

Abstract
We examined the hypothesis that learning to write Chinese characters influences the brain's reading network for characters. Students from a college Chinese class learned 30 characters in a character‐writing condition and 30 characters in a pinyin‐writing condition. After learning, functional magnetic resonance imaging collected during passive viewing showed different networks for reading Chinese characters and English words, suggesting accommodation to the demands of the new writing system through short‐term learning. Beyond these expected differences, we found specific effects of character writing in greater activation (relative to pinyin writing) in bilateral superior parietal lobules and bilateral lingual gyri in both a lexical decision and an implicit writing task. These findings suggest that character writing establishes a higher quality representation of the visual–spatial structure of the character and its orthography. We found a greater involvement of bilateral sensori‐motor cortex (SMC) for character‐writing trained characters than pinyin‐writing trained characters in the lexical decision task, suggesting that learning by doing invokes greater interaction with sensori‐motor information during character recognition. Furthermore, we found a correlation of recognition accuracy with activation in right superior parietal lobule, right lingual gyrus, and left SMC, suggesting that these areas support the facilitative effect character writing has on reading. Finally, consistent with previous behavioral studies, we found character‐writing training facilitates connections with semantics by producing greater activation in bilateral middle temporal gyri, whereas pinyin‐writing training facilitates connections with phonology by producing greater activation in right inferior frontal gyrus. Hum Brain Mapp, 2013. © 2012 Wiley Periodicals, Inc.
Keywords: writing, character, pinyin, sensori‐motor, orthography, phonology, semantics
INTRODUCTION
Skilled reading depends in part on high quality representations of written word forms. Writing, because it can help establish these forms, can be an important support for learning to read. For alphabetic reading, the importance of writing for establishing written representations for specific words may seem relatively unimportant, because orthographic representations are intimately connected and partly dependent on the phonological forms they represent. Success in reading, accordingly, is very dependent on establishing the phonological connections to orthography [Rayner et al., 2001; Vellutino et al., 2004; Ziegler and Goswami, 2005]. In reading Chinese, the case is slightly different. Although Chinese characters are also connected to phonological representations, these connections are less intimate, instantiated at the syllabic level rather than the phoneme level. Most important for the question of writing effects on reading, character connections to both phonology and meaning depend crucially on establishing a high quality representation of the written character form. Indeed, some studies suggest that orthographic awareness, rather than phonological awareness, is the most predictive variable for Chinese reading achievement [Chung et al., 2010; Siok and Fletcher, 2001].
In addition to its coarser mapping of phonology, Chinese orthography relies on characters with complex visual–spatial configurations. The Chinese character consists of any or all of eight basic strokes interwoven in patterns to form component radical(s) in a two‐dimensional square. Writing characters involves the coupling of writing‐related visual and motor systems. This coupling may help establish the spatial configuration of strokes and radicals, which along with a temporal sequence of motor movements associated with stroke composition, completely defines the shape of the character. These features of Chinese orthography are evident in the emphasis placed on writing in Chinese literacy instruction [Wu et al., 1999].
Several studies have suggested that writing skills are highly correlated with reading scores in native Chinese‐speaking children. One study found that performance on a delayed copying task (i.e. writing previously presented characters from memory) differs in young children with high or low overall literacy skill in Hong Kong [Pak et al., 2005]. Another study found that skill in copying Chinese pseudocharacters was highly correlated with reading scores in children, even with phonological processing skills statistically controlled [Tan et al., 2005b]. A study of Chinese dyslexic children also found that writing and reading are highly correlated [Chan et al., 2006].
More direct evidence of a causal link between writing and reading comes from learning studies. For example, Longcamp et al. (2005) found handwriting practice produced higher performance on a letter recognition task compared with typing practice, presumably because handwriting improves the quality of visual–spatial representation of letters to a greater degree than typing [Longcamp et al., 2005b]. Naka (1998) found that a writing condition produced better recall of pseudocharacters than a visual‐only condition in Japanese [Naka, 1998]. Most relevant to our study, a recent study of adult learners of Chinese found that a handwriting condition produced greater accuracy in a lexical decision task and a semantic task than pinyin‐typing and reading‐only conditions [Guan et al., 2011]. Pinyin, an alphabetic system that indicates pronunciation of a character, provided an advantage on phonological tasks.
The value of writing for Chinese reading may be to more strongly establish a visual–spatial memory that also has a motor memory trace. A motor memory that becomes stabilized through practice can last for a very long period of time [Shadmehr and Holcomb, 1997]. Motor memories for writing can be accessed during character recognition [Flores d'Arcais, 1994] and letter perception [Parkinson et al., 2010]. Both Flores d'Arcais (1994) and Parkinson et al. (2010) found a facilitative effect during reading when the stroke presentation order was consistent with the order in which people usually write the character/letter. This facilitation may arise from perceptual predictions triggered by the previous action‐consistent stroke sequence. Neuroimaging studies have also provided evidence that motor memory trace is wired with the visual perceptual representation. For example, two studies have found that visual perception of single letters activated premotor cortex and precentral gyrus, suggesting that writing‐related motor information is also involved in the process of visual recognition. [James and Gauthier, 2006; Longcamp et al., 2003].
To summarize, we assume that writing facilitates recognition for both Chinese characters and English letters because (1) writing adds additional motor‐related information to the representation system, which is wired together with visual input and enhances the activation of visual information during the recognition stage and (2) writing improves the quality of visual representation itself, which results in greater visual–orthographic activation during recognition. The question for our study is whether we can identify a writing effect on the neural network for Chinese reading.
The reading network for Chinese includes both regions shared with alphabetic reading and regions distinct neural mechanisms for reading across different writing systems [Bolger et al., 2005; Chen et al., 2002; Kuo et al., 2004; Paulesu et al., 2000; Tan et al., 2005a]. Among the common neural mechanisms, the left temporo‐occipital cortex and left inferior parietal lobule (IPL) are involved in both languages for orthographic analysis and conversion between orthography and phonology/semantics, respectively [Bolger et al., 2005; Booth et al., 2006; Tan et al., 2003]. Among the distinctive mechanisms, right temporo‐occipital cortex and bilateral superior parietal lobules are more involved in Chinese than English, presumably due to the complex holistic visual–spatial analysis required by Chinese characters [Cao et al., 2010; Tan et al., 2005a]. Left dorsal inferior/middle frontal gyrus (BA 9) is also more involved in Chinese than in English, which is probably associated with the retrieval of phonology at the whole syllable level and with lexical selection required by the extensive homophony in Chinese [Bolger et al., 2005; Tan et al., 2005a]. Left inferior frontal gyrus and left temporo‐parietal cortex are more involved in English than Chinese, because these two regions are involved in phonological segmentation and grapheme–phoneme conversion, respectively, both of which are important for alphabetic reading [Bolger et al., 2005; Tan et al., 2005a].
Neuroimaging studies on English‐speaking adults learning Chinese have examined the accommodation made by the alphabetic neural network to the demands of Chinese writing [Liu et al., 2007; Nelson et al., 2009]. Liu et al. (2007) and Nelson et al. (2009) found that learners showed greater activation in right fusiform gyrus and left dorsal inferior frontal gyrus for Chinese characters than English words, suggesting they accommodate the special features of Chinese during learning. These results allow a general conclusion about accommodation when writing was not a part of instruction. The additional accommodating effect of writing practice is the focus of this study.
In this study, we compare, using functional magnetic resonance imaging (fMRI), the neural correlates of the learning that occurs with two methods of character instruction for English‐speaking adults learning Chinese. One of the learning methods is character writing and the other is pinyin writing. Character writing may establish a high‐quality representation of orthography and strong connections to semantics, whereas pinyin writing may establish a high‐quality representation of phonology, while controlling for the activity of handwriting. We adopted a passive viewing task, which allowed us to examine brain activation for lexical processing without any additional task factors, a lexical decision task, in which we examined the process of visual–orthographic recognition, and an implicit writing task, which allowed us to directly examine the brain activation patterns associated with a mental simulation of writing. The key purposes of this study are (1) to examine whether character writing produces greater accommodation as evidenced by greater activation in the “Chinese network,” including bilateral temporo‐occipital cortex, bilateral superior parietal lobules, left dorsal inferior frontal gyrus (BA 9), and left IPL; (2) to examine whether character writing invokes greater involvement of sensori‐motor cortex (SMC) during character recognition; (3) to examine whether character writing creates stronger connections between orthography and semantics by showing greater activation in bilateral middle temporal gyri; and (4) to examine whether pinyin writing creates stronger connections between orthography and phonology by producing greater activation in bilateral inferior frontal gyri and left superior temporal gyrus.
METHODS
Participants
Seventeen undergraduate students enrolled in introductory Chinese at Carnegie Mellon University or the University of Pittsburgh (M age = 21.58, range: 19–24; 10 males) participated in the study. According to an informal interview, all participants met the following criteria: (1) monolingual English speaker, (2) not from a Chinese heritage family, (3) right‐handed, (4) free of neurological disease or psychiatric disorders, (5) no Attention Deficit Hyperactivity Disorder, and (6) no learning disability. The Institutional Review Board at the University of Pittsburgh reviewed and approved this study.
Procedures
Behavioral training and testing
Ninety characters that had not been taught before the experiment were chosen from the participants' Chinese textbook. These characters were evenly assigned to one of three conditions: character‐writing training, pinyin‐writing training, or novel. The characters were matched on variables of spatial structure (left‐right, up‐down, and simple), number of strokes, and frequency of the English translation [Kucera and Francis, 1967] across conditions. A pretest was given before learning started on the first day of training to make sure that none of the 90 characters were known. Participants learned 60 characters in all, with 30 appearing in each training condition for 5 days. The learning sequence of the two conditions was counterbalanced between participants and days via a Latin square arrangement. Training of each character was divided into three 800‐ms segments. For the first 800 ms, participants saw a character in the center of a computer screen; for the second 800 ms, they saw that character's pinyin and listened to a recording of its pronunciation by a native Chinese speaker; for the final 800 ms, they saw the English translation of the character. This sequence was followed by a 15‐s pause, during which the participant was asked to write down either the character or its pinyin, depending on the condition, three times. The entire sequence was repeated three times in a row for each character.
On each day of training, learning was followed by a testing session that included a lexical decision task. In this task, participants were asked to judge whether the stimulus presented on the screen was a real character or not. There were 30 stimuli of each type: characters learned in the character‐writing training condition, characters learned in the pinyin‐writing training condition, and novel characters. Participants were told to press a button with their index finger for a “yes” response and another button with their middle finger for a “no” response. Each stimulus was presented for 800 ms followed by a blank of 1,200 ms. False characters were created by deleting a stroke, adding a stroke, or switching the position of two radicals in a character. The presentation order of the different types of stimuli was randomized. The testing session included three other tasks: a character‐sound matching task, a character‐meaning matching task, and a partial character recognition task for the Carnegie Mellon students on the first 3 days. (For details about the procedures of these tasks, see Guan et al., 2011.)
After learning and testing on the last day of training, participants completed a post‐test to assess their proficiency on trained characters. The post‐test required participants to provide the pinyin and meaning of each of the 60 characters they had learned. The pretest and post‐test were paper–pen tests, and the learning and testing were computerized using E‐Prime.
The six Carnegie Mellon University students had 2 months previously taken part in a behavioral study of Chinese learning that involved 3 days of training and four tests (lexical decision, character‐sound matching, character‐meaning matching, and partial character recognition) after training on each day. To assure that all 17 participants had learned the characters sufficiently, these six students received two additional days of training before the scans. They completed two tasks during the fMRI session: passive viewing and lexical decision. The eleven University of Pittsburgh students received 5 days of training within a 2‐week interval. The materials and learning and testing procedures were the same for the Carnegie Mellon University students and University of Pittsburgh students, except that during training, the lexical decision task occurred on 5 days for the 11 University of Pittsburgh students and on only the first 3 days for the Carnegie Mellon University students. The two samples were comparable on the post‐test.
fMRI session
Within 1 week of the last training day, the fMRI session was administered, consisting of a passive viewing task followed by a lexical decision task. An implicit writing task then followed for the 11 University of Pittsburgh students. To reduce the repetition effect across tasks, we believe that the passive viewing task should go first due to the least requirement of encoding and that the implicit writing task should go last, because it requires deep coding. We used a block design for the passive viewing task and an event‐related design for the lexical decision and implicit writing tasks. There was a 12‐s equilibration period at the beginning of each fMRI run and a 22‐s period at the end to be able to deconvolve the hemodynamic response function (HRF) for the last trial.
Passive viewing
In this task, participants were asked to view a stimulus presented in the center of the screen. There were four types of stimuli, including 30 Chinese characters learned in the character‐writing condition, 30 characters learned in the pinyin‐writing condition, 30 novel characters, and 30 English words. Six five‐item blocks were presented for each stimulus type; each item was presented for 800 ms followed by a 200 ms blank. The four experimental blocks were interleaved with baseline blocks, during which a fixation (+) was presented using the same procedure as the experimental stimuli. The passive viewing task lasted 4 min and 34 s.
Lexical decision
This task was the same as the one in the testing session during training, except that we added 45 null trials with a fixation (+) serving as a baseline. The null trials were presented using the same procedure as the experimental trials, and participants were asked to press the “yes” button for null trials. The lexical decision task in the scanner lasted 5 min and 4 s.
Implicit writing
Participants were asked to imagine writing with their index finger the stimulus they had seen presented briefly on the screen. There were 30 stimuli of each type: characters learned in the character‐writing training condition, characters learned in the pinyin‐writing training condition, and novel characters. There were also 45 null trials with a fixation (+) serving as a baseline, during which participants were asked to imagine writing the fixation (+) with their index finger. Each stimulus was presented for 200 ms followed by a blank of 1,800 ms. The presentation order of different types of stimuli was randomized. This task also lasted 5 min and 4 s.
MRI data acquisition
After informed consent was obtained, an informal interview was administered to confirm each participant's language background, hand dominance, and neuropathological and psychiatric history. The participant then practiced abbreviated versions of the experimental tasks to become familiarized with the tasks. Different stimuli (matched on key characteristics) were used in the practice and fMRI sessions.
All images were acquired using a 3 T Siemens scanner. Gradient‐echo localizer images were acquired to determine the placement of the functional slices. For the functional imaging studies, a susceptibility weighted single‐shot echo planar imaging (EPI) blood oxygenation level‐dependent method was used. Functional images were interleaved from bottom to top in a whole brain EPI acquisition. The following scan parameters were used: TR = 2,000 ms, TE = 25 ms, flip angle = 79°, matrix size = 64 × 64, field of view = 205 mm, slice thickness = 3.2 mm, and number of slices = 38. These scanning parameters resulted in a 3.2 mm × 3.2 mm × 3.2 mm voxel size. At the end of the functional imaging session, a high resolution, T1 weighted 3D image was acquired (magnetization prepared rapid gradient echo, TR = 1,640 ms, TE = 2.48 ms, TI = 800 ms, flip angle = 8°, matrix size = 256 × 256, field of view = 249 mm, slice thickness = 0.8 mm, and number of slices = 256). The orientation of the 3D volume was identical to the functional slices.
Image data analysis
Data analysis was performed using SPM5 (Statistical Parametric Mapping; http://www.fil.ion.ucl.ac.uk/spm). The functional images were corrected for differences in slice‐acquisition time to the middle volume and were realigned to the last volume in the scanning session using affine transformations. No individual runs had more than 4 mm maximum movement for any subject in the x‐plane (M = 0.44, range = 0.08–1.57), y‐plane (M = 0.91, range = 0.25–2.77), or z‐plane (M = 1.89, range = 0.27‐3.66). Furthermore, no individual runs had more than 3° of maximum displacement in rotation for pitch, yaw, or roll. An analysis of variance (ANOVA) with task as an independent variable showed no significant main effects on any of the above six dependent variables. All statistical analyses were conducted on movement‐corrected images. Co‐registered images were normalized to the Montreal Neurological Institute average template (12 linear affine parameters for brain size and position, eight nonlinear iterations, and 2 × 2 × 2 nonlinear basis functions). Statistical analyses were calculated on the smoothed data (9 mm Gaussian kernel).
Data from each subject were entered into a general linear model using a block analysis procedure for the passive viewing task and an event‐related analysis procedure for the lexical decision and implicit writing tasks with a canonical HRF. Statistics were calculated with a high pass filter (128 s cutoff period). We used global normalization to scale the mean of each scan to a common value. Parameter estimates from contrasts of the canonical HRF in single subject models were entered into random‐effects analyses. All whole brain results are reported at P < 0.001 uncorrected and contain 10 or more voxels.
To determine differences between viewing English words and Chinese characters, we employed paired t‐tests to examine the contrasts of Chinese characters versus English words in the passive viewing task. To determine differences between character writing‐training and pinyin‐writing training, we analyzed the contrasts of character‐writing training versus pinyin‐writing training in all three tasks. To determine differences between learned characters and novel characters, we analyzed the contrasts of learned characters (combined character‐writing training and pinyin‐writing training conditions) versus novel characters in all three tasks.
In a volume of interest (VOI) analysis for the lexical decision task, we further examined whether the character‐writing effect can be transferred to new characters by comparing novel characters to character‐writing training and pinyin‐writing training in one ANOVA model. If novel equals character writing, then the writing effect can be generalized. If novel equals pinyin writing and smaller than character writing, then the writing effect is specifically about writing. Two VOIs were created at the peaks in the contrasts of character‐writing training minus pinyin‐writing training with a radius of 6 mm. The two VOIs were left superior parietal lobule (−32, −54, 54) and right postcentral gyrus (38, −38, 48). We then used the implicit writing task to create a conjunction map (with an exclusive mask) of all three conditions (character writing, pinyin writing, and novel). The conjunction map was created using SPM at a threshold of P < 0.001, uncorrected. The conjunction of all three conditions in the implicit writing task provides a map dedicated to the nature of the task of “implicit writing,” which may include retrieving visual configuration of the character, recalling the stroke sequence, and writing‐related motor planning. If the character‐writing effect found in the lexical decision task is caused by the actual “writing,” and if actual writing is partly simulated by imagined writing, then there should be nothing left when the exclusive mask from the conjunction analysis is applied.
At the whole brain level, we carried out brain behavior correlation analyses for each condition in the lexical decision task. We correlated accuracy on character‐writing trained, pinyin‐writing trained, and novel characters with their corresponding brain activation in the contrast of lexical minus null.
RESULTS
Behavioral performance during the training
As shown in Table I, lexical decision accuracy and decision times improved over the course of the 5‐day training for both character‐writing and pinyin‐writing conditions. An ANOVA of condition (character‐writing training, pinyin‐writing training, and novel) by day (1–5) on accuracy found significant main effects of condition (F(2, 8) = 34.823, P = 0.000 for accuracy) and day (F(4, 16) = 15.213, P = 0.000 for accuracy), with no significant interaction. Accuracy for the character‐writing trained characters was significantly higher than for the pinyin‐writing trained characters (t(16) = 4.210, P = 0.001) and the novel characters (t(16) = 7.469, P = 0.000). The accuracy on the pinyin‐writing trained characters was higher than that on the novel characters (t(16) = 4.916, P = 0.000). Multiple comparisons on day found that accuracy on Day 4 was significantly higher than that on Day 3 (t(16) = 3.785, P = 0.004), corrected for multiple comparisons (P < 0.005).
Table I.
Means and standard deviations of accuracy and reaction time for each condition on each day on the lexical decision task
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
|---|---|---|---|---|---|
| Accuracy | |||||
| Character‐writing trained | 0.60 (0.21) | 0.67 (0.21) | 0.78 (0.11) | 0.82 (0.07) | 0.82 (0.11) |
| Pinyin‐writing trained | 0.57 (0.21) | 0.62 (0.19) | 0.70 (0.11) | 0.73 (0.11) | 0.73 (0.10) |
| Novel | 0.51 (0.23) | 0.52 (0.18) | 0.56 (0.10) | 0.62 (0.15) | 0.59 (0.09) |
| Reaction time | |||||
| Character‐writing trained | 716 (119) | 659 (109) | 659 (91) | 591 (37) | 583 (35) |
| Pinyin‐writing trained | 701 (97) | 660 (117) | 672 (102) | 591 (38) | 583 (47) |
| Novel | 725 (120) | 675 (125) | 668 (98) | 609 (37) | 577 (60) |
Table II presents accuracy on meaning and pinyin production in the post‐test after training on the last day. An ANOVA of condition (character‐writing training and pinyin‐writing training) by measurement (meaning production and pinyin production) found a significant main effect of measurement (F(1, 15) = 5.724, P = 0.03), with higher accuracy on meaning production than on pinyin production. The main effect of condition and the interaction of condition by measurement were not significant. Thus, before the scans, character‐writing trained and pinyin‐writing trained characters had been learned equally well.
Table II.
Means and standard deviations of accuracy on the meaning production and pinyin production in the post‐test for each learning condition
| Meaning | Pinyin | |
|---|---|---|
| Character‐writing training | 0.95 (0.10) | 0.89 (0.17) |
| Pinyin‐writing training | 0.96 (0.13) | 0.92 (0.17) |
Behavioral performance in the scanner
Table III presents accuracy and reaction time data for each condition on the lexical decision task in the scanner. Separate ANOVAs of condition effects (character writing, pinyin writing, novel, and baseline) for accuracy and reaction time on correct trials showed main effects of condition for accuracy (F(3,48) = 67.237, P < 0.001) and reaction time (F(3,48) = 93.916, P < 0.001). Multiple comparisons found that accuracy on the character‐writing condition was higher than that on the pinyin‐writing condition (t(16) = 7.853, P < 0.001) and on the novel condition (t(16) = 8.025, P < 0.001). Accuracy on pinyin writing was higher than that on novel (t(16) = 5.533, P < 0.001). Accuracy on the baseline was higher than that on the character‐writing condition (t(16) = 3.166, P < 0.008 corrected for multiple comparisons P = 0.05/6), the pinyin‐writing condition (t(16) = 9.218, P < 0.001), and the novel condition (t(16) = 11.203, P < 0.001). Multiple comparisons found that reaction time on the baseline was faster than that on the character‐writing condition (t(16) = 12.234, P < 0.001), the pinyin‐writing condition (t(16) = 12.448, P < 0.001), and the novel condition (t(16) = 10.205, P < 0.001). The differences between the character writing, pinyin writing, and novel were not significant.
Table III.
Means and standard deviations of accuracy and reaction time for each condition on the lexical decision task and the baseline task in the scanner
| Character‐writing trained | Pinyin‐writing trained | Novel | Baseline | |
|---|---|---|---|---|
| Accuracy | 0.91 (0.09) | 0.80 (0.08) | 0.66 (0.14) | 0.97 (0.04) |
| Reaction time | 755 (72) | 759 (82) | 781 (96) | 563 (69) |
Brain activation patterns
Passive viewing
In the passive viewing task, we found greater activation for viewing Chinese characters than for viewing English words in left dorsal inferior frontal gyrus/middle frontal gyrus (BA 9), bilateral precuneus, bilateral superior parietal lobules, bilateral middle occipital gyri, and bilateral fusiform gyri. We found greater activation for viewing English words than for viewing Chinese characters in bilateral inferior frontal gyrus (BA 45, 47) and bilateral superior temporal gyri (BA 22, 21) (see Table IV and Fig. 1). We found no differences between viewing characters learned in the character‐writing training condition and the pinyin‐writing training condition. We found greater activation for learned characters than novel characters in left precentral gyrus, left precuneus, and left superior frontal gyrus (see Appendix). Thus, passive viewing reflected language differences and general learning effects, but not specific training condition effects.
Table IV.
Brain activation for the comparisons between Chinese characters and English words in the passive viewing task
| Anatomical region | H | BA | Voxels | Z‐Score | x | y | z |
|---|---|---|---|---|---|---|---|
| Chinese > English | |||||||
| Precuneus, superior parietal lobule | L | 7 | 2464 | 5.42** | −14 | −74 | 42 |
| Precuneus | R | 7 | 1417 | 4.74** | 20 | −64 | 36 |
| Middle occipital gyrus, inferior temporal gyrus, fusiform gyrus | L | 19, 37 | 1090 | 5.38** | −52 | −64 | −8 |
| Middle occipital gyrus, inferior temporal gyrus, fusiform gyrus | R | 19, 18, 37 | 3350 | 5.23** | 44 | −74 | −16 |
| Superior frontal gyrus | L | 6 | 527 | 4.98** | −4 | 4 | 58 |
| Middle frontal gyrus | L | 6 | 23 | 3.37* | −24 | −4 | 54 |
| Dorsal Inferior frontal gyrus | L | 9 | 24 | 3.43** | −50 | 2 | 34 |
| Supramarginal gyrus | L | 40 | 19 | 3.22* | −36 | −48 | 38 |
| Insula | L | ‐‐ | 230 | 4.19** | −30 | 14 | 8 |
| Putamen | R | ‐‐ | 91 | 4.10** | 30 | 16 | 2 |
| Cerebellar tonsil | R | ‐‐ | 21 | 3.87** | 24 | −36 | −44 |
| Posterior cingulated | L | ‐‐ | 306 | 3.74** | −4 | −36 | 24 |
| English > Chinese | |||||||
| Inferior frontal gyrus | L | 47, 45 | 299 | 4.12* | −46 | 36 | −14 |
| Inferior frontal gyrus | R | 47 | 137 | 3.69* | 46 | 36 | −8 |
| Superior temporal gyrus, middle temporal gyrus, | L | 22 | 2329 | 4.24* | −38 | −54 | 20 |
| Inferior temporal gyrus | L | 20 | 2329 | 4.58* | −54 | −28 | −22 |
| Superior temporal gyrus | R | 21 | 651 | 4.83* | 60 | −62 | 20 |
| Middle temporal gyrus | R | 21 | 24 | 3.34* | 64 | −6 | −12 |
| Inferior temporal gyrus | R | 20 | 10 | 3.18* | 50 | −16 | −24 |
| Medial frontal gyrus | L | 25 | 45 | 3.61* | −2 | 28 | −18 |
| Superior frontal gyrus | L | 8 | 137 | 4.43* | −12 | 42 | 46 |
| Precentral gyrus | R | 6 | 17 | 3.50* | 60 | 12 | 10 |
| Parahippocampal gyrus | L | 34 | 17 | 3.38* | −16 | −10 | −22 |
| Cerebellum | R | ‐‐ | 95 | 4.21* | 22 | −80 | −36 |
Brain regions listed in bold are the peak coordinate for areas spanning different regions. H, hemisphere; L, left; R, right; BA, Broadmann's area.
**P < 0.01, FDR corrected.
P < 0.05, FDR corrected.
Figure 1.
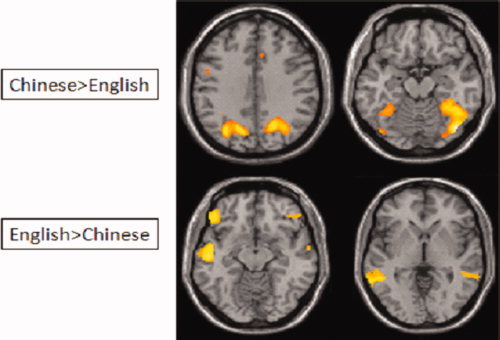
Comparisons between languages in the passive viewing task. There was greater activation in left dorsal inferior/middle frontal gyrus (MFG), and bilateral fusiform gyrus (FG) for viewing Chinese characters than viewing English words. There was greater activation in left inferior frontal gyrus (IFG) and left superior temporal gyrus (STG) for viewing English words than viewing Chinese characters.
Lexical decision
In the lexical decision task, we found greater activation for the character‐writing condition than the pinyin‐writing condition in bilateral superior parietal lobules, right IPL and postcentral gyrus (see Table V and Fig. 2). When an exclusive mask was applied, no regions showed greater activation for the character‐writing training than the pinyin‐writing training. The mask was a conjunction analysis of character‐writing training, pinyin‐writing training, and novel in the implicit writing task which provided a network involved in the activity of character writing (see Table AII). This analysis suggests that the recognition of a character learned in the character‐writing training condition invokes activation of a network involved in the previous writing training. We found greater activation for the pinyin‐writing condition than the character‐writing condition in right inferior frontal gyrus (see Table IV). We found greater activation in bilateral middle occipital gyri, precuneus, and left middle temporal gyrus for learned characters than novel characters (see Table AI).
Table V.
Brain activations for the comparisons between the two learning conditions in the lexical decision task and Correlations between accuracy and brain activation
| Anatomical region | H | BA | Voxels | Z‐Score | x | y | z |
|---|---|---|---|---|---|---|---|
| Character‐writing trained > Pinyin‐writing trained | |||||||
| Superior parietal lobule, precentral gyrus | R | 40, 3 | 151 | 3.87 | 38 | −38 | 48 |
| Inferior parietal lobule, postcentral gyrus | R | 40, 5 | 49 | 3.86 | 42 | −48 | 56 |
| Superior parietal lobule, postcentral gyrus | L | 7 | 78 | 3.62 | −32 | −54 | 54 |
| Pinyin‐writing trained > Character‐writing trained | |||||||
| Inferior frontal gyrus | R | 47 | 15 | 3.44 | 34 | 20 | −6 |
| Higher accuracy correlated with greater activation for the character‐writing trained characters in the lexical decision task | |||||||
| Lingual gyrus | R | 19, 18, 37 | 225 | 4.02* | 26 | −76 | −12 |
| Postcentral gyrus, precentral gyrus | L | 3 | 41 | 3.92* | −36 | −24 | 44 |
| Superior parietal lobule | R | 7 | 13 | 3.29 | 32 | −72 | 44 |
| Cerebellum | L | — | 206 | 4.70* | −14 | −58 | −16 |
Figure 2.
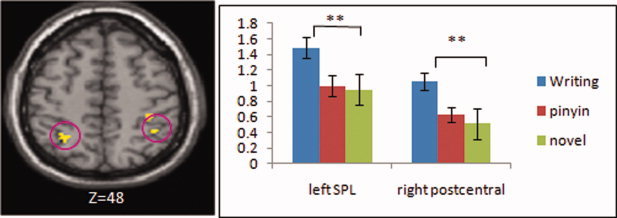
Comparisons between conditions in the lexical decision task. The activation at bilateral superior parietal lobule (SPL) and right postcentral gyrus was greater for character writing than pinyin writing and novel (character writing > pinyin writing = novel). Volumes of interest used in the VOI analysis in the bar graphs on the right were indicated in the pink circles on the left.
An ANOVA of condition (character‐writing training, pinyin‐writing training, and novel) at the VOI analysis found a significant main effect of condition for left superior parietal lobule (F(2,32) = 13.555, P = 0.000) and for right postcentral gyrus (F(2,32) = 18.103, P = 0.000). Multiple comparisons found that writing‐trained characters showed greater activation than novel characters, (t(16) = 4.380, P = 0.000) at the left superior parietal lobule, and (t(16) = 5.834, P = 0.000) at the right postcentral gyrus. There were no differences between the pinyin‐writing training condition and the novel condition (t(16) = 0.431, P = 0.672) for the left superior parietal lobule, or (t(16) = 1.163, P = 0.262) for the right postcentral gyrus (see Fig. 2).
Implicit writing
In the implicit writing task, we found greater activation in bilateral middle temporal gyri, left IPL, bilateral lingual gyri, left superior/middle frontal gyrus (BA 9, 8, 6), and left precentral gyrus for the character‐writing condition than for the pinyin‐writing condition (Table VI and Fig. 4). We found no greater activation for pinyin writing than for character writing. We also found greater activation for learned characters than novel character in left precuneus, precentral gyrus and right precuneus and superior parietal lobule (see Table AI). The “Discussion section” will focus on the differences between character‐writing training and pinyin‐writing training.
Table VI.
Brain activations for the comparisons between the two learning conditions in the implicit writing task
| Anatomical region | H | BA | Voxels | Z‐Score | x | y | z |
|---|---|---|---|---|---|---|---|
| Character‐writing trained > Pinyin‐writing trained | |||||||
| Inferior parietal lobule, supramarginal gyrus | L | 40 | 79 | 4.20* | −64 | −44 | 36 |
| Middle temporal gyrus | L | 22 | 129 | 5.16** | −48 | −46 | 6 |
| Superior temporal gyrus | L | 22 | 229 | 4.68* | −40 | −36 | 4 |
| Middle temporal gyrus | L | 21 | 80 | 3.91* | −68 | −26 | 2 |
| Superior temporal gyrus | R | 41 | 66 | 4.27* | 46 | −36 | 12 |
| Superior temporal gyrus | R | 22 | 78 | 4.07* | 58 | −40 | 18 |
| Middle temporal gyrus | R | 22 | 242 | 4.18* | 66 | −34 | 4 |
| Middle temporal gyrus | R | 39 | 110 | 4.02* | 46 | −60 | 24 |
| Middle temporal gyrus | R | 22 | 38 | 3.57* | 40 | −60 | 10 |
| Lingual gyrus | L | 37 | 99 | 3.67* | −22 | −46 | −16 |
| Lingual gyrus | R | 19, 37 | 20 | 3.65* | 32 | −74 | −14 |
| Middle frontal gyrus, | L | 8 | 37 | 3.98* | −32 | 22 | 44 |
| Superior frontal gyrus, Middle frontal gyrus | L | 9,10 | 297 | 4.39* | −16 | 52 | 38 |
| Postcentral gyrus | L | 3, 4 | 11 | 3.44* | −18 | −38 | 66 |
| Posterior cingulated | R | 29 | 1294 | 5.19* | 4 | −58 | 10 |
| Pinyin‐writing trained > Character‐writing trained | |||||||
| Pons | L | — | 82 | 3.99 | −8 | −38 | −36 |
Brain regions listed in bold are the peak coordinate for areas spanning different regions. H, hemisphere; L, left; R, right; BA, Broadmann's area.
**P < 0.01, FDR corrected.
P < 0.05, FDR corrected.
Figure 4.
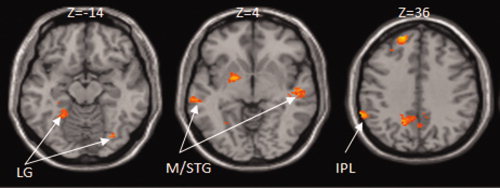
Comparisons between conditions in the implicit writing task. There was greater activation in bilateral lingual gyrus (LG), bilateral middle/superior temporal gyrus (M/STG), and left inferior parietal lobule (IPL) for the character‐writing condition than for the pinyin‐writing condition.
Brain‐behavior correlations
For the lexical decision task, we carried out brain‐behavior correlation analyses on the whole brain level and we found that greater activations in right superior parietal lobule (rSPL), right lingual gyrus (rLG), and left precentral gyrus were correlated with higher accuracy in the character‐writing trained condition (see Table V, and Fig. 3). We found no significant correlation for the pinyin‐writing trained or novel character conditions.
Figure 3.
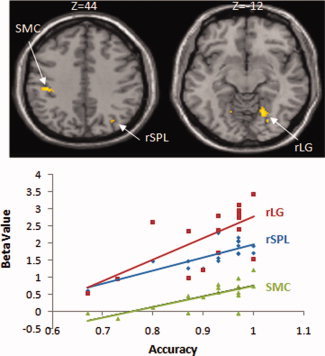
Correlation of accuracy in the character‐writing condition in the lexical decision task with brain activations in left sensori‐motor cortex (SMC) (r = 0.729, P = 0.001), right superior parietal lobule (rSPL) (r = 0.687, P = 0.002), and right lingual gyrus (rLG) (r = 0.832, P = 0.000).
DISCUSSION
Differences between Chinese and English
We found greater activation in bilateral superior parietal lobules, bilateral middle occipital gyri, bilateral fusiform gyri, and left dorsal inferior frontal gyrus for viewing Chinese characters than for viewing English words. On the other hand, we found greater activation in ventral left inferior frontal gyrus and left superior posterior temporal gyrus for viewing English words than Chinese characters.
For English, our results converge with findings concerning the left superior temporal gyrus and left inferior frontal gyrus, which are associated with the route of assembled phonology in English. The left posterior superior temporal gyrus and adjacent angular and supramarginal gyrus are involved in grapheme–phoneme conversion and fine‐grained phonemic analysis for alphabetic reading [Booth et al., 2003a; Eden et al., 2004; Poldrack et al., 2001; Temple et al., 2003] and the left inferior frontal gyrus (BA 45/46/47) is involved in phonological manipulation [Fiez et al., 1999] and subvocal rehearsal in phonological processing [Chein and Fiez, 2001]. Both of these processes are more involved in English reading than Chinese reading.
For Chinese, our results converge with results showing greater involvement of bilateral visual–spatial areas and visual–orthographic areas [Bolger et al., 2005; Tan et al., 2005a]. Previous research has also found greater involvement for Chinese reading in a dorsal posterior region of the left inferior frontal gyrus bordering on the precentral gyrus (BA 6/9) [Bolger et al., 2005; Tan et al., 2003, 2005a]. In our study, we observed activation during character reading near this same “Chinese region,” at the left dorsal inferior frontal gyrus (BA9). The peak was at (−50, 2, 34), very close to the peaks of (−48, 4, 35) and (−44, 4, 33) for learners in Nelson et al. (2009) and Liu et al. (2007), respectively, and to the peaks of (−48, 9, 30) and (−46, 8, 34) for native Chinese speakers in Bolger (2005) and Cao (2010), respectively. This convergence of findings suggests that English speakers can adopt the procedures of addressed retrieval of phonology and semantics after only a short period learning Chinese.
There are diverse hypotheses about the function of this region in Chinese reading: the first three are variations on the observation that this region holds character information to coordinate orthography, phonology, and semantics [Perfetti et al., 2005]: (1) visuo‐spatial analysis of Chinese characters that allows syllable level mapping [Tan et al., 2001]; (2) a long‐term storage center for addressed phonology [Tan et al., 2005a]; (3) a character memory to allow retrieval of associated character constituents [Perfetti et al., 2007], allowing lexical selection and integration [Li, 2010]; and (4) writing involvement, reflecting its location just anterior to the premotor cortex that governs motor functions and the writing practice that is part of Chinese reading acquisition [Tan et al., 2005a]. This region is involved in English reading as well under some circumstances [Binder et al., 2003; Bolger et al., 2008], e.g. when there is grapheme–phoneme inconsistency. A control function for selecting and integrating across competing activations could be a function that supports both Chinese and alphabetic reading.
That learners with just over one term of study show activation patterns typical of skilled native language Chinese readers demonstrates the accommodation of the brain's reading network to the demands of the writing system [Liu et al., 2007; Nelson et al., 2009]. For example, the bilateral activation of fusiform gyri and middle occipital gyri for Chinese suggests a role for holistic visual configuration in the right hemisphere, in addition to the left hemisphere role in more fine‐grain (e.g. stroke patterns) orthographic processing. As suggested by Liu and Perfetti (2003), the right hemisphere, responding to low spatial frequencies, may be important in identifying the spatial configuration of radicals [Liu and Perfetti, 2003]. The fact that learners as well as L1 Chinese readers show this pattern, suggest that this right hemisphere involvement reflects a relatively rapid accommodation to the specific graphic demands of the writing system [Liu et al., 2007]. Another accommodation may be seen in the role of the bilateral superior parietal lobules, which may support memories for the visuo‐spatial configuration of components within a character. The superior parietal lobules, as well as the bilateral middle occipital gyri, show age‐related increases in Chinese reading over development, suggesting a role in the acquisition of Chinese reading skill [Cao et al., 2010; Cao et al., 2009]. Finally, as we observed above, learners also showed activation in the left dorsal frontal region (BA9) that suggest an accommodation to the properties of Chinese characters that call on that region.
Differences between the two learning conditions
The most interesting contrast in this study is between the two learning conditions. Consistent with our expectations, we found that character writing facilitates accommodation in learning Chinese at both the script level and the mapping level. For facilitation at the script level we have both behavioral and neurological evidence. Behaviorally, we found greater accuracy for character‐writing trained characters than pinyin‐writing trained characters in the lexical decision task, which directly reflects orthographic recognition. Neurologically, we found greater activation in bilateral superior parietal lobules during the lexical decision task, and greater activation in bilateral lingual gyri during the implicit writing task for character‐writing trained characters than pinyin‐writing trained characters. We also found that higher lexical decision accuracy was correlated with greater activation in rLG and rSPL for character‐writing trained characters. Moreover, the area around the rLG (26, −76, −12) that produced the correlation with accuracy overlaps with the character‐writing effect observed in the implicit writing task (32, −74, −14). Further, the area around the rSPL (32, −72, 44) that correlated with accuracy is near the area that showed the character‐writing effect in the lexical decision task (38, −38, 48). Overall, character‐writing training produced greater activation in bilateral superior parietal lobules and bilateral lingual gyri and the right homologues of these two regions are especially sensitive to the effectiveness of the training.
A previous study suggests that superior parietal cortex may play a crucial role in learning the visual–spatial aspects of characters, because its activation is correlated with behavioral improvement in learning and with better long‐term retention [Deng et al., 2008]. Our study adds the idea that character‐writing training helps this learning process, because during character writing, which is a process of producing the visual forms from memory, attention is explicitly drawn to the visuo‐spatial structure of the character. Beyond the global spatial layout, the learner needs to acquire a precise representation of character, including its strokes and the spatial relations among them. Our finding of greater activation in bilateral lingual gyri for the character‐writing trained characters suggests that character writing may help establish these more precise orthographic representations, perhaps through specifying the character‐writing stroke sequence. James (2010) found that writing training but not visual‐only training produced increased activation in bilateral fusiform/lingual gyri from pretraining to post‐training scans in a letter recognition task in children of 4–5 years of age, suggesting greater neural specialization of letters in fusiform gyri [James, 2010]. Another study also suggests that left fusiform gyrus is involved in not only orthographic configuration but also in the dynamic aspect of the writing sequence of Chinese characters by showing greater activation in this region for viewing incorrect sequences than for viewing correct sequences of character writing [Yu et al., 2011]. This finding suggests that writing training facilitates neural specialization of orthographic recognition for both Chinese characters and English letters by establishing a more elaborated visuo‐spatial and orthographic representation of the graphs.
When participants read a character in a lexical decision task, there was increased involvement of bilateral SMC for characters learned through character writing than for characters learned with pinyin writing. We also found that higher accuracy was correlated with greater activation in the left SMC for character‐writing trained characters in the lexical decision task. According to the common‐coding principle proposed by Prinz (1997), “event codes” and “action codes,” the functional bases of percepts and action plans, respectively, share the same representational domain and are therefore commensurate. Consistent with this principle, two previous studies found that visual presentation of letters produced activation in ventral left premotor area (BA 6) involved in writing for right‐handed writers and in right premotor area for left‐handed writers [Longcamp et al., 2003, 2005a]. James and Atwood (2009) found that writing training, but not visual‐only training, produced increased activation in left precentral gyrus in adults during a pseudoletter recognition task, suggesting that regions involved in writing were activated during recognition [James and Atwood, 2009]. Our study provides another piece of evidence for the common‐coding principle, and further suggests that the more strongly the SMC is wired with the recognition network, the greater performance will be. Sensori‐motor learning enriches the representation system by introducing into the network a motor component that is highly interactive with the other components and may become especially helpful for perception when other components are impaired or weak. This finding therefore has implications for intervention studies on dyslexia. Further research is needed to determine how writing aids in word recognition in children with reading disability.
Beyond these visual–spatial accommodations, we found that character writing also supports accommodation to the mapping level of the Chinese writing system. The clearest indicator for this is the greater activation in left IPL in the implicit writing task. The left IPL is associated with the mapping between orthography and phonology in Chinese [Cao et al., 2010; Tan et al., 2005a] and English [Booth et al., 2002, 2003b]. The left IPL involved in Chinese is more dorsal and lateral than the one involved in English, which may indicates the whole‐syllable level mapping in Chinese versus the phoneme level mapping in English. Other studies have also suggested that this region is involved in the mapping between orthography and semantics [Chou et al., 2006b]. Another piece of evidence for the benefit of character writing on mapping is that our study identified a semantic advantage for character writing for adult learners of Chinese. We found greater activation in bilateral middle/superior temporal gyri for the character‐writing condition than the pinyin‐writing condition in the implicit writing task, a region associated with verbal semantic representation and storage [Chou et al., 2006a; Howard et al., 1992; Shaywitz et al., 2001]. Our result is consistent with previous behavioral findings that writing practice is more helpful for semantic tasks than for reading only and typing tasks [Guan et al., 2011]. In general, English speakers learning Chinese without an emphasis on spoken language show greater accuracy in memorizing character meanings than pronunciations.
We also report significantly higher accuracy for meaning production than pinyin production on the post‐test in this study. It appears that when English speakers learn Chinese characters, they tend to attach meaning to the visual form before pronunciation. In behavioral data collected during character learning, it has been found that the benefits of character writing for orthographic recognition appears on the first day, whereas the benefit for semantics becomes significant on the third day. This finding suggests that character writing helps establish a high‐quality orthographic representation and that when visual recognition is automatic, memory and attention resources can be focused on building up the connections first between orthography and semantics and then between orthography and phonology. On the other hand, we also identified a phonological advantage for pinyin writing. There was greater activation in right inferior frontal gyrus for the pinyin‐writing condition than the character‐writing condition in the lexical decision task. A previous transcranial magnetic stimulation study found that right posterior inferior frontal gyrus is necessary for accurate and efficient phonological decisions [Hartwigsen et al., 2010]. Our study suggests that pinyin training helps to establish a more elaborated phonological representation. This is consistent with behavioral results [Guan et al., 2011] in which pinyin‐typing training led to an advantage in identifying the pinyin and tone of the characters. The behavioral data from this study, which will be reported in a separate paper, also suggests that pinyin writing is especially beneficial for tone production. Other researchers have found that right inferior frontal gyrus is involved in tone production in Mandarin Chinese in both native speakers [Liu et al., 2006] and English learners [Wang et al., 2003].
The writing‐specific nature of the character‐writing effect
In the VOI analyses at left superior parietal lobule and right SMC for the lexical decision task we found that the pinyin‐writing trained characters induced the same amount of activation as novel characters, even though accuracy was significantly higher in the pinyin‐writing training condition than in the novel condition. The character‐writing trained characters induced significantly greater activation at those two regions than the pinyin‐writing trained and novel conditions. This result suggests that these regions are sensitive not to accuracy, but to the specific experience of character writing. The application of an exclusive mask from a conjunction analysis of the implicit writing task also bolsters the idea that the character‐writing effect is due to the specific experience of “writing.” The present results might be due to the fact that only 10% of the characters in the pinyin‐writing and novel conditions share a radical with those in the character‐writing condition. A parametric manipulation of the overlap of radicals in different conditions would be helpful in understanding whether the writing effect can be generalized to other characters comprised of the same radicals. One might also expect the proficiency level of Chinese and the amount of writing training to influence generalization.
CONCLUSIONS
English speakers are able to accommodate to the special features of the visual form of Chinese characters and the relatively arbitrary mapping from the visual form to pronunciation and meaning by showing greater activation for Chinese characters than English words in bilateral superior parietal lobules, bilateral lingual gyri, and left dorsal inferior frontal gyrus (BA 9). Character writing facilitates the accommodation at both the visual form level and the mapping level. Writing improves the quality of visual–orthographic representation, showing increased activation in bilateral lingual gyri and bilateral superior parietal lobules. The increased precision of orthography boosts the processes of connecting orthography to semantics and phonology, indicated by increased activation in left IPL and bilateral middle/superior temporal gyri. Character writing also increases the interactivity with the SMC during character recognition, suggesting that learning by doing involves more neural resources.
Table AI.
Regions that showed greater activation for learned (combined character writing and Pinyin writing) than novel in the passive viewing and implicit writing tasks
| Anatomical region | H | BA | Voxels | Z‐Score | x | y | z |
|---|---|---|---|---|---|---|---|
| Passive viewing Learned > novel | |||||||
| Inferior parietal lobule | L | 40, 39 | 35 | 3.90 | −46 | −28 | 24 |
| Superior frontal gyrus | L | 6, 4 | 54 | 3.86 | −10 | −2 | 70 |
| Medial frontal gyrus | R | 6, 4 | 30 | 3.85 | 12 | −18 | 60 |
| Medial frontal gyrus | L | 6 | 16 | 3.84 | −10 | −12 | 52 |
| Precuneus | L | 7 | 112 | 3.81 | −12 | −60 | 40 |
| Inferior parietal lobule | L | 40, 6 | 17 | 3.68 | −38 | −24 | 28 |
| Superior frontal gyrus | L | 11 | 16 | 3.48 | −20 | 48 | −10 |
| Claustrum | L | 13 | 12 | 3.30 | −34 | 6 | 0 |
| Lexical decision Learned > novel | |||||||
| Middle occipital gyrus | R | 18, 19 | 143 | 4.94 | 30 | −90 | 10 |
| Superior occipital gyrus, precuneus | R | 19 | 43 | 3.57 | 38 | −78 | 24 |
| Posterior cingulate | R | 30 | 499 | 4.67 | 4 | −58 | 14 |
| Inferior parietal lobule | R | 40 | 160 | 4.55 | 42 | −38 | 50 |
| Middle frontal gyrus | L | 6 | 22 | 4.17 | −20 | 6 | 54 |
| Superior parietal lobule, precuneus | R | 7 | 237 | 4.08 | 14 | −68 | 54 |
| Precuneus | L | 7 | 42 | 3.82 | −6 | −60 | 60 |
| Middle occipital gyrus, precuneus | L | 18, 19 | 78 | 4.02 | −32 | −76 | 28 |
| Inferior parietal lobule | L | 40 | 63 | 3.62 | −42 | −38 | 48 |
| Precuneus | L | 18, 19 | 35 | 3.59 | −18 | −50 | 54 |
| Implicit writing Learned > novel | |||||||
| Cuneus | L | 18, 19 | 15 | 3.54 | −20 | −88 | 2 |
| Middle temporal gyrus, parahippocampal gyrus | L | 30, 42 | 13 | 3.44 | −58 | −54 | −2 |
| Precuneus | L | 7 | 177 | 4.43 | −18 | −76 | 44 |
| Superior frontal gyrus, precentral gyrus | L | 6 | 330 | 4.21 | −16 | −12 | 70 |
| Lentiform nucleus | R | Putamen | 57 | 4.08 | 18 | 14 | 2 |
| Superior parietal lobule, precuneus | R | 7 | 239 | 4.00 | 24 | −60 | 62 |
| Middle frontal gyrus | R | 6 | 30 | 3.69 | 28 | −14 | 50 |
| Declive | R | — | 12 | 3.61 | 18 | −62 | −24 |
| Superior occipital gyrus | L | 19 | 20 | 3.60 | −34 | −82 | 24 |
| Medial frontal gyrus | L | 6 | 18 | 3.58 | −2 | −4 | 54 |
| Cingulate gyrus | L | 32 | 14 | 3.56 | −12 | 10 | 32 |
| Postcentral gyrus | R | 2 | 12 | 3.44 | 46 | −30 | 48 |
Table AII.
Conjunction mask of all three conditions in the implicit writing task
| Anatomical region | H | BA | Voxels | Z‐Score | x | y | z |
|---|---|---|---|---|---|---|---|
| Precuneus, superior parietal lobule | R | 7, 40 | 3545 | inf | 16 | −74 | 52 |
| Superior parietal lobule, precuneus, inferior parietal lobule | L | 7, 40 | 3580 | inf | −30 | −56 | 54 |
| Middle frontal gyrus, cingulate gyrus | L, R | 6, 4, 9, 8 | 3104 | 7.34 | −28 | −10 | 60 |
| Middle occipital gyrus, inferior temporal gyrus, culmen | L | 19, 18, 17 | 1682 | 6.40 | −46 | −68 | −8 |
| Inferior temporal gyrus, culmen, declive | R | 20 | 2287 | 6.27 | 56 | −56 | −18 |
| Precentral gyrus, inferior frontal gyrus | L | 6, 9, 4, 44 | 759 | 5.52 | −48 | 2 | 30 |
| Inferior frontal gyrus | R | 44, 45 | 363 | 4.71 | 54 | 6 | 28 |
| Corpus callosum | — | — | 78 | 3.90 | 0 | 4 | 26 |
| Insula | L | 13 | 44 | 3.72 | −32 | 16 | 6 |
| Insula | R | 13 | 21 | 3.42 | 36 | 18 | 6 |
Contributor Information
Fan Cao, Email: fcao@ntu.edu.sg.
Charles A. Perfetti, Email: perfetti@pitt.edu.
REFERENCES
- Binder JR, McKiernan KA, Parsons ME, Westbury CF, Possing ET, Kaufman JN, Buchanan L ( 2003): Neural correlates of lexical access during visual word recognition. J Cogn Neurosci 15: 372–393. [DOI] [PubMed] [Google Scholar]
- Bolger DJ, Minas J, Burman DD, Booth JR ( 2008): Differential effects of orthographic and phonological consistency in cortex for children with and without reading impairment. Neuropsychologia 46: 3210–3224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger DJ, Perfetti CA, Schneider W ( 2005): Cross‐cultural effect on the brain revisited: Universal structures plus writing system variation. Hum Brain Mapp 25: 92–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TR, Mesulam MM ( 2002): Functional anatomy of intra‐ and cross‐modal lexical tasks. NeuroImage 16: 7–22. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TR, Mesulam MM ( 2003a): The relation between brain activation and lexical performance. Hum Brain Mapp 19: 155–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Zhang L, Choy J, Gitelman DR, Parrish TR, Mesulam MM ( 2003b): Modality‐specific and ‐independent developmental differences in the neural substrate for lexical processing. J Neurolinguistics 16: 383–405. [Google Scholar]
- Booth JR, Lu D, Burman DD, Chou TL, Jin Z, Peng DL, Zhang L, Ding GS, Deng Y, Liu L ( 2006): Specialization of phonological and semantic processing in Chinese word reading. Brain Res 1071: 197–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Lee R, Shu H, Yang Y, Xu G, Li K, Booth JR ( 2010): Cultural constraints on brain development: Evidence from a developmental study of visual word processing in Mandarin Chinese. Cereb Cortex 20: 1223–1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Peng D, Liu L, Jin Z, Fan N, Deng Y, Booth JR ( 2009): Developmental differences of neurocognitive networks for phonological and semantic processing in Chinese word reading. Hum Brain Mapp 30: 797–809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan D, Ho CS, Tsang S‐M, Lee S‐H, Chung K ( 2006): Exploring the reading‐writing connection in Chinese children with dyslexia in Hong Kong. Read Writ 19: 543–561. [Google Scholar]
- Chein JM, Fiez JA ( 2001): Dissociation of verbal working memory system components using a delayed serial recall task. Cereb Cortex 11: 1003–1014. [DOI] [PubMed] [Google Scholar]
- Chen Y, Fu S, Iversen SD, Smith SM, Matthews PM ( 2002): Testing for dual brain processing routes in reading: A direct contrast of Chinese character and pinyin reading using FMRI. J Cogn Neurosci 14: 1088–1098. [DOI] [PubMed] [Google Scholar]
- Chou TL, Booth JR, Bitan T, Burman DD, Bigio JD, Cone NE, Dong L, Cao F ( 2006a): Developmental and skill effects on the neural correlates of semantic processing to visually presented words. Hum Brain Mapp 27: 915–924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TL, Booth JR, Burman DD, Bitan T, Bigio JD, Dong L, Cone NE ( 2006b): Developmental changes in the neural correlates of semantic processing. Neuroimage 29: 1141–1149. [DOI] [PubMed] [Google Scholar]
- Chung K, Ho CS‐H, Chan D, Tsang S‐M, Lee S‐H ( 2010): Cognitive skills and literacy performance of Chinese adolescents with and without dyslexia. Read Writ 16: 2–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng Y, Booth JR, Chou T‐L, Ding G‐S, Peng D‐L ( 2008): Item‐specific and generalization effects on brain activation when learning Chinese characters. Neuropsychologia 46: 1864–1876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eden GF, Jones KM, Cappell K, Gareau L, Wood FB, Zeffiro TA, Dietz NA, Agnew JA, Flowers DL ( 2004): Neural changes following remediation in adult developmental dyslexia. Neuron 44: 411–422. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Balota DA, Raichle ME, Petersen SE ( 1999): Effects of lexicality, frequency, and spelling‐to‐sound consistency on the functional anatomy of reading. Neuron 24: 205–218. [DOI] [PubMed] [Google Scholar]
- Flores d'Arcais G ( 1994): Order of strokes writing as a cue for retrieval in reading Chinese characters. Eur J Cogn Psychol 6: 337–355. [Google Scholar]
- Guan Q, Liu Y, Chan H, Perfetti C ( 2011): Writing strengthens orthography and alphabetic‐coding strengthens phonology in learning to read Chinese. J Educ Psychol 103: 509–522. [Google Scholar]
- Hartwigsen G, Price CJ, Baumgaertner A, Geiss G, Koehnke M, Ulmer S, Siebner HR ( 2010): The right posterior inferior frontal gyrus contributes to phonological word decisions in the healthy brain: Evidence from dual‐site TMS. Neuropsychologia 48: 3155–3163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howard D, Patterson K, Wise R, Brown WD, Friston K, Weiller C, Frackowiak R ( 1992): The cortical localization of the lexicons: Positron emission tomography evidence. Brain 115: 1769–1782. [DOI] [PubMed] [Google Scholar]
- James KH ( 2010): Sensori‐motor experience leads to changes in visual processing in the developing brain. Dev Sci 13: 279–288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James KH, Atwood TP ( 2009): The role of sensorimotor learning in the perception of letter‐like forms: Tracking the causes of neural specialization for letters. Cogn Neuropsychol 26: 91–110. [DOI] [PubMed] [Google Scholar]
- James KH, Gauthier I ( 2006): Letter processing automatically recruits a sensory‐motor brain network. Neuropsychologia 44: 2937–2949. [DOI] [PubMed] [Google Scholar]
- Kucera H, Francis WN ( 1967): Computational Analysis of Present Day American English. Providence, RI: Brown University Press. [Google Scholar]
- Kuo WJ, Yeh TC, Lee JR, Chen LF, Lee PL, Chen SS, Ho LT, Hung DL, Tzeng OJ, Hsieh JC ( 2004): Orthographic and phonological processing of Chinese characters: An fMRI study. Neuroimage 21: 1721–1731. [DOI] [PubMed] [Google Scholar]
- Li Q, Bi, HY , Zhang, JX ( 2010): Neural correlates of the orthographic neighborhood size effect in Chinese. Eur J Neurosci. 32: 866–872. [DOI] [PubMed] [Google Scholar]
- Liu L, Peng D, Ding G, Jin Z, Zhang L, Li K, Chen C ( 2006): Dissociation in the neural basis underlying Chinese tone and vowel production. Neuroimage 29: 515–523. [DOI] [PubMed] [Google Scholar]
- Liu Y, Dunlap S, Fiez J, Perfetti C ( 2007): Evidence for neural accommodation to a writing system following learning. Hum Brain Mapp 28: 1223–1234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Perfetti CA ( 2003): The time course of brain activity in reading English and Chinese: An ERP study of Chinese bilinguals. Hum Brain Mapp 18: 167–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Longcamp M, Anton JL, Roth M, Velay JL ( 2003): Visual presentation of single letters activates a premotor area involved in writing. Neuroimage 19: 1492–1500. [DOI] [PubMed] [Google Scholar]
- Longcamp M, Anton JL, Roth M, Velay JL ( 2005a): Premotor activations in response to visually presented single letters depend on the hand used to write: A study on left‐handers. Neuropsychologia 43: 1801–1809. [DOI] [PubMed] [Google Scholar]
- Longcamp M, Zerbato‐Poudou MT, Velay JL ( 2005b): The influence of writing practice on letter recognition in preschool children: A comparison between handwriting and typing. Acta Psychol (Amst) 119: 67–79. [DOI] [PubMed] [Google Scholar]
- Naka M ( 1998): Repeated writing facilitates children's memory for pseudocharacters and foreign letters. Mem Cognit 26: 804–809. [DOI] [PubMed] [Google Scholar]
- Nelson JR, Liu Y, Fiez J, Perfetti CA ( 2009): Assimilation and accommodation patterns in ventral occipitotemporal cortex in learning a second writing system. Hum Brain Mapp 30: 810–820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pak A, Cheng‐Lai A, Tso I, Shu H, Li W, Anderson RC ( 2005): Visual chunking skills of Hong Kong children. Read Writ 18: 437–454. [Google Scholar]
- Parkinson J, Dyson BJ, Khurana B ( 2010): Line by line: The ERP correlates of stroke order priming in letters. Exp Brain Res 201: 575–586. [DOI] [PubMed] [Google Scholar]
- Paulesu E, McCrory E, Fazio F, Menoncello L, Brunswick N, Cappa SF, Cotelli M, Cossu G, Corte F, Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith CD, Frith U ( 2000): A cultural effect on brain function. Nat Neurosci 3: 91–96. [DOI] [PubMed] [Google Scholar]
- Perfetti CA, Liu Y, Fiez J, Nelson J, Bolger DJ, Tan L‐H ( 2007): Reading in two writing systems: Accommodation and assimilation in the brain's reading network. Biling Lang Cogn 10: 131–146. [Google Scholar]
- Perfetti CA, Liu Y, Tan LH ( 2005): The lexical constituency model: Some implications of research on Chinese for general theories of reading. Psychol Rev 112: 43–59. [DOI] [PubMed] [Google Scholar]
- Poldrack RA, Temple E, Protopapas A, Nagarajan S, Tallal P, Merzenich M, Gabrieli JD ( 2001): Relations between the neural bases of dynamic auditory processing and phonological processing: Evidence from fMRI. J Cogn Neurosci 13: 687–697. [DOI] [PubMed] [Google Scholar]
- Prinz W ( 1997): Perception and action planning. Eur J Cogn Psychol. 9: 129–154. [Google Scholar]
- Rayner K, Foorman BR, Perfetti CA, Pesetsky D, Seidenberg MS ( 2001): How psychological science informs the teaching of reading. Psychol Sci 2( 2 Suppl ): 31–74. [DOI] [PubMed] [Google Scholar]
- Shadmehr R, Holcomb HH ( 1997): Neural correlates of motor memory consolidation. Science 277: 821–825. [DOI] [PubMed] [Google Scholar]
- Shaywitz BA, Shaywitz SE, Pugh KR, Fulbright RK, Skudlarski P, Mencl WE, Constable RT, Marchione KE, Fletcher JM, Klorman R, Lacadie C, Gore JC ( 2001): The functional neural architecture of components of attention in language‐processing tasks. Neuroimage 13: 601–612. [DOI] [PubMed] [Google Scholar]
- Siok WT, Fletcher P ( 2001): The role of phonological awareness and visual–orthographic skills in Chinese reading acquisition. Dev Psychol 37: 886–899. [PubMed] [Google Scholar]
- Tan LH, Feng CM, Fox PT, Gao J‐H ( 2001): An fMRI study with written Chinese. Neuroreport 12: 83–88. [DOI] [PubMed] [Google Scholar]
- Tan LH, Laird AR, Karl L, Fox PT ( 2005a): Neuroanatomical correlates of phonological processing of Chinese characters and alphabetic words: A meta‐analysis. Hum Brain Mapp 25: 83–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Spinks JA, Eden GF, Perfetti CA, Siok WT ( 2005b): Reading depends on writing, in Chinese. Proc Natl Acad Sci USA 102: 8781–8785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Spinks JA, Feng CM, Siok WT, Perfetti CA, Xiong J, Fox PT, Gao JH, Kalogirou E ( 2003): Neural systems of second language reading are shaped by native language. Hum Brain Mapp 18: 158–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Temple E, Deutsch GK, Poldrack RA, Miller SL, Tallal P, Merzenich MM, Gabrieli JD ( 2003): Neural deficits in children with dyslexia ameliorated by behavioral remediation: Evidence from functional MRI. Proc Natl Acad Sci USA 100: 2860–2865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vellutino FR, Fletcher JM, Snowling MJ, Scanlon DM ( 2004): Specific reading disability (dyslexia): What have we learned in the past four decades? J Child Psychol Psychiatry 45: 2–40. [DOI] [PubMed] [Google Scholar]
- Wang Y, Sereno JA, Jongman A, Hirsch J ( 2003): fMRI evidence for cortical modification during learning of Mandarin lexical tone. J Cogn Neurosci 15: 1019–1027. [DOI] [PubMed] [Google Scholar]
- Wu X, Li W, Anderson R ( 1999): Reading instruction in China. J Curric Stud 31: 571–586. [Google Scholar]
- Yu H, Gong L, Qiu Y, Zhou X ( 2011): Seeing Chinese characters in action: An fMRI study of the perception of writing sequences. Brain Lang 119: 60–67. [DOI] [PubMed] [Google Scholar]
- Ziegler JC, Goswami U ( 2005): Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychol Bull 131: 3–29. [DOI] [PubMed] [Google Scholar]