

Abstract
Sentence comprehension is a complex process. Besides identifying the meaning of each word and processing the syntactic structure of a sentence, it requires the computation of thematic information, that is, information about who did what to whom. The present fMRI study investigated the neural basis for thematic reanalysis (reanalysis of the thematic roles initially assigned to noun phrases in a sentence) and its interplay with syntactic reanalysis (reanalysis of the underlying syntactic structure originally constructed for a sentence). Thematic reanalysis recruited a network consisting of Broca's area, that is, the left pars triangularis (LPT), and the left posterior superior temporal gyrus, whereas only LPT showed greater sensitivity to syntactic reanalysis. These data provide direct evidence for a functional neuroanatomical basis for two linguistically motivated reanalysis processes during sentence comprehension. Hum Brain Mapp , 2011. © 2010 Wiley‐Liss, Inc.
Keywords: BA 45, left pars triangularis, Broca's area, left inferior gyrus, superior temporal gyrus, syntactic processing, thematic reanalysis
INTRODUCTION
To understand a sentence, the information about who did what to whom, that is, thematic information, has to be computed. For example, comprehension of the sentence “John gave Mary a present” involves, besides identifying the meaning of each word and processing syntactic relations between the words, the assignment of the following thematic roles to each phrase in the sentence: “John” is the agent of a present‐giving action, “Mary” is a recipient of that action, and “a present” is the theme undergoing the action. Previous evidence in both psycholinguistics and event‐related brain potentials (ERPs) suggests that thematic information (i.e., assignment of thematic roles to a sentence) is processed online, immediately upon the reception of each word. It further suggests that there is a default (or initial canonical) interpretation about which thematic role is assigned to which phrase during online comprehension of a sentence [Bornkessel and Schlesewsky, 2006; Bornkessel et al., 2002, 2003; Kamide et al., 2003; Kuperberg, 2007; Kuperberg et al., 2003, 2006, 2007; Mazuka and Ito, 1995]. In some sentence constructions, the initial assignment of thematic roles (e.g., agent role before patient role) turns out to be wrong at a later point during sentence comprehension. In such cases, the original assignment of thematic roles must undergo revision, “thematic reanalysis.” Consider, for example, the passive sentence “John was given a present by Mary.” When presented in an out‐of‐the‐blue context and read or listened to online, “John” is initially assigned an agent role (at the time when “John” is heard or read), but when the predicate “was given” is processed, the thematic role must be reanalyzed from the agent role to a recipient role, so that the sentence is interpreted correctly as a passive sentence. Numerous psycholinguistic studies provide evidence of a processing cost induced by thematic reanalysis (e.g., see Clifton et al. [ 2003] for an overview). Furthermore, recent ERP findings provide clear evidence that thematic (re)analysis is a distinct linguistic processes, different from other kinds of linguistic process such as that required to comprehend a complex structure as opposed to a simple structure [Bornkessel and Schlesewsky, 2006; Bornkessel et al., 2003; Kuperberg, 2007; Kuperberg et al., 2007]. The thematic reanalysis process, if psychologically real as suggested by previous psycholinguistic research [Altmann, 1999; Carlson and Tanenhaus, 1988; Clifton et al., 2003; Kamide et al., 2003; MacDonald et al., 1994; McRae et al., 1997; Rayner et al., 1983; Scheepers et al., 2000; Trueswell et al., 1994] and neurophysiological studies [Bornkessel and Schlesewsky, 2006; Bornkessel et al., 2002, 2003, 2005; Kuperberg, 2007; Kuperberg et al., 2003, 2006, 2007], should involve as its neural basis some regions of the brain related to language processing.
General Aims and Previous Findings in Related Research Areas
The present work aims to identify the neural basis of a particular type of linguistic process, that is, thematic reanalysis during sentence comprehension and its interplay with syntactic (“movement”) reanalysis. Identification of a functional–neuroanatomical basis for such linguistic processes is critical in understanding the function of language‐related areas in the human brain. When it comes to the investigation of complex processes such as the reanalysis processes in sentence comprehension, theoretical development as well as experimental findings in the relevant interdisciplinary research areas (e.g., linguistics, neurolinguistics, and neurocognitive models of language) must be considered at the same time. In what follows, previous proposals and findings related to key components of the present study (e.g., thematic roles, syntactic “movement” and thematic processes, and reanalysis processes) in the relevant research areas will be briefly discussed.
First, concerning thematic roles, linguistic theories—at least some—assume that thematic role assignments are syntactic as well as semantic in nature (see, e.g., Chomsky [1981]). According to these theories, verbs specify thematic role information by assigning thematic roles to the noun phrases in a sentence. In this view, thematic information is closely tied to the syntactic structure of a sentence; a sentence's basic structure is determined by a verb and a noun or noun phrases, which are linked to thematic roles specified by the verb. For example, in a sentence like “John gave Mary a present,” the verb “give” asks for three noun phrases, “John” (a subject noun phrase = agent), “Mary” (an indirect object noun phrase = recipient), and “a present” (a direct object noun phrase = theme). Thematic information is also semantic in nature, because it indicates “semantic” (or “thematic”) relations between the entities denoted by each of the noun phrases in a sentence. In the previous sentence, the assigned thematic roles, agent, recipient, and theme identify semantic relations between “John,” “Mary,” and “a present.” If the linguistic assumptions laid out here are directly connected to the nature of language processing and its brain basis, thematic role (re)assignments should exhibit brain activation related to syntactic and semantic processing.
Second, turning to neurolinguistic theories, with respect to syntactic processes, a process called “movement,” hypothesized to apply to sentences with noncanonical word order, lies in Broca's area in the left inferior frontal gyrus (IFG) [Grodzinsky, 2000]. Previous neuroimaging studies have provided support for such a proposal by showing that different subregions of Broca's area, for example, the left pars triangularis (LPT; BA 45) and the left pars opercularis (BA 44), are responsible for processing different linguistic input. Although the processing of lexical‐semantic information relies on BA 45/47 in addition to the middle and posterior portion of the left superior temporal gyrus (STG) and the middle portion of temporal gyrus [Vigneau et al., 2006], syntactic processes recruit Broca's area (BA 44/45) together with anterior and posterior portions of the left STG (for an overview, see Bookheimer [ 2002], Friederici [ 2002], and Grodzinsky and Friederici [ 2006]; see also Ben‐Shachar [ 2003, 2004] and Wartenburger et al. [ 2004]). Even within the Broca's area, recent studies have suggested that syntactic structures that require “movement” recruit BA 45 ([Santi and Grodzinsky, 2007a, b]; see also Ben‐Shachar et al. [ 2003, 2004] and Kinno et al. 2008), whereas structural complexity generated by the number of permutations of noun phrases or by the number of embedded phrases within a sentence induces greater activation of BA 44 [Friederici et al., 2006; Makuuchi et al., 2009]. Furthermore, directly related to the investigation of thematic processing and thematic reanalysis, Bornkessel et al. [ 2005] found that the complexity associated with “argument hierarchization” (i.e., linearization of a hierarchical structure such as placing a subject (agent) before an object (theme)) elicited increased activity in a fronto‐temporal network, with the processing of a (thematic) hierarchical structure resulting in activation in BA 44 and the processing of morphological information relevant for linearization (i.e., case markers in German) recruiting left posterior superior temporal sulcus (STS) (see also Grewe et al. [ 2005]).
Third, previous ERP evidence suggests that thematic and syntactic reanalyses are distinct neurocognitive processes reflected in different ERP components (i.e., N400 and P600 effects), occurring at different time points in language processing. Frisch and Schlesewsky [ 2005] have shown that problems of semantic–thematic integration are indicated by an N400 effect, whereas the cost of syntactic integration arriving at a well‐formed structure for a sentence elicits a posterior P600 effect (cf., Hagoort et al. [ 1993], Osterhout and Holcomb [ 1992]; see Friederici [ 2002] for an overview). A previous study by the same researchers [Frisch and Schlesewsky, 2001] provides additional support for the N400 effect not only being associated with the cost attributed to the general lexical semantic integration, as widely assumed in the ERP literature, but also the thematic problem (for an overview, see Kutas and Federmeier [ 2000]; for recent theoretical development concerning the N400 effect, see, e.g., Bornkessel et al. [ 2002, 2003, 2005], Bornkessel and Schlesewsky [ 2006], Kuperberg [ 2007], and Kuperberg et al. [ 2003, 2006, 2007]). As for a P600 effect, two different syntactic subprocesses, that is, reanalysis and repair, are associated with different topographical distributions. A P600 effect with an anterior distribution is associated with an increased integration cost for processing complex syntactic structures (or syntactic reanalysis), whereas a P600 with a central–parietal distribution reflects a repair process for sentences with syntactic violations (Friederici et al. [ 2002]; see also Kaan and Swaab [ 2002, 2003]). In summary, the ERP studies seems to have provided evidence for two reanalysis processes, thematic reanalysis (correlated with an N400) preceding syntactic reanalysis (correlated with a P600) in online sentence comprehension.
Fourth, neurocognitive models must also be looked into given an interesting case examined in this study. The notion of “reanalysis” in sentence comprehension implies that the structure of a sentence initially processed (i.e., the initial syntactic structure or a canonical structure of sentence, as well as the thematic roles originally assigned to the phrases based on the available input) has to be given up when contradictory information concerning the syntactic structure or thematic roles of the phrases is signaled by the input, and received and processed by listeners or readers. In other words, thematic reanalysis may be viewed as a case in which there is a mismatch between the thematic structure constructed in the listeners or readers' mind up to some point in time (based on language input available up to that point) and the actual (or correct) thematic roles that must be assigned to the phrases using information such as morphological markers, which may come in at a later point in time. Likewise, syntactic reanalysis may be viewed as a case in which conflicting information regarding the syntactic structure of a sentence is involved. Many models proposed in the domain of cognitive neuroscience, especially those concerning inhibition processes, generally assume that the prefrontal cortex is recruited for inhibition and related processes. Some of these hold that BA 45 is associated with the selection of task relevant items and the inhibition of irrelevant items [Badre and Wagner, 2007; Rodd et al., 2005]. If one hypothesizes that the reanalysis process in sentence processing mirrors the inhibition of the preferred canonical structure, one would predict that increased activation in BA 45 should be observed as a function of inhibition cost.
The Present Study in Japanese: Aims and Hypotheses
The central goal of the present work is to determine the neural basis for thematic reanalysis and its relation to another crucial process, syntactic (“movement”) reanalysis, which also plays an important role during sentence comprehension. To pursue this, the present study addresses the question of whether different brain activation patterns can be observed when two linguistically distinct processes are computed: thematic versus syntactic “movement” reanalysis. Japanese serves as an excellent testing ground for studying these distinct linguistic processes. As detailed below, Japanese allows the processes in question to occur exactly at the same location within a sentence, that is, at the verb that appears consistently at the sentence final position. Despite the fact that a verb always appears at the end of a sentence, Japanese sentences are processed incrementally, that is, input is processed on a word by word basis without delay even before encountering the verb [Aoshima et al., 2004; Kamide and Mitchell, 1999; Wolff et al., 2008]. Relevant to the current study, case markers attached to the noun phrase, which indicate a grammatical relation of noun phrases to the verb such as subject and object, are used to assign thematic roles to each phrase as the phrases are processed [Kamide et al., 2003; Mazuka and Ito, 1995]. That is, as depicted in Figure 1, before the verb is heard or read, each noun phrase receives a thematic role (agent, recipient, and theme) via case marking information [e.g., nominative (NOM), dative (DAT), and accusative (ACC)] (see black arrows in Fig. 1).
Figure 1.
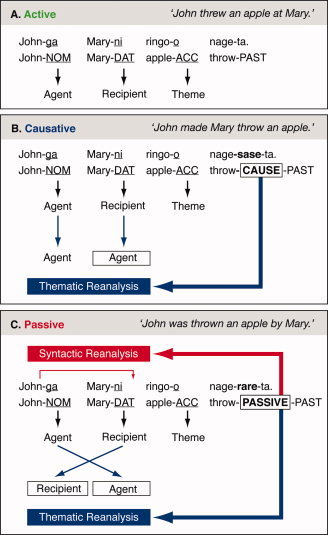
Predicted thematic and syntactic reanalysis. Black arrows indicate a default assignment of thematic roles, blue arrows thematic reanalysis, and red arrows syntactic reanalysis. As a default thematic role assignment (A), a phrase with a nominative marker (NOM) is assigned to “Agent” (an actor of an event), one with a dative marker (DAT) “Recipient” of an event, and one with an accusative marker (ACC) “Theme” (an object that undergoes an event). In Causative sentence (B), the causative morpheme sase triggers the second phrase “Mary‐DAT” for thematic reanalysis from “Agent” to “Recipient” and in Passive sentence (C), the passive morpheme rare requires thematic roles for the first two phrases, “John‐NOM” and “Mary‐DAT,” to be interchanged (“Agent” to “Recipient” and vice versa). In addition, Passive sentence (C) undergoes a syntactic reanalysis “movement,” that is, an additional structural change relating the first phrase “John‐NOM” to the syntactic position before the accusative phrase “apple‐ACC.”
The current fRMI study investigated the following three linguistic processes: (a) the default assignment of thematic roles, (b) thematic reanalysis, and (c) syntactic reanalysis. For an active sentence (Fig. 1A), the default assignment of thematic roles (i.e., “agent” first, “recipient” second, and “theme” third) is correct and, hence, no thematic reanalysis is required at the end of the sentence. In contrast, both causative (Fig. 1B) and passive sentences (Fig. 1C) require thematic reanalysis at the sentence final verb (i.e., at the causative and passive markers or morphemes attached to the verb). In the causative and passive sentences, the initial assignment of thematic roles guided by case marking information of the noun phrases (NOM, DAT, and ACC) does not coincide with the thematic roles which the special causative and passive morphemes at the sentence final verb call for. (For the predicted thematic reanalysis (see Fig. 1B,C); thematic reanalysis is indicated by blue arrows). A passive sentence, in addition to the expected thematic reanalysis described earlier, requires syntactic reanalysis. “Syntactic reanalysis” here corresponds to the process that requires the phrase John to “move” to a different syntactic position within a sentence to construct the correct underlying structure for the passive sentence (Fig. 1C; see red arrows). (For recent literature supporting the movement analysis for Japanese passive sentences adopted here, see, e.g., Hoshi [ 1991], Shibatani [ 1990], and Terada [ 1990]; for an alternative proposal, refer to Kuroda [ 1979]). Crucially, passive and causative sentences differ in that in the former, but not in the latter, a syntactic “movement” reanalysis process is involved (cf. Fig. 1B,C) (For the linguistic assumptions relied on here, see Hoshi [ 1999] and Miyagawa [ 1999]). As illustrated in Figure 1, both thematic and syntactic reanalyses are triggered exactly at the same position within a sentence for both causative and passive sentences, that is, where the critical morphemes appear at the sentence final verb. This feature of Japanese allows us to test these different types of reanalysis processes in a well‐controlled manner, as sentences are identical up to the critical position.
In summary, both causative and passive sentences undergo thematic reanalysis, while passive sentences, but not causative sentences, are subject to syntactic reanalysis. Active sentences are free from any of these reanalysis processes. These properties of Japanese, all together, create interesting test sentences to study brain responses for linguistically distinct reanalysis processes, that is, thematic and syntactic reanalysis processes.
MATERIALS AND METHODS
Participants
Sixteen native speakers of Japanese (12 females, mean age 25.63 years, SD 3.26) participated in the present experiment after receiving written informed consent. No participant had any history of speech, hearing, neurological, or psychiatric disorders. All participants had normal or corrected vision and were right handed (mean laterality quotient 95.44; Oldfield [ 1971]).
Materials
Participants listened to Japanese sentences of the form in Table I. To investigate brain areas supporting thematic reanalysis, the comparison for activation elicited by sentences requiring thematic reanalysis (Causative and Passive) and those requiring no thematic reanalysis (Active) was crucial (see Table I and Fig. 1; see blue arrows for Causative and Passive in Fig. 1). In addition, Passive against Causative provided a critical comparison to investigate the effect of syntactic “movement” reanalysis. Passive, but not Causative, requires syntactic reanalysis in addition to thematic reanalysis (see red arrows in Fig. 1).
Table I.
Example sentences
| Conditions | Example sentences | |||||
|---|---|---|---|---|---|---|
| A. | Active | Mikkamae | John‐ga | Mary‐ni | Ball‐o | Nageta |
| “Three days ago John threw a ball at Mary” | ||||||
| B. | Causative (thematic reanalysis) | Mikkamae | John‐ga | Mary‐ni | Ball‐o | Nagesaseta |
| “Three days ago John made Mary throw a ball” | ||||||
| C. | Passive (thematic and syntactic reanalysis) | Mikkamae | John‐ga | Mary‐ni | Ball‐o | Nagerareta |
| “Three days ago John was thrown a ball by Mary” | ||||||
Procedures
A total of 34 spoken sentences for each condition (Active, Causative, and Passive) constituted the critical materials in this experiment. A temporal adverbial phrase (e.g., the other day, around noon, in the evening, and two years ago) was added at the beginning of each sentence. One hundred and two experimental sentences were combined with 68 filler sentences. The filler sentences took the form of either active sentences with different morphemes attached to the verb or passive sentences requiring no thematic reanalysis at the sentence final position. The filler sentences were included to balance the ratio of active versus nonactive sentences and the ratio of sentences requiring reanalysis process versus those requiring no reanalysis process. Sentence materials (both experimental sentences and fillers) were recorded by a native speaker of Japanese. All sentences were spoken as naturally as possible. Acoustic analyses for the sentences were provided to ensure that the experimental sentences did not differ from each other prosodically. The length of sentences was systematically controlled (∼5 s). Table II presents the results of the acoustic analyses, that is, F0 maxima and the duration of each phrase in the experimental sentences (note that the intensity level for the sentences was normalized to less than 73 db and therefore, the acoustic analysis on this measure was not included.) In addition, Figure 2 provides pitch (F0) information in hertz for each condition. One‐way ANOVAs with the factor CONDITION (Active, Causative, and Passive) were conducted for each phrase of the sentences on each acoustic measurement reported in Table II. The results showed no significant difference among the three conditions (all F's < 0.79).
Table II.
Acoustic analyses
| Condition | Adverbial phrase | Nominative phrase | Dative phrase | Accusative phrase | Verb | |
|---|---|---|---|---|---|---|
| Stem | Critical morpheme | |||||
| F0 maxima of each phrase (Hz) | ||||||
| A. Active | 280 | 316 | 280 | 267 | 236 | 217 |
| B. Causative | 279 | 316 | 277 | 266 | 240 | 218 |
| C. Passive | 279 | 319 | 276 | 266 | 238 | 219 |
| Duration of each phrase (s) | ||||||
| A. Active | 1.48 | 0.71 | 0.79 | 1.17 | 0.19 | 0.62 |
| B. Causative | 1.42 | 0.67 | 0.79 | 1.17 | 0.19 | 0.78 |
| C. Passive | 1.45 | 0.70 | 0.79 | 1.17 | 0.18 | 0.76 |
Figure 2.
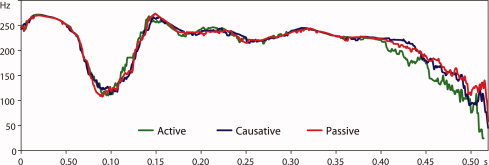
Time course pitch (F0) information for sentence materials. F0 (Hz) information was extracted from each sentence, and the average F0 value for every 10–ms period for each of the conditions was plotted in the figure. The figure shows that the sentence materials did not differ prosodically among the three conditions (Active, Causative, and Passive) up to the critical morpheme appearing at the sentence final verb.
All the sentences (both experimental sentences and fillers) were systematically distributed across two lists, each containing a total of 170 sentences and divided into three blocks, so that no one sentence appeared in more than one condition in one block of an experimental session. Within each list, sentences were pseudo‐randomized using constraints such that the same type of sentences (e.g., active sentence and active filler) did not appear more than twice in a sequence. Each of the lists was randomized twice and as a result, a total of four lists were created (two lists × two randomizations). Each participant was randomly assigned to one of the four lists.
At the beginning of each trial, a small fixation star appeared in the middle of a screen, which participants saw via a small mirror placed in front of their eyes. After 500 ms, a sentence was presented to participants over headphones. The fixation star remained on the screen until the end of the sentence. Following 20% of the total number of trials, a comprehension question about the content of the previous sentence was presented auditorily. For example, a Japanese sentence corresponding to “John threw a ball” was played as a question sentence to the Japanese test stimuli “John made Mary throw a ball.” Two choices (“yes” or “no” written in Japanese) to the question appeared on the screen until one of the choices was selected by pressing one of the buttons on a key pad or a time‐out period of 3,000 ms was reached.
fMRI Data Acquisition
Twenty axial slices (4‐mm thickness, 1‐mm interslice distance, FOV 19.2 cm, data matrix of 64 × 64 voxels, in‐plane resolution of 3 × 3 mm) were acquired every 2 s during functional measurements (gradient‐echo EPI sequence, TR = 2 s, TE = 30 ms, flip angle 90°, and acquisition bandwidth 116 kHz) with a 3 Tesla Siemens TRIO scanner (Siemens, TRIO, Erlangen). Before functional imaging, T1‐weighted MDEFT images (data matrix 256 × 256, TR 1.3 s, TE 10 ms) were obtained with a non–slice‐selective inversion pulse followed by a single excitation of each slice [Norris, 2000]. These were used to co‐register functional scans with previously obtained high‐resolution whole‐head 3D brain scans (128 sagittal slices, 1.5‐mm thickness, FOV 25.0 × 25.0 × 19.2 cm, and data matrix of 256 × 256 voxels) [Lee et al., 1995].
fMRI Data Analysis
The functional imaging data were processed with the software package SPM5 (available at http://www.fil.ion.ucl.ac.uk/spm/). The first five volumes of each fMRI session were discarded to eliminate magnetic saturation effects, resulting in a total of 1,200 volumes. Structural images were corrected for a signal intensity bias due to the magnetic field inhomogeneity using the bias correction tool in SPM5. As a preprocessing step, the EPI images were realigned to the first image, and the slice time correction was applied. EPI images were coregistered to the participants' T1 and then to 3D high‐resolution structural images. The normalization of an individual structural image to the SPM5 T1 brain template was processed in two steps: (1) estimation of the normalization parameters and (2) writing the normalized images with the obtained parameters. These parameters transformed the structural images and all the EPI volumes into a common stereotaxic space to allow multiparticipant analyses. The EPI images were resampled into 3 × 3 × 3 mm3 voxels and the structural images into 1 × 1 × 1 mm3 voxels with the seventh degree B‐spline interpolation.
The statistical evaluation was based on a least‐squares estimation using the general linear model for serially autocorrelated observations [Worsley and Friston, 1995]. The design matrix was generated with a synthetic hemodynamic response function [Friston et al., 1998; Josephs et al., 1997]. All the test sentences were treated as a common condition from the beginning of the sentence to the critical morpheme, that is, the point in each sentence at which thematic information was disambiguated. The critical morphemes, which differed among the different sentence conditions, were modeled as different conditions, that is, Active, Causative, and Passive (see Table I for examples; the critical morphemes are highlighted in different colors). More specifically, the onset of the critical morpheme was treated as a single event and the time difference between the onset of the critical morpheme and the end of the sentence was discarded. Questions were modeled as an additional, separate condition. For each participant, three contrast images were generated. These images represented the direct contrast in the processing of (1) Causative versus Active, (2) Passive versus Active, and (3) Passive versus Causative. Single‐participant contrast images were entered into a second‐level random effects analysis for each of the contrasts (i.e., Causative vs. Active, Passive vs. Active, and Passive vs. Causative). The group analysis consisted of a one‐sample t‐test across the contrast images of all the participants. To protect against false‐positive activation, a double threshold was applied: for t‐value threshold, voxels with a t‐score above 2.60 (P < 0.01, uncorrected) were selected, and survived clusters that had a volume exceeding 80 voxels (2,160 mm3) were considered as significantly activated at P < 0.05, corrected for multiple comparisons.
To further characterize the activated regions, the time course for each of the trials for the left posterior temporal gyrus (LpSTG) and the LPT was plotted. First, the volumes of interest (VOI) were defined as 6‐mm radius spheres with the local maxima of individual participants nearest to the group (all participants) maxima (−42 −57 21) and (−42 60 24) for LpSTG and (−54 27 6) and (−42 27 0) for LPT, which were computed based on Causative versus Active and Passive versus Active (see Table III). Second, time series data were extracted as eigenvariates (by SPM5), and the time course for each of the trials was estimated using preprocessed time series data for each participant. More specifically, the time course for each trial for each condition (i.e., sentences from the onset to the critical morpheme, three different morphemes, and questions) were modeled with 17 variables representing the BOLD signals obtained in every second of the trials starting from the onset of the critical morpheme (i.e., 0, 1, 2,…, 16 s). The simultaneous equations were solved against the time series data of VOIs, assuming a linear time invariant system. Paired t‐tests between the three conditions (i.e., Active, Causative, and Passive) were carried out for the BOLD signals at each time point (i.e., every second up to 16 s from the onset of the critical morpheme) in the trials.
Table III.
Brain regions activated by Causative vs. Active and Passive vs. Active comparisons
| Region | Tmax | Cluster size | Location | ||
|---|---|---|---|---|---|
| (number of voxles) | x | y | z | ||
| Causative > Active | |||||
| Left | |||||
| pars triangularis | 6.95 | a706 | −54 | 27 | 6 |
| dorsal premotor area | 5.87 | a706 | −42 | 3 | 18 |
| putamen | 5.13 | a706 | −12 | 6 | 3 |
| superior temporal gyrus | 6.63 | b499 | −42 | −57 | 21 |
| heschel gyrus | 6.02 | b499 | −63 | −18 | 6 |
| Right | |||||
| superior temporal gyrus | 6.26 | 181 | 66 | −21 | 9 |
| anterior cingulate cortex | 6.05 | 293 | 9 | 21 | 48 |
| putamen | 6.1 | 98 | 18 | 12 | 0 |
| cerebellum | 4.73 | 110 | 36 | −60 | −30 |
| Passive > Active | |||||
| Left | |||||
| pars triangularis | 5.89 | 380 | −45 | 27 | 0 |
| superior temporal gyrus | 4.58 | 84 | −42 | −60 | 24 |
| middle temporal gyrus | 5.96 | 160 | −54 | −36 | −3 |
| putamen | 6.79 | c615 | −15 | 9 | 15 |
| thalamus | 5.25 | c615 | −9 | −9 | −3 |
| Right | |||||
| dorsal premotor area | 4.87 | d153 | 36 | 12 | 36 |
| anterior insula | 4.9 | d153 | 39 | 18 | −9 |
| putamen | 5.05 | e615 | 15 | −9 | 6 |
| thalamus | 5.02 | e615 | 15 | 6 | 6 |
| anterior cingulate cortex | 5.59 | 287 | 9 | 18 | 48 |
Note: T‐values are reported for voxels of greatest activity within activated clusters. Locations of these voxels are given in MNI coordinates.
a‐e: Five distinct anatomical regions were selected.
RESULTS
Behavioral Data
One‐way ANOVAs with the factor CONDITION (Active, Causative, and Passive) were carried out for accuracy rates and reaction times for the comprehension questions, using error terms based on participant variability. The mean percent accuracy rates (and their SDs in parentheses) for the comprehension questions were 83 (0.14) for Active sentences, 85 (0.18) for Causative sentences, and 83 (0.18) for Passive sentences. The one‐way ANOVA of the accuracy rates showed no significant effect of CONDITTION [F(2,30) = 0.12, P = 0.89], which probably suggests that sentences of all different conditions were understood equally well. In addition, mean response times (ms) for the comprehension questions did not indicate significance of CONDITION (SDs in parentheses): Active 659.49 (209.07), Causative 752.66 (295.03), and Passive 708.49 (262.51) [F(2,30) = 1.24, P = 0.3].
fMRI Data
The whole‐brain analysis for the contrasts of Causative versus Active and Passive versus Active showed significant activation in the regions listed in Table III. No significant difference was found for the Passive versus Causative contrast in the whole brain analysis. Both Causatives and Passives, when contrasted with Active sentences, showed a large activation cluster in the left IFG expanding into the dorsal premotor cortex and moreover in the left middle and posterior temporal cortex. According to our hypothesis, the most important regions within these activation clusters are the LPT and the LpSTG. For these brain regions, region of interest (ROI) analyses were conducted (see Methods section for information regarding how the analyses were conducted). Specifically, analyses were carried out to test whether those two regions (LPT and LpSTG) showed different effects for different comparisons, that is, Causative versus Active, Passive versus Active, and Passive versus Causative. The time course for activation in LPT and LpSTG for all the conditions starting from the critical morphemes was plotted and paired t‐tests were conducted between the conditions. As presented in Figure 3, a significant signal change was observed for both Causative > Active and Passive > Active in both LPT and LpSTG between 4 and 6 s (see the time windows highlighted in sky blue on the left and right graphs). In addition, the LPT revealed an additional effect, which was significant at 8 s for Passive > Causative only (see the orange arrow pointing at 8 s in the graph for Passive > Causative). That is, in addition to the significant activation increase in LPT and LpSTG for Causative and Passive each against Active, there was a critical time course difference in the activation of LPT for Passive versus Causative.
Figure 3.
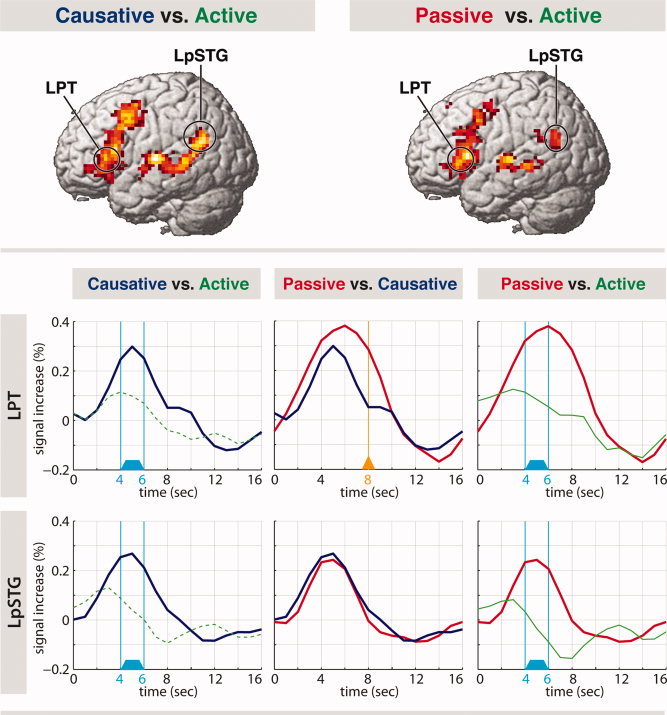
Brain regions sensitive to thematic and syntactic reanalyses. Top: Brain activation maps. Causative versus Active and Passive versus Active contrasts with the threshold of corrected P < 0.05. Bottom: Time course plots of brain activation for the LPT and LpSTG VOI. Time 0 corresponds to the onset of the critical morpheme. Active is represented in green (a dashed line using the Causative VOI and a solid line using the Passive VOI), Causative in blue, and Passive in red. LPT (BA 45) is involved in both thematic and syntactic reanalyses. Thematic reanalysis, required for both Causative and Passive, leads to increased activation in LPT between 4 and 6 s (indicated by sky blue) and precedes syntactic reanalysis, which elicits increased activation for Passive in the same region at 8 s (indicated by orange arrow). LpSTG is recruited for thematic reanalysis as indicated by the increased activation for both Causative and Passive between 4 and 6 s (indicated by sky blue).
DISCUSSION
The aim of the present fMRI study was to investigate the functional neuroanatomical basis for thematic reanalysis and its relation to syntactic reanalysis during sentence comprehension. To this end, the study tested Japanese causative and passive sentences, in which the relevant reanalysis processes, that is, thematic reanalysis and/or syntactic reanalysis, took place. Passive sentences in Japanese, when compared with their active counterparts, require not only thematic reanalysis but also syntactic reanalysis, which specifically calls for restructuring of the syntactic position of a phrase (“movement”) within a sentence. Causative sentences, in contrast, only require thematic reanalysis (see Fig. 1). Thus, any activation observed for both causative versus active and passive versus active is likely to be an effect reflecting thematic reanalysis. Likewise, any activation in the perisylvian language regions specific for passive versus causative (but not for other contrasts) can be attributed to the additional syntactic reanalysis required for passive sentences. Recall that both thematic and syntactic reanalyses are predicted to occur at exactly the same position in each of the sentences across all the conditions, that is, at the position of the critical morpheme appearing at the sentence final verb (see Fig. 1). This unique property of Japanese made it possible to create controlled stimuli for the experiment. Because the sentence materials did not differ prosodically up to the occurrence of the critical morpheme (see Table II and Fig. 2 for the acoustic analysis), any difference observed between the conditions is likely due to the difference in the processing cost associated with each of the conditions. In other words, the sentence materials were created in such a way that listeners could not identify the sentence structure before encountering the critical morpheme at the sentence final verb. Accordingly, fMRI data were analyzed by having the BOLD signals time‐locked to the onset of the critical morpheme (see Methods section for information about the data analyses).
The present data suggest that the LpSTG is recruited for thematic reanalysis. The LpSTG showed increased activation for processing both causative and passive sentences, as predicted by the theoretical view that both causatives and passive sentences require thematic reanalysis. In addition, it was found that the LPT in Broca's area supports not only thematic reanalysis but also syntactic reanalysis. Furthermore, the study demonstrated the crucial involvement of LPT during syntactic reanalysis. This was shown by a prolonged activation in time for passive sentences compared with causative sentences.
Overall, the results of the reported fMRI experiment suggest that thematic reanalysis is subserved by a neural network comprising LPT and LpSTG, whereas the LPT is crucial for syntactic reanalysis. In what follows, the brain activations associated with thematic and syntactic reanalyses, along with implications for the relevant fields, will be discussed.
Thematic Reanalysis: Broca's Area and LpSTG
Thematic reanalysis process was supported by a neural network consisting of the left Broca's area (BA 45), that is, the LPT and the left posterior STG (LpSTG), as revealed by the ROI analysis. In the current study on Japanese sentence comprehension, the thematic reanalysis required the retrieval of the already assigned thematic roles and proper reassignment of roles to phrases in the sentence. This process was triggered by special morphological markers attached to the verb and not by an explicit change in the syntactic structure of the sentence. The study by Bornkessel et al. [ 2005] has also reported the left posterior superior temporal region (pSTS) as part of the network subserving thematic hierarchization with this region being particularly sensitive to morphological information and verb class. The neural networks identified for thematic processes in German and Japanese are quite similar. The network described for thematic hierarchization in German [Bornkessel et al., 2005] is defined as consisting of the left Broca's area (pars opercularis, BA 44) and the left pSTS, whereas the neural network for thematic reanalysis in the present study on Japanese relies on the left Broca's area (pars triangularis, BA 45) and the left pSTG with the respective activation maxima, however, only lying millimeters apart.
Posterior STG has been implicated previously in the processes of sentence evaluation and of sentential integration ([Friederici et al., 2003]; see Vigneau et al. [2007] for a review). This area, moreover, is known to be integrative across different modalities, that is, auditory and visual [Hickok and Poeppel, 2007] and across different linguistic domains, that is, integration of syntax and semantics [Ben‐Shachar et al., 2003; Bornkessel et al., 2005; Friederici et al., 2003; Grodzinsky and Friederici, 2006]. Such a view on the posterior STG is consistent with the current findings on thematic reanalysis in Japanese. The thematic reanalysis process requires extracting correct thematic roles at the sentence final verb position and integrating them to the entire sentence, that is, integration of reassigned thematic roles to each phrase in the sentence. In addition, in the case of passive sentences, not only thematic reanalysis but also syntactic reanalysis, that is, moving a phrase to the correct syntactic position, must be carried out.
Syntactic Reanalysis: The Role of Broca's Area
The activation observed in the left Broca's area (BA 45), that is, the LPT, for syntactic “movement” reanalysis in Japanese is an important new finding. The ROI analysis of BA 45 indicated that this area is involved in the processing of both passive and causative sentences. Crucially, a time course ROI analysis of BA 45 revealed a differential effect between the two reanalysis types. Although both passives and causatives showed a significant activation enhancement from 4 to 6 s, passives, but not causatives, had an enhancement at 8 s (see Fig. 3). The latter effect is probably attributed to an additional syntactic reanalysis process required for passives. These results may imply that syntactic reanalysis reaches the activation peak in LPT later or lasts longer than thematic reanalysis. Such a finding is consistent with the ERP evidence, suggesting that thematic reanalysis (N400) occurs before syntactic reanalysis (P600) during online sentence comprehension. The present results can be integrated nicely into a general theory on the role of Broca's area in language processing.
Previous work has viewed Broca's area as an area supporting syntactic processes as well as thematic processes. Across different studies in different languages using different experimental paradigms, BA 44 and BA 45 were interpreted to reflect different aspects of language processing [Bookheimer et al., 2002; Friederici, 2002; Hagoort, 2005; Poldrack et al., 2001; Vigneau et al., 2006]. For example, Hagoort [ 2005] and Newman et al. [ 2003] reported that BA 44 is critical in syntactic processing and BA 45 in thematic processing. Santi and Grodzinsky [2007], however, proposed that BA 45 is recruited for the syntactic operation of movement, whereas BA 44 is involved for binding two elements in a sentence, for example, a reflexive pronoun and its referent. In their study, the movement operation, which showed sensitivity in BA 45, was realized in English sentences with a relative clause in which the object noun phrase was moved in front of the subject noun phrase. Because English is a strict word‐order language, such a movement operation associated with a relative clause probably made the correct assignment of thematic roles more difficult. A recent study investigating the assignment of thematic roles to noun phrases in German argued for a crucial involvement of BA 44 rather than BA 45 [Bornkessel et al., 2005]. In their study, a verb class of the sentence final verb, word order, and morphological marking were manipulated. Word‐order manipulations have commonly been seen to modulate activity within BA 44 [Ben‐Shachar, 2003, 2004], so the focus of activity within Bornkessel et al.'s study may be driven by the word order manipulation. Those previous studies all together suggest that both BA 44 and BA 45 are involved in both syntactic and thematic processing, although they seem to point to different sensitivities to different aspects of language processing. The present data on processing different types of Japanese sentences have demonstrated that BA 45 (or LPT) is involved both in syntactic and thematic reanalysis, but with syntactic reanalysis following thematic reanalysis in time. This result is in line with the previous studies and may further explain why BA 45 was found to be activated for both syntactic and thematic reanalysis in the previous studies.
Of course, a considerable degree of caution must be taken when a time‐course analysis of fMRI data is interpreted. Alternatively, one could assume that both thematic and syntactic processes started at the same time, triggered by the critical morpheme, which appeared at the identical position of the sentence. While thematic reanalysis (causatives) ended early, syntactic reanalysis (passives) or a combination of thematic and syntactic reanalyses may have taken longer to complete. Although the present study cannot rule out such an alternative account (besides relying on independent support from the previous ERP studies), the data are in line with our prediction: An additional linguistic process, that is, syntactic reanalysis, is responsible for later and longer lasting activation of the left Broca's area (LPT) for passives. The crucial difference in passives versus causatives lies in the time course analysis of LPT; a similar time modulation was not observed in LpSTG. Because the interpretation of current data is limited to the case investigated in this study (i.e., passive and causative sentences in Japanese), further studies are needed in the future.
Other Areas of Activation
A number of brain areas both inside and outside of the perisylvian cortex were activated for causatives and passive sentences, when each of the conditions compared with active sentences. These areas were the left Heschl's gyrus and the putamen and thalamus bilaterally, in addition to some of the right temporal and frontal areas (see Table III). The temporal region in the right hemisphere may be considered as homolog activations to the left hemisphere, as often described in language studies, in particular, auditory studies in language comprehension [Friederici et al., 2000; Meyer et al., 2004]. The additional hemispheric activations cannot be functionally specified on the basis of the present data, and further research is clearly needed in the future.
Left Heschl's gyrus
Both causative and passive sentences activated the left Heschl's gyrus. This activation may reflect a specific type of low‐level linguistic processing. The present study, together with previous evidence [Herrmann et al., 2009], may suggest that the left Heschl's gyrus is activated when syntactic processes (not specific to those requiring “movement”) are triggered by morphological information in linguistic input. A recent MEG study has reported the activation of the left primary auditory cortex under a specific condition in which listeners' predictions for the syntactic structure of a sentence did not match the morphological markers that appeared in the sentence [Herrmann et al., 2009]. Such a case is analogous to causative and passive sentences in the current study. In both causative and passive sentences, the critical morpheme (“sase” and “rare” illustrated in Table I) attached to the verb signaled to the listeners that sentence reanalysis was needed.
Subcortical areas
The activations observed in the subcortical areas are also interesting. The putamen, as part of the basal ganglia, was activated in both causative and passive sentences. In contrast, the thalamus was activated for passive sentences only. Previous studies identified the involvement of the basal ganglia in the processing of syntactic and morphological information [Moro et al., 2001; Ni et al., 2000; Ullman, 2001, 2004] but also phonological [Tettamanti et al., 2005] and lexical information [Crosson et al., 2003]. The present data are consistent with these previous findings concerning the role of basal ganglia in language processing and may further suggest that the basal ganglia are sensitive to reanalysis processes, which are thematic as well as syntactic in nature (evidenced by both causative and passive sentences). In contrast, the selective activation of the thalamus by passive sentences, that is, its activation by passive sentences only and not causative sentences, may imply that the thalamus has some involvement in syntactic reanalysis, or syntactic process in general, but no other types of linguistic processes. The active role of the thalamus in the processing of syntactic information has recently been reported in a study using intracranial recordings [Wahl et al., 2008]. It will be interesting to test further what specific role that the thalamus plays in sentence comprehension.
CONCLUSION
The present fMRI study is the first to provide direct evidence for a functional neuroanatomical basis for thematic reanalysis and its interplay with syntactic reanalysis in Japanese sentence comprehension. Thematic reanalysis activated the network comprising the left Broca's area, in particular, BA 45, and the left posterior STG, whereas syntactic reanalysis led to increased activation in BA 45 only. The data, furthermore, suggest that the time course of activation of BA 45 is driven by the nature of distinct reanalysis processes with thematic reanalysis preceding syntactic reanalysis in time, thereby functionally specifying the dynamics of different brain regions within the frontotemporal language network.
Acknowledgements
The authors thank Jens Brauer, Ina Bornkessel‐Schlesewsky, Takene Ito, and two anonymous reviewers for their helpful comments about the present work. Many thanks go to Derek V.M. Ott for his support carrying out the study, Petra Erz, Mandy Naumann, Domenica Wilfling, and Simone Wipper for data collection, and Andrea Gast‐Sandmann and Kerstin Flake for creating figures.
REFERENCES
- Altmann GTM ( 1999): Thematic role assignment in context. J Mem Lang 41: 124–145. [Google Scholar]
- Aoshima S, Phillips C, Weinberg A ( 2004): Processing filler‐gap dependencies in a head‐final language. J Mem Lang 51: 23–54. [Google Scholar]
- Badre D, Wagner AD ( 2007): Left ventrolateral prefrontal cortex and the control of memory. Neuropsychologia 45: 2883–2901. [DOI] [PubMed] [Google Scholar]
- Ben‐Shachar M, Hendler T, Kahn I, Ben‐Bashat D, Grodzinsky Y ( 2003): The neural reality of syntactic transformations: Evidence from functional magnetic resonance imaging. Psychol Sci 14: 433–440. [DOI] [PubMed] [Google Scholar]
- Ben‐Shachar M, Palti D, Grodzinsky Y ( 2004): Neural correlates of syntactic movement: Converging evidence from two fMRI experiments. Neuroimage 21: 1320–1336. [DOI] [PubMed] [Google Scholar]
- Bookheimer SY ( 2002): Functional MRI of Language: New approaches to understanding the cortical organization of semantic processing. Annu Rev Neurosci 25: 151–188. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Schlesewsky M ( 2006): The extended argument dependency model: A neurocognitive approach to sentence comprehension across languages. Psychol Rev 4: 787–821. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Schlesewsky M, Friederici AD ( 2002): Beyond syntax: Language‐related positivities reflect the revision of hierarchies. Neuroreport 3: 361–364. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Schlesewsky M, Friederici AD ( 2003): Eliciting thematic reanalysis effects: The role of syntax‐independent information during parsing. Lang Cognit Process 18: 268–298. [Google Scholar]
- Bornkessel I, Zysset S, Friederici AD, von Cramon DY, Schlesewsky M ( 2005): Who did what to whom? The neural basis of argument hierarchies during language comprehension. Neuroimage 26: 221–233. [DOI] [PubMed] [Google Scholar]
- Carlson G, Tanenhaus MK ( 1988): Thematic roles and language comprehension In: Wilkins W, editor. Syntax and Semantics, Vol. 21: Thematic Relations. New York: Academic Press; pp 263–288. [Google Scholar]
- Clifton C Jr, Traxler MJ, Mohamed MT, Williams RS, Morris RK, Rayner K ( 2003): The use of thematic role information in parsing: Syntactic processing autonomy revised. J Mem Lang 49: 317–334. [Google Scholar]
- Crosson B, Benefield H, Cato MA, Sadek JR, Moore AB, Wierenga CE, Gopinath K, Soltysik D, Bauer RM, Auerbach EJ, Gokcay D, Leonard CM, Briggs RW ( 2003): Left and right basal ganglia and frontal activity during language generation: Contributions to lexical, semantic, and phonological processes. J Int Neuropsychol Soc 9: 1061–1077. [DOI] [PubMed] [Google Scholar]
- Frisch S, Schlesewsky M ( 2001): The N400 reflects problems of thematic hierarchizing. Neuroreport 12: 3391–3394. [DOI] [PubMed] [Google Scholar]
- Frisch S, Schlesewsky M ( 2005): The resolution of case conflicts from a neurophysiological perspective. Cogn Brain Res 25: 484–498. [DOI] [PubMed] [Google Scholar]
- Friederici AD ( 2002): Towards a neural basis of auditory sentence processing. Trends Cogn Sci 6: 78–84. [DOI] [PubMed] [Google Scholar]
- Friederici AD ( 2006): The neural basis of sentence processing: Inferior frontal and temporal contributions In: Grodzinsky Y, Amunts K, editors. Broca's Region. Oxford: Oxford University Press; pp 196–217. [Google Scholar]
- Friederici AD, Steinhauer K, Pfeifer E ( 2002): Brain signatures of artificial language processing: evidence challenging the critical period hypothesis. Proc Natl Acad Sci U S A 99: 529–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederici AD, Wang Y, Herrmann CS, Maess B, Oertel U ( 2000): Localization of early syntactic processes in frontal and temporal cortical areas: A magnetoencephalographic study. Hum Brain Mapp 11: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederici AD, Rüschemeyer SA, Hahne A, Fiebach CJ ( 2003): The role of left inferior frontal and superior temporal cortex in sentence comprehension: Localizing syntactic and semantic processes. Cereb Cortex 13: 170–177. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Fiebach CJ, Schlesewsky M, Bornkessel ID, von Cramon YD ( 2006): Processing linguistic complexity and grammaticality in the left frontal cortex. Cereb Cortex 16: 1709–1717. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Fletcher P, Josephs O, Holmes A, Rugg MD, Turner R ( 1998): Event‐related fMRI: Characterizing differential responses. Neuroimage 7: 30–40. [DOI] [PubMed] [Google Scholar]
- Grewe T, Bornkessel I, Zysset S, Wiese R, von Cramon DY, Schlesewsky M ( 2005): The emergence of the unmarked: A new perspective on the language‐specific function of Broca's area. Hum Brain Mapp 26: 178–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grodzinsky Y ( 2000): The neurology of syntax: language use without Broca s area. Behav Brain Sci 23: 1–71. [DOI] [PubMed] [Google Scholar]
- Grodzinsky J, Friederici AD ( 2006): Neuroimaging of syntax and syntactic processing. Curr Opin Neurobiol 16: 240–246. [DOI] [PubMed] [Google Scholar]
- Hagoort P ( 2005): On broca, brain, and binding: A new framework. Trends Cogn Sci 9: 416–423. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Brown C, Groothusen J ( 1993): The syntactic positive shift (SPS) as an ERP measure of syntactic processing. Lang Cognit Process 8: 439–483. [Google Scholar]
- Herrmann B, Maess B, Hasting AS, Friederici AD ( 2009): Localization of the syntactic mismatch negativity in the temporal cortex: An MEG study. NeuroImage 48: 590–600. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D ( 2007): The cortical organization of speech processing. Nat Rev Neurosci 8: 393–402. [DOI] [PubMed] [Google Scholar]
- Hoshi H ( 1991): The generalized projection principle and its implications for passive constructions. J Japan Ling 13: 53–89. [Google Scholar]
- Hoshi H ( 1999): Passives In: Tsujimura N, editor. The Handbook of Japanese Linguistics. Malden: Blackwell; p 191–225. [Google Scholar]
- Josephs O, Turner R, Friston K ( 1997): Event‐related fMRI. Hum Brain Mapp 5: 243–248. [DOI] [PubMed] [Google Scholar]
- Kaan E, Swaab TY ( 2002): The brain circuitry of syntactic comprehension. Trends Cogn Sci 6: 350–356. [DOI] [PubMed] [Google Scholar]
- Kaan E, Swaab TY ( 2003): Repair, revision, and complexity in syntactic analysis: An electrophysiological differentiation. J Cogn Neurosci 15: 98–110. [DOI] [PubMed] [Google Scholar]
- Kamide Y, Mitchell DC ( 1999): Incremental pre‐head attachment in Japanese parsing. Lang Cognit Process 14: 631–662. [Google Scholar]
- Kamide Y, Altmann GTM, Haywood SL ( 2003): Prediction and thematic information in incremental sentence processing: Evidence from anticipatory eye movements. J Mem Lang 49: 133–156. [Google Scholar]
- Kinno R, Kawamura M, Shioda S, Sakai KL ( 2008): Neural correlates of non‐canonical syntactic processing revealed by a picture‐sentence matching task. Hum Brain Mapp 29: 1015–1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuperberg G ( 2007): Neural mechanisms of language comprehension: Challenges to syntax. Brain Res 1146: 23–49. [DOI] [PubMed] [Google Scholar]
- Kuperberg G, Sitnikova T, Caplan D, Holcomb PJ ( 2003): Electrophysiological distinctions in processing conceptual relationships within simple sentences. Cogn Brain Res 17: 117–129. [DOI] [PubMed] [Google Scholar]
- Kuperberg G, Caplan D, Sitnikova T, Eddy M, Holcomb PJ ( 2006): Neural correlates of processing syntactic, semantic, and thematic relationships in sentences. Lang Cognit Process 21: 489–530. [Google Scholar]
- Kuperberg G, Kreher DA, Sitnikova T, Caplan D, Holcomb PJ ( 2007): The role of animacy and thematic relationships in processing active English sentences: Evidence from eventrelated potentials. Brain Lang 100: 223–238. [DOI] [PubMed] [Google Scholar]
- Kuroda S‐Y ( 1979): On Japanese passives In: Bedell G, Kobayashi E, Muraki M, editors. Exloration in Linguistics: Papers in Honor of Kazuko Inoue. Tokyo: Kenkyusha; p 305–347. [Google Scholar]
- Kutas M, Federmeier KD ( 2000): Electrophysiology reveals semantic memory use in language comprehension. Trends Cogn Sci 4: 463–470. [DOI] [PubMed] [Google Scholar]
- Lee J, Garwood M, Menon R, Adriany G, Andersen P, Truwit C, Ugurbil K ( 1995): High contrast and fast three dimensional magnetic resonance imaging at high fields. Magn Reson Med 34: 308. [DOI] [PubMed] [Google Scholar]
- MacDonald MC, Pearlmutter NJ, Seidenberg MS ( 1994): The lexical nature of syntactic ambiguity resolution. Psychol Rev 101: 676–703. [DOI] [PubMed] [Google Scholar]
- Makuuchi M, Bahlmann J, Anwander A, Friederici AD ( 2009): Segregating the core computational faculty of human language from working memory. Proc Natl Acad Sci U S A 106: 8362–8367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazuka R, Itoh K ( 1995): Can Japanese speakers be led down the garden‐path? In: Mazuka R, Nagai N, editors. Japanese Sentence Processing. Hillsdale: Lawrence Erlbaum; p 295–329. [Google Scholar]
- McRae K, Ferretti TR, Amyote L ( 1997): Thematic roles as verb‐specific concepts. Lang Cognit Process 12: 137–176. [Google Scholar]
- Meyer M, Steinhauer K, Alter K, Friederici AD, von Cramon DY ( 2004): Brain activity varies with modulation of dynamic pitch variance in sentence melody. Brain Lang 89: 277–289. [DOI] [PubMed] [Google Scholar]
- Miyagawa S ( 1999): Causatives In: Tsujimura N, editor. The Handbook of Japanese Linguistics. Malden: Blackwell; p 236–268. [Google Scholar]
- Moro A, Tettamanti M, Perani D, Donati C, Cappa SF, Fazio F ( 2001): Syntax and the brain: Disentangling grammar by selective anomalies. NeuroImage 13: 110–118. [DOI] [PubMed] [Google Scholar]
- Ni W, Constable RT, Menci WE, Pugh KR, Fulbright RK, Shaywitz SE, Gore JC, Shankweiler D ( 2000): An event‐related neuroimaging study distinguishing form and content in sentence processing. J Cogn Neurosci 12: 120–133. [DOI] [PubMed] [Google Scholar]
- Newman SD, Just MA, Keller TA, Roth J, Carpenter PA ( 2003): Differential effects of syntactic and semantic processing on the subregions of Broca's area. Cogn Brain Res 16: 297–307. [DOI] [PubMed] [Google Scholar]
- Norris DG ( 2000): Reduced power multislice MDEFT imaging. Magn Reson 11: 445–451. [DOI] [PubMed] [Google Scholar]
- Oldfield RC ( 1971): The assessment and analysis of handedness. The Edinburgh inventory. Neuropsychologia 9: 97–113. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Holcomb PJ ( 1992): Event‐related brain potentials elicited by syntactic anomaly. J Mem Lang 31: 785–806. [Google Scholar]
- Poldrack RA, Temple E, Protopapas A, Nagarajan S, Tallal P, Merzenich M, Gabrieli JDE ( 2001): Relations between the neural bases of dynamic auditory processing and phonological processing: Evidence from fMRI. J Cogn Neurosci 13: 687–697. [DOI] [PubMed] [Google Scholar]
- Rayner K, Carlson M, Frazier L ( 1983): The interaction of syntax and semantics during sentence processing: Eye movements in the analysis of semantically biased sentences. J Verbal Learn Verbal Behav 22: 358–374. [Google Scholar]
- Rodd JM, Davis MH, Johnsrude IS ( 2005): The neural mechanism of speech comprehension: fMRI studies of semantic ambiguity. Cerebral Cortex 15: 1261–1269. [DOI] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y ( 2007a): Taxing working memory with syntax: Bi‐hemispheric modulations. Hum Brain Mapp 28: 1089–1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y ( 2007b): Working memory and syntax interact in Broca's area. Neuroimage 37: 8–17. [DOI] [PubMed] [Google Scholar]
- Scheepers C, Hemforth B, Konieczny L ( 2000): Linking syntactic functions with thematic roles: Psych‐verbs and the resolution of subject‐object ambiguity In: Hemforth B, Konieczny L, editors. German Sentence Processing. Dordrecht: Kluwer; p 95–135. [Google Scholar]
- Shibatani M. 1990. The Languages of Japan. Cambridge: Cambridge University Press. [Google Scholar]
- Talairach J, Tournoux P. 1988. Co‐Planar Stereotaxic Atlas of the Human Brain. Stuttgart: Thieme. [Google Scholar]
- Terada M. 1990. Incorporation and Argument Structure in Japanese. Amherst: Graduate Student Linguistics Association, University of Massachusetts Amherst. [Google Scholar]
- Tettamanti M, Buccino G, Saccuman M, Gallese V, Danna M, Scifo P ( 2005): Listening to action‐related sentences activates fronto‐parietal motor circuits. J Cogn Neurosci 17: 273–281. [DOI] [PubMed] [Google Scholar]
- Trueswell JC, Tanenhaus MK, Garnsey SM ( 1994): Semantic influences on parsing: Use of thematic role information in syntactic disambiguation. J Mem Lang 33: 285–318. [Google Scholar]
- Ullman MT ( 2001): A neurocognitive perspective on language: The declarative/procedural model. Nat Rev Neurosci 2: 717–726. [DOI] [PubMed] [Google Scholar]
- Ullman MT ( 2004): Contributions of memory circuits to language: The declarative/procedural model. Cognition 92: 231–270. [DOI] [PubMed] [Google Scholar]
- Vigneau M, Beaucousin V, Herve PY, Duffau H, Crivello F, Houde O, Mazoyer B, Tzourio‐Mazoyer N ( 2006): Meta‐analyzing left hemisphere language areas: Phonology, semantics, and sentence processing. Neuroimage 30: 1414–1432. [DOI] [PubMed] [Google Scholar]
- Wahl M, Marzinzik F, Friederici AD, Hahne A, Kupsch A, Schneider GH, Saddy D, Curio G, Klostermann F ( 2008): The role of the human thalamus in syntactic language processing. Neuron 59: 695–707. [DOI] [PubMed] [Google Scholar]
- Wartenburger I, Heekeren HR, Burchert F, Heinemann S, Bleser DR, Villringer A ( 2004): Neural correlates of syntactic transformations. Hum Brain Mapp 22: 72–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolff S, Schlesewsky M, Hirotani M, Bornkessel‐Schlesewsky I ( 2008): The neural mechanisms of word order processing revisited: Electrophysiological evidence from Japanese. Brain Lang 107: 133–157. [DOI] [PubMed] [Google Scholar]
- Worsley KJ, Friston KJ ( 1995): Analysis of fMRI time‐series revisited‐again. Neuroimange 2: 173–181. [DOI] [PubMed] [Google Scholar]