

Abstract
Brain imaging studies have identified a left‐lateralized network of regions that are engaged when monolinguals read. However, for individuals who are native speakers of two languages, it is unclear whether this pattern of activity is maintained across both languages or if it deviates according to language‐specific properties. We used functional magnetic resonance imaging to investigate single‐word processing in Spanish and in English in 12 proficient early Spanish–English bilinguals matched in skill level in both languages. Word processing in Spanish engaged the left inferior frontal and left middle temporal gyri. Word processing in English activated the left inferior frontal, middle frontal, and fusiform gyri extending to inferior temporal gyrus and the right middle temporal gyrus extending into superior temporal sulcus. The comparison of reading in Spanish greater than reading in English revealed involvement of the left middle temporal gyrus extending into the superior temporal sulcus. English greater than Spanish, however, demonstrated greater engagement of the left middle frontal gyrus extending into the superior frontal gyrus. We conclude that although word processing in either language activates classical areas associated with reading, there are language‐specific differences, which can be attributed to the disparity in orthographic transparency. English, an orthographically deep language, may require greater engagement of the frontal regions for phonological coding, whereas Spanish allows increased access to semantic processing via the left middle temporal areas. Together, these results suggest that bilinguals will show adjustments to the typical neural representation of reading as necessitated by the demands of the orthography. Hum Brain Mapp, 2012. © 2010 Wiley Periodicals, Inc.
Keywords: fMRI, bilingualism, orthography, proficiency, written language
INTRODUCTION
Two decades of research using brain imaging technology have shed light on the functional anatomy of word processing and reading. These studies have revealed a network of left‐lateralized regions in the occipitotemporal, temporoparietal, and frontal cortices that participate in aloud and silent reading [Fiez and Peterson,1998; Price et al.,1996; Pugh et al.,1996; Rumsey et al.,1997; Turkeltaub et al.,2002]. Based on these findings, Pugh et al. [Pugh et al.,2001; Sandak et al., 2004a] have devised a brain‐based model of reading focusing on a dorsal and ventral dichotomy within left hemisphere posterior systems as well as a left frontal region, each of which demonstrates changes in activation depending on the type of word being processed. Specifically, this model proposes that the occipitotemporal region, which contains the putative visual word form area [VWFA; Cohen et al.,2002; however, see Price and Devlin,2003], is principally responsible for word (real and pseudoword) identification [Pugh et al.,2001; Sandak et al., 2004b]. In response to frequently encountered words, this ventral area makes available the memory representations corresponding to a word's orthography. However, when a new word is encountered for which no orthographic representation exists in the occipitotemporal cortex, the word must be decoded, or “sounded out,” by the reader. The word is thus assigned to the temporoparietal region, which integrates orthographic, phonological, lexical, and semantic rules. This dorsal area, encompassing Wernicke's Wortschatz [Brunswick et al.,1999], shows increased activation during tasks invoking phonological processing [Démonet et al.,1992] and, compared to the occipitotemporal region, is considered to be relatively slower in its temporal onset [Pugh et al.,2001]. Access to the meaning of a word is thought to occur in the middle and inferior temporal gyri within this region [Pugh et al.,2005; Sandak et al., 2004a, 2004b]. A third participant in the process, the inferior frontal gyrus, which incorporates Broca's area, is proposed to provide more detailed analysis of word processing, supporting articulatory recoding, or “assembled phonology” [Pugh et al.,2001]. These regions are believed to act in concert, with the dorsal circuit enabling advancement of the ventral circuit as reading develops in typical development [Sandak et al., 2004a]. Specifically, during learning, the supramarginal gyrus (within the temporoparietal system) and the inferior frontal gyrus integrate orthographic and phonological features of words, whereas the angular gyrus (within the temporoparietal system) transfers these features to distributed cortical sites of semantic knowledge, allowing a “phonologically tuned” occipitotemporal skill zone to develop for functional pattern‐identification. Once learning of words improves, this region, in turn, relies on lexical and semantic processing in the middle and inferior temporal gyri to facilitate the emergence of rapid word identification in the ventral circuit. The degree to which this pattern of brain activity adheres when reading in different languages [Paulesu et al.,2000] or in nonalphabetic writing systems [Tan et al.,2000] has been an area of active research. Of particular interest is how language is organized in individuals who have more than one language at their disposal, either because they learned more than one language early on (i.e., no later than age 6) or because they acquired an additional language to a high level of proficiency later in life. The debate central to both lesion studies of neurological patients [Karanth,1981; Ohno,2002; Raman and Weekes,2005; Wechsler,1977] and imaging studies of multilingual healthy subjects [Perani and Abutalebi,2005] is the extent to which multiple languages spoken by an individual share cortical representation.
Most investigations examining the neural substrates responsible for representing first (L1) and second (L2) languages in late bilinguals have been conducted in the context of oral language skills rather than in the context of reading. Although many of these have used experimental approaches using written stimuli, to date, only one study has directly addressed the question of reading in bilinguals [Meschyan and Hernandez,2006]. This investigation provided insights into reading in Spanish and English. However, because the participants were not equally skilled in both languages (their proficiency in English was weaker on both the actual task and on a self‐report of language proficiency), the results on the brain's response to processing English versus Spanish words is somewhat open to interpretation due to differences in both orthography and proficiency. In the present investigation, we ask if different brain regions are used when an early bilingual reads words in Spanish and English under conditions in which proficiency on the task is matched and participants are matched on their language proficiency for both languages.
Evidence for a common cortical system for L1 and L2 exists at both the single‐word level [Chee et al.,1999,2000,2001; Illes et al.,1999; Tan et al., 1999,2003] and sentence‐processing level [Chee et al.1999; Yokoyama et al.,2006]. Together, these studies of written languages cover a range of paradigms that includes cued word generation, rhyme judgments, and semantic judgments. Other findings, however, suggest distinct patterns of activation for L1 and L2 [Luke et al.,2002; Meschyan and Hernandez,2006; Tham et al.,2005]. These conflicting results can be attributed to various factors. First, variability in age of L2 acquisition and L2 proficiency among subjects may account for these discrepancies [for review, see Perani and Abutalebi,2005]. Indeed, Wartenburger et al. [2003] found that whether L1 and L2 activate the same regions when bilinguals make semantic and syntactic judgments of visually presented sentences depends on L2 proficiency and age of L2 acquisition, respectively. Second, disparities in the orthographic transparency (i.e., consistency in mapping letters to sounds) of the languages tested may elucidate the discordant findings. Specifically, orthographies vary across languages in the consistency of their grapheme‐to‐phoneme correspondences. Pronunciation in German, for example, is very consistent, whereas the mapping from print to sound in English is not [Goswami et al.,2005]. These distinctions lead to differences in rate of reading acquisition in beginning readers [Goswami,2008; Ziegler and Goswami,2005] and have also been proposed as a source of between‐language variations in the functional anatomy of reading [Paulesu et al.,2000]. Third, disparities may be even more extensive for different writing systems. For example, Chinese uses a logographic or morphosyllabic writing system in which characters representing meaningful monosyllables serve as the basic units, whereas English has an alphabetic writing system in which letters indicating phonemes are the primary units [Tan et al.,2003]. Finally, variations in syntax may also explain the contradictory findings. As Luke et al. [2002] point out, English and Chinese vary significantly in syntax even though both are subject‐verb‐object languages. Thus, dissimilarities seen in the neural representations of English and Chinese may reflect the separate syntactic processing needed to read sentences in each language.
Although all of the aforementioned investigations used the visual presentation of words as an experimental paradigm, only one focused explicitly on the functional anatomy of reading rather than linguistic aspects of language processing. Meschyan and Hernandez [2006] studied subjects during a covert word‐reading task and reported that early Spanish–English bilinguals produced greater activation in the inferior parietal region and along the occiptoparietal border when reading words in English than when reading in Spanish. Spanish, on the other hand, elicited greater activation in the right supplementary motor area, right putamen, right insula, and left superior temporal gyrus. As noted above, subjects in this study differed in proficiency in and experience with Spanish and English. Consequently, whether the between‐group findings observed during the reading of English versus Spanish words were due to differences in orthography or due to discrepancies in proficiency and experience were not clear. Here, we build on this work by asking if individuals who are early users of Spanish and English map two written languages that use alphabetic writing systems with mainly shared Roman letters to the same brain regions despite the differences in the transparency of their orthographies. To allow for clear interpretation of any between‐language differences, it was required that subjects (1) had acquired both languages at an early age, (2) be matched on language and reading proficiency, and (3) perform the task equally well during the acquisition of scans. Only adults who had learned both English and Spanish at or before age 6 and were highly proficient in both languages were included in the study. Objective measures were used to confirm each subject's reading ability in both languages. An “implicit” reading task involving feature detection in words [Price et al.,1996; Turkeltaub et al.,2003] was used to identify the patterns of cortical activation for reading in English and Spanish. This paradigm has previously been shown to elicit brain activity in areas commonly seen in explicit reading paradigms, reflecting orthographic, phonological, and semantic processing [Price et al.,1996; Turkeltaub et al.,2003]. Importantly, this task has a distinct advantage for the study of bilingual reading in that the same task can be given in two languages and equivalent performance levels during the scanning session can be achieved.
Based on previous studies [Chee et al.,1999; Wartenburger et al.,2003], we predicted that reading by proficient early Spanish–English bilinguals in either language would invoke a similar cortical network consistent with that reported for reading in monolinguals of alphabetic languages in the literature [Pugh et al.,2001]. At the same time, we hypothesized that there would be language‐specific brain regions, reflecting the substantial differences in the orthographies of English and Spanish. A dual‐route cascaded (DRC) computational model of reading [Coltheart et al.,2001] purports that orthographic input can be processed via an indirect phonological route or via a direct lexical route to access semantics. Similarly, a computational connectionist model of reading [Harm and Seidenberg,2004] asserts that semantic activation occurs via visual and phonological pathways that normally contribute simultaneously but differentially, with division of labor dependent on various factors (e.g., skill acquisition, frequency, and consistency). If fewer resources for phonological processing are needed, as would be expected in Spanish because phonological mapping is more straightforward, there may be less use of this pathway and, at the same time, greater use of the semantic route. On the other hand, as English is orthographically deep, left temporoparietal and frontal regions that support phonological interpretation may show greater activation when subjects are processing English words compared with Spanish words.
METHODS
Subjects
Twelve healthy, right‐handed Spanish–English bilinguals (eight female, four male) aged 18–29 years (mean = 22.67 years) participated in this study. All subjects acquired both languages not later than age 6 and had minimal exposure to languages other than English and Spanish. None of the subjects reported a significant personal history of neurological impairment or learning disorders. Experimental procedures were approved by the Georgetown University Institutional Review Board; written informed consent was secured from each participant.
Behavioral Testing Battery
All subjects were given a battery of standard neuropsychological tests to measure single‐word reading skills in English and in Spanish as well as other cognitive skills that have been shown to provide an index of reading efficiency [Wagner and Torgesen,1987]. In English, the letter‐word identification and word attack subtests from the Woodcock–Johnson III Tests of Achievement [Woodcock et al.,2001] were used to generate the basic reading skills score, which represents a broader measure of reading that combines both sight word reading and decoding skills. The Batería III Woodcock‐Muñoz: Pruebas de aprovechamiento [Muñoz‐Sandoval et al.,2005] subtests of Identificacion de Letras y Palabras and Analisis de Palabras, the Spanish equivalents of the English tests, were used to provide the same measures in Spanish. Normal scores are considered to be in the range of 85–115, representing one standard deviation from the mean of 100.
In addition, to ensure that all study participants demonstrated a typical reading profile in English (i.e., no signs of a reading disability), specific cognitive aspects known to be related to reading proficiency in English were also evaluated [Wagner and Torgesen,1987]. Naming fluency (phonological recoding in lexical access) was determined by the rapid automatized naming test [Denckla and Rudel,1974,1976a,b], and phonetic recoding in auditory working memory was evaluated by the digit span subtest from the Wechsler Adult Intelligence Test, Third Edition (WAIS‐III) [Wechsler,1997]. Each participant also completed a language self‐assessment based on Meschyan and Hernandez [2006], including questions about exposure to both languages and current usage of Spanish and English (Table I). The Edinburgh Handedness Inventory [Oldfield,1971] was used to verify right‐handedness for each participant. Good visual acuity was confirmed for all subjects using the Sloan Letters Chart (Good‐Lite, Forest Park, IL).
Table I.
Language background and self‐assessment of subjects (n = 12)
| Parameter | Spanish (M, SD) | English (M, SD) | P value |
|---|---|---|---|
| Language background | |||
| Age of first exposure (years) | 0.0 (0.0) | 3.79 (2.21) | 9.67 × 10−5 |
| Formal study (years) | 12.96 (8.69) | 14.71 (4.79) | ns |
| Percent currently spoken per day | 22.92 (15.14) | 76.92 (15.00) | 6.65 × 10−5 |
| Language self‐assessment (Scale 1–7; 7=native‐like competence) | |||
| Listening comprehension | 6.75 (0.45) | 6.75 (0.45) | ns |
| Reading comprehension | 6.33 (0.78) | 6.67 (0.49) | ns |
| Speaking | 6.5 (0.52) | 6.67 (0.49) | ns |
| Writing | 6.08 (1.00) | 6.50 (0.80) | ns |
Functional Magnetic Resonance Imaging Data Acquisition
MRI images were acquired on a Siemens Vision Magnetom 3.0‐Tesla scanner with a circularly polarized head coil at Georgetown University's Center for Functional and Molecular Imaging. One three‐dimensional (3D) T1‐weighted image was obtained for every subject. Functional runs comprised an echo‐planar imaging (EPI) sequence (TR = 3.0 s, TE = 30 ms, flip angle = 90°, FOV = 192 mm, 64 × 64 matrix, 50 axial slices, interleaved and descending acquisition order, 3.0 mm × 3.0 mm × 2.8 mm cubic voxels).
Stimuli and Task
As reported in previous studies of reading [Turkeltaub et al.,2003,2004], and adapted from a study by Price et al. [1996], the experimental paradigm required subjects to view single words and press one of two buttons to indicate the presence (right thumb) or absence (left thumb) of an ascender, or tall letter, in each word (e.g., arroz, babor in Spanish; manor, vowel in English). Subjects also performed the same task on false font strings, or “pseudofonts.” As false font strings are associated with neither phonological nor semantic representations, contrasting activations for words with those for false font strings isolates the neural substrates specifically responsible for word processing from those concerned with nonlinguistic computations (e.g., motor planning, motor execution, response selection, visual processing, and spatial processing). As first described by Price at al. [1996], this experimental approach leads to the “implicit” or incidental activation of regions involved in orthography, phonology, and semantics but does not preferentially emphasize any of these aspects of reading.
Before being scanned, subjects were trained on the task. Each subject was scanned during four experimental runs (each run duration = 4 min, 27 sec), twice with English stimuli and twice with Spanish stimuli. Order of runs was counterbalanced across participants. Alternating blocks of words, false font strings, and fixation were presented in each run. Periods of crosshair fixation occurred at the beginning (24 s) and end (21 s) of each run and between (18 s) the other blocks. Stimulus presentation rate was 1 per 4.2 s (stimulus duration = 1.2 s, interstimulus duration = 3.0 s). For each condition, 40 whole‐head EPI volumes were compiled.
Stimuli consisted of 80 words (40 English, 40 Spanish) and 80 false font strings (40 English equivalents and 40 Spanish equivalents) presented in separate runs according to language. For the “words” condition, based on prior studies [Price et al., 1997; Turkeltaub et al.,2003], five‐letter, low‐frequency (English: KF = 8.05, SD = 6.03; Spanish: LEXESP = 7.95, SD = 5.16) words containing one or two syllables were shown individually in black Arial font. English stimuli were selected from the MRC Psycholinguistic Database [Coltheart,1981]; Spanish ones were chosen from the BuscaPalabras database [Davis and Perea,2005]. Cognates, words with the letters i or j, and words containing accents were excluded to eliminate ambiguity.
For the “false font strings” condition, unfamiliar characters in modified black Arial font were matched to words for length and position of ascenders (e.g., l, t, b) and descenders, or hanging letters, (e.g., y, p, g). Each English word had a false font counterpart, as did each Spanish word. To prevent subjects from assigning phonological meaning to false font strings, a strict correspondence between false font characters and letters was not applied, however (i.e., no false font character represented a unique letter). Ascenders were located in half of the stimuli.
Functional Magnetic Resonance Imaging Data Analysis
MRI images were analyzed with MEDx (Sensor Systems). After being motion‐corrected and globally normalized, data underwent Gaussian spatial smoothing (8 mm), high‐pass temporal filtering (240 s), and spatial normalization to the SPM99 MNI‐205 coordinate system [Woods et al.,1998a]. Using automated image registration, head motion correction was completed on scans [Woods et al.,1998b]; a cutoff of 0.6 mm (20% of voxel size) for 3D intrascan translation was used to exclude any scan in which excessive movement was found. To generate statistical maps of task‐related signal changes for every subject, the “real‐word” condition and corresponding “false‐font strings” condition were contrasted for each run. The maps were then rated by two blind evaluators for overall image quality and head motion. The better of the two runs acquired was submitted for group analysis. Random‐effects group analyses were then conducted to generate statistical maps of word processing in: (1) Spanish, (2) English, (3) Spanish greater than English, and (4) English greater than Spanish. As in previous reports using this task, for all statistical analyses Z‐maps were generated with a threshold of Z > 3.1 (P < 0.001) and 25 contiguous voxels [Turkeltaub et al.,2003]. The statistical maps of word processing in English and Spanish were overlaid to generate a colocalization map to determine common areas of activation underlying Spanish and English word processing. Brain renderings for figures were created with VolView 2.0. The brain was made transparent to allow activation to be seen below the surface.
RESULTS
Behavioral Results
All behavioral measures indicate that the subjects were equally proficient in English and Spanish. Subjects' self‐reported language skills did not vary significantly between the two languages (Table I). Although subjects indicate more current use of English than Spanish, their overall language experience, as signified by formal years of study, was similar in Spanish and English. Neuropsychological testing confirmed this: As shown in Table II, the basic reading skills cluster scores did not significantly differ between languages. In‐scanner data showed that subjects performed the feature detection task equally well in both languages (Table III).
Table II.
Neuropsychological measures of subjects' reading ability
| Neuropsychological measure | Spanish (M, SD) | English (M, SD) | P value |
|---|---|---|---|
| Basic Reading Skills Cluster | 114.83 (16.25) | 104 (7.03) | ns |
| Digit Span | 113.33 (17.23) | ||
| RAN Letters and Numbers | 110.25 (11.09) | ||
| RAN Colors and Objects | 100.17 (10.19) |
Basic reading skills cluster is a composite score based on word ID and word attack performance. P <0.05.
RAN = Rapid Automatized Naming. RH = right‐handed.
Table III.
Performance inside scanner during implicit reading task (word – false font string)
| Parameter | Spanish (M, SD) | English (M, SD) | P value |
|---|---|---|---|
| Overall accuracy (% correct) | 91.04 (7.94) | 91.46 (5.59) | ns |
| Overall response time (ms) | 751.58 (78.98) | 726.52 (74.70) | ns |
| Word / false font accuracy difference (% correct) | 2.92 (8.65) | −0.42 (3.96) | ns |
| Word / false font response time difference (ms) | −37.40 (60.29) | −13.11 (38.87) | ns |
Functional Magnetic Resonance Imaging Results
Within Group Comparisons
Spanish
Regions involved in word processing in Spanish (i.e., contrast between Spanish words and Spanish‐matched false font strings) are shown in Figure 1A and Table IV. Consistent with prior results in monolinguals, reading in Spanish engaged the left middle temporal gyrus (BA 21) and left inferior frontal gyrus (BA 45/47).
Figure 1.
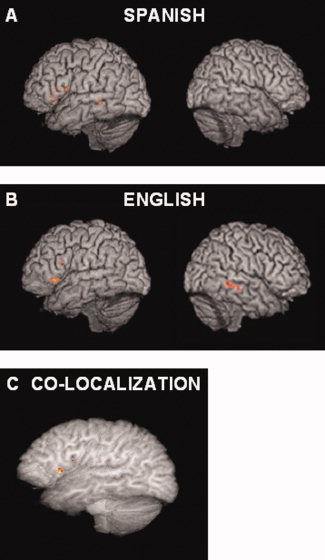
Individual languages. (A) Word processing in Spanish elicited activation in the left middle temporal and inferior frontal gyri (Z > 3.1, P < 0.001). (B) Word processing in English engaged left hemisphere fusiform gyrus extending to inferior temporal gyrus as well as inferior frontal and middle frontal gyri. In the right hemisphere, activation was seen in middle temporal gyrus extending into superior temporal sulcus. (C) Word processing in both Spanish and English activated an eight‐voxel cluster in the left inferior frontal gyrus. Brain renderings created with VolView 2.0 used templates with transparency values on a scale on which 0 equals complete transparency and 100 equals complete opacity. For (A) and (B), images were set to a transparency value of 75. For (C), a transparency value of 55 was used.
Table IV.
Activation during implicit reading task (word – false font string)
| Region | Brodmann area | Number of voxels | Max Z | Talairach coordinates | ||
|---|---|---|---|---|---|---|
| x | y | z | ||||
| Spanish | ||||||
| Left | ||||||
| Middle temporal gyrus | 21 | 60 | 3.67 | −64 | −34 | −2 |
| Inferior frontal gyrus | 45, 47 | 295 | 4.13 | −52 | 26 | 8 |
| English | ||||||
| Left | ||||||
| Fusiform gyrus/Inferior temporal gyrus | 37, 20 | 31 | 3.69 | −48 | −56 | −26 |
| Inferior frontal gyrus/Middle frontal gyrus | 9, 46 | 55 | 3.58 | −46 | 14 | 26 |
| Inferior frontal gyrus | 45, 46 | 65 | 3.81 | −58 | 22 | 12 |
| Inferior frontal gyrus | 47 | 98 | 4.33 | −50 | 32 | −10 |
| Right | ||||||
| Middle temporal gyrus/Superior temporal gyrus | 21, 22 | 137 | 3.69 | 72 | −32 | −4 |
| English and Spanish | ||||||
| Left | ||||||
| Inferior frontal gyrus | 47 | 8 | 3.87 | −50 | 30 | −8 |
| Spanish > English | ||||||
| Left | ||||||
| Middle temporal gyrus/Superior temporal sulcus | 21, 22 | 36 | 3.85 | −50 | −40 | 6 |
| Inferior colliculus | N/A | 25 | 3.38 | −8 | −30 | −4 |
| English > Spanish | ||||||
| Left | ||||||
| Middle frontal gyrus/Superior frontal gyrus | 10 | 46 | 3.50 | −32 | 54 | 4 |
Threshold of Z > 3.1 (P < 0.001) and 25 contiguous voxels
English
Word processing in English‐activated regions in the left hemisphere fusiform gyrus (BA 37/20) extending into the inferior temporal gyrus (Fig. 1B, Table IV). Other areas included the inferior frontal gyrus (BA 45/46/47) extending into the middle frontal gyrus as well as additional regions in the inferior frontal gyrus (BA 45, 46, and 47). Right‐hemispheric regions consisted of middle temporal gyrus extending into the superior temporal sulcus (BA 21/22).
Colocalization of Spanish and English
Common involvement in word processing in Spanish (i.e., contrast between Spanish words and Spanish‐matched false font strings) and English is indicated in Figure 1C and Table IV. Both languages activated a cluster in the left inferior frontal gyrus (BA 47).
Between Group Comparisons
Spanish Greater than English
The contrast of Spanish greater than English revealed relatively more activation for Spanish in the left middle temporal gyrus extending into the superior temporal sulcus (BA 21/22; Fig. 2, Table IV) as well as the inferior colliculus.
Figure 2.
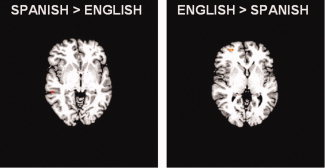
Comparisons. Contrast of Spanish greater than English showed activation in the left middle temporal gyrus extending into the superior temporal sulcus (Z > 3.1, P < 0.001). The opposite comparison, English greater than Spanish, found more engagement of the left middle frontal gyrus extending into the superior frontal gyrus.
English Greater than Spanish
The comparison of English greater than Spanish yielded relatively greater activity for English in the left middle frontal gyrus extending into the superior frontal gyrus (BA 10; Fig. 2, Table IV).
DISCUSSION
This study investigated single‐word reading in proficient early Spanish–English bilinguals to determine the functional anatomy of reading in each language. The results demonstrate that while both languages engage left hemisphere temporal and frontal cortices, word processing of Spanish and English rely on partially distinct brain regions. Because the participants were early bilinguals with equal language proficiency and comparable reading scores in both languages and because performance on the experimental task was the same for Spanish and English, these differences in activation patterns can be attributed to the nature of the orthographies of these two languages.
To interpret these language‐specific differences, the impact of orthographic transparency on reading must be considered within models that describe the roles of phonological and semantic processing in response to the orthographic information that is presented. By only examining reading at the single‐word level, we averted any potential effects of differing syntactic processing between languages. English has a deep orthography (i.e., the mapping between graphemes and phonemes is not consistent), whereas Spanish has a shallow orthography. The comparison between word processing in Spanish versus word processing in English revealed greater activation for Spanish in the left middle temporal gyrus that extends into the superior temporal sulcus (BA 21/22) and the inferior colliculus, a region involved in auditory processing. Meschyan and Hernandez (2006) also detected enhanced activity for Spanish relative to English words in a left superior temporal gyrus (BA22) region near the one reported presently and concluded that this finding is driven by orthographic transparency of reading in Spanish. However, in this study the maxima of the focus emanates from the left middle temporal gyrus, suggesting that this more inferior region may play a greater role in the reading of Spanish compared with English. Differences in the tasks used by this study and by Meschyan and Hernandez [2006] may account for this discrepancy in the results. The superior temporal gyrus may be more engaged if a covert reading task, like the one used by Meschyan and Hernandez, evokes associated acoustic properties.
The relatively increased recruitment of the left middle temporal gyrus for Spanish observed in this study may be due to this region's involvement in semantic processing. Support for this interpretation is offered by Price and Mechelli [2005] in their review of the literature, which reveals that middle temporal gyrus activity is greater during semantic processing than phonological processing. For example, a study of sentence comprehension by Dapretto and Bookheimer [1999] found the left middle temporal gyrus (BA 21) to be engaged in the semantic condition (single‐word meaning) but not the syntactic condition. In a meta‐analysis on semantic processing of words, Binder et al. [2009] found the left middle temporal gyrus to be a key component of the semantic network. One possible explanation is that because Spanish is currently used less often by the participants, difficulty in recruiting semantic representations could have resulted in greater activation in this area. However, Katz et al. [2005] found stable activation in the middle temporal gyrus extending into the superior temporal gyrus for both repeated and new words in lexical and naming tasks, indicating that engagement of this region does not depend on vocabulary and therefore making this interpretation less likely. Our results instead suggest that as Spanish is more orthographically transparent than English, resources for semantic coding are more readily accessible. This idea is consistent with models of reading, such as the DRC computational model [Coltheart et al.,2001], which describes orthographic input proceeding to either phonological output or semantics, and the computational connectionist model of reading [Harm and Seidenberg,2004], which contends that visual and phonological routes concurrently but unequally contribute to semantic activation. Hence, the orthographic transparency of Spanish may enable semantic processing to occur more easily in parallel because fewer resources for phonological processing are needed.
During the reverse comparison, English greater than Spanish, we found that English makes greater demands on the left middle and superior frontal gyri. We believe that this finding is compatible with this region's role in phonological mapping, which is invoked more deeply in English, a language replete with orthographic inconsistencies. Numerous experiments with English monolingual readers have demonstrated the role of the left inferior frontal cortex in tasks that require explicit mapping of graphemes to phonemes, including those involving pseudoword reading [Brunswick et al.,1999; Burton et al.,2005; Fiez et al.,1999; Hagoort et al.,1999; Herbster et al.,1997; Poldrack et al.,1999; Pugh et al.,1996; Rumsey et al.,1997; Zurowski et al.,2002]. Although the region identified in our study is more superior to the area observed in these studies, the same region has been shown to be responsive to the orthography of words. Specifically, in an event‐related functional magnetic resonance imaging study of visually presented English words, Binder et al. [2003] investigated lexicality and manipulated orthographic neighborhood size. They found that the left middle frontal gyrus (BAs 10/9, 6 and 8) showed a significant interaction of lexicality and neighborhood. This region was more engaged for words without any neighbors than words with many neighbors. In other words, activation in this region increased with orthographic complexity, which is consistent with our finding of this region's greater involvement in English than Spanish. In another study, Bolger et al. [2008] showed that the left middle frontal gyrus was more involved when words with greater phonological inconsistency were presented in a rhyming task, a task that entails direct phonological processing. As the left middle frontal gyrus in monolingual readers has heightened sensitivity to phonologically inconsistent words, one might anticipate that this region is more engaged when a bilingual reader is operating in the language with the deeper orthography.
Turning to ventral cortex, the occipitotemporal region, near the putative VWFA, was engaged during the reading of English words but not during the reading of Spanish words. According to the model proposed by Pugh et al. [2001], activation of the ventral circuit, which includes the occipitotemporal region, is frequency‐dependent. That is, the ventral stream shows greater engagement for familiar, high‐frequency words. As we selected low‐frequency words in both languages, it is somewhat surprising that the occipitotemporal region was even activated above threshold in the English condition, as this finding has not been reported in previous studies using this implicit reading task [Turkeltaub et al2003; Price et al,1996]. A possible explanation for this activation in the current sample is that in bilinguals, the presence of word form representations in Spanish affects the perceived frequency of words in English, the written language currently used more extensively, thereby resulting in activations in the left occipitotemporal region as if the words were of low frequency in nature.
The current study did not replicate the finding of right inferior parietal lobe seen in the English versus Spanish contrast by Meschyan and Hernandez [2006]. The authors argued that the disparity in orthographic transparency of the subjects' first (Spanish) and second (English) languages accounted for the differential activation in this region. It is noteworthy that Wartenburger et al. [2003] also reported a similar right hemisphere difference when comparing two languages in bilinguals. However, they interpreted these findings in terms of language proficiency, not orthography. In that study, late Italian (L1) – German (L2) bilinguals highly proficient in L2 showed greater activation in the right inferior parietal lobe when compared to late Italian (L1) – German (L2) bilinguals with low L2 proficiency. As both Italian and German are orthographically shallow, this result cannot be attributed to an orthography effect. As the participants in the present investigation were equally proficient in both languages, the absence of right hemisphere differences in the English greater than Spanish contrast is therefore not surprising. Together, these studies suggest that the findings reported by Meschyan and Hernandez in the right parietal cortex are more likely due to the differences in proficiency in their participants rather than the orthographic characteristics of English and Spanish.
The present investigation also did not reproduce other right hemispheric findings of the Meschyan and Hernandez [2006] study. Meschyan and Hernandez found greater engagement of the right insula and putamen for Spanish versus English. The authors concluded that this greater activation for Spanish resulted from being less proficient in that language. As our subjects were equally proficient in both languages, the lack of activation in these regions in our results is not unexpected.
Previous studies on bilingualism have focused specifically on the age of acquisition of language, particularly the second language. Consistent with prior studies [e.g., Chee et al.1999], our investigation used a cutoff age of 6 years as an inclusion criterion to classify participants as “early” bilinguals. In the end, however, the mean age of acquisition of each language did differ. We do not believe that this plays a significant role in the outcome of our study. To date, studies on age of acquisition differences have focused on individuals who acquired L2 at a much later age. For example, the mean age of initial exposure for early bilinguals and late bilinguals in Kim et al. [1997] and Wartenburger et al. [2003] differed, respectively, by 11.2 and 18.9 years, both significantly larger differences than that reported by subjects in this study. Further, Wartenburger et al. [2003] found that age of acquisition plays a role in grammatical, but not semantic, processing. As single‐word processing, the target of the present study, is influenced by semantics rather than grammar, age of acquisition should not have affected the results. Moreover, written language is typically taught around age 6, making this issue less likely to explain any between‐group findings.
Similarly, the role of language proficiency has been considered to play an important role vis‐à‐vis colocalization of two languages in bilinguals. Although subjects in this study currently spend the majority of their day speaking English, they are equally proficient in both languages as indicated in their self‐assessments. Objective standardized measures of reading established not only that they were good readers but also that they had equivalent reading skills in both languages. These measures make it possible to conclude that language proficiency is not the reason for the differences observed during reading of Spanish and English. Although standard neuropsychological tests confirmed that subjects were equally proficient in English and Spanish on measures of reading, the tests available in both English and Spanish were limited to these domains and did not include higher‐level skills, such as comprehension and vocabulary. Given the present findings, which suggest facilitated semantic processing for Spanish words, future studies would benefit from expanding language‐based testing in both languages.
Finally, the current study contributes to the bilingual imaging literature by extending the debate of cortical representation to reading. As reviewed in Perani and Abutalebi [2005], the predominant view holds that a common neural network underlies L1 and L2 processing and that differences in activation during language tasks are related to disparities in age of acquisition, proficiency, and linguistic features between languages. Although many investigations have contributed results in support of this hypothesis, nearly all focused on the spoken modality even though the paradigms involved written stimuli. This study therefore offers a novel contribution in that it demonstrates specifically that single‐word reading in proficient early bilinguals engages a shared cortical system within which activation is modulated by orthography. As subjects had early acquisition of and matched proficiency in two languages with similar writing systems, involvement of partially distinct brain regions during reading can be attributed to differences in written language (versus spoken language). The development of a similar neural system for reading in early bilinguals proficient in both alphabetic and nonalphabetic writing systems remains to be seen.
In summary, this investigation of single‐word reading in proficient early bilinguals found that while both languages engage the cortical network previously reported for reading in monolinguals, there are language‐specific differences. Spanish, an orthographically transparent language, elicits greater involvement of the left middle temporal gyrus and superior temporal sulcus, possibly due to demands on semantic processing. English, an orthographically opaque language, yields more activity in the left middle and superior frontal gyri, which have previously been shown to play a role in phonological processing. Overall, this study reveals the presence of language‐selective representations of reading in proficient early bilinguals who are matched on proficiency in both languages and relates this finding to the intrinsic orthographic properties of the two languages.
Acknowledgements
The authors would like to thank Virginia Spaventa and Lynn Flowers for recording instructions in Spanish and discussing criteria for subject selection, Natasha Tokowicz for offering insight about the study design, and Hima Ekanadham for creating the language self‐assessment questionnaire. The authors also appreciate the help of Emma Cole, Joe Maisog, and Jenni Rosenberg and the input of the two anonymous reviewers.
REFERENCES
- Binder JR, Desai RH, Graves WW, Conant LL ( 2009): Where is the semantic system? A critical review and meta‐analysis of 120 functional neuroimaging studies. Cereb Cortex 19: 2767–2796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder JR, McKiernan KA, Parsons ME, Westbury CF, Possing ET, Kaufman JN, Buchanan L ( 2003): Neural correlates of lexical access during visual word recognition. J Cogn Neurosci 15: 372–393. [DOI] [PubMed] [Google Scholar]
- Bolger DJ, Hornickel J, Cone NE, Burman DD, Booth JR ( 2008): Neural correlates of orthographic and phonological consistency effects in children. Hum Brain Mapp 29: 1416–1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunswick N, McCrory E, Price CJ, Frith CD, Frith U ( 1999): Explicit and implicit processing of words and pseudowords by adult developmental dyslexics: A search for Wernicke's Wortschatz? Brain 122: 1901–1917. [DOI] [PubMed] [Google Scholar]
- Burton MW, Locasto PC, Krebs‐Noble D, Gullapalli RP ( 2005): A systematic investigation of the functional neuroanatomy of auditory and visual phonological processing. NeuroImage 26: 647–661. [DOI] [PubMed] [Google Scholar]
- Chee MW, Caplan D, Soon CS, Sriram N, Tan EW, Thiel T, Weekes B ( 1999): Processing of visually presented sentences in Mandarin and English studied with fMRI. Neuron 23: 127–137. [DOI] [PubMed] [Google Scholar]
- Chee MW, Hon N, Lee HL, Soon CS ( 2001): Relative language proficiency modulates BOLD signal change when bilinguals perform semantic judgments. Blood oxygen level dependent. NeuroImage 13: 1155–1163. [DOI] [PubMed] [Google Scholar]
- Chee MW, Tan EW, Thiel T ( 1999): Mandarin and English single word processing studied with functional magnetic resonance imaging. J Neurosci 19: 3050–3056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chee MW, Weekes B, Lee KM, Soon CS, Schreiber A, Hoon JJ, Chee M ( 2000): Overlap and dissociation of semantic processing of Chinese characters, English words, and pictures: Evidence from fMRI. NeuroImage 12: 392–403. [DOI] [PubMed] [Google Scholar]
- Cohen L, Lehéricy S, Chochon F, Lemer C, Rivaud S, Dehaene S ( 2002): Language‐specific tuning of visual cortex? Functional properties of the Visual Word Form Area. Brain 125: 1054–1069. [DOI] [PubMed] [Google Scholar]
- Coltheart M ( 1981). The MRC Psycholinguistic Database. Quart J Exp Psychol A 33: 497–505. [Google Scholar]
- Coltheart M, Rastle K, Perry C, Langdon R, Ziegler J ( 2001): DRC: A dual route cascaded model of visual word recognition and reading aloud. Psychol Rev 108: 204–256. [DOI] [PubMed] [Google Scholar]
- Dapretto M, Bookheimer SY ( 1999): Form and content: dissociating syntax and semantics in sentence comprehension. Neuron 24: 427–432. [DOI] [PubMed] [Google Scholar]
- Davis CJ, Perea M ( 2005): BuscaPalabras: A program for deriving orthographic and phonological neighborhood statistics and other psycholinguistic indices in Spanish. Behav Res Methods 37: 665–671. [DOI] [PubMed] [Google Scholar]
- Démonet JF, Chollet F, Ramsay S, Cardebat D, Nespoulous JL, Wise R, Rascol A, Frackowiak R ( 1992): The anatomy of phonological and semantic processing in normal subjects. Brain 115: 1753–1768. [DOI] [PubMed] [Google Scholar]
- Denckla MB, Rudel R ( 1974): Rapid “automatized” naming of pictured objects, colors, letters and numbers by normal children. Cortex 10: 186–202. [DOI] [PubMed] [Google Scholar]
- Denckla MB, Rudel RG ( 1976a): Naming of object‐drawings by dyslexic and other learning disabled children. Brain Lang 3: 1–15. [DOI] [PubMed] [Google Scholar]
- Denckla, MB , Rudel, RG ( 1976b): Rapid “automatized” naming (R.A.N): Dyslexia differentiated from other learning disabilities. Neuropsychologia 14: 471–479. [DOI] [PubMed] [Google Scholar]
- Dijkstra T ( 2005): Bilingual word recognition and lexical access In: Kroll JF, De Groot AMB, editors. Handbook of Bilingualism: Psycholinguistic Approaches. New York: Oxford University Press; pp 179–201. [Google Scholar]
- Fabbro F, Skrap M, Aglioti S ( 2000): Pathological switching between languages after frontal lesions in a bilingual patient. J Neurol Neurosurg Psychiatry 68: 650–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiez, JA , Balota DA, Raichle M E, Petersen SE ( 1999): Effects of lexicality, frequency, and spelling‐to‐sound consistency on the functional anatomy of reading. Neuron 24: 205–218. [DOI] [PubMed] [Google Scholar]
- Fiez, JA , Petersen SE ( 1998): Neuroimaging studies of word reading. Proc Natl Acad Sci USA 95: 914–921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goswami U ( 2008): The development of reading across languages. Ann NY Acad Sci 1145: 1–12. [DOI] [PubMed] [Google Scholar]
- Goswami U, Ziegler JC, Richardson U ( 2005): The effects of spelling consistency on phonological awareness: a comparison of English and German. J Exp Child Psychol 92: 345–365. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Indefrey P, Brown C, Herzog H, Steinmetz H, Seitz RJ ( 1999): The neural circuitry involved in the reading of German words and pseudowords: A PET study. J Cogn Neurosci 11: 383–398. [DOI] [PubMed] [Google Scholar]
- Harm MW, Seidenberg MS ( 2004): Computing the meanings of words in reading: Cooperative division of labor between visual and phonological processes. Psychol Rev 111: 662–720. [DOI] [PubMed] [Google Scholar]
- Herbster AN, Mintun MA, Nebes RD, Becker JT ( 1997): Regional cerebral blood flow during word and nonword reading. Hum Brain Mapp 5: 84–92. [DOI] [PubMed] [Google Scholar]
- Illes J, Francis WS, Desmond JE, Gabrieli JD, Glover GH, Poldrack R, Lee CJ, Wagner AD ( 1999): Convergent cortical representation of semantic processing in bilinguals. Brain Lang 70: 347–363. [DOI] [PubMed] [Google Scholar]
- Jared D, Kroll JF ( 2001): Do bilinguals activate phonological representations in one or both of their languages when naming words? J Mem Lang 44: 2–31. [Google Scholar]
- Karanth P ( 1981): Pure alexia in a Kannada‐English bilingual. Cortex 17: 187–198. [DOI] [PubMed] [Google Scholar]
- Katz L, Lee CH, Tabor W, Frost SJ, Mencl WE, Sandak R, Rueckl J, Pugh KR ( 2005): Behavioral and neurobiological effects of printed word repetition in lexical decision and naming. Neuropsychologia 43: 2068–2083. [DOI] [PubMed] [Google Scholar]
- Kim KHS, Relkin NR, Lee K‐M, Hirsch J ( 1997): Distinct cortical areas associated with native and second languages. Nature 388: 171. [DOI] [PubMed] [Google Scholar]
- Kroll JF, Bobb S, Wodniecka Z ( 2006): Language selectivity is the exception, not the rule: Arguments against a fixed locus of language selection in bilingual speech. Biling Lang Cognit 9: 119–135. [Google Scholar]
- Luke K, Liu H, Wai Y, Wan Y, Tan LH ( 2002): Functional anatomy of syntactic and semantic processing in language comprehension. Hum Brain Mapp 16: 133–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marian V, Spivey M, Hirsch J ( 2003): Shared and separate systems in bilingual language processing: converging evidence from eyetracking and brain imaging. Brain Lang 86: 70–82. [DOI] [PubMed] [Google Scholar]
- Meschyan G, Hernandez AE ( 2006): Impact of language proficiency and orthographic transparency on bilingual word reading: An fMRI investigation. NeuroImage 29: 1135–1140. [DOI] [PubMed] [Google Scholar]
- Muñoz‐Sandoval AF, Woodcock RW, McGrew KS, Mather N ( 2005): Batería III Woodcock‐Muñoz: Pruebas de aprovechamiento. Itasca, IL: The Riverside Publishing Company. [Google Scholar]
- Ohno T, Takeda K, Kato S, Hirai S ( 2002): Pure alexia in a Japanese‐English bilingual: Dissociation between the two languages. J Neurol 249: 105–107. [DOI] [PubMed] [Google Scholar]
- Oldfield RC ( 1971): The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 9: 97–113. [DOI] [PubMed] [Google Scholar]
- Paulesu E, McCrory E, Fazio F, Menoncello, L , Brunswick N, Cappa SF, Cotelli M, Cossu G, Corte F, Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith CD, Frith U ( 2000): A cultural effect on brain function. Nat Neurosci 3: 91–96. [DOI] [PubMed] [Google Scholar]
- Perani D, Abutalebi J ( 2005): The neural basis of first and second language processing. Curr Opin Neurobiol 15: 202–206. [DOI] [PubMed] [Google Scholar]
- Poldrack RA, Wagner AD, Prull MW, Desmond JE, Glover GH, Gabrieli JD ( 1999): Functional specialization for semantic and phonological processing in the left inferior prefrontal cortex. NeuroImage 10: 15–35. [DOI] [PubMed] [Google Scholar]
- Price CJ, Devlin JT ( 2003): The myth of the visual word form area. NeuroImage 19: 473–481. [DOI] [PubMed] [Google Scholar]
- Price CJ, Green DW, von Studnitz R ( 1999): A functional imaging study of translation and language switching. Brain 122: 2221–2235. [DOI] [PubMed] [Google Scholar]
- Price CJ, Mechelli A ( 2005): Reading and reading disturbance. Curr Opin Neurobiol 15: 231–238. [DOI] [PubMed] [Google Scholar]
- Price CJ, Wise RJ, Frackowiak RS ( 1996): Demonstrating the implicit processing of visually presented words and pseudowords. Cereb Cortex 6: 62–70. [DOI] [PubMed] [Google Scholar]
- Pugh KR, Mencl WE, Jenner AR, Katz L, Frost SJ, Lee JR, Shaywitz SE, Shaywitz BA ( 2001): Neurobiological studies of reading and reading disability. J Commun Disord 34: 479–492. [DOI] [PubMed] [Google Scholar]
- Pugh KR, Sandak R, Frost SJ, Moore D, Mencl WE ( 2005): Examining reading development and reading disability in English language learners: Potential contributions from functional neuroimaging. Learn Disabil Res Pract 20: 24–30. [Google Scholar]
- Pugh KR, Shaywitz BA, Shaywitz SE, Constable RT, Skudlarski P, Fulbright RK, Bronen RA, Shankweiler DP, Katz L, Fletcher JM, Gore JC ( 1996): Cerebral organization of component processes in reading. Brain 119: 1221–1238. [DOI] [PubMed] [Google Scholar]
- Raman I, Weekes BS ( 2005): Acquired dyslexia in a Turkish‐English speaker. Ann Dyslexia 55: 79–104. [DOI] [PubMed] [Google Scholar]
- Rumsey JM, Horwitz B, Donohue BC, Nace K, Maisog JM, Andreason P ( 1997): Phonological and orthographic components of word recognition. A PET‐rCBF study. Brain 120: 739–759. [DOI] [PubMed] [Google Scholar]
- Sandak R, Mencl WE, Frost SJ, Pugh KR ( 2004): The neurobiological basis of skilled and impaired reading: Recent findings and new directions. Sci Stud Read 8: 273–292. [Google Scholar]
- Sandak R, Mencl WE, Katz L, Moore DL, Pugh KR ( 2004): The neurobiology of adaptive learning in reading: A contrast of different training conditions. Cogn Affect Behav Neurosci 4: 67–88. [DOI] [PubMed] [Google Scholar]
- Schwartz AI, Kroll JF ( 2006): Bilingual lexical activation in sentence context. J Mem Lang 55: 197–212. [Google Scholar]
- Talairach J, Tournoux P ( 1998): Co‐Planar Stereotaxic Atlas of the Human Brain. Stuttgart, Germany: Thieme. [Google Scholar]
- Tan LH, Spinks JA, Feng C, Siok WT, Perfetti CA, Xiong J, Fox PT, Gao JH ( 2003): Neural systems of second language reading are shaped by native language. Hum Brain Mapp 18: 158–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Spinks JA, Gao JH, Liu HL, Perfetti CA, Xiong J, Stofer KA, Pu Y, Liu Y, Fox PT ( 2000): Brain activation in the processing of Chinese characters and words: A functional MRI study. Hum Brain Mapp 10: 16–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tham WWP, Rickard Liow SJ, Rajapakse JC, Choong Leong, T , Ng SES, Lim WEH, Ho LG ( 2005): Phonological processing in Chinese‐English bilingual biscriptals: An fMRI study. NeuroImage 28: 579–587. [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA ( 2002): Meta‐analysis of the functional neuroanatomy of single‐word reading: Method and validation. NeuroImage 16: 765–780. [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF ( 2003): Development of neural mechanisms for reading. Nat Neurosci 6: 767–73. [DOI] [PubMed] [Google Scholar]
- Turkeltaub PE, Flowers DL, Verbalis A, Miranda M, Gareau L, Eden GF ( 2004): The neural basis of hyperlexic reading: An FMRI case study. Neuron 41: 11–25. [DOI] [PubMed] [Google Scholar]
- Wartenburger I, Heekeren HR, Abutalebi J, Cappa SF, Villringer A, Perani D ( 2003): Early setting of grammatical processing in the bilingual brain. Neuron 37: 159–170. [DOI] [PubMed] [Google Scholar]
- Wagner RK, Torgeson JK ( 1987): The nature of phonological awareness and its causal role in the acquisition of reading skills. Psychol Bull 101: 192–212. [Google Scholar]
- Wechsler AF ( 1977): Dissociative alexia. Arch Neurol 34: 257. [DOI] [PubMed] [Google Scholar]
- Wechsler D ( 1997): Wechsler Adult Intelligence Scale‐Third Edition. San Antonio: The Psychological Corporation. [Google Scholar]
- Woodcock RW, McGrew KS, Mather N ( 2001): Woodcock‐Johnson III: Tests of Achievement. Itasca, IL: The Riverside Publishing Company. [Google Scholar]
- Woods RP, Grafton ST, Watson JD, Sicotte NL, Mazziotta JC ( 1998a): Automated image registration: II. Intersubject validation of linear and nonlinear models. J Comput Assist Tomogr 22: 153–165. [DOI] [PubMed] [Google Scholar]
- Woods RP, Grafton ST, Holmes CJ, Cherry SR, Mazziotta JC ( 1998b): Automated image registration: I. General methods and intrasubject, intramodality validation. J Comput Assist Tomogr 22: 139–152. [DOI] [PubMed] [Google Scholar]
- Yokoyama S, Okamoto H, Miyamoto T, Yoshimoto K, Kim J, Iwata K, Jeong H, Uchida S, Ikuta N, Sassa Y, Nakamura W, Horie K, Sato S, Kawashima R ( 2006): Cortical activation in the processing of passive sentences in L1 and L2: An fMRI study. NeuroImage 30: 570–579. [DOI] [PubMed] [Google Scholar]
- Ziegler JC, Goswami U ( 2005): Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychol Bull 131: 3–29. [DOI] [PubMed] [Google Scholar]
- Zurowski B, Gostomzyk J, Gron G, Weller R, Schirrmeister H, Neumeier B, Spitzer M, Reske S, Walter H ( 2002): Dissociating a common working memory network from different neural substrates of phonological and spatial stimulus processing. NeuroImage 15: 45–57. [DOI] [PubMed] [Google Scholar]