

Abstract
Objectives
The anatomical organization of the insular cortex is characterized by its rich and heterogeneous cytoarchitecture and its wide network of connections. However, only limited knowledge is available regarding the intrainsular connections subserving the complex integrative role of the insular cortex. The aim of this study was to analyze the functional connectivity within‐ and across‐insular subregions, at both gyral and functional levels.
Experimental design
We performed intracerebral electrical stimulation in 10 patients with refractory epilepsy investigated with depth electrodes, 38 of which were inserted in the insula. Bipolar electrical stimulation, consisting of two series of 20 pulses of 1‐ms duration, 0.2‐Hz frequency, and 1‐mA intensity, was delivered at each insular contact. For each stimulated insular anatomical region, we calculated a rate of connectivity, reflecting the proportion of other insular contacts, showing significant evoked potentials.
Results
Statistically significant evoked potentials were recorded in 74% of tested connections, with an average latency of 26 ± 3 ms. All insular gyri were interconnected, except the anterior and posterior short gyri. Most connections were reciprocal, showing no clear anterior to posterior directionality. No connection was observed between the right and the left insula.
Conclusions
These findings point to specific features of human insula connectivity as compared to non‐Human primates, and remain consistent with the complex integration role devoted to the human insula in many cognitive domains. Hum Brain Mapp 35:2779–2788, 2014. © 2013 Wiley Periodicals, Inc.
Keywords: insular, functional connectivity, intra‐cranial electrical stimulation, evoked potential, human
INTRODUCTION
The insula is a functionally complex region providing multimodal integration and involved in various verbal and nonverbal activities, including speech motor control [Bohland and Guenther, 2006], emotional processing [Adolphs et al., 2003; Calder et al., 2000; Wicker et al., 2003], pain perception [Alkire et al., 2004; Mazzola et al., 2006; Ostrowsky et al., 2002], somatosensory integration [Burton et al., 1993], auditory processing [Bieser, 1998], as well as taste and olfactory perceptions [Kurth et al., 2010; Small, 2010; Stephani et al., 2011; Yaxley et al.,1990].
The anatomical organization of the insular cortex is characterized by its rich and heterogeneous cytoarchitecture [Mesulam and Mufson, 1985] and its wide network of connections [Augustine, 1996]. However, only limited knowledge is available regarding the intrainsular connections subserving the complex integrative role of the insular cortex. The nonhuman primate insula demonstrates abundant bidirectional intrainsular connections, with stronger connectivity from anterior to posterior regions [Mesulam and Mufson, 1982b; Seltzer and Pandya, 1991], providing a potential pathway for conveying olfactory and gustatory information into the posterior insula [Mesulam and Mufson, 1982b]. In human, a single in vivo probabilistic tractography has suggested strong within‐subregion structural connectivity in both anterior and posterior insular regions [Cloutman et al., 2011].
Human cerebral functional connectivity can be studied in patients with refractory epilepsy undergoing intracerebral EEG (icEEG) investigation, by applying electrical stimulation to cortical areas and recording evoked potentials (EPs) in distant‐connected structures [Brazier, 1964; Buser and Bancaud, 1983; Buser et al., 1992; Catenoix et al., 2005, 2011; Lacruz et al., 2007; Matsumoto et al., 2004, 2007; Rosenberg et al., 2009; Rutecki et al., 1989; Wilson et al., 1990]. This approach offers a high level of temporal and spatial resolution for mapping brain connectivity, providing complementary findings to those depicted by currently available neuroimaging methods. Taking advantage of clinical icEEG investigations of the insula, we applied this method to analyze the functional connectivity within‐ and across‐insular subregions, at both gyral and functional levels.
MATERIALS AND METHODS
Patients
Ten patients with drug‐resistant partial epilepsy contemplating epilepsy surgery and undergoing icEEG were included in this study. Inclusion criteria included the presence of at least two different electrodes inserted within the same insula to look at EP in at least one insular electrode while stimulating another. Exclusion criteria included the presence of any morphological abnormality of the insula. All patients gave their informed consent to participate in this study.
Stereotaxic Implantation of Depth Electrodes
IcEEG was performed according to the technique described by Talairach and Bancaud [1973], a procedure used routinely in our department [Guenot et al., 2001]. The brain regions to be investigated were determined for each patient, based on individual presurgical data, and most likely origin of seizure onset. In general, about one‐third of the electrodes target the most likely zone of seizure onset, whereas the remaining electrodes probe alternative but less likely hypothesis, or help to define the borders of the epileptogenic zone. As a result, a significant number of recording contacts are not affected by the epileptic process. This is particularly true for the insula, which is often investigated owing to its potential to mimic temporal or frontal lobe seizures [Isnard et al., 2001, 2004, Ryvlin et al., 2006], but eventually found not to be affected in the majority of cases.
Electrodes were implanted perpendicular to the mid‐sagittal plane with the patient's head fixed in the Talairach's stereotactic frame, providing Talairach's coordinates for each electrode in relation to the anterior commissure/posterior commissure plane. The exact location of each electrode and recording lead was further verified with a postimplantation MRI. The two deepest contacts of the electrodes targeting the insula were located within that structure, whereas the next more lateral contact was located either in the sylvian fissure or within the deepest portion of the adjacent operculum (temporal, frontal, or parietal). For the purpose of illustrating our findings on a single template, we used the above information to position the insular leads inserted in our 10 patients onto a T1‐MRI sagittal slice of the insula derived from a normal subject.
In total, 11–16 semi‐rigid intracerebral electrodes were implanted per patient, either unilaterally (n = 8) or bilaterally (n = 2). Each electrode was 0.8 mm in diameter and included 5, 12, or 15 contacts 2 mm in length, 1.5 mm apart (Dixi, Besançon, France), depending on the target region. No morbidity was related to cerebral electrode implantation.
Brain Stimulation and EP Recordings
Intracerebral EP recordings were performed using a video‐EEG monitoring system (Micromed, Treviso, Italy) that allowed to simultaneously record up to 128 contacts at a sampling rate of 1,024 Hz. As part of the clinical icEEG investigation, electrical brain stimulation is systematically performed to assess the epileptogenicity and functionality of the implanted brain regions. This procedure includes 50, 1, and 0.2 Hz stimulations, with 50 and 1 Hz, aiming at triggering ictal signs, epileptic discharge, or full‐blown seizure [Kahane et al., 1993, 2004; Munari et al., 1993], whereas 0.2 Hz are used to trigger abnormal cortical‐evoked responses (i.e., delayed to more than 100 ms or repetitive) suggestive of an underlying epileptogenic cortex [Valentin et al., 2002; 2005a,2005b; van't Klooster et al., 2011]. In addition to their clinical utility, 0.2‐Hz stimulations also allow to measure physiological early responses, referred to as EPs in this study, and reflecting brain connectivity [Brazier, 1964; Buser and Bancaud, 1983; Buser et al., 1992; Catenoix et al., 2005, 2011; Lacruz et al., 2007; Matsumoto et al., 2004, 2007; Rosenberg et al., 2009; Rutecki et al., 1989; Wilson et al., 1990].
We performed 0.2‐Hz stimulations at least 4 days after electrodes implantation, once the patient has fully recovered from the surgical procedure. We used bipolar stimulation of adjacent contacts from the same electrode, known to deliver current within 5 mm around the stimulated bipole [Nathan et al., 1993]. Electrical stimulation was delivered using a current‐regulated neurostimulator (Micromed, Treviso, Italy), with parameters ensuring patient's safety and effective generation of EPs [Catenoix et al., 2005, 2011; Gordon et al., 1990]. We used monophasic pulse of 1 ms width and 1 mA intensity, resulting in an electrical charge of 1 μC. The latter was delivered over a contact surface of 0.05 cm2 (0.8 mm diameter × 2 mm length × π), resulting in a total charge density of 20 μC/cm2/phase, thus significantly lower than the maximum safe value of 60 μC/cm2/phase [Gordon et al., 1990]. Two consecutive series of 20 pulses were delivered at each pair of contacts.
Data Analysis
EEG data were analyzed with the software package for electrophysiological analysis (ELAN‐pack) developed at the DYCOG laboratory of Lyon Neuroscience Research Centre (CRNL, Lyon, France) [Aguera et al., 2011]. We first performed an automatic detection of the pulse artifact generated on the stimulated contacts, and systematically verified the accuracy of the generated marker. Thanks to the very reproducible shape and amplitude of artifacts, as well as the associated high signal‐to‐noise ratio, this procedure did not suffer from false‐positive or ‐negative detection. We used the stimulation marker for averaging each block of 20 consecutive pulse stimulations, and then calculated grand averages from the two blocks. Both visual and statistical analyses were used to conclude on the presence of significant EPs over each recording contact. EPs were first selected on the basis of visual analysis if detected and found comparable in each of the two consecutive series. Statistical analysis of the selected EPs was then performed using the nonparametric statistical function of ELAN‐pack for single trails (Wilcoxon test), with significance set at P 0.001. In brief, this statistical analysis compared each time point of the poststimulation period to the 1,000‐ms prestimulation baseline, providing a curve, superimposed on the EP, illustrating the P‐value associated with each component of the EP. The first 10‐ms poststimulations were not evaluated owing to the presence of residual stimulation‐induced artifact. EPs were considered significant when they reached the statistical threshold of P ≤ 0.001 during at least five consecutive milliseconds. The latency of the first peak of each significant potential was measured on the grand average of the two series.
Insular regions targeted by intracerebral electrodes were divided into those involved at seizure onset, those invaded during seizure propagation, and those not affected by the epileptic discharge.
For each stimulated insular anatomical region, we calculated a rate of connectivity, reflecting the proportion of other insular contacts showing significant EPs. This analysis was performed at the gyral level, considering the three short and two long gyri as five distinct anatomical regions, and at a functional level, distinguishing the social–emotional, cognitive, chemical sensory, and sensorimotor insular subregions as identified in a recent functional neuroimaging meta‐analysis of the human insula [Kurth et al., 2010].
RESULTS
A total number of 38 electrodes were implanted into 12 insulae (two patients had bilateral implantation), with 29 electrodes placed in the left insula (76%) and 9 in the right (24%). The gyral location of these electrodes was as follows: three electrodes in the anterior short gyrus (ASG), four in the middle short gyrus (MSG), seven in the posterior short gyrus (PSG), 10 in the anterior long gyrus (ALG), and 14 in the posterior long gyrus (PLG). In terms of functional areas, three electrodes sampled the cognitive, four the chemical sensory, and 26 the sensorimotor subregions of the insula, whereas no electrode was placed in its social emotional portion (five other electrodes fell out of the functional areas determined by Kurth et al., 2010).
Stimulations were performed at all 38 insular electrodes, giving rise to significant EPs on other insular contacts in 74% of the 110 tested connections. Recorded EPs typically corresponded to biphasic early responses, with an average ± SD latency of the first peak of 26 ± 3 ms (range, 20–31 ms) (Fig. 1).
Figure 1.
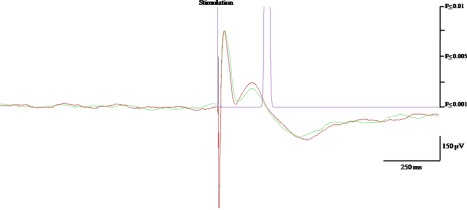
Illustration of EP recorded after insular stimulation. Negative polarity is upward. The green and red color superimposed curves are the average of two 20 trials, showing similar N1 and N2 peaks. The purple curve represents the P‐statistic whose value is 0.001 threshold (i.e., significant response) when reaching the abscissa.
The epileptogenic zone was located in the temporal lobe in five patients, frontal lobe in three, parietal lobe in one, and in the insula in one. In this patient, four out of the six insular electrodes were involved at seizure onset, whereas the other two were affected only during seizure propagation. Another two patients with a temporal or a frontal epileptogenic zone had one of their insular electrodes affected by seizure propagation. The proportion, morphology, and latency of EPs triggered or recorded at each insular contact were comparable for those involved at seizure onset, those affected during seizure propagation, and those not affected by the epileptic discharge. Thus, further results pooled data triggered and recorded at all insular contacts.
Gyral Connectivity
Connectivity between the different insular gyri is shown in Table 1 and Figures 2 and 3. The ASG demonstrated limited connectivity (30%, N = 20), with most of its connections targeting the adjacent MSG and being unidirectional from anterior to posterior. The MSG showed a 67% rate of connectivity (N = 24), distributed over all other gyri, with most connections being reciprocal. The PSG showed the highest rate of connectivity (88%, N = 40), primarily targeting the MSG and the two long gyri with reciprocal connections. ALG showed a 71% rate of connectivity (N = 52), mostly with the adjacent PSG and PLG. All connections with PSG were reciprocal, whereas nearly half of those with PLG were unidirectional. Among the latter, half were anterior to posterior, whereas the other half were posterior to anterior. PLG showed a 75% rate of connectivity (N = 60), which qualitative pattern was similar to that described for ALG.
Table 1.
Connectivity across and within the five insular gyri
| Recorded leads | ||||||
|---|---|---|---|---|---|---|
| Anterior short gyrus | Middle short gyrus | Posterior short gyrus | Anterior long gyrus | Posterior long gyrus | ||
| Stimulated leads | Anterior short gyrus | – | 100% (N = 2) 24 ± 7 | 0 (N = 2) | 33% (N = 3) 21 ± 1 | 33% (N = 3) 28 ± 1 |
| Middle short gyrus | 50% (N = 2) 24 ± 5 | – | 100% (N = 2) 30 ± 1 | 50% (N = 4) 26 ± 5 | 75% (N = 4) 34 ± 1 | |
| Posterior short gyrus | 0 (N = 2) | 100% (N = 2) 29 ± 4 | 100% (N = 4) 30 ± 4 | 50% (N = 6) 25 ± 3 | 75% (N = 10) 28 ± 4 | |
| Anterior long gyrus | 0 (N = 3) | 50% (N‐4) 26 ± 1 | 100% (N‐6) 27 ± 5 | – | 77% (N‐13) 27 ± 2 | |
| Posterior long gyrus | 0 (N = 3) | 50% (N = 4) 27 ± 5 | 90% (N = 10) 28 ± 3 | 77% (N = 13) 26 ± 5 | 100% (N = 8) 28 ± 2 | |
For each tested connection: 1) top number respresents connectivity rate (responding connections/total tested connections); 2) middle number in brackets (N) is the number of connections tested; 3) bottom number is the mean latency in ms ± SD.
Empty cells: no tested connection.
Figure 2.
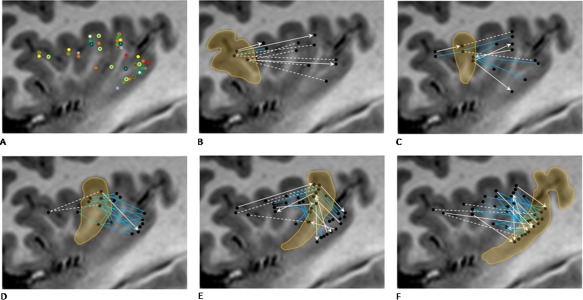
Electrode location and gyral connectivity pattern. A: Electrode location in all 10 patients; different color is used for each patient. B: ASG connectivity. C: MSG connectivity. D: PSG connectivity. E: ALG connectivity. F: PLG connectivity. For graphs B–F, gyrus of interest is encircled and highlighted, blue solid lines indicate bidirectional connectivity, white solid arrows indicate unidirectional connectivity, and dotted white lines indicate lack of detectable connection.
Figure 3.
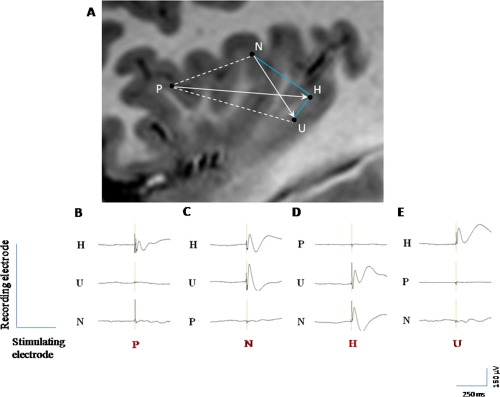
Illustration of the various patterns of connectivity in one patient. A: Connectivity between four insular leads located within the ASG (P), PSG (N), and PLG (H, U). Blue solid lines indicate bidirectional connectivity, white solid arrows indicate unidirectional connectivity, and dotted white lines indicate lack of detectable connection. (B–E) EPs evoked by stimulating P, N, H, and U leads, respectively. B: the stimulation of P (ASG) generate EPs only over H (PLG), which are not reciprocal; (C) the stimulation of N (PSG) generate EPs over H and U (PLG), only one of which is reciprocal (H); (D) the stimulation of U (PLG) generate EPs only over H, which are reciprocal; and (E) the stimulation of H (PLG) generate EPs over N (PSG), and U (PLG), both of which are reciprocal.
Latencies of the earliest EP varied from 21 ± 1 ms for connections between the anterior short and long gyri, to 34 ± 1ms for connections between the MSG and the PLG (Table 1).
Intragyral connectivity could be tested only across a limited number of connections within the PSG (N = 4) and the PLG (N = 8), showing 100% connectivity rate in these two gyri.
Functional Connectivity
Connectivity between the different functional areas of the insula is shown in Table 2 and Figure 4. The cognitive subregion had a low rate of unidirectional anterior to posterior connectivity with sensorimotor areas (29%) (N = 7). In contrast, the chemical sensory and sensorimotor areas demonstrated a 63% rate of reciprocal connectivity (N = 8). Only two connections could be tested between the cognitive and the chemical sensory areas, showing bidirectional connectivity. Connectivity within the same functional subregion could be assessed only within the sensorimotor area where 93% of connections proved functional (N = 44).
Table 2.
Connectivity across and within main insular functional subregions
| Recorded leads | ||||
|---|---|---|---|---|
| Cognitive | Chemical sensory | Sensory‐motor | ||
| Stimulated leads | Cognitive | – | 100% (N = 2) 25 ± 3 | 29% (N = 7) 27 ± 4 |
| Chemical sensory | 50% (N = 2) 26 ± 2 | – | 63% (N = 8) 28 ± 5 | |
| Sensory‐motor | 0 (N = 7) | 63% (N = 8) 28 ± 2 | 93% (N = 44) 27 ± 3 | |
For each tested connection: 1) top number respresents connectivity rate (responding connections/total tested connections); 2) middle number in brackets (N) is the number of connections tested; 3) bottom number is the mean latency in ms ± SD.
Empty cells: no tested connection.
Figure 4.
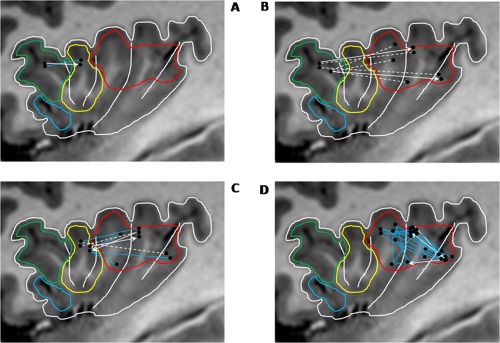
Connectivity across‐ and within‐main insular functional subregions. A: Connectivity between cognitive and chemical sensory regions. B: Connectivity between cognitive and sensorimotor regions. C: Connectivity between chemical sensory and sensorimotor regions. D: Connectivity within sensorimotor region. Functional regions as delineated by Kurth et al. [2010]: social–emotional (blue), cognitive (green), chemical sensory (yellow), and sensorimotor (red). For graphs (A–D), blue lines indicate bidirectional connectivity, white arrows indicate unidirectional connectivity, and dotted white lines indicate lack of detectable connection.
Latencies of the earliest EP were 25 ± 3 ms for connections between the cognitive and the chemical sensory regions, 28 ± 5ms for connections between the chemical sensory and the sensorimotor regions, and 27 ± 3ms for connections within the sensorimotor area (Table 2).
Contralateral Connectivity
Two patients had bilateral insular implantation, both of whom had three electrodes on one side and a single electrode on the opposite side. In one patient, contralateral electrode had an exact counterpart in the opposite insula, located in the same portion of the PLG. However, no EP could be recorded over the 12 potential interhemispheric connections tested in these patients.
DISCUSSION
This study provides the first human electrophysiological data exploring intrainsular functional connectivity, offering clues to the complex integrative role of the insular cortex.
Our knowledge of intrainsular connectivity primarily relies on data from nonhuman primates [Mesulam and Mufson, 1982b; Seltzer and Pandya, 1991]. The relevance of these to human insular connectivity is suggested by the similarities observed in the gyration, cytoarchitectony, and functional anatomy of the insula between the two species, leading some authors to suggest that the human insula has a plan of organization virtually identical to that of the rhesus monkey [Mesulam and Mufson, 1982a]. Both display an anterior–posterior pattern evolving from agranular to granular cortices through a transitional dysgranular area and a similar pattern of connectivity with extrainsular regions, primarily governed by the cytoarchitectonic features of the connected brain regions [Cerliani et al., 2011]. In line with the above observations, a recent in vivo tractography study reported an anterior–posterior connectivity predominating over the transitional area of the human insula, particularly between the PSG and the ALG [Cloutman et al., 2011]. However, whether intrainsular functional connectivity overlaps with these structural data remained to be determined. So far, only indirect evidence of intrainsular functional connectivity in human is available, based on the pattern of insular activation observed in a meta‐analysis of 1,768 functional neuroimaging experiments [Kurth et al., 2010].
Our study used direct electrical stimulation of the human insula and associated EPs to assess intrainsular functional connectivity. This method has been used to study the connectivity of various brain regions such as the mesial temporal structures [Brazier, 1964; Buser and Bancaud, 1983; Catenoix et al., 2005, 2011; Lacruz et al., 2007; Rutecki et al., 1989; Wilson et al., 1990], temporal neocortex including language areas [Matsumoto et al., 2004], frontal cortex [Buser et al., 1992; Lacruz et al., 2007], motor system [Matsumoto et al., 2007], and thalamic medial pulvinar nucleus [Rosenberg et al., 2009]. Based on this method, we showed that the human insula was characterized by rich connections between various insular gyri which can be summarized as follows: (1) all gyri were interconnected, except the ASG and PSG for which limited sample size hampers any firm conclusion; (2) most intrainsular connections proved reciprocal, in contrast with the anterior–posterior preferential direction described in the macaque monkey [Mesulam et al., 1982b; Seltzer and Pandya, 1991].
This latter finding raises the possibility that the human insula might have specific functional characteristics, but could also reflect differences between structural and functional methods on the one hand, and nonhuman and human studies on the other hand. Indeed, divergences between human neuroimaging functional and structural connectivity data have been reported in other brain regions [Damoiseaux and Greicius, 2009; Zhang et al., 2010]. The anterograde transport methods used in monkeys have offered the possibility to clearly determine the directionality of connections within the insular cortex of nonhuman primates [Mesulam et al., 1982b], an information not assessable with in vivo tractography in human. Whether connection directionality can be reliably tested by intracerebral electrical stimulation studies remains disputed. Indeed, such stimulation is thought to primarily trigger action potentials within bypassing axons with both orthodromic and antidromic propagation [Matsumoto et al., 2004; Wilson et al., 1990; Zhang and Oppenheimer, 2000]. Although this mechanism might well occur, it remains that 14% of the connections observed in our study were unidirectional, demonstrating that bidirectional stimulation of the same axons is not a generalized feature of corticocortical EPs. Furthermore, some insular contacts showed bidirectional connectivity with one insular region and unidirectional connectivity with another (Fig. 3). Finally, latencies of reciprocal EPs usually differ between the two directions, supporting the view that distinct neurons are stimulated to produce EPs of varying latency.
Although our data suggest a more reciprocal intrainsular connectivity pattern than that described in monkeys, some anterior‐to‐posterior preferential direction was observed for connections in the ASG and cognitive subregion. However, only a few electrodes were available in those regions, with very low connectivity rates, hampering any robust conclusion regarding this finding.
The morphology of EPs recorded in our series was consistent with those observed in the above studies. Latencies of the first detectable peak were consistent with the average values found in the previous studies, typically ranging between 20 and 30 ms [Matsumoto et al., 2004, 2007]. However, 10–20 ms latencies were reported for short‐distance EPs recorded within the motor and language brain regions [Matsumoto et al., 2004, 2007], in line with the observation that EP latency increases with the distance between the stimulating and the recording electrodes [Matsumoto et al., 2012]. Thus, latencies of intrainsular EPs could be considered excessive according to the relatively short distance of the tested connections. One possibility could be that we failed at detecting earlier EPs occurring during the first 10‐ms poststimulation owing to stimulus‐induced artifact. Another hypothesis would be that different from the motor and language networks that require fast processing monosynaptic pathways, the more integrative role of the insula is subtended by slower polysynaptic connections.
This hypothesis is consistent with the greater connectivity rate observed within the insula as compared to other brain regions assessed using the same methodology [Lacruz et al., 2007; Wilson et al., 1990], as well as with the dense connections observed between the insular cortex and many cortical and subcortical regions [Brooks et al., 2005; Büttner‐Ennever and Gerrits, 2004; Mesulam and Mufson, 1985; Pritchard and Norgren, 2004]. The human insula is thought to ensure a complex integration role in many cognitive domains such as emotional processing [Adolphs et al., 2003; Calder et al., 2000; Wicker et al., 2003], somatosensory and pain integration [Alkire et al, 2004; Burton et al., 1993; Ostrowsky et al., 2002; Mazzola et al., 2006], taste and olfaction [Kurth et al., 2010; Small, 2010; Stephani et al., 2011; Yaxley et al., 1990], and auditory and speech processing [Bieser, 1998; Bohland and Guenther, 2006]. Specifically, we observed dense functional connectivity within the sensorimotor portion of the insula, a region thought to provide a primary interoceptive representation of the physiological condition of the body which is then associated with input from multiple other sources in the mid‐insula [Craig, 2010]. Rich bidirectional connectivity between cortical areas that receive sensory afferents from different sources might underlie the build‐up of complex representation of sensory environment.
On the other hand, the low connectivity pattern observed between the ASG or cognitive subregion of the insula and the other insular regions is surprising. Indeed, it has been proposed that multimodal sensory information converge to the most anterior portion of the insula in a way that would contribute to self‐recognition and emotional awareness [Craig, 2009]. Conversely, other authors view that the anterior insula as an anatomically and functionally distinct structure from the reminder of the insula [Nelson et al., 2010]. In any event, our study suffers from clear undersampling of the most anterior portion of the insula, and further data will be needed to conclude on this issue.
Some other limitations need to be acknowledged. First, icEEG‐based studies of functional connectivity are necessarily performed in patients with epilepsy whose brain connections might be altered, especially within the epileptogenic zone. In fact, no difference between the EPs recorded from contacts included in the epileptic network and those recorded from nonepileptic tissue was observed as described previously by others [Lacruz et al., 2007; Wilson et al., 1990]. Furthermore, the insula was not part of the epileptogenic zone in nine out of our 10 patients, with the majority of tested contacts not even affected by the epileptic discharge during its propagation. Thus, we believe that our main findings are likely to apply to healthy individuals. A second issue relates to the sampling limitations of electrodes implanted orthogonally in the insula, with vascular constraints hampering access to the ventral portion of the anterior insula, leaving this region unexplored by our study. In addition, the clinical profiles of our patients resulted in greater need for exploring the PSG and PLG rather than the anterior and MSG, leading to less robust findings for these latter structures.
We failed to detect contralateral EPs to insular stimulation although we could test 12 potential connections in two patients, including one pair of electrodes placed in a very homologous position within both insulae. This negative finding is in contrast with the evidence of structural and functional connections between the two insulae in human [Anderson et al., 2011; Ebisch et al., 2010]. Although most likely reflecting insufficient insular sampling, our findings also suggest that the density of connections between the two insulae is much lower than that observed within each insula.
Overall, electrically induced corticocortical EPs demonstrate that the human insula is characterized by rich reciprocal connections within and between its mid and posterior aspects, in particular throughout the regions underlying sensorimotor integration. Further studies should confirm these findings in larger population and look at their relationship to the insular afferents and efferents as delineated by the same electrophysiological method.
REFERENCES
- Adolphs R, Tranel D, Damasio AR (2003): Dissociable neural systems for recognizing emotions. Brain Cogn 52:61–69. [DOI] [PubMed] [Google Scholar]
- Aguera PE, Jerbi K, Caclin A, Bertrand O (2011): ELAN: A software package for analysis and visualization of MEG, EEG, and LFP signals. Comput Intell Neurosci 2011:158970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alkire MT, White NS, Hsieh R, Haier RJ (2004): Dissociable brain activation responses to 5‐Hz electrical pain stimulation: A high‐field functional magnetic resonance imaging study. Anesthesiology 100:939–946. [DOI] [PubMed] [Google Scholar]
- Anderson JS, Druzgal TJ, Froehlich A, DuBray MB, Lange N, Alexander AL, Abildskov T, Nielsen JA, Cariello AN, Cooperrider JR, Bigler ED, Lainhart JE (2010): Decreased interhemispheric functional connectivity in autism. Cereb Cortex 21:1134–1146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Augustine JR. (1996): Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res Rev 22:229–244. [DOI] [PubMed] [Google Scholar]
- Bieser, A (1998): Processing of twitter‐call fundamental frequencies in insula and auditory cortex of squirrel monkeys. Exp Brain Res 122:139–148. [DOI] [PubMed] [Google Scholar]
- Bohland JW, Guenther FH (2006): An fMRI investigation of syllable sequence production. Neuroimage 32:821–841. [DOI] [PubMed] [Google Scholar]
- Brazier MA (1964): Evoked responses recorded from the depths of the human brain. Ann N Y Acad Sci 112:33–59. [DOI] [PubMed] [Google Scholar]
- Brooks JC, Zambreanu L, Godinez A, Craig AD, Tracey I (2005): Somatotopic organisation of the human insula to painful heat studied with high resolution functional imaging. Neuroimage 27:201–209. [DOI] [PubMed] [Google Scholar]
- Burton H, Videen TO, Raichle ME (1993): Tactile‐vibration‐activated foci in insular and parietal‐opercular cortex studied with positron emission tomography: Mapping the second somatosensory area in humans. Somatosen Mot Res 10:297–308. [DOI] [PubMed] [Google Scholar]
- Buser P, Bancaud J (1983): Unilateral connections between amygdala and hippocampus in man. A study of epileptic patients with depth electrodes. Electroencephalogr Clin Neurophysiol 55:1–12. [DOI] [PubMed] [Google Scholar]
- Buser P, Bancaud J, Chauvel P (1992): Callosal transfer between mesial frontal areas in frontal lobe epilepsies. Adv Neurol 57:589–604. [PubMed] [Google Scholar]
- Büttner‐Ennever JA, Gerrits NM (2004): Vestibular system In Paxinos G, Mai JK, editors. The Human Nervous System. Amsterdam: Elsevier; pp 1212–1240. [Google Scholar]
- Calder AJ, Keane J, Manes F, Antoun N, Young AW (2000): Impaired recognition and experience of disgust following brain injury. Nat Neurosci 3:1077–1078. [DOI] [PubMed] [Google Scholar]
- Catenoix H, Magnin M, Guenot M, Isnard J, Mauguiere F, Ryvlin P (2005): Hippocampal‐orbitofrontal connectivity in human: An electrical stimulation study. Clin Neurophysiol 116:1779–1784. [DOI] [PubMed] [Google Scholar]
- Catenoix H, Magnin M, Mauguiere F, Ryvlin P (2011): Evoked potential study of hippocampal efferent projections in the human brain. Clin Neurophysiol 122:2488–2497. [DOI] [PubMed] [Google Scholar]
- Cerliani L, Thomas RM, Jbabdi S, Siero JC, Nanetti L, Crippa A, Gazzola V, D'Arceuil H, Keysers C (2011): Probabilistic tractography recovers a rostrocaudal trajectory of connectivity variability in the human insular cortex. Hum Brain Mapp 33:2005–2034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cloutman LL, Binney RJ, Drakesmith M, Parker GJ, Ralph MA (2011): The variation of function across the human insula mirrors its patterns of structural connectivity: Evidence from in vivo probabilistic tractography. NeuroImage 59:3514–3521. [DOI] [PubMed] [Google Scholar]
- Craig AD (2009): How do you feel—Now? The anterior insula and human awareness. Nat Rev Neurosci. 10:59–70. [DOI] [PubMed] [Google Scholar]
- Craig AD (2010): The sentient self. Brain Struct Funct 214:563–577. [DOI] [PubMed] [Google Scholar]
- Damoiseaux JS, Greicius MD (2009): Greater than the sum of its parts: A review of studies combining structural connectivity and resting‐state functional connectivity. Brain Struct Funct 213:525–533. [DOI] [PubMed] [Google Scholar]
- Ebisch SJH, Gallese V, Willems RM, Mantini D, Groen WB, Romani GL, Buitelaar JK, Bekkering H (2010): Altered intrinsic functional connectivity of anterior and posterior insula regions in high‐functioning participants with autism spectrum disorder. Hum Brain Mapp 32:1013–1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon B, Lesser RP, Rance NE, Hart J Jr, Webber R, Uematsu S, Fisher RS (1990): Parameters for direct cortical electrical stimulation in the human: Histopathologic confirmation. Electroencephalogr Clin Neurophysiol 75:371–377. [DOI] [PubMed] [Google Scholar]
- Guenot M, Isnard J, Ryvlin P, Fischer C, Ostrowsky K, Mauguiere F, Sindou M (2001): Neurophysiological monitoring for epilepsy surgery: The Talairach SEEG method. Stereotact Funct Neurosurg 77:29–32. [DOI] [PubMed] [Google Scholar]
- Isnard J, Guenot M, Ostrowsky K, Sindou M, Mauguiere F (2001): The role of the insular cortex in temporal lobe epilepsy. Ann Neurol 48:614–623. [PubMed] [Google Scholar]
- Isnard J, Guenot M, Sindou M, Mauguiere F (2004): Clinical manifestations of insular lobe seizures: A stereo‐electroencephalographic study. Epilepsia 45:1079–1090. [DOI] [PubMed] [Google Scholar]
- Kahane P, Tassi L, Francionne S, Hoffmann D, Lo Russo G, Munari C (1993): Manifestations électrocliniques induites par la stimulation électrique intracérébrale par chocs dans les épilepsies temporales. Clin Neurophysiol 22:305–326. [DOI] [PubMed] [Google Scholar]
- Kahane P, Minotti L, Hoffmann D, Lachaux JP, Ryvlin P (2004): Invasive EEG in the definition of the seizure onset zone: Depth electrodes In: Rosenow F, Lüders HO, editors. Handbook of Clinical Neurophysiology, Vol. 3 Presurgical assessment of the epilepsies with clinical neurophysiology and functional imaging. Amsterdam: Elsevier BV; pp 109–133. [Google Scholar]
- Kurth F, Zilles K, Fox PT, Laird AR, Eickhoff SB (2010): A link between the systems: Functional differentiation and integration within the human insula revealed by meta‐analysis. Brain Struct Funct 214:519–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacruz ME, Garcia Seoane JJ, Valentin A, Selway R, Alarcon G (2007): Frontal and temporal functional connections of the living human brain. Eur J Neurosci 26:1357–1370. [DOI] [PubMed] [Google Scholar]
- Matsumoto R, Nair DR, LaPresto E, Najm I, Bingaman W, Shibasaki H, Luders HO (2004): Functional connectivity in the human language system: A cortico‐cortical evoked potential study. Brain 127:2316–2330. [DOI] [PubMed] [Google Scholar]
- Matsumoto R, Nair DR, LaPresto E, Bingaman W, Shibasaki H, Luders HO (2007): Functional connectivity in human cortical motor system: A cortico‐cortical evoked potential study. Brain 130:181–197. [DOI] [PubMed] [Google Scholar]
- Matsumoto R, Nair DR, Ikeda A, Fumuro T, Lapresto E, Mikuni N, Bigaman W, Miamoto S, Fukuyama H, Takahashi R, Najm I, Shibasaki H, Luders HO (2012): Parieto‐frontal network in Human studied by cortico‐cortical evoked potential. Hum Brain Mapp 33:2856–2872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazzola L, Isnard J, Mauguiere F (2006): Somatosensory and pain responses to stimulation of the second somatosensory area (SII) in humans. A comparison with SI and insular responses. Cereb Cortex 16:960–968. [DOI] [PubMed] [Google Scholar]
- Mesulam MM, Mufson EJ (1982a): Insula of the old world monkey. I. Architectonics in the insulo‐orbito‐temporal component of the paralimbic brain. J Comp Neurol 212:1–22. [DOI] [PubMed] [Google Scholar]
- Mesulam M, Mufson EJ (1982b): Insula of the old world monkey. III: Efferent cortical output and comments on function. J Comp Neurol 212:38–52. [DOI] [PubMed] [Google Scholar]
- Mesulam MM, Mufson EJ (1985): The insula of Reil in man and monkey In Peters A, Jones EG, editors. Association and Auditory Cortices. New York: Plenum; pp 179–226. [Google Scholar]
- Munari C, Kahane P, Tassi L, Francione S, Hoffmann D, Lo RG, Benabid AL (1993): Intracerebral low frequency electrical stimulation: A new tool for the definition of the “epileptogenic area”. Acta Neurochirurg Suppl 58:181–185. [DOI] [PubMed] [Google Scholar]
- Nathan SS, Sinha SR, Gordon B, Lesser RP, Thakor NV (1993): Determination of current density distributions generated by electrical stimulation of the human cerebral cortex. Electroencephalogr Clin Neurophysiol 86:183–192. [DOI] [PubMed] [Google Scholar]
- Nelson SM, Dosenbach NU, Cohen AL, Wheeler ME, Schlaggar BL, Petersen SE (2010): Role of the anterior insula in task‐level control and focal attention. Brain Struct Funct 214:669–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostrowsky K, Magnin M, Ryvlin P, Isnard J, Guenot M, Mauguiere F (2002): Representation of pain and somatic sensation in the human insula: A study of responses to direct electrical cortical stimulation. Cereb Cortex 12:376–385. [DOI] [PubMed] [Google Scholar]
- Pritchard TC, Norgren, R (2004): Gustatory system In Paxinos G, Mai JK, editors. The Human Nervous System. Amsterdam: Elsevier; pp 1171–1196. [Google Scholar]
- Rosenberg DS, Mauguiere F, Catenoix H, Faillenot I, Magnin M (2009): Reciprocal thalamocortical connectivity of the medial pulvinar: A depth stimulation and evoked potential study in human brain. Cereb Cortex 19:1462–1473. [DOI] [PubMed] [Google Scholar]
- Rutecki PA, Grossman RG, Armstrong D, Irish‐Loewen S (1989): Electrophysiological connections between the hippocampus and entorhinal cortex in patients with complex partial seizures. J Neurosurg 70:667–675. [DOI] [PubMed] [Google Scholar]
- Ryvlin P, Minotti L, Demarquay G, Hirsch E, Arzimanoglou A, Hoffman D, Guenot M, Picard F, Rheims S, Kahane P (2006): Nocturnal hypermotor seizures, suggesting frontal lobe epilepsy, can originate in the insula. Epilepsia 47:755–765. [DOI] [PubMed] [Google Scholar]
- Seltzer B, Pandya DN (1991): Post‐rolandic cortical projections of the superior temporal sulcus in the rhesus monkey. J Comp Neurol 312:625–640. [DOI] [PubMed] [Google Scholar]
- Small DM (2010): Taste representation in the human insula. Brain Struct Funct 214:551–561. [DOI] [PubMed] [Google Scholar]
- Stephani C, Fernandez‐Baca Vaca G, Maciunas R, Koubeissi M, Luders HO (2011): Functional neuroanatomy of the insular lobe. Brain Struct Funct 216:137–149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talairach J, Bancaud J (1973): Stereotaxic approach to epilepsy. Methodology of anatomo‐functional stereotaxic investigations. Prog Neurol Surg 5:297–354. [Google Scholar]
- Valentin A, Anderson M, Alarcon G, Seoane JJ, Selway R, Binnie CD, Polkey CE (2002): Responses to single pulse electrical stimulation identify epileptogenesis in the human brain in vivo. Brain 125:1709–1718. [DOI] [PubMed] [Google Scholar]
- Valentin A, Alarcon G, Garcia‐Seoane JJ, Lacruz ME, Nayak SD, Honavar M, Selway RP, Binnie CD, Polkey CE (2005a): Single‐pulse electrical stimulation identifies epileptogenic frontal cortex in the human brain. Neurology 65:426–435. [DOI] [PubMed] [Google Scholar]
- Valentin A, Alarcon G, Honavar M, Garcia Seoane JJ, Selway RP, Polkey CE, Binnie CD (2005b): Single pulse electrical stimulation for identification of structural abnormalities and prediction of seizure outcome after epilepsy surgery: A prospective study. Lancet Neurol 4:718–726. [DOI] [PubMed] [Google Scholar]
- van't Klooster MA, Zijlmans M, Leijten FS, Ferrier CH, van Putten MJ, Huiskamp GJ (2011): Time‐‐frequency analysis of single pulse electrical stimulation to assist delineation of epileptogenic cortex. Brain 134:2855–2866. [DOI] [PubMed] [Google Scholar]
- Wicker B, Keysers C, Plailly J, Royet JP, Gallese V, Rizzolatti G (2003): Both of us disgusted in my insula: The common neural basis of seeing and feeling disgust. Neuron 40:655–664. [DOI] [PubMed] [Google Scholar]
- Wilson CL, Isokawa M, Babb TL, Crandall PH (1990) Functional connections in the human temporal lobe. Exp Brain Res 82:279–292. [DOI] [PubMed] [Google Scholar]
- Yaxley S, Rolls ET, Sienkiewicz ZJ (1990): Gustatory responses of single neurons in the insula of the macaque monkey. J Neurophysiol 63:689–700. [DOI] [PubMed] [Google Scholar]
- Zhang D, Snyder AZ, Shimony JS, Fox MD, Raichle ME (2010): Noninvasive functional and structural connectivity mapping of the human thalamocortical system. Cereb Cortex 20:1187–1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang ZH, Oppenheimer SM (2000): Electrophysiological evidence for reciprocal insulo‐insular connectivity of baroreceptor‐related neurons. Brain Res 863: 25–41. [DOI] [PubMed] [Google Scholar]