

Abstract
A wide range of essential reasoning tasks rely on contradiction identification, a cornerstone of human rationality, communication and debate founded on the inversion of the logical operators “Every” and “Some.” A high‐density electroencephalographic (EEG) study was performed in 11 normal young adults. The cerebral network involved in the identification of contradiction included the orbito‐frontal and anterior‐cingulate cortices and the temporo‐polar cortices. The event‐related dynamic of this network showed an early negative deflection lasting 500 ms after sentence presentation. This was followed by a positive deflection lasting 1.5 s, which was different for the two logical operators. A lesser degree of network activation (either in neuron number or their level of phase locking or both) occurred while processing statements with “Some,” suggesting that this was a relatively simpler scenario with one example to be figured out, instead of the many examples or the absence of a counterexample searched for while processing statements with “Every.” A self‐generated reward system seemed to resonate the recruited circuitry when the contradictory task is successfully completed. Hum Brain Mapp, 2009. © 2009 Wiley‐Liss, Inc.
Keywords: contradictory reasoning, electroencephalography (EEG), independent component analysis (ICA), logical operators
INTRODUCTION
Reasoning is one of the core expressions of human intelligence and is pivotal to effective social interactions, as well as to the solution of everyday practical problems. The identification of contradiction, which is crucial for logical reasoning, plays an important role in communication, understanding, and learning. Western culture, which largely derives its models from Aristotelian logic, relies on a model that polarizes contradictory perspectives, i.e., deciding if two apparently divergent propositions are contradictory. In philosophical and historical perspective, contradictions are embedded in the social structure of society and can inherently lead to economic and cultural crises and eventually revolutions. In conversations, when trying to prove the contradiction of a general statement we frequently search for a counterexample. To refute the statement “Every man is tall,” it is therefore sufficient to state “Napoleon is short.” In logic, the same statement would be contradicted with the formulation “Some men are short.” Above and beyond being fundamental structures in logic, “Every” and “Some” are key conceptual categories of human reasoning. We rely on these operators when generalizing properties of objects or phenomena or alternatively to describe their specific attributes. “Every” and “Some,” pervade natural language, provide a means of communicating quantities and are central to conducting a debate. A profound cultural awareness of the importance of these categories promotes communication and dialogue aptitude across the whole of society.
Considerable progress has been made over the past decade in understanding the neural basis of human reasoning [Goel, 2007; Kroger et al., 2008] with the aid of noninvasive brain imaging tools. Techniques like functional magnetic resonance (fMRI) [Goel, 2003, 2007; Goel and Dolan, 2001, 2003, 2004; Goel et al., 2000, 2007; Newstead et al., 1997; Prado and Noveck, 2007] and electroencephalography (EEG) [Luo et al., 2008; Prado et al., 2008; Qiu et al., 2007] offer a window on brain function that is complementary to more traditional measurements of psychophysical performance, such as reaction times and other psychometric parameters. Extensive and distributed brain networks organized with a high degree of modularity have been so far identified as sustaining logical reasoning, with a consistent involvement of frontal areas [Goel, 2007; Greene et al., 2004].
In this study we have used high resolution EEG to investigate the electrophysiological correlates of cognitive processing while identifying whether a conclusion correctly contradicted a premise. Given the complexity of the putative processing networks, we decided to approach the reasoning network recognition through independent component analysis (ICA). Surprisingly, to researchers accustomed to thinking that EEG sources can only be separated by solving the biophysical inverse problem, ICA does not require any “a priori” assumptions on network properties or head model during the separation step. This algorithm is able to identify the brain sources supporting the investigated function by exploiting the most accurate information provided by the electrophysiological techniques, i.e. the dynamical properties of the recorded signal [Barbati et al., 2004; Jung et al., 2000; Makeig et al., 2002; Porcaro et al., 2009]. Beyond separating stereotyped nonbrain artifact signals including eye movements, line noise, cardiac artifacts, and muscle activities [Makeig et al., 2004], ICA can identify large or small neuronal pool activities with diverse physiological and functional roles [Hyvärinen and Oja, 2000; Makeig et al., 1999; Vorobyov and Cichocki, 2002]. Appropriate procedures must be applied to identify the ICs representing brain sources of interest, generally with a set of ICs corresponding to one brain source with specific spatio‐temporal properties. After IC separation on the basis of time signal statistical properties, and after the second step of IC identification on the basis of the expected spatio‐temporal characteristics, proper algorithms must be run to characterize the 3D configuration of these sources. In other words, after IC separation, ICA allows the use of source localization algorithms having isolated the field distribution generated by the specific source of interest.
We used typical forms within the Aristotelian categorical proposition structure in the context of a naturalistic debate, presenting premise‐conclusion pairs like “Every man is mortal—Some men are immortal.” The two sentences were presented in a dialogue‐like fashion, with a proponent formulating a statement, and an opponent attempting its refutation. The subject was asked to recognize the correctness of the opponent refutation independent of the correctness of the sentence content (see Fig. 1). We investigated whether identification of contradiction corresponded to different activation dynamics for the universal (“Every”), as opposed to the particular (“Some”) operator. To this end, the 200 premise‐conclusion couples were evenly divided between contradictory (C) and not contradictory (NC), half of them presenting a premise with the “Every” logical operator (UNIVERSAL) and the other half with the “Some” logical operator (PARTICULAR).
Figure 1.
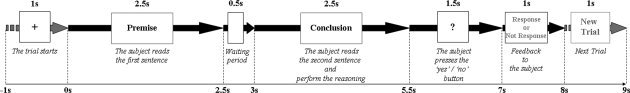
Experimental setup. The beginning of the trial was signaled by a “+” appearing in the centre of the screen for 1 s, followed by the presentation of the premise categorical proposition for 2.5 s. Thereafter the sentence disappeared for 0.5 s (Waiting period), followed by the presentation of the conclusion categorical proposition for 2.5 s. Then, a question mark appeared for 1.5 s, requiring the subject to press one push button if the conclusion contradicted the premise and a different push button if the conclusion did not. If they were unable to make a clear choice, they were told to omit the response (i.e., to not push either button). Finally, a feedback advised the subject whether any button had been pressed or not, with no indication about the correctness of the contradiction identification. “New Trial” appearing on the screen advised the subject that the next trial was starting.
MATERIALS AND METHODS
Subjects
Subjects consisted of 11 healthy native Italian speakers (mean age: 30.5 years; range: 24–38 years; five men, six women) took part in this experiment. None of the subjects had specific background knowledge or experience in logic. Subjects gave their informed written consent after the nature of the study was explained to them. The study occurred at Aston University and was authorized by the Aston University Ethics Committee. All subjects were healthy, right‐handed, and had normal or corrected‐to‐normal vision.
EEG Recordings
Scalp EEG signals were recorded continuously during the protocol using an EGI data acquisition system (Electrical Geodesics, Eugene, OR, USA; http://www.egi.com) with the third generation of EGI dense array net, the HydroCel Geodesic Sensor Net® [128‐channel HCGSN—Tucker, 1993]. Gain and zero calibration were performed before the start of each EEG recording; channel impedances were kept below 50 kΩ for all net sensors [Ferree et al., 2001]. EEG amplified signals were sampled at 500 Hz (pre‐sampling analogical filter 0.1–200 Hz) and collected for off‐line processing. All channels were referenced to the vertex (Cz) electrode.
Stimuli
Subjects were comfortably seated at a distance of 1 m from the front of a 19‐in. monitor. Stimulus presentation was carried out via a Dell PC using E‐Prime experimental programming software (Psychology Software Tools; http://www.pstnet.com), which provides triggers for the EGI recording apparatus with highly accurate timing. The protocol contained 200 pairs of sentences, evenly divided into the following four forms: 50 Every Some‐Contradictory (ES‐C), 50 Some Every‐Contradictory (SE‐C), and 50 Every Some‐Not Contradictory (ES‐NC), 50 Some Every‐Not Contradictory (SE‐NC). In the ES‐C and ES‐NC the premise is universal (E: Every) and the conclusion is particular (S: Some), with the conclusion contradictory for the first set (ES‐C) and not contradictory for the other (ES‐NC). Similar conditions were used for the SE‐C and SE‐NC.
For example, one set of the four conditions is provided in Table I. The four conditions were randomly presented.
Table I.
Stimuli example
| Contradictoriness | Logical operator | |
|---|---|---|
| Universal | Particular | |
| Contradictory | Every man is mortal | Some men are mortal |
| Some men are immortal | Every man is immortal | |
| Not contradictory | Every man is mortal | Some men are mortal |
| Some men are mortal | Every man is mortal | |
An example of the four experimental conditions. ES‐C: Universal‐Contradictory; ES‐NC: Universal‐Not Contradictory; SE‐C: Particular‐Contradictory; SE‐NC: Particular‐Not Contradictory.
Experimental Setup
Prior to recording, the experiment was described to the subjects (see Fig. 1) and they were given a test training session. Subjects were asked to evaluate whether the conclusion was contradictory or not, with respect to the premise independent of the correctness of the sentence content. After the conclusion was reached, and at the appearance of a go signal (a question mark), subjects had to press one push button if the conclusion contradicted the premise and a different push button if the conclusion did not. If they were unable to make a clear choice, they were told to omit the response (i.e., to not push either button).
Data Analysis
The data were low pass filtered (100 Hz) prior to the off‐line analysis. The analysis strategy aimed at extracting signals generated only in response to the task. For this purpose, we assumed the set of EEG signals to be generated by the mixing model:
| (1) |
where t = 0,1,2,… is the discrete sampling time; x(t) = [x 1(t),…,x m(t)] is the m‐dimensional vector of the observed signal recorded by m sensors; A is an n ≤ m (with) unknown full‐rank mixing matrix; s(t) = [s 1(t),…,s n(t)]T is the n‐dimensional unknown vector of the sources (see Fig. 2). The model is approached by processing sensor signals by an ICA demixing system described in the form:
| (2) |
where IC(t) = [IC 1(t),…,IC n(t)]T is the n‐dimensional vector of the estimated Independent Components (ICs) and W is the separation matrix, i.e., the estimate of the inverse of the unknown mixing matrix A, up to permutation and scaling:
| (3) |
Figure 2.
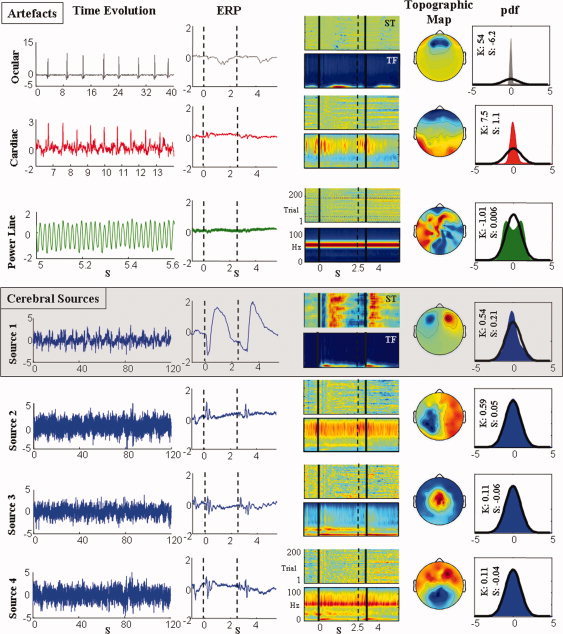
Reasoning Network Identification. For representative ICs in one paradigmatic subject, the criteria used to classify them as artifacts (upper panel) and cerebral sources (bottom panel) are shown. From the left column: an exemplificative segment of IC signal (Time Evolution), the average triggered by the premise presentation across all 200 trails (ERP); Single Trail image with each epoch including premise‐conclusion pair represented in color code in the abscissa with successive epochs in the ordinate (ST); and the Time Frequency plot representing each time point in the abscissa, the color coded spectral content in the ordinate (TF), vertical solid lines indicate the presentation of each sentence, the dashed line the end of the premise; spatial distribution obtained by representing the corresponding IC weights (Topographic map); the probability density function of the IC signal (pdf) with the black line indicating the normal probability density, the values of kurtosis (K) and skewness (S) are provided. As the ICA procedure performs whitened preprocessing, each quantity is expressed in arbitrary unit (a.u.). Among the artifacts, the ocular artifact is in the first row, the cardiac artifact is in the second and line power is in the third. Among IC classified as cerebral sources, that represented in the first row, the only one showing clear event‐related activity later than 150 ms, was consequently selected as the Reasoning Network. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
We used the FastICA algorithm proposed by Hyvärinen [1999] and Hyvärinen et al. [2001]
In the case of a high number of channels (128 in our case), a direct extraction of all the ICs would have been extremely time consuming and component selection extremely challenging. We pursued a dimensionality reduction by applying an optimized procedure to select a k ICs‐subset such that the corresponding explained variance will be at least 95% [Salustri et al., 2005]. This was proven to be a suitable procedure for subspace reduction and avoided the time consuming estimation of all ICs in the case of high space dimensionality corresponding to the high number of channels. In our case, on average, 39 ICs (range 27–49) resulted in a mean explained variance ± standard deviation of 97.58% ± 1.12%.
Selection criteria of reasoning network (RN) ICs
As determined by the improvements of ICA model applications, introduced for example in the context of fMRI [Beckmann and Smith, 2004] and Foetal Magnetoencephalography [Porcaro et al., 2006], we selected among the above described IC subset (of about 39 ICs) of those components showing appropriate spatio‐temporal characteristics. We required significant event‐related responses triggered by the conclusion presentation. To this end, each IC dynamic was averaged, triggered on the second sentence presentation for the time window including the premise, and lasting until the start of the next trial. As we used a constant inter‐stimulus interval between the premise and conclusion (3 s) in our paradigm, a single average evidenced the phase locked response of the IC to both sentences. The single trial and time‐frequency behaviors, as well as the topographical distribution, were used to identify ICs of interest (RN ICs, Fig. 2).
Reasoning network localization
After RN identification, its position was estimated in each subject by retro projecting only the RN ICs and obtaining the electric potential distribution:
| (4) |
where A k is the estimated mixing vector [matrix A of Eq. (1)] for the source IC k and EEGreck is the resulting IC k retro‐projection on the channels space. We submitted EEGreck to a source localization algorithm (sLORETA) [Pascual‐Marqui, 2002] as implemented on CURRY® software (http://www.neuroscan.com).
Reasoning network dynamics
The RN IC dynamics were studied in response to the presentation of the premise and the conclusion, i.e., analyzing the two time periods lasting (−0.5, 2.5) s, with 0 being the sentence presentation. The RN dynamics were studied in the four different experimental conditions ES‐C, ES‐NC, SE‐C, and SE‐NC, referring to trials with correct identification.
Statistical Analysis
Behavioral data
While the distribution of errors did not differ from a Gaussian fit (Kolmogorov‐Smirnov P > 0.200 consistently), the distribution of reaction times differed (P < 0.001) and they were logarithmically transformed obtaining a suitable fit (P > 0.200 consistently). Log‐transformed reaction times were analyzed by univariate general linear model (GLM) with Logical Operator (UNIVERSAL, PARTICULAR) and Contradiction (CONTRADICTORY, NOT CONTRADICTORY) as fixed factors and Subject as Random factor. Percentages of errors were submitted to an analysis of variance (ANOVA) for repeated measures with Logical Operator (UNIVERSAL, PARTICULAR) and Contradiction (CONTRADICTORY, NOT CONTRADICTORY) as within‐subject factors. Absence of either Logical Operator or Contradiction main factor for Errors was checked, indicating that the accuracy in performing the tasks did not change in relationship to the presentation of the different sentences.
Reasoning network behavior
The dynamics of the RN‐evoked response were submitted to statistical analyses to assess amplitude differences in different tasks. The mean amplitude areas included between the onset of the early negative stage and the return to baseline and the positive stage onset and the return to baseline were considered for RN‐evoked response, both following the premise and conclusion presentations. Since the distributions of these parameters did not differ from a Gaussian fit (Kolmogorov‐Smirnov P > 0.200 consistently), they were submitted to an analysis of variance (ANOVA) without correcting transformations. The full model included Sentence (PREMISE, CONCLUSION), Logical Operator (UNIVERSAL, PARTICULAR), and Contradiction (CONTRADICTORY, NOT CONTRADICTORY) as within‐subject factors.
A result was reported only if it was statistically significant (P < 0.050 Greenhouse‐Geisser corrected whenever the sphericity assumption for ANOVA was violated).
RESULTS
Behavioral Data
Subjects performed the PARTICULAR and UNIVERSAL tasks with comparable accuracy both for the contradictory and not contradictory conditions, as demonstrated by the absence of any main effect from the ANOVA design for the number of errors (P > 0.200) (Table II).
Table II.
Behavioral data
| Contradictoriness | Logical operator | |
|---|---|---|
| Universal | Particular | |
| Correct responses (%) | ||
| Contradictory | 98.0 ± 3.2 | 97.5 ± 1.9 |
| Not contradictory | 97.9 ± 2.2 | 98.5 ± 1.7 |
| Reaction times (log s) | ||
| Contradictory | 2.74 ± 0.007 (550 ms) | 2.69 ± 0.006 (490 ms) |
| Not contradictory | 2.71 ± 0.006 (513 ms) | 2.70 ± 0.006 (501 ms) |
In each of the four experimental tasks, mean across subjects (±S.D.) of the percentage of correct responses and of the reaction times, i.e. the time between the question mark appearance and the subject response, after logarithmic transformation. In squared parenthesis the exponential inverse transformation of the mean is indicated to give an idea of reaction time dimension (ms).
On the contrary, reaction times showed a strong main Logical Operator effect [logarithmically transformed time F(1, 10.098) = 32.535, P < 0.001, Table II], with longer reaction times for UNIVERSAL tasks than for PARTICULAR ones. A Contradictory effect was not observed (P = 0.441).
Identification of Reasoning Network (RN)
In every subject a single independent component (IC) was associated with an event‐related response occurring at latencies subsequent to those traditionally associated with primary sensory processing (see Fig. 2). This IC accounted for the majority of the variance of the whole non‐artifactual scalp EEG signals along the whole 40‐min session in each subject (Table III, second column). As said before, we refer to this IC as the reasoning network (RN).
Table III.
Structures involved in reasoning networks
| Subject | ICs EV (%) | Temporo‐polar cortex | Orbitofrontal cortex (OFC) | Anterior cingulated cortex (ACC) | |||
|---|---|---|---|---|---|---|---|
| L | R | L | R | L | R | ||
| AI | 60 | 20; 21; 38 | 47 | ||||
| AS | 82 | 38 | 21; 22; 38 | 10; 11; 47 | 10;11; 47 | 32 | |
| CP | 84 | 38 | 21; 22; 38 | 11; 47 | 11; 47 | ||
| DB | 82 | 38 | 20; 21; 22; 38 | 10;11; 47 | 10; 11; 47 | 32 | 32 |
| DF | 38 | 21; 38 | 20; 21; 38 | ||||
| ET | 88 | 20; 21; 38 | 20; 21; 38 | 10; 11; 47 | 10; 11; 47 | 32 | 32 |
| FS | 56 | 21; 38 | 38 | 10; 11; 47 | 10; 11; 47 | 32 | 32 |
| GDL | 41 | 38 | 20; 21; 22; 38 | 11; 47 | 11; 47 | 32 | 32 |
| LDL | 62 | 21; 38 | 21; 38 | 11; 47 | 11; 47 | 32 | 32 |
| MTM | 87 | 20; 21; 38 | 20; 21; 22; 38 | 11; 47 | 11; 47 | ||
| SC | 90 | 20; 21; 22; 38 | 20; 21; 22; 38 | 10; 11; 47 | 10; 11; 47 | 32 | 32 |
| Average | 70 | 21; 38 | 21; 38 | 10; 11; 47 | 10; 11; 47 | 32 | 32 |
For each subject, the variance of the signal from all channels throughout the 40 min explained by the identified IC is expressed as percentage of the total variance (IC EV). Structures involved within the network are expressed by the corresponding Brodmann areas classified by the anatomical site.
Consistently across subjects, the RN included the bilateral Temporo‐polar cortex (middle temporal gyrus Brodmann Area (BA) 21 and the most rostral part of the superior and middle temporal gyri BA 38), the bilateral Orbitofrontal cortex (BA 10, 11, and 47), and the bilateral Anterior Cingulate Cortex (BA 32) (Fig. 3, Table III).
Figure 3.
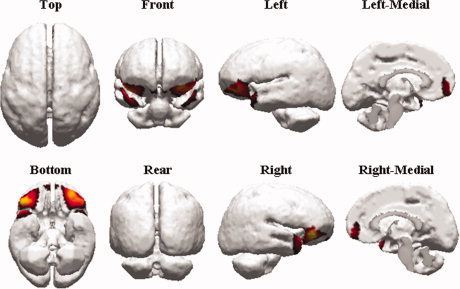
Reasoning Network position. Topography of the reasoning network, averaged across subjects, localized as described in the methods section on a standard model. It is to be noted that the potential distribution obtained by retro‐projecting only one IC is time‐invariant up to a scale factor, consequently, the subtending current distribution shape is time‐independent. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Reasoning Network Behavior
At the presentation of the two sentences (Fig. 4a,b), RN activated much more strongly after the conclusion (mean 1.62 ± S.D. 0.69 averaged over the four conditions) than the premise (1.33 ± 0.53, paired t test t(10) = −3.396, P = 0.007). Moreover, the processing of the conclusion lasted much longer (1663 ± 651 ms) than that of the premise (1233 ± 454 ms, t(10) = −3.842, P = 0.003). Remarkably, the RN activated much faster after the conclusion presentation (280 ± 87 ms) than after the premise (481 ± 101 ms, t(10) = 12.225, P < 0.001). This last feature underlines the fact that the reasoning for contradiction identification starts with the premise processing. On this basis, we developed the statistical modeling to include both premise and conclusion as a whole reasoning process.
Figure 4.
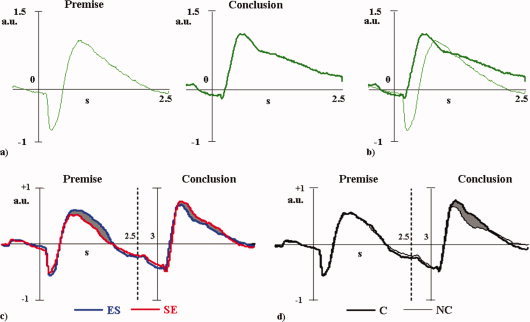
Reasoning Network Behavior. (a) RN activity after Premise and Conclusion presentations during the time periods lasting (−0.5, 2.5) s, 0 being the sentence presentation. (b) The RN activity after Premise and Conclusion are superimposed to facilitate comparison of their dynamics. (c) RN activity compared during the UNIVERSAL (ES‐C, ES‐NC) vs. PARTICULAR (SE‐C, SE‐NC) premise‐conclusion pairs. The grey areas indicate significant change between the two task groups. (d) As case (c) for the Contradictory vs. Not Contradictory task comparison. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
As for the involved anatomical structures mentioned in the above section, the RN time‐frequency characteristics were also highly consistent across subjects. RN dynamics consistently showed an early negative stage in the 150–500 ms time period after sentence presentation (Early Negative Stage) across trials (Fig. 5, first row). With a similar inter‐trial stability, a positive stage was subsequently found reaching maximal amplitude between 500 and 2,000 ms after appearance of both the premise and the conclusion sentences (Positive Stage). In terms of spectral characteristics, the RN activity was strongly represented in the 0.1–15 Hz frequency range (Fig. 5 second row). Moreover, a clear dynamic of oscillating activity in the gamma band was evident for the duration of the task (Fig. 5 second row).
Figure 5.
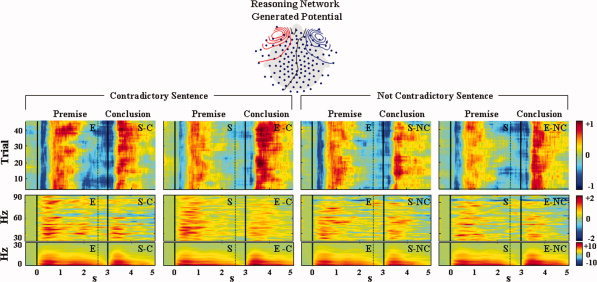
Reasoning Network time‐frequency behavior. Up: Average across subjects of the topographical distribution of the electric potential generated by the RN. Middle: Average across subjects of single trail image as described in Figure 2 showing the RN time evolution along the premise‐conclusion presentations corresponding to the four experimental conditions. Bottom: Again for each of the four tasks, time frequency plot as described in Figure 2, averaged across subjects and referred to as the 500 ms pre‐premise period. The green color code represents values nonsignificantly differing from 0 using a bootstrap technique with threshold at P = 0.01. Note that different scales hold for the oscillatory activity amplitude in the two (0–30) Hz and (30–90) Hz frequency bands. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Early negative stage
Full model ANOVA did not display any main or interaction effect.
Positive stage
Full model ANOVA indicated clear interaction effects Sentence × Contradiction [F(1,10) = 14.879, P = 0.003] and Sentence × Logical Operator [F(1,10) = 5.854, P = 0.036], indicating that the reasoning network was activated in a different way during the presentation of the premise and the conclusion depending on the presence or absence of Contradiction, and whether a UNIVERSAL or PARTICULAR premise had to be contradicted. To better investigate the subtended phenomena, the two sentences were submitted to respective ANOVA reduced models.
Premise
While a Contradictory effect was completely absent (P = 0.878), a strong Logical Operator effect was found [F(1,10) = 10.616, P = 0.009], corresponding to a stronger reasoning network activation during the presentation of UNIVERSAL as opposed to the PARTICULAR sentences (Fig. 4c).
CONCLUSION
A clear Contradictory effect appeared [F(1,10) = 7.402, P = 0.022], corresponding to a stronger reasoning network activation when the CONTRADICTORY conclusions were presented than when the NOT CONTRADICTORY ones were (Fig. 4d). Logical Operator effect did not reach statistical significant (p = 0.193).
To investigate, in greater detail, dynamic properties of different stages of information processing, the RN activity periods during the presentation of premises and conclusions were subdivided into earlier ascending and later descending activation phases (Fig. 4c,d), by introducing into the statistical design the corresponding within‐subject factor Phase (ASCENDING, DESCENDING). The amplitude of RN activity was calculated as the mean area between activation onset and the maximum for the ascending phase, and between the maximum and the return to baseline for the descending phase.
The analysis of the premise showed that the Logical Operator effect appeared only during the descending phase [F(1,10) = 5.148, P = 0.047] (Fig. 4c).
The analysis of the conclusion showed, in addition to the above‐mentioned Contradictory effect, the interaction Phase × Logical Operator effect [F(1,10) = 4.942, P = 0.050]. While RN activations did not differ during the ASCENDING phase, during the DESCENDING period UNIVERSAL conclusions induced stronger RN activation than PARTICULAR ones [reduced model in descending phase F(1,10) = 5.448, P = 0.042, Fig. 4c,d].
DISCUSSION
Contradiction is a cornerstone concept in logic and is considered to be the basis for any reasoning process. In more general terms, contradiction is inherently associated with a “truth” criterion. Contradictory entities are present in every significant human experience as the two faces of a coin: light and darkness, joy and sorrow, passion and suffering, life and death. Our study on contradiction offers a novel angle to the discussion on a human reasoning aspect intimately connected with the historical tradition of mathematicians, philosophers, and physicists and sheds light on a fundamental aspect of cognition.
RN Activity Universal > Particular
During reasoning, to identify whether a conclusion contradicts a premise, our reasoning network shows a significantly greater activation when a UNIVERSAL statement is processed. The more limited computational load required to process PARTICULAR statements could be due to the more direct process required to explore a single example in the case of a “Some” sentence, as opposed to the multiple examples or the absence of a counterexample searched for during processing of an “Every” sentence. The longer reaction times found for UNIVERSAL as opposed to PARTICULAR statements seem to support this hypothesis (Table II). When reasoning about familiar and concrete situations—as opposed to unfamiliar and abstract ones like “Every A is B …”—our brain automatically utilizes, in parallel with formal methods, situation‐specific heuristics based on prior knowledge and experience [Goel et al., 2007]. Previous evidence suggests that the response of the frontal‐temporal system to familiar situations is—at least to some degree—content specific. In particular, the middle temporal lobe regions as part of object‐based knowledge networks are more active when reasoning on statements like “Every tree is tall” and were structures well represented in our reasoning network. In our experiment, when processing statements commonly encountered in natural language, the fronto‐temporal system was challenged more by universal than particular forms. Our result suggested that in processing familiar sentences, generalization requirements recruit a wider network than would be required when assigning specific attributes within a limited field of action.
In spite of an identical inversion required by both logical operators, we found that universal sentences required stronger reasoning network activation than particular ones. This could be explained by a different internal representation of the logical operators themselves rather than by the effect of the applied rule.
RN Activity Dynamics
The early negative stage of the reasoning network event‐related dynamics (Figs. 4 and 5), peaking around 150–500 ms after sentence presentation in correspondence with the reading phase, was remarkably consistent with intra‐cortical recordings from lateral prefrontal cortex during word reading [Lachaux et al., 2008]. This seems to support the idea that the reading phase is parallel to cognitive processes possibly mediated by a coordinated interaction between regional synchronizations and desynchronizations of neuronal oscillations, reflecting long‐range network engagement [Lachaux et al., 2008]. This initial phase is followed by a strong positive stage lasting ∼1.5 s when the presence of a contradictory conclusion is identified. This is associated with a consistent pattern of an ascending phase lasting about 300 ms and a successive decay starting at about 800–900 ms from sentence presentation. This latter phase corresponds to the period in which the logical operator is classified during premises and conclusions.
RN Activity Contradictory > Not Contradictory
Subjects had to target the contradictory sentences and these evoked stronger reasoning network activation than the control not contradictory ones. As opposed to what is observed when comparing Universal and Particular sentences, reaction times were similar for contradictory and not contradictory pairs, suggesting that the higher RN amplitude does not reflect a higher processing load during the identification of contradictoriness. The activated reasoning network partially overlaps with the cortical component of the central system sustaining decision‐making, strongly involving the orbitofrontal and cingulate cortices. It has, in fact, been recently determined that, in addition to the striatum and dopamine systems, activity in these regions is predictive of the choices of animals and people [Rushworth and Behrens, 2008]. In particular, enhanced activity in the orbitofrontal cortex (OFC) is associated with risk‐taking behavior [Doya, 2008]. The anterior cingulate cortex (ACC) plays an important role in motivating subjects to act based on a cost/benefit estimate [Denk et al., 2005] The ACC has a strong topographical specificity, with the gyral component implicated in social learning and the sulcal in reward‐based learning [Rudebeck et al., 2006; Walton et al., 2002]. In our experiment based on identifying contradictions, stronger sulcal ACC and OFC activities were found while deciding in favor of the required task. The feedback consequent to the positive and negative response was the same; the same behavioral response was required as a consequence of the decision and subjects performed with similar reaction times in both cases. The absence of any external feedback after the identification of the contradictory form suggests that neural structures can develop an intrinsic ability to enhance synchronization phenomena as a consequence of performing the required task, without the need of a reward from the external environment. Our data could be explained by hypothesizing that human cognitive structures involved in reward‐based training and decision‐making are able—as a result of reinforcement‐based learning to intrinsically induce an activity that had previously been dependent on external reward during the learning process This internal ability is further supported by recent hypotheses of the rostral prefrontal cortex (PFC, approximating BA 10) as the key attention‐switching structure focusing either on environmental stimuli, or by contrast, on self‐generated or maintained representations [i.e. the “thoughts in our head,” “gateway hypothesis,” Burgess et al., 2007].
RN Constituted by “Expected Areas”
We found reasoning network activation in every subject. This involved the temporo‐polar, orbito‐frontal, and anterior cingulate cortices. Moreover, single trial analysis showed strong repeatability of network recruitment throughout the task performance, indicating a fundamental role of searching for contradiction in human reasoning. Cognitive neuroscience data are not in keeping with a unitary system being the basis of logical reasoning, suggesting instead a distributed system that includes dynamic reconfiguration of the fronto‐temporal and anterior cingulate regions in response to specific tasks and environmental cues [Goel, 2007]. Language structures are not included in the networks activated by our paradigm, probably because reasoning about familiar situations automatically utilizes situation‐specific heuristics, mainly involving orbito‐frontal cortex and inferior‐middle temporal regions [Goel, et al. 2007]. Since in all subjects a single component explained the majority of the signal power throughout the experiment, and very simple common words were used for the reasoning, it is conceivable that structures processing semantic‐syntactic content were not significantly engaged.
RN Identification Procedure
Brain electric fields recordable from the scalp through EEG are macroscopic post‐synaptic potentials created by clusters of apical dendrites of neocortical pyramidal cells firing synchronously [Nunez and Srinivasan, 2006]. The absence of any appreciable delay in the scalp sensor signals with respect to neural current dynamics and frequency components, such that quasi‐static approximation of Maxwell equations holds throughout the whole spectrum of interest, strongly supports the superposition principle, according to which the relation between EEG scalp potentials and generating cerebral sources may be reasonably approximated by a system of linear equations [Sarvas, 1987]. Under these conditions the use of a blind source separation technique such as ICA seems most advantageous for identifying cerebral activities of interest. ICA decomposes the data into sources with independent time courses and scalp maps, without taking into account any information about head and cerebral current physics and geometry. ICA, combined with time/frequency analysis and trial‐by‐trial visualization, is able to recover components indexing physiologically distinct processes. Moreover, the complexity of networks sustaining reasoning strengthens the suitability of source identification methods based on source dynamic behavior instead of the mere localization of their spatial properties.
In conclusion, a consistent reasoning network was clearly identified in each healthy volunteer that involved the temporo‐polar, orbito‐frontal and anterior cingulated cortices. After an early negative period lasting about 500 ms, this network was more activated for about 1.5 s, when processing universal statements and contradictory conclusions. Simpler cerebral processing in the heuristic figuring of a single example instead of many, or searching for the absence of a counterexample, could be the reason why the logical operator “Some” induced smaller reasoning network activation than “Every.” A self‐generated reward system appeared to be present in adult humans, with the ability to resonate the recruited circuitry when a task is accomplished in the required form.
Acknowledgements
The authors thank Prof. Vito Michele Abrusci and Prof. Claudia Casadio for their continuous support and for valuable comments and Dr. Paolo Roselli for scientific discussion.
REFERENCES
- Barbati G, Porcaro C, Zappasodi F, Rossini PM, Tecchio F ( 2004): Optimization of an independent component analysis approach for artifact identification and removal in magnetoencephalographic signals. Clin Neurophysiol 115: 1220–1232. [DOI] [PubMed] [Google Scholar]
- Beckmann CF, Smith SM ( 2004): Probabilistic independent component analysis for functional magnetic resonance imaging. IEEE Trans Med Image 23: 137–152. [DOI] [PubMed] [Google Scholar]
- Burgess PW, Dumontheil I, Gilbert SJ ( 2007): The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends Cogn Sci 7: 290–298. [DOI] [PubMed] [Google Scholar]
- Denk F, Walton ME, Jennings KA, Sharp T, Rushworth MF, Bannerman DM ( 2005): Differential involvement of serotonin and dopamine systems in cost‐benefit decisions about delay or effort. Psychopharmacology 179: 587–596. [DOI] [PubMed] [Google Scholar]
- Doya K ( 2008): Modulators of decision making. Nat Neurosci 11: 410–416. [DOI] [PubMed] [Google Scholar]
- Ferree T, Luu P, Russel JS, Tucker DM ( 2001): Scalp electrode impedance, infection risk, and EEG data quality. Clin Neurophysiol 112: 536–544. [DOI] [PubMed] [Google Scholar]
- Goel V ( 2003): Evidence for dual neural pathways for syllogistic reasoning. Psychologica 32: 301–309. [Google Scholar]
- Goel V ( 2007): Anatomy of deductive reasoning. Trends Cogn Sci 11: 435–441. [DOI] [PubMed] [Google Scholar]
- Goel V, Dolan RJ ( 2001): Functional neuroanatomy of three‐term relational reasoning. Neuropsychologia 39: 901–909. [DOI] [PubMed] [Google Scholar]
- Goel V, Dolan RJ ( 2003): Reciprocal neural response within lateral and ventral medial prefrontal cortex during hot and cold reasoning. Neuroimage 20: 2314–2321. [DOI] [PubMed] [Google Scholar]
- Goel V, Dolan RJ ( 2004): Differential involvement of left prefrontal cortex in inductive and deductive reasoning. Cognition 93: B109–B121. [DOI] [PubMed] [Google Scholar]
- Goel V, Buchel C, Frith C, Dolan RJ ( 2000): Dissociation of mechanisms underlying syllogistic reasoning. Neuroimage 12: 504–514. [DOI] [PubMed] [Google Scholar]
- Goel V, Tierney M, Sheesley L, Bartolo A, Vartanian O, Grafman J ( 2007): Hemispheric specialization in human prefrontal cortex for resolving certain and uncertain inferences. Cereb Cortex 17: 2245–2250. [DOI] [PubMed] [Google Scholar]
- Greene JD, Nystrom LE, Engell AD, Darley JM, Cohen JD ( 2004): The neural bases of cognitive conflict and control in moral judgment. Neuron 44: 389–400. [DOI] [PubMed] [Google Scholar]
- Hyvärinen A ( 1999): Fast and robust fixed point algorithms for independent component analysis. IEEE Trans Neural Netw 10: 126–634. [DOI] [PubMed] [Google Scholar]
- Hyvärinen A, Oja E ( 2000): Independent component analysis: Algorithms and applications. Neural Netw 13: 411–430. [DOI] [PubMed] [Google Scholar]
- Hyvärinen A, Karhunen J, Oja E ( 2001): Independent Component Analysis. New York: Wiley. [Google Scholar]
- Jung TP, Makeig S, Humphries C, Lee TW, McKeown MJ, Iragui V, Sejnowski TJ ( 2000): Removing electroencephalographic artifacts by blind source separation. Psychophysiology 37: 163–178. [PubMed] [Google Scholar]
- Kroger JK, Nystrom LE, Cohen JD, Johnson‐Laird PN ( 2008): Distinct neural substrates for deductive and mathematical processing. Brain Res 1243: 86–103. [DOI] [PubMed] [Google Scholar]
- Lachaux JP, Jung J, Mainy N, Dreher JC, Bertrand O, Baciu M, Minotti L, Hoffmann D, Kahane P ( 2008): Silence is golden: Transient neural deactivation in the prefrontal cortex during attentive reading. Cereb Cortex 18: 443–450. [DOI] [PubMed] [Google Scholar]
- Luo J, Yuan J, Qiu J, Zhang Q, Zhong J, Huai Z ( 2008): Neural correlates of the belief‐bias effect in syllogistic reasoning: An event‐related potential study. Neuroreport 10: 1073–1078. [DOI] [PubMed] [Google Scholar]
- Makeig S, Westerfield M, Jung TP, Covington J, Townsend J, Sejnowski TJ, Courchesne E ( 1999): Functionally independent components of the late positive event‐related potential during visual spatial attention. J Neurosci 19: 2665–2680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makeig S, Westerfield M, Jung TP, Enghoff S, Townsend J, Courchesne E, Sejnowski TJ ( 2002): Dynamic brain sources of visual evoked responses. Science 295: 690–693. [DOI] [PubMed] [Google Scholar]
- Makeig S, Debener S, Onton J, Delorme A ( 2004): Mining event‐related brain dynamics. Trends Cogn Sci 8: 204–210. [DOI] [PubMed] [Google Scholar]
- Newstead SE, Ellis C, Evans JStBT, Dennis I ( 1997): Conditional reasoning with realistic material. Think Reason 3: 49–76. [Google Scholar]
- Nunez PL, Srinivasan R ( 2006): Electric Field of the Brain, 2nd ed New York: Oxford University Press. [Google Scholar]
- Pascual‐Marqui RD ( 2002): Standardized low‐resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find Exp Clin Pharmacol 24D: 5–12. [PubMed] [Google Scholar]
- Porcaro C, Zappasodi F, Barbati G, Salustri C, Pizzella V, Rossini PM, Tecchio F ( 2006): Fetal auditory responses to external sounds and mother's heart beat: Detection improved by independent component analysis. Brain Res 1101: 51–58. [DOI] [PubMed] [Google Scholar]
- Porcaro C, Coppola G, Di Lorenzo G, Zappasodi F, Siracusano A, Pierelli F, Rossini PM, Tecchio F, Seri S ( 2009): Hand somatosensory subcortical and cortical sources assessed by functional source separation: An EEG study. Hum Brain Mapp 30: 660–674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prado J, Noveck IA ( 2007): Overcoming perceptual features in logical reasoning: A parametric functional magnetic resonance imaging study. J Cogn Neurosci 19: 642–657. [DOI] [PubMed] [Google Scholar]
- Prado J, Kaliuzhna M, Cheylus A, Noveck IA ( 2008): Overcoming perceptual features in logical reasoning: An event‐related potentials study. Neuropsychologia 46: 2629–2637. [DOI] [PubMed] [Google Scholar]
- Qiu J, Li H, Huang X, Zhang F, Chen A, Luo Y, Zhang Q, Yuan H ( 2007): The neural basis of conditional reasoning: An event‐related potential study. Neuropsychologia 45: 1533–1539. [DOI] [PubMed] [Google Scholar]
- Rudebeck PH, Walton ME, Smyth AN, Bannerman DM, Rushworth MF ( 2006): Separate neural pathways process different decision costs. Nat Neurosci 9: 1161–1168. [DOI] [PubMed] [Google Scholar]
- Rushworth MF, Behrens TE ( 2008): Choice, uncertainty and value in prefrontal and cingulate cortex. Nat Neurosci 4: 389–397. [DOI] [PubMed] [Google Scholar]
- Salustri C, Barbati G, Porcaro C ( 2005): Fetal magnetocardiographic signals extracted by “signal subspace” blind source separation. IEEE Trans Biomed Eng 52: 1140–1142. [DOI] [PubMed] [Google Scholar]
- Sarvas J ( 1987): Basic mathematical and electromagnetic concepts of the biomagnetic inverse problem. Phys Med Biol 32: 11–22. [DOI] [PubMed] [Google Scholar]
- Tucker DM ( 1993): Spatial sampling of head electrical fields: the geodesic sensor net. Electroencephalogr Clin Neurophysiol 87: 145–163. [DOI] [PubMed] [Google Scholar]
- Vorobyov S, Cichocki A ( 2002): Blind noise reduction for multisensory signals using ICA and subspace filtering, with application to EEG analysis. Biol Cybern 86: 293–303. [DOI] [PubMed] [Google Scholar]
- Walton ME, Bannerman DM, Rushworth MF ( 2002): The role of rat medial frontal cortex in effort‐based decision making. J Neurosci 22: 10996–11003. [DOI] [PMC free article] [PubMed] [Google Scholar]