrasts.
Table 2.
ASD > CON for fearful 40 versus neutral
| Structure | Side | Cluster size | Z | x | y | z |
|---|---|---|---|---|---|---|
| 1,910 | ||||||
| Middle Frontal Gyrus | R | 4.26 | 36 | 16 | 50 | |
| Precentral Gyrus | R | 3.69 | 34 | −14 | 40 | |
| Angular Gyrus | R | 3.19 | 44 | −46 | 20 | |
| Postcentral Gyrus | R | 3.12 | 22 | −18 | 54 | |
| 1,500 | ||||||
| Caudate | L | 3.58 | −20 | 18 | 8 | |
| Middle Frontal Gyrus | L | 3.55 | −26 | 32 | 28 | |
| Frontal Pole | L | 3.34 | −22 | 38 | 30 | |
| Paracingulate Gyrus | L | 2.98 | −12 | 30 | 34 | |
| 1,340 | ||||||
| Precentral Gyrus | L | 3.5 | −4 | −30 | 60 | |
| Postcentral Gyrus | L | 3.45 | −14 | −36 | 60 | |
| Cingulate Gyrus | R | 2.71 | 6 | −30 | 38 | |
| L | 3.09 | −14 | −26 | 36 | ||
| Pallidum | L | 2.66 | −20 | −10 | 6 | |
| Putamen | L | 2.63 | −24 | −10 | 12 | |
| 1,168 | ||||||
| Paracingulate Gyrus | R | 3.55 | 8 | 40 | 32 | |
| L | 3.36 | 0 | 40 | 24 | ||
| Cingulate Gyrus | R | 3.4 | 10 | 30 | 24 | |
| L | 2.96 | −4 | 38 | 20 | ||
| Frontal Pole | R | 3.12 | 16 | 40 | 44 | |
| Caudate | R | 3.1 | 10 | 10 | 18 | |
| 415 | ||||||
| Middle Temporal Gyrus | R | 3.98 | 68 | −12 | −10 | |
| STG | R | 3.74 | 66 | −8 | −6 | |
| 390 | ||||||
| Brainstem | 3.4 | 0 | −22 | −40 | ||
| 320 | ||||||
| Postcentral Gyrus | L | 3.25 | −60 | −8 | 34 | |
| Precentral Gyrus | L | 3.14 | −52 | −8 | 48 | |
| 295 | ||||||
| Parahippocampal Gyrus | R | 3.72 | 24 | −8 | −30 | |
| Temporal Fusiform Cortex | R | 3.01 | 36 | −12 | −38 | |
| 138 | ||||||
| Planum Temporale | L | 3.35 | −40 | −34 | 6 | |
| 112 | ||||||
| Parahippocampal Gyrus | L | 2.97 | −22 | −4 | −32 | |
| Hippocampus | L | 2.94 | −24 | −10 | −22 | |
| Amygdala | L | 2.86 | −20 | −10 | −16 | |
| 109 | ||||||
| Thalamus | R | 3.61 | 4 | −10 | 2 | |
| L | 2.88 | −8 | −8 | 2 | ||
| 107 | ||||||
| Precuneous Cortex | L | 3.31 | −14 | −68 | 36 | |
| 101 | ||||||
| Temporal Fusiform Cortex | R | 2.99 | 32 | −28 | −26 | |
| Parahippocampal Gyrus | R | 2.79 | 30 | −26 | −18 |
Brain regions more activated in participants with autism spectrum disorder than in controls (ASD > CON) for fearful faces at low intensity vs. neutral faces (P < 0.05).
Figure 1.
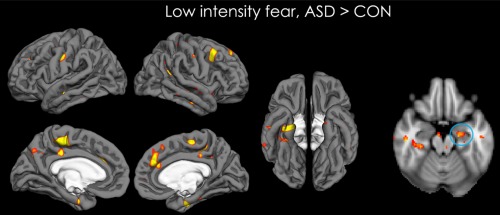
Map of activation showing the brain areas that are more activated for ASD than CON participants, for the low intensity fearful condition. The rightmost image is a transverse view of the brain showing increased activation in the amygdala for ASD vs. CON participants, in this condition. Data are shown for P < 0.05 (unclustered). [Color figure can be viewed at http://wileyonlinelibrary.com]
Table 3.
CON > ASD for happy 100 versus neutral
| Structure | Side | Cluster size | Z | x | y | z |
|---|---|---|---|---|---|---|
| 3,540 | ||||||
| Frontal orbital cortex | L | 4.39 | −34 | 24 | −24 | |
| Inferior frontal gyrus | L | 4.38 | −52 | 10 | 6 | |
| Middle frontal gyrus | L | 4.32 | −48 | 36 | 20 | |
| Frontal pole | L | 4.26 | −48 | 52 | −6 | |
| Precentral gyrus | L | 4.12 | −52 | 10 | 30 | |
| Insular cortex | L | 3.96 | −36 | 4 | −14 | |
| Amygdala | L | 3.45 | −30 | 0 | −12 | |
| Pallidum | L | 3.28 | −22 | 0 | −4 | |
| Temporal pole | L | 3.09 | −46 | 12 | −10 | |
| Frontal operculum cortex | L | 2.68 | −36 | 16 | 8 | |
| 2071 | ||||||
| Occipital pole | R | 3.87 | 24 | −94 | 20 | |
| Lingual gyrus | R | 3.76 | 28 | −60 | −6 | |
| Occipital fusiform gyrus | R | 3.46 | 22 | −74 | −4 | |
| Intracalcarine cortex | R | 3.35 | 20 | −72 | 6 | |
| Temporal occipital fusiform cortex | R | 3.31 | 30 | −54 | −16 | |
| Lateral occipital cortex | R | 3.12 | 42 | −64 | 0 | |
| Middle temporal gyrus | R | 3.08 | 46 | −60 | 2 | |
| 1674 | ||||||
| Parahippocampal gyrus | R | 3.99 | 16 | −30 | −18 | |
| L | 2.64 | −16 | −30 | −16 | ||
| Brainstem | 3.92 | 0 | −22 | −20 | ||
| Hippocampus | R | 3.08 | 18 | −32 | −6 | |
| L | 3.34 | −18 | −34 | −2 | ||
| Putamen | R | 3.33 | 16 | 12 | −12 | |
| Amygdala | R | 3.28 | 30 | −4 | −16 | |
| Frontal orbital cortex | R | 3.28 | 18 | 10 | −24 | |
| Pallidum | R | 3.26 | 26 | −16 | −4 | |
| Lingual gyrus | R | 3.09 | 12 | −46 | −10 | |
| Thalamus | R | 3.07 | 20 | −32 | −2 | |
| 1,149 | ||||||
| Inferior temporal gyrus | L | 3.69 | −50 | −52 | −8 | |
| Lateral occipital cortex | L | 3.67 | −40 | −68 | −6 | |
| Occipital fusiform gyrus | L | 3.66 | −26 | −82 | −6 | |
| Middle temporal gyrus | L | 3.45 | −50 | −56 | 10 | |
| Angular gyrus | L | 3.28 | −54 | −56 | 18 | |
| Temporal occipital fusiform cortex | L | 2.84 | −32 | −54 | −8 | |
| 590 | ||||||
| Central opercular cortex | R | 3.7 | 48 | 8 | 4 | |
| Precentral gyrus | R | 3.66 | 40 | 4 | 24 | |
| Insular cortex | R | 3.46 | 44 | −8 | 6 | |
| Frontal operculum cortex | R | 3.34 | 48 | 12 | 2 | |
| Inferior frontal gyrus | R | 2.6 | 58 | 12 | 8 | |
| 382 | ||||||
| Inferior frontal gyrus | R | 3.47 | 56 | 18 | 22 | |
| Precentral gyrus | R | 3.43 | 42 | −4 | 44 | |
| 355 | ||||||
| Lateral occipital cortex | R | 3.41 | 44 | −90 | 4 | |
| 244 | ||||||
| Occipital pole | L | 3.16 | −22 | −98 | 14 | |
| 183 | ||||||
| Precentral gyrus | R | 3.86 | 12 | −28 | 78 | |
| Postcentral gyrus | L | 3.24 | 20 | −32 | 78 | |
| 179 | ||||||
| Precuneous cortex | L | 3.61 | −2 | −58 | 8 | |
| Lingual gyrus | L | 3.04 | −6 | −62 | 6 | |
| 156 | ||||||
| Temporal pole | R | 3.33 | 54 | 20 | −24 | |
| STG | R | 3.25 | 64 | 2 | −12 | |
| 149 | ||||||
| Lateral occipital cortex | L | 3.36 | −24 | −76 | 46 | |
| 137 | ||||||
| Postcentral gyrus | L | 3.52 | −20 | −30 | 66 | |
| Precentral gyrus | L | 2.96 | −18 | −24 | 74 | |
| 136 | ||||||
| Lingual gyrus | L | 3.24 | −20 | −58 | −2 | |
| 107 | ||||||
| Juxtapositional lobule cortex | L | 2.9 | −6 | 0 | 54 | |
| Paracingulate gyrus | R | 2.85 | 8 | 14 | 50 | |
| Superior frontal gyrus | R | 2.81 | 8 | 12 | 54 | |
| 104 | ||||||
| Superior frontal gyrus | L | 3.55 | −4 | 18 | 62 | |
| 100 | ||||||
| Inferior frontal gyrus | R | 3.29 | 44 | 32 | 2 |
Brain regions more activated in participants with ASD than in controls (CON > ASD) for happy faces at high intensity vs. neutral faces (P< 0.05).
Figure 2.

Map of activation showing the brain areas that are more activated for CON than ASD participants, for the high intensity happy condition. Data are shown for P < 0.05 (unclustered). [Color figure can be viewed at http://wileyonlinelibrary.com]
ROI results
We computed activity for the following ROIs, all part of the social brain network: the amygdala, the vmPFC, the ACC, the right STS/TPJ, and the FFA. Two types of statistical analyses were conducted with those ROIs. For both, age was used as a covariate. First, for each of the ROIs, a two (groups: ASD, CON) by three (emotions: fearful, angry, happy) by two (intensities: high, low) repeated measure analysis of covariance (ANCOVA) was performed. The Greenhouse‐Geisser correction was used when the assumption of sphericity was violated. The alpha level for significance was set to 0.05. When the ANCOVA yielded significant interactions, simple effects were investigated with post‐hoc tests and the Bonferroni correction for multiple comparisons was applied. We also tested the hypothesis of an abnormal coupling between the amygdala and the vmPFC in ASD. For that, we used partial correlations (controlling for the effect of age) to investigate, for each contrast and each group, whether (1) activation in the amygdala and in the vmPFC were correlated and (2) anxiety (as measured by the State‐Trait Anxiety Inventory: STAI‐T) [Spielberger et al., 1983] and activation in the amygdala were correlated.
ANCOVA
In the amygdala, there was a main effect of intensity (F(1,45) = 6.87, MSE = 32946.53, P = 0.01, η p 2 = 0.13; high > low). No other effects were significant. In the vmPFC, the effects of emotion and intensity were not significant and did not interact with group. However, the emotion*intensity*group interaction was significant (F(2,90) = 4.18, MSE = 36999.36, P = 0.02, η p 2 = 0.09). This was characterized by an intensity*group interaction for angry and happy faces (F(1,45) = 5.31, MSE = 36905.04, P = 0.03, η p 2 = 0.11 and F(1,45) = 4.36, MSE = 36012.56, P = 0.04, η p 2 = 0.09) but not for fearful faces (F = 0.13, P = 0.72). For angry faces the intensity effect (high >low) was significant in ASD participants (P bonf = 0.03) but not in controls (P bonf = 0.26). For happy faces, the intensity effect (high > low) was marginally significant in CON participants (P bonf = 0.09) but nonsignificant in ASD participants (P bonf = 0.26). No further effects were significant. In the FFA, there was a significant effect of intensity (F(1,45) = 9.51, MSE = 111069.00, P < 0.01, η p 2 = 0.17; high > low), and significant intensity*emotion interaction (F(2,90) = 4.82, MSE = 50451.06, P < 0.01, η p 2 = 0.11) such that the intensity effect was significant for angry but not happy and fearful faces (F = 22.21, P < 0.01; F = 2.54, P = 0.12; and F = 0.01, P = 0.91, respectively). No other effects were significant, notably there was no group effect. No effects were significant in the ACC and the right STS/TPJ.
Correlations2
For fear, the vmPFC/amygdala correlation was significant in CON participants at low and high intensity (r p = 0.51, P = 0.02; r p = 0.75, P < 0.01), but not in ASD participants. For anger, there was a significant vmPFC/amygdala correlation at low and high intensity in ASD participants (r p = 0.67, P < 0.01; r p = 0.65, P < 0.01), but only a weak correlation at low intensity in CON participants (r p = 0.43, P = 0.06). Finally, for happiness, the vmPFC/amygdala correlation was significant at high intensity (r p = 0.46, P = 0.02) for ASD participants but was non‐significant for CON participants (Fig. 3). No partial correlation between anxiety and amygdala activation reached significance.
Figure 3.

Partial correlations (with age being controlled for) between the amygdala activity (displayed on the x‐axis) and the vmPFC activity (displayed on the y‐axis) for angry (leftmost panel), happy (central panel), and fearful faces (rightmost panel). Low intensity emotions are featured in the upper part of the figure while high intensity emotions are featured in the lower part of the picture. ASD data is represented in blue and CON data is represented in orange. Significant correlation P < 0.05 are indicated with an asterisk (*), P < 0.01 with two asterisks (**), and trending correlations P < 0.07 with a tilde (∼). [Color figure can be viewed at http://wileyonlinelibrary.com]
DISCUSSION
In this study, we demonstrate that (1) ASD participants are hypersensitive to low intensity fearful expressions; (2) the greater intensity of emotional expression is reflected by greater neural activation, although this pattern varies as a function of emotion and group; (3) while there is a vmPFC/amygdala correlation for fearful faces in controls, such a functional correlation is absent in ASD participants; and (4) ASD participants do not differ from controls in the FFA activation when constrained to look in the eye region.
The deficits in social skills of people with ASD have been linked to their impairment in reading others' emotions [Harms et al., 2010] and to their abnormal social brain network activation in response to facial emotion [Ashwin et al., 2007; Critchley et al., 2000b; Dalton et al., 2005; Dapretto et al., 2006; Kim et al., 2015; Monk et al., 2010; Pelphrey et al., 2002]. Many past studies have found that those with ASD hypoactivated critical regions of the social brain such as the amygdala and the FFA when watching emotional faces [Ashwin et al., 2007; Critchley et al., 2000a; Dapretto et al., 2006; Kim et al., 2015; Pelphrey et al., 2002]. However, these studies did not monitor participants' gaze. One study in which gaze was monitored during the presentation of face stimuli found that the extent to which ASD participants activate their social brain in response to observing emotional faces correlated to the amount of time they spent looking at the eyes [Dalton et al., 2005]. In addition, we have recently reported that constraining individuals with ASD to look in the eyes, compared with free‐viewing of dynamic emotional face stimuli, increases activation for all the emotions studied (neutral, happy, angry, and fear) in the subcortical face‐processing system, including the amygdala [Hadjikhani et al., 2017a]. Since the eyes are highly important for decoding emotions [Smith and Schyns, 2009], the difficulties of people with ASD in accurately reading facial expressions could result from their lack of attention to the eyes. These results also raise the possibility that the social brain hypoactivity found in some studies of people with ASD may reflect their lack of attention to the eyes of the face stimuli.
In this study, we used emotional face stimuli with a fixation cross between the eyes that served as a gaze constrain for participants, and investigated whether people with ASD differed from controls in their social brain activation, when fixation was maintained to the eyes. Using such methodological control, we found that, although ASD participants have decreased social brain activations for high intensity happy faces relative to controls, they have increased social brain activations for low intensity fearful faces compared with controls. These results have important methodological implications: in fMRI experiments, stimuli are shown assuming that all participants will process them similarly. For ASD individuals, evidence has shown that when examining face perception, it is important to control what part of the stimulus participants are attending to [Dalton et al., 2005; Hadjikhani et al., 2004], and our findings suggest that some form of control needs to be applied to gain valid results when examining emotion perception.
The notion that people with ASD are oversensitive to fearful faces when cued to the eye region is consistent with several theoretical accounts. This is in agreement with the eye avoidance hypothesis of Tanaka and Sung [2016], according to which people with ASD actively avoid the eye region of faces to cope with the overwhelming physiological response that looking at this region provokes in them [e.g., Kylliainen and Hietanen, 2006] (for a study showing increased skin conductance for direct gaze in ASD vs. CON participants). According to Tanaka and Sung [2016], eye contact is uncomfortable for people with ASD regardless of whether the face is neutral or emotional. However, when faces display a fearful expression, eyes could even be more arousing than when faces are neutral or expressing another emotion, as they signal the presence of an environmental threat, although our group recently reported the presence of an eye‐contact effect for all emotions in ASD [Hadjikhani et al., 2017a]. Further supporting this hypothesis, it was shown that, contrary to controls, ASD participants reflexively shift their attention away from the eye region when perceiving emotional faces [Kliemann et al., 2012]. Gernsbacher and Frymiare [2005] reported that people with ASD find it painful to make eye‐contact. Although a coping strategy, this eye region avoidance might have a profound effect on the ability of people with ASD to decode others' emotion, since emotion are often signaled through subtle changes in the eye region [Adolphs et al., 1994, 2005]. According to Smith [2009], autistic behavior may be an adaptive response to an over‐arousal created by the perception of other's threatening emotions, together with a lack of cognitive empathy/theory of mind. We believe that not only cognitive empathy, but also emotion regulatory processes are crucial to be able to cope with others' emotions, and not feel personal distress when confronted with negative emotions in social situations [Hadjikhani et al., 2014]. It has repeatedly been shown that people with ASD have less efficient emotion regulatory behaviors [Mazefsky et al., 2013; Samson et al., 2015] and our present data are in line with these previous findings.
To cope with a strong emotional reaction (indexed by increased activity in the social brain region), one can either look away, or try to down‐regulate feelings, using reappraisal or suppression. If these strategies do not work, an emotional “meltdown,” so often observed in ASD, can happen. Interestingly, in this study we found that, in CON participants, there was a strong vmPFC/amygdala correlation in response to fearful faces, which could be interpreted as an attempt to regulate emotional reactivity to threat. However, such a correlation was absent in ASD. For anger, the vmPFC/amygdala correlation was strong at both intensities in ASD participants, but was only trending at low intensity in CON participants. These results are tentatively suggestive of more efficient capacities for ASD participants to downregulate their perception of anger (a social threat), but not their perception of fear, which may just be too much, even at low intensities. This oversensitivity could also be clinically linked to the tendency in ASD to look bewildered by fearful expressions that other people would not even notice. There was also an interesting vmPFC/amygdala correlation for happy faces in ASD but not CON participants. An increased regulation of emotional reactivity to happy faces in people ASD could explain their hypoactivation to high intensity happy faces.
We speculate that our results could reflect an imbalance between excitatory and inhibitory (E/I) influences in the socio‐affective processing system in ASD, mirroring E/I imbalance for lower level stimulus processing [Freyberg et al., 2015; Robertson et al., 2013, 2016; Rosenberg et al., 2015]. In a brain with an overactive subcortical system [Hadjikhani et al., 2017a; Zürcher et al., 2013b], and hypersensitive to emotions, a way to decrease emotional reactivity would be to use the frontal parts of the brain to inhibit the emotional response, or to disengage from the face, as previously hypothesized in ASD [Tanaka and Sung, 2016]. Our results have important implications for understanding the socio‐affective abnormalities in ASD. They bring into question the concept that people with ASD are indifferent towards others, and are, as Kanner described them “self‐absorbed, emotionally cold, distanced, and retracted” [Kanner and Eisenberg, 1957]. Quite the contrary, our results indicate that people with ASD may have an oversensitive emotional processing system, at least for the perception of feelings of pain [Hadjikhani et al., 2014] or distress/fear (present data), as well as for positive emotions [Hadjikhani et al., 2017a]. In real‐life settings, this oversensitivity, coupled with less well‐developed emotional regulation, might be expressed as an avoidant behavior and misinterpreted as socio‐affective indifference.
However, it is noteworthy that the oversensitivity of people with ASD to low intensity fearful faces could also simply be due to an increased sensitivity to the eye widening of the fearful expression. This alternative interpretation would be consistent with our finding of hyposensitivity to happy faces, given that happy eyes are smaller than fearful and angry eyes [Bayless et al., 2011]. In a previous study, control participants showed more amygdala activity for masked fearful eyes than for masked happy eyes [Whalen et al., 2004]. Although this remains to be investigated, the modulation of the amygdala activity with the widening of the eyes could be more important in ASD than in CON participants, leading to the observed effects. In addition, fearful and angry eyes are often confused in people with ASD [Wallace et al., 2008]. If people with ASD did confuse angry and fearful faces in this experiment where they had to remain fixated on the eye region, this may explain their increased regulatory response for angry faces relative to controls. However, this hypothesis is put into question by our recent finding of an abnormally high subcortical activation in participants with ASD fixating the eye region of dynamic emotional face stimuli regardless of the expression (happy, angry, fearful) shown on the face [Hadjikhani et al., 2017a]. To clarify this issue, future studies should continue to investigate the way in which the heightened neural response to emotional faces in ASD is modulated by the expression of the face.
There was no group difference in FFA activation in this study. These findings are in contrast with the results of many previous studies [Critchley et al., 2000b; Pelphrey et al., 2002; Pierce et al., 2001; Pinkham et al., 2008; Schultz et al., 2000], but these studies did not monitor the participants' gaze behavior, and FFA activation has been shown to be correlated to the time spent fixating the eyes [Dalton et al., 2005]. A recent study showed that participants with ASD hypoactivate the FFA relative to controls in the free‐viewing condition, but that it normalizes when they look at the eyes [Perlman et al., 2011]. This is in line with our finding that the FFA is activated just as much in people with and without ASD when fixation is constrained in the eye region of emotional faces and with the growing body of studies reporting normal FFA activation in ASD [Bird et al., 2006; Bookheimer et al., 2008; Dapretto et al., 2006; Hadjikhani et al., 2004, 2007; Kleinhans et al., 2008, 2009; Pierce et al., 2004; Piggot et al., 2004; Pinkham et al., 2008; Sato et al., 2012; Wang et al., 2004; Wicker et al., 2008; Zürcher et al., 2013a; Zürcher et al., 2013b].
Similarly, in this study, no group differences in STS/TPJ activation were observed. In contrast to our results, most previous studies that did not constrain fixation on the eye region [Ashwin et al., 2006; Baron‐Cohen et al., 1999; Critchley et al., 2000b; Pelphrey et al., 2002] but see [Kim et al., 2015] found an over‐activation of the STS/TPJ in people with ASD. The STS/TPJ is involved in perspective taking [Ruby and Decety, 2003], empathy [Jackson et al., 2006; Lamm et al., 2007], and theory of mind [Lawrence et al., 2006; Saxe and Wexler, 2005], and the abnormal activation of this region in people with ASD when eyes are not fixated may be linked to their deficits in those domains. Our results suggest that fixation on the eye region normalizes STS/TPJ activation in people with ASD, and we speculate that this normalization could have a positive impact on the functions carried out by this structure. Although previous studies have considered the “typical” scanpath of faces to involve frequent fixation to the eyes [Morris et al., 2007; Perlman et al., 2011], constraining participants' fixation to the eye region may result in a scanpath that is not entirely natural. However, this method has been previously successfully used to demonstrate that people with and without ASD activated their FFA to the same extent when perceiving neutral faces [Hadjikhani et al., 2004], and we recently reported that comparing constrained scanpath with free viewing was resulting in increased activation on the subcortical system in ASD compared with controls [Hadjikhani et al., 2017a], and that it also resulted to increased activation in the social brain in typical controls [Hadjikhani et al., 2017b].
Given the high comorbidity between anxiety and ASD [van Steensel et al., 2011], it is unsurprising that our ASD participants exhibited more trait anxiety than controls. It is not excluded that anxiety played a role in our finding that ASD participants are oversensitive to low intensity fear. Indeed, amygdala activation in response to fearful emotions was previously shown to be increased in participants with clinical anxiety [Phan et al., 2006; Shin and Liberzon, 2010]. In contrast, here, amygdala activity and anxiety did not correlate in either group. However, only a small subset of our participants were assessed for anxiety in this study. Future studies should investigate the possibility of a link between social brain abnormality and anxiety in ASD using neuroimaging methods in a larger sample.
One of the limitations of our study is our use of static emotional face stimuli. Although those have been used extensively to study emotion processing in people with and without ASD, facial expressions are inherently dynamic in natural settings. In addition, dynamic emotional face stimuli are better recognized and activate the social brain more than static emotions [Kamachi et al., 2013; Kilts et al., 2003; Sato et al., 2004; Trautmann et al., 2009]. Of particular interest here is the study by Pelphrey et al. [2002] that found differential social brain activation in ASD participants depending on whether static or dynamic emotional face stimuli were used. For instance, although ASD participants had an increased STS/TPJ activation in response to static emotional faces, they showed a normalization of their STS/TPJ activation, when the facial emotions were presented dynamically. Similarly, FFA and amygdala were found to be hypoactivated in ASD participants when the faces expressed emotions dynamically but not when emotions were static. Although the present study is a first step toward determining the way in which social brain activations are affected by eye fixation in ASD, those previous results [Pelphrey et al., 2002] highlight the importance of replicating our findings using dynamic emotional stimuli.
Another limitation is that we did not compare constrained versus unconstrained gaze during emotion perception, nor did we use an eye‐tracking to monitor whether participants were indeed looking in between the eyes of the emotional face stimuli. Those controls should be included in future studies attempting to replicate our findings. However, given that the decrease in emotion‐elicited social brain activation that was previously reported in people with ASD was correlated to their lack of eye fixation [Dalton et al., 2005], it is reasonable to hypothesize that the normalization of their social brain activation may be due to them fixating the eye region in this experiment, using the fixation cross as a placeholder. We attempted to place the fixation cross in the area of the face typically looked‐at in free‐viewing [Bindemann et al., 2009], so as to ensure a natural scanpath for participants.
A third limitation is its inclusion of participants with a wide age range. Although we have attempted to address this limitation by controlling for age in all analyses, it is possible that the developmental trajectory of facial emotion processing differs between people with and without ASD. However, although the processing of facial expression still matures between early adolescence and adulthood [Somerville et al., 2011] in typically developing participants, no study to date has investigated whether the maturing of emotion processing during this time window is affected by ASD. Future studies should consider investigating this critical research question.
Finally, we have contrasted emotional faces with neutral faces. This has been common practice in previous studies investigating emotion processing in the general population [see Fusar‐Poli et al., 2009 for a review] and in ASD [e.g., Critchley et al., 2000b]. This allows investigating brain activations specific to emotion processing without including activations associated with face processing, contrary to studies contrasting emotional faces with scrambled display [e.g., Ashwin et al., 2007]. Future studies investigating emotion processing in ASD should consider how a neutral baseline might affect their results.
To conclude, our results show that, when constrained to look at the eyes, ASD participants have increased social brain activations to low intensity fearful faces relative to controls. This hypersensitivity could reflect over‐arousal to distressing emotions and explain why people with ASD tend to avoid looing in the eyes. This over‐arousal could be due to hyperexcitable amygdala and to a faulty regulatory system in ASD, since we found that the vmPFC/amygdala coupling was absent in ASD participants viewing fearful faces. Overall, our results support the presence of an E/I imbalance in the socio‐affective processing system in ASD, where social deficits can be explained by an oversensitivity to distressing emotions, and a concomitant lack of emotional regulation. Social disengagement and avoidance of eye‐contact may be potential mechanisms to cope with this in daily life.
Supporting information
Supporting Information
ACKNOWLEDGMENT
The authors want to thank Ophélie Rogier for her help in data acquisition, Karine Métrailler and Carole Burget for their support in participant's recruitment and administrative assistance, and Anthony Lissot and Torsten Ruest for their help in data analysis. The funders had no role in the design and conduct of the study; collection, management, analysis, and interpretation of the data; and preparation, review, or approval of the manuscript. None of the authors has a conflict of interest to declare.
Footnotes
All ASD participants (N = 8) who did not have an ADOS and/or an ADI‐R were diagnosed using the DISCO.
The correlation results are presented without correction for multiple comparison. When the Bonferroni correction is applied, only the functional correlation between activity in the amygdala and the vmPFC for anger at high and low intensity in ASD participants remain significant.
REFERENCES
- Adolphs R, Tranel D, Damasio H, Damasio A (1994): Impaired recognition of emotion in facial expressions following bilateral damage to the human amygdala. Nature 372:669–672. [DOI] [PubMed] [Google Scholar]
- Adolphs R, Gosselin F, Buchanan TW, Tranel D, Schyns P, Damasio AR (2005): A mechanism for impaired fear recognition after amygdala damage. Nature 433:68–72. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association (2013): Diagnostic and statistical manual of mental disorders (5th ed.): DSM‐5. Arlington, VA: American Psychiatric Association. [Google Scholar]
- APA (2000) Diagnostic and Statistical Manual of Mental Disorders, DSM‐IV‐TR. APA. [DOI] [PubMed]
- Armony JL, Dolan RJ (2002): Modulation of spatial attention by fear‐conditioned stimuli: An event‐related fMRI study. Neuropsychologia 40:817–826. [DOI] [PubMed] [Google Scholar]
- Ashwin C, Chapman E, Colle L, Baron‐Cohen S (2006): Impaired recognition of negative basic emotions in autism: A test of the amygdala theory. Soc Neurosci 1:349–363. [DOI] [PubMed] [Google Scholar]
- Ashwin C, Baron‐Cohen S, Wheelwright S, O'Riordan M, Bullmore ET (2007): Differential activation of the amygdala and the 'social brain' during fearful face‐processing in Asperger Syndrome. Neuropsychologia 45:2–14. [DOI] [PubMed] [Google Scholar]
- Baron‐Cohen S, Ring HA, Wheelwright S, Bullmore ET, Brammer MJ, Simmons A, Williams SC (1999): Social intelligence in the normal and autistic brain: An fMRI study. Eur J Neurosci 11:1891–1898. [DOI] [PubMed] [Google Scholar]
- Bayless SJ, Glover M, Taylor MJ, Itier RJ (2011): Is it in the eyes? Dissociating the role of emotion and perceptual features of emotionally expressive faces in modulating orienting to eye gaze. Vis Cogn 19:483–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bindemann M, Scheepers C, Burton AM (2009): Viewpoint and center of gravity affect eye movements to human faces. J Vis 9:7 1–16. [DOI] [PubMed] [Google Scholar]
- Bird G, Catmur C, Silani G, Frith C, Frith U (2006): Attention does not modulate neural responses to social stimuli in autism spectrum disorders. NeuroImage 31:1614–1624. [DOI] [PubMed] [Google Scholar]
- Bookheimer SY, Wang AT, Scott A, Sigman M, Dapretto M (2008): Frontal contributions to face processing differences in autism: Evidence from fMRI of inverted face processing. J Int Neuropsychol Soc 14:922–932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brothers L (1990): The social brain: A project for integrating primate behaviour and neurophysiology in a new domain. Concepts Neurosci 1:27–151. [Google Scholar]
- Castelli F, Frith C, Happe F, Frith U (2002): Autism, Asperger syndrome and brain mechanisms for the attribution of mental states to animated shapes. Brain 125:1839–1849. [DOI] [PubMed] [Google Scholar]
- Critchley H, Daly E, Phillips M, Brammer M, Bullmore E, Williams S, Van Amelsvoort T, Robertson D, David A, Murphy D (2000a): Explicit and implicit neural mechanisms for processing of social information from facial expressions: A functional magnetic resonance imaging study. Hum Brain Mapp 9:93–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Critchley HD, Daly EM, Bullmore ET, Williams SC, Van Amelsvoort T, Robertson DM, Rowe A, Phillips M, McAlonan G, Howlin P, Murphy DG (2000b): The functional neuroanatomy of social behaviour: Changes in cerebral blood flow when people with autistic disorder process facial expressions. Brain 123:2203–2212. [DOI] [PubMed] [Google Scholar]
- Dalton KM, Nacewicz BM, Johnstone T, Schaefer HS, Gernsbacher MA, Goldsmith HH, Alexander AL, Davidson RJ (2005): Gaze fixation and the neural circuitry of face processing in autism. Nat Neurosci 8:519–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dapretto M, Davies MS, Pfeifer JH, Scott AA, Sigman M, Bookheimer SY, Iacoboni M (2006): Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders. Nat Neurosci 9:28–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dichter GS, Felder JN, Green SR, Rittenberg AM, Sasson NJ, Bodfish JW (2012): Reward circuitry function in autism spectrum disorders. Soc Cogn Affect Neurosci 7:160–172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engell AD, Haxby JV (2007): Facial expression and gaze‐direction in human superior temporal sulcus. Neuropsychologia 45:3234–3241. [DOI] [PubMed] [Google Scholar]
- Etkin A, Buchel C, Gross JJ (2015): The neural bases of emotion regulation. Nat Rev Neurosci 16:693–700. [DOI] [PubMed] [Google Scholar]
- Falck‐Ytter T, von Hofsten C (2011): How special is social looking in ASD: A review. Prog Brain Res 189:209–222. [DOI] [PubMed] [Google Scholar]
- Freyberg J, Robertson CE, Baron‐Cohen S (2015): Reduced perceptual exclusivity during object and grating rivalry in autism. J Vis 15:11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fusar‐Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S, Benedetti F, Abbamonte M, Gasparotti R, Barale F, Perez J, McGuire P, Politi P (2009): Functional atlas of emotional faces processing: a voxel‐based meta‐analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci 34:418–432. [PMC free article] [PubMed] [Google Scholar]
- Gernsbacher MA, Frymiare JL (2005): Does the Autistic Brain Lack Core Modules? J Dev Learn Disord 9:3–16. [PMC free article] [PubMed] [Google Scholar]
- Gotham K, Risi S, Pickles A, Lord C (2007): The Autism Diagnostic Observation Schedule: Revised algorithms for improved diagnostic validity. J Autism Dev Disord 37:613–627. [DOI] [PubMed] [Google Scholar]
- Grelotti DJ, Klin AJ, Gauthier I, Skudlarski P, Cohen DJ, Gore JC, Volkmar FR, Schultz RT (2005): fMRI activation of the fusiform gyrus and amygdala to cartoon characters but not to faces in a boy with autism. Neuropsychologia 43:373–385. [DOI] [PubMed] [Google Scholar]
- Hadjikhani N, Joseph RM, Snyder J, Chabris CF, Clark J, Steele S, McGrath L, Vangel M, Aharon I, Feczko E, Harris G, Tager‐Flusberg H (2004): Activation of the fusiform gyrus when individuals with autism spectrum disorder view faces. NeuroImage 22:1141–1150. [DOI] [PubMed] [Google Scholar]
- Hadjikhani N, Joseph RM, Snyder J, Tager‐Flusberg H (2007): Abnormal activation of the social brain during face perception in autism. Hum Brain Mapp 28:441–449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadjikhani N, Zürcher NR, Rogier O, Hippolyte L, Lemonnier E, Ruest T, Ward N, Lassalle A, Gillberg N, Billstedt E, Helles A, Gillberg C, Solomon P, Prkachin KM, Gillberg C (2014): Emotional contagion for pain is intact in autism spectrum disorders. Transl Psychiatry 4:e343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadjikhani N, Åsberg‐Johnels J, Zürcher NR, Lassalle A, Guillon Q, Hippolyte L, Billstedt E, Ward N, Lemonnier E, Gillberg C (2017a): Look me in the eyes: Constraining gaze in the eye‐region provokes abnormally high subcortical activation in autism. Sci Rep 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadjikhani N, Zürcher NR, Lassalle A, Hippolyte L, Ward N, Åsberg‐Johnels J (2017b): The effect of constraining eye‐contact during dynamic emotional face perception ‐ an fMRI study. Soc Cogn Affect Neurosci 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harms MB, Martin A, Wallace GL (2010): Facial emotion recognition in autism spectrum disorders: a review of behavioral and neuroimaging studies. Neuropsychol Rev 20:290–322. [DOI] [PubMed] [Google Scholar]
- Hobson, P. (1994) Understanding persons: The role of affect In: Baron‐Cohen S, Tager‐Flusberg H, Cohen D, editors. Understaning other minds. London: Oxford University Press. [Google Scholar]
- Jackson PL, Brunet E, Meltzoff AN, Decety J (2006): Empathy examined through the neural mechanisms involved in imagining how I feel versus how you feel pain. Neuropsychologia 44:752–761. [DOI] [PubMed] [Google Scholar]
- Kamachi M, Bruce V, Mukaida S, Gyoba J, Yoshikawa S, Akamatsu S (2013): Dynamic properties influence the perception of facial expressions. Perception 42:1266–1278. [DOI] [PubMed] [Google Scholar]
- Kanner L, Eisenberg L (1957): Early infantile autism, 1943–1955. Psychiatr Res Rep Am Psychiatr Assoc 55–65. [DOI] [PubMed] [Google Scholar]
- Kanwisher N, McDermott J, Chun MM (1997): The fusiform face area: A module in human extrastriate cortex specialized for face perception. J Neurosci 17:4302–4311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilts CD, Egan G, Gideon DA, Ely TD, Hoffman JM (2003): Dissociable neural pathways are involved in the recognition of emotion in static and dynamic facial expressions. NeuroImage 18:156–168. [DOI] [PubMed] [Google Scholar]
- Kim SY, Choi US, Park SY, Oh SH, Yoon HW, Koh YJ, Im WY, Park JI, Song DH, Cheon KA, Lee CU (2015): Abnormal activation of the social brain network in children with autism spectrum disorder: An FMRI study. Psychiatry Investig 12:37–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleinhans NM, Richards T, Sterling L, Stegbauer KC, Mahurin R, Johnson LC, Greenson J, Dawson G, Aylward E. (2008) Abnormal functional connectivity in autism spectrum disorders during face processing. Brain 131:1000–1012. [DOI] [PubMed] [Google Scholar]
- Kleinhans NM, Johnson LC, Richards T, Mahurin R, Greenson J, Dawson G, Aylward E (2009): Reduced Neural Habituation in the Amygdala and Social Impairments in Autism Spectrum Disorders. Am J Psychiatry 166:467–475. [DOI] [PubMed] [Google Scholar]
- Kliemann D, Dziobek I, Hatri A, Baudewig J, Heekeren HR (2012): The role of the amygdala in atypical gaze on emotional faces in autism spectrum disorders. J Neurosci 32:9469–9476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klin A, Jones W, Schultz R, Volkmar F, Cohen D (2002): Visual fixation patterns during viewing of naturalistic social situations as predictors of social competence in individuals with autism. Arch Gen Psychiatry 59:809–816. [DOI] [PubMed] [Google Scholar]
- Kylliainen A, Hietanen JK (2006): Skin conductance responses to another person's gaze in children with autism. J Autism Dev Disord 36:517–525. [DOI] [PubMed] [Google Scholar]
- Lamm C, Batson CD, Decety J (2007): The neural substrate of human empathy: Effects of perspective‐taking and cognitive appraisal. J Cogn Neurosci 19:42–58. [DOI] [PubMed] [Google Scholar]
- Lawrence EJ, Shaw P, Giampietro VP, Surguladze S, Brammer MJ, David AS (2006): The role of 'shared representations' in social perception and empathy: An fMRI study. Neuroimage 29:1173–1184. [DOI] [PubMed] [Google Scholar]
- Leekam SR, Libby SJ, Wing L, Gould J, Taylor C (2002): The Diagnostic Interview for Social and Communication Disorders: algorithms for ICD‐10 childhood autism and Wing and Gould autistic spectrum disorder. J Child Psychol Psychiatry 43:327–342. [DOI] [PubMed] [Google Scholar]
- Lord C, Rutter M, Le Couteur A (1994): Autism Diagnostic Interview‐Revised: A revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord 24:659–685. [DOI] [PubMed] [Google Scholar]
- Lord C, Risi S, Lambrecht L, Cook EH Jr., Leventhal BL, DiLavore PC, Pickles A, Rutter M (2000): The autism diagnostic observation schedule‐generic: A standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord 30:205–223. [PubMed] [Google Scholar]
- Mazefsky CA, Herrington J, Siegel M, Scarpa A, Maddox BB, Scahill L, White SW (2013): The role of emotion regulation in autism spectrum disorder. J Am Acad Child Adolesc Psychiatry 52:679–688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monk CS, Peltier SJ, Wiggins JL, Weng SJ, Carrasco M, Risi S, Lord C (2009): Abnormalities of intrinsic functional connectivity in autism spectrum disorders. Neuroimage 47:764–772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monk CS, Weng SJ, Wiggins JL, Kurapati N, Louro HM, Carrasco M, Maslowsky J, Risi S, Lord C (2010): Neural circuitry of emotional face processing in autism spectrum disorders. J Psychiatry Neurosci 35:105–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JP, Pelphrey KA, McCarthy G (2007): Controlled scanpath variation alters fusiform face activation. Soc Cogn Affect Neurosci 2:31–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelphrey KA, Sasson NJ, Reznick JS, Paul G, Goldman BD, Piven J (2002): Visual scanning of faces in autism. J Autism Dev Disord 32:249–261. [DOI] [PubMed] [Google Scholar]
- Perlman SB, Hudac CM, Pegors T, Minshew NJ, Pelphrey KA (2011): Experimental manipulation of face‐evoked activity in the fusiform gyrus of individuals with autism. Soc Neurosci 6:22–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Tancer ME (2006): Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia. Biol Psychiatry 59:424–429. [DOI] [PubMed] [Google Scholar]
- Pierce K, Muller RA, Ambrose J, Allen G, Courchesne E (2001): Face processing occurs outside the fusiform 'face area' in autism: Evidence from functional MRI. Brain 124:2059–2073. [DOI] [PubMed] [Google Scholar]
- Pierce K, Haist F, Sedaghat F, Courchesne E (2004): The brain response to personally familiar faces in autism: findings of fusiform activity and beyond. Brain 127:2703–2716. [DOI] [PubMed] [Google Scholar]
- Piggot J, Kwon H, Mobbs D, Blasey C, Lotspeich L, Menon V, Bookheimer S, Reiss AL (2004): Emotional attribution in high‐functioning individuals with autistic spectrum disorder: A functional imaging study. J Am Acad Child Adolesc Psychiatry 43:473–480. [DOI] [PubMed] [Google Scholar]
- Pinkham AE, Hopfinger JB, Pelphrey KA, Piven J, Penn DL (2008): Neural bases for impaired social cognition in schizophrenia and autism spectrum disorders. Schizophr Res 99:164–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puce A, Allison T, Bentin S, Gore JC, McCarthy G (1998): Temporal cortex activation in humans viewing eye and mouth movements. J Neurosci 18:2188–2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richey JA, Damiano CR, Sabatino A, Rittenberg A, Petty C, Bizzell J, Voyvodic J, Heller AS, Coffman MC, Smoski M, Davidson RJ, Dichter GS (2015): Neural Mechanisms of Emotion Regulation in Autism Spectrum Disorder. J Autism Dev Disord 45:3409–3423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson CE, Kravitz DJ, Freyberg J, Baron‐Cohen S, Baker CI (2013): Slower rate of binocular rivalry in autism. J Neurosci 33:16983–16991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson CE, Ratai EM, Kanwisher N (2016): Reduced GABAergic Action in the Autistic Brain. Curr Biol 26:80–85. [DOI] [PubMed] [Google Scholar]
- Rosenberg A, Patterson JS, Angelaki DE (2015): A computational perspective on autism. Proc Natl Acad Sci U S A 112:9158–9165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossignol M, Philippot P, Douilliez C, Crommelinck M, Campanella S (2005): The perception of fearful and happy facial expression is modulated by anxiety: an event‐related potential study. Neurosci Lett 377:115–120. [DOI] [PubMed] [Google Scholar]
- Ruby P, Decety J (2003): What you believe versus what you think they believe: A neuroimaging study of conceptual perspective‐taking. Eur J Neurosci 17:2475–2480. [DOI] [PubMed] [Google Scholar]
- Samson AC, Hardan AY, Lee IA, Phillips JM, Gross JJ (2015): Maladaptive Behavior in Autism Spectrum Disorder: The Role of Emotion Experience and Emotion Regulation. J Autism Dev Disord 45:3424–3432. [DOI] [PubMed] [Google Scholar]
- Sato W, Yoshikawa S, Kochiyama T, Matsumura M (2004): The amygdala processes the emotional significance of facial expressions: An fMRI investigation using the interaction between expression and face direction. Neuroimage 22:1006–1013. [DOI] [PubMed] [Google Scholar]
- Sato W, Toichi M, Uono S, Kochiyama T (2012): Impaired social brain network for processing dynamic facial expressions in autism spectrum disorders. BMC Neurosci 13:99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saxe R, Kanwisher N (2003): People thinking about thinking people. The role of the temporo‐parietal junction in “theory of mind”. Neuroimage 19:1835–1842. [DOI] [PubMed] [Google Scholar]
- Saxe R, Wexler A (2005): Making sense of another mind: the role of the right temporo‐parietal junction. Neuropsychologia 43:1391–1399. [DOI] [PubMed] [Google Scholar]
- Schultz RT, Gauthier I, Klin A, Fulbright RK, Anderson AW, Volkmar F, Skudlarski P, Lacadie C, Cohen DJ, Gore JC (2000): Abnormal ventral temporal cortical activity during face discrimination among individuals with autism and Asperger syndrome. Arch Gen Psychiatry 57:331–340. [DOI] [PubMed] [Google Scholar]
- Schyns PG, Petro LS, Smith ML (2007): Dynamics of visual information integration in the brain for categorizing facial expressions. Curr Biol 17:1580–1585. [DOI] [PubMed] [Google Scholar]
- Senju A (2013): Atypical development of spontaneous social cognition in autism spectrum disorders. Brain Dev 35:96–101. [DOI] [PubMed] [Google Scholar]
- Senju A, Johnson MH (2009): Atypical eye contact in autism: Models, mechanisms and development. Neurosci Biobehav Rev 33:1204–1214. [DOI] [PubMed] [Google Scholar]
- Shin LM, Liberzon I (2010): The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacology: Official publication of the American College of. Neuropsychopharmacology 35:169–191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shumway S, Farmer C, Thurm A, Joseph L, Black D, Golden C (2012): The ADOS calibrated severity score: relationship to phenotypic variables and stability over time. Autism Res 5:267–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegel JS, Power JD, Dubis JW, Vogel AC, Church JA, Schlaggar BL, Petersen SE (2014): Statistical improvements in functional magnetic resonance imaging analyses produced by censoring high‐motion data points. Hum Brain Mapp 35:1981–1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith A (2009): The Empathy Imbalance Hypothesis of Autism: A Theoretical Approach to Cognitive and Emotional Empathy in Autistic Development. Psychol Rec 59:489–510. [Google Scholar]
- Smith FW, Schyns PG (2009): Smile through your fear and sadness: Transmitting and identifying facial expression signals over. A range of viewing distances. Psychol Sci 20:1202–1208. [DOI] [PubMed] [Google Scholar]
- Somerville LH, Fani N, McClure‐Tone EB (2011): Behavioral and neural representation of emotional facial expressions across the lifespan. Dev Neuropsychol 36:408–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spezio ML, Adolphs R, Hurley RS, Piven J (2006): Analysis of face gaze in autism using “Bubbles”. Neuropsychologia. [DOI] [PubMed] [Google Scholar]
- Spielberger CD, Gorsuch RL, Lushene R, Vagg PR, Jacobs GA (1983): Manual for the State‐Trait Anxiety Inventory. Palo Alto, CA: Consulting Psychologists Press. [Google Scholar]
- Surguladze SA, Brammer MJ, Young AW, Andrew C, Travis MJ, Williams SC, Phillips ML (2003): A preferential increase in the extrastriate response to signals of danger. Neuroimage 19:1317–1328. [DOI] [PubMed] [Google Scholar]
- Tanaka JW, Sung A (2016): The “Eye Avoidance” Hypothesis of Autism Face Processing. J Autism Dev Disord 46:1538–1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tottenham N, Tanaka JW, Leon AC, McCarry T, Nurse M, Hare TA, Marcus DJ, Westerlund A, Casey BJ, Nelson C (2009): The NimStim set of facial expressions: Judgments from untrained research participants. Psychiatry Res 168:242–249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trautmann SA, Fehr T, Herrmann M (2009): Emotions in motion: dynamic compared to static facial expressions of disgust and happiness reveal more widespread emotion‐specific activations. Brain Res 1284:100–115. [DOI] [PubMed] [Google Scholar]
- van Steensel FJ, Bogels SM, Perrin S (2011): Anxiety disorders in children and adolescents with autistic spectrum disorders: A meta‐analysis. Clin Child Fam Psychol Rev 14:302–317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vuilleumier P, Pourtois G (2007): Distributed and interactive brain mechanisms during emotion face perception: Evidence from functional neuroimaging. Neuropsychologia 45:174–194. [DOI] [PubMed] [Google Scholar]
- Vuilleumier P, Armony JL, Driver J, Dolan RJ (2001): Effects of attention and emotion on face processing in the human brain: An event‐related fMRI study. Neuron 30:829–841. [DOI] [PubMed] [Google Scholar]
- Vuilleumier P, Richardson MP, Armony JL, Driver J, Dolan RJ (2004): Distant influences of amygdala lesion on visual cortical activation during emotional face processing. Nat Neurosci 7:1271–1278. [DOI] [PubMed] [Google Scholar]
- Wallace S, Coleman M, Bailey A (2008): An investigation of basic facial expression recognition in autism spectrum disorders. Cogn Emotion 22:1353–1380. [Google Scholar]
- Wang AT, Dapretto M, Hariri AR, Sigman M, Bookheimer SY (2004): Neural correlates of facial affect processing in children and adolescents with autism spectrum disorder. J Am Acad Child Adolesc Psychiatry 43:481–490. [DOI] [PubMed] [Google Scholar]
- Wechsler, D. (1999) Wechsler Abbreviated Scale of Intelligence (WASI). San Antonio, TX: Harcourt Assessment. [Google Scholar]
- Wechsler, D. , Naglieri, J. (2006) Wechsler Nonverbal Scale of Ability. San Antonio, TX: PsychCorp Edition, A Brand of Harcourt Assessment. [Google Scholar]
- Whalen PJ, Kagan J, Cook RG, Davis FC, Kim H, Polis S, McLaren DG, Somerville LH, McLean AA, Maxwell JS, Johnstone T (2004): Human amygdala responsivity to masked fearful eye whites. Science 306:2061–2061. [DOI] [PubMed] [Google Scholar]
- Wicker B, Fonlupt P, Hubert B, Tardif C, Gepner B, Deruelle C (2008): Abnormal cerebral effective connectivity during explicit emotional processing in adults with autism spectrum disorder. Soc Cogn Affect Neurosci 3:135–143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zürcher NR, Donnelly N, Rogier O, Russo B, Hippolyte L, Hadwin J, Lemonnier E, Hadjikhani N (2013a): It's all in the eyes: Subcortical and cortical activation during grotesqueness perception in autism. PloS One 8:e54313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zürcher NR, Rogier O, Boshyan J, Hippolyte L, Russo B, Gillberg N, Helles A, Ruest T, Lemonnier E, Gillberg C, Hadjikhani N (2013b): Perception of social cues of danger in autism spectrum disorders. PloS One 8:e81206. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Information