

Abstract
Cortical activity in the EEG alpha band has proven to be particularly sensitive to creativity‐related demands, but its functional meaning in the context of creative cognition has not been clarified yet. Specifically, increases in alpha activity (i.e., alpha synchronisation) in response to creative thinking can be interpreted in different ways: As a functional correlate of cortical idling, as a sign of internal top‐down activity or, more specifically, as selective inhibition of brain regions. We measured brain activity during creative thinking in two studies employing different neurophysiological measurement methods (EEG and fMRI). In both studies, participants worked on four verbal tasks differentially drawing on creative idea generation. The EEG study revealed that the generation of original ideas was associated with alpha synchronisation in frontal brain regions and with a diffuse and widespread pattern of alpha synchronisation over parietal cortical regions. The fMRI study revealed that task performance was associated with strong activation in frontal regions of the left hemisphere. In addition, we found task‐specific effects in parietotemporal brain areas. The findings suggest that EEG alpha band synchronisation during creative thinking can be interpreted as a sign of active cognitive processes rather than cortical idling. Hum Brain Mapp, 2009. © 2008 Wiley‐Liss, Inc.
Keywords: alpha, creativity, EEG, fMRI, originality
INTRODUCTION
The ability to think creatively plays an important role in almost all areas of our life: It is essential in education, in the arts or in the scientific domain. Likewise, the generation of novel concepts or ideas is advantageous in engineering or in the economic sector. However, notwithstanding its crucial importance in many areas of our life, no conclusive scientific understanding of this mental ability construct has been achieved yet. Creativity is commonly defined as the ability to produce work that is novel (original, unique), useful, and generative [e.g., Sternberg and Lubart, 1996]. According to Guilford [1950], creative people can be characterized by the ability to produce a large quantity of ideas (i.e., ideational fluency), to produce novel output (unique/original ideas) or to think flexibly (i.e., the ability to produce different types of ideas). Stimulated by Guilford's definitions of creative people, psychometric measures of creative thinking such as the Torrance Tests of Creative Thinking [TTCT; Torrance, 1966] have been developed and empirically tested [see Plucker and Renzulli, 1999].
The availability of psychometric measures has stimulated research on creativity in different scientific disciplines. It has, for instance, been studied in the cognitive sciences [Ward, 2007], in pedagogy or in the educational domain [Sawyer, 2006] and most recently also in the field of neurosciences [e.g., Bowden and Jung‐Beeman, 2007; Bowden et al., 2005; Dietrich, 2004; Fink et al., 2007; Kounios et al., 2006]. In applying a variety of tasks and experimental procedures and in using different neurophysiological measurement methods (such as fMRI, PET, NIRS, EEG), neuroscientific studies have yielded evidence of possible brain correlates underlying creative thinking [for an overview see Fink et al., 2007]. For instance, brain activity has been investigated in response to divergent (as opposed to convergent) thinking [Mölle et al., 1999; Razumnikova, 2000], during insightful problem solving or the subjective experience of “AHA!” [Jung‐Beeman et al., 2004], likewise during the performance of classic creativity tasks such as the alternate or unusual uses test [Folley and Park, 2005; Martindale and Hines, 1975] or during match problem solving tasks [Goel and Vartanian, 2005]. In addition to this, neuroscientific research on creativity has also focused on musical creativity [Petsche, 1996], imagery or visual art [Bhattacharya and Petsche, 2005]. Moreover, the ability to think creatively has been investigated in relation to resting EEG brain activity [Jaušovec and Jaušovec, 2000a].
Martindale's [1999] work provides valuable insights into possible brain correlates underlying creative thinking. In his so‐called low arousal theory of creativity he refers to early psychological concepts in this field of research: Kris' [1952] supposition of primary process cognition, Mendelsohn's [1976] hypothesis of defocused attention, and Mednick's [1962] assumption of individual differences in associative hierarchies. Accordingly, creative people are thought to be more capable of shifting between secondary (abstract, analytical) and primary (dreaming, reverie) modes of thinking, or to “regress” to primary process cognition which is necessary in the generation of novel, original ideas. Creative individuals can also be characterized by “flat” (more and broader associations to a given stimulus) instead of “steep” associational hierarchies (just a few, common associations to a given stimulus), and can attend to more things at the same time (i.e., defocused attention) instead of just narrowly attending to a single task or event. According to Martindale [1999], primary process cognition, defocused attention and flat associational hierarchies are more likely to occur if an individual is in a state of low cortical arousal. Empirically, in using EEG alpha activity as an index of cortical arousal, Martindale reports evidence that highly creative (as opposed to low creative) individuals exhibited a comparatively low cortical arousal during the performance of the Alternate Uses Test which is known as a good measure of creativity [Martindale and Hines, 1975]. Similarly, in Martindale and Hasenfus [1978] highly creative individuals showed lower levels of cortical arousal than less creative subjects while thinking of a story (i.e., inspirational phase) but not during an elaboration phase (i.e., writing down the story). Hence, Martindale's [1999] work suggests that the production of novel, original ideas more likely occurs when the individual is in a state of low cortical arousal or high alpha activity, respectively.
The high sensitivity of EEG activity in the alpha frequency range to creativity‐related demands has been corroborated in several studies on creative cognition [e.g., Jaušovec, 2000; Jaušovec and Jaušovec, 2000b; Jung‐ Beeman et al., 2004; Martindale, 1999; Razumnikova, 2000]. We also investigated brain activity patterns during creative thinking [Fink et al., 2006; Fink and Neubauer, 2006, 2008; Grabner et al., 2007] and focused on event‐ or task‐related changes of EEG alpha activity as this measure has proven to be a reliable and valid EEG correlate of cognition [Neubauer et al., 2006; Neuper and Klimesch, 2006]. In the studies of our laboratory, the EEG of the participants was recorded while they were engaged in the performance of different creative idea generation tasks. The employed tasks were adapted from well‐known creativity tests such as the Torrance Tests of Creative Thinking [TTCT; Torrance, 1966], or from well‐established German creativity tests [i.e., verbal creativity test by Schoppe, 1975; imagination subscales of the Berlin Intelligence Test, Jäger et al., 1997]. Participants were requested to think of original causes or consequences to hypothetical or utopian situations. Furthermore, they were required to name original uses of conventional, everyday objects or to complete German suffixes in an original way [see Fink et al., 2007]. Our findings revealed that creative idea generation is generally accompanied by relatively strong increases in EEG alpha activity relative to a prestimulus resting condition [i.e., in the following referred to as synchronisation of alpha activity; cf. Pfurtscheller and Lopes da Silva, 2005]. Interestingly, alpha synchronisation during creative thinking was higher in response to more “free‐associative” tasks such as responding creatively to hypothetical, utopian situations or generating unusual uses of everyday objects as opposed to completing suffixes originally [Fink et al., 2007]. In addition to this, we also found evidence that more original ideas were associated with stronger increases in alpha activity than less original, conventional ideas during self‐rated [Grabner et al., 2007; cf. also Jung‐Beeman et al., 2004] as well as during external‐rated originality of ideas [Fink and Neubauer, 2006, 2008]. Moreover, synchronisation of alpha activity has even shown to increase as a result of a creative thinking training [Fink et al., 2006]. But what do these findings tell us about possible brain correlates of creative thinking?
Task‐ or event‐related alpha synchronisation has traditionally been considered as a cortical idling phenomenon, presumably reflecting a reduced state of active information processing in the underlying neuronal networks [Pfurtscheller et al., 1996]. In applying this viewpoint to our EEG results a possible interpretation of our findings could be that the generation of novel, original ideas is accompanied by a lower arousal or activity level of the brain [cf. Martindale's low arousal theory; Martindale, 1999]. However, recent evidence in the neuroscientific study of cognition suggests that synchronisation of alpha activity does not merely reflect cortical deactivation or cortical idling [e.g., Klimesch et al., 2007; Knyazev, 2007; Ray and Cole, 1985]. Contrary to the usual finding that alpha power desynchronizes when individuals are engaged in the performance of cognitively demanding tasks, Klimesch et al. [1999], for instance, reported a study in which event‐related synchronisation of alpha activity has been observed during the retention interval of a memory task, i.e. when individuals were instructed to temporarily hold information in mind. Similar findings have been reported by Jensen et al. [2002] and Sauseng et al. [2005] who also investigated alpha power changes while participants were engaged in working memory processing or by Cooper et al. [2003] and Rihs et al. [2007], who analyzed alpha activity during the performance of attentional tasks. In all of these studies, alpha activity has been observed to synchronize (increase) in response to task performance which has been interpreted as a functional correlate of inhibition or top‐down control [Klimesch et al., 2007; Sauseng et al., 2005]. In this context, it is important to note that the term “inhibition” is not used in a physiological but rather in a functional, cognitive sense [for a physiological theory on the normal waking EEG, see Miller, 2007]. Specifically, alpha synchronisation may reflect an inhibition of cognitive processes that are not directly relevant for task performance [e.g., retrieval of interfering information during retention or the processing of interfering sensory input in working memory tasks; Klimesch et al., 2007]. In a similar vein, von Stein and Sarnthein [2000] argue that alpha activity reflects the absence of bottom up stimulation and thus “a pure form of top‐down activity” (p. 311). The generation of novel, original ideas certainly requires such a mental state that is not driven or influenced by external bottom up stimulation. This could also explain the particular role of EEG alpha activity in the context of creative thinking.
To learn more about possible brain correlates underlying creative thinking, the research presented in this article was designed to study the functional meaning of alpha synchronisation in the particular context of creative cognition more thoroughly. To this end, brain activity during creative idea generation was measured in two studies employing different neurophysiological measurement methods. In study 1, brain activity was—as we did in our former studies—quantified by means of task‐ or event‐related changes of EEG alpha activity. In study 2, brain activity during creative thinking was studied by means of functional MRI. Experimental design and tasks (timing, stimuli, response modality, etc.) were exactly the same in both studies: Participants worked on different idea generation tasks, viz. the classic unusual uses test (i.e., generation of original uses of conventional everyday objects) and the name invention task (i.e., generating names to given abbreviations). Along with these rather “free‐associative” tasks participants were also confronted with more verbal ability‐related demands by administering an object characteristics task (i.e., name typical attributes of conventional objects) and a task requiring the completion of German suffixes. Guided by recent work in this field of research, we might generally assume that different types of thinking (such as convergent vs. divergent, verbal ability‐related vs. free‐associative thinking) are associated with different activity patterns of the brain [Carlsson et al., 2000; Goel and Vartanian, 2005; Jaušovec, 2000; Martindale and Hines, 1975; Mölle et al., 1999; Razumnikova, 2000]. Particularly, when participants are engaged in the generation of unusual uses or in inventing original names they are expected to show a stronger synchronisation of EEG alpha activity than during the performance of tasks with more verbal ability‐related demands such as the completion of suffixes [cf. Fink et al., 2006, 2007; Grabner et al., 2007]. Depending on whether the topographical synchronisation of EEG alpha activity in study 1 corresponds to an activation or deactivation of certain brain areas in fMRI (study 2), we may then be able to assess whether alpha synchronisation in the context of creative idea generation is rather to be understood as a functional correlate of cortical idling or of active cognitive processes, respectively.
STUDY 1
Method
Participants
Out of a larger pool of participants who were screened with respect to personality, intelligence and trait creativity (see later) 50 participants (25 females and 25 males) took part in the EEG study. Because of the extensive EEG artifacts the data of three persons had to be excluded from further analyses. The remaining sample (n = 47) consisted of 22 females and 25 males. Their age ranged between 18 and 32 years (M = 24.09, SD = 2.95). All participants were healthy, right‐handed and gave written informed consent prior to the EEG recording session. They were paid for their participation in the EEG session.
Psychometric tests
Prior to the EEG sessions participants' intellectual abilities were tested by means of a well‐established German intelligence test, the “Berliner Intelligenz‐Struktur‐Test” [BIS; Jäger et al., 1997] which provides scores for specific intellectual abilities such as verbal, numerical and visuospatial abilities. We administered the Neuroticism Extraversion Openness to new experiences Five Factor Inventory (NEO‐FFI) by Costa and McCrae [translated into German by Borkenau and Ostendorf, 1993] to assess participants' personality traits. During the EEG test session, we also measured temporary mood of the participants by means of a self‐report questionnaire (e.g., activation, anger, calmness, weakness) and anxiety by means of a German version of Spielberger's state‐trait anxiety inventory [STAI; Laux et al., 1981].
Experimental tasks
In both the EEG (study 1) and the fMRI study (study 2) four experimental tasks were given to the participants. The construction or selection of tasks was guided by the main objective to contrast more free‐associative task demands such as those involved in the unusual uses test (which displayed a comparatively strong synchronisation of alpha in our former EEG studies) with more intelligence‐related tasks which require participants to operate with verbal stimulus material [such as completing word ends which exhibited the lowest level of alpha synchronisation; cf. Fink et al., 2006, 2007]. Given that these tasks do not only differ with respect to creativity‐related demands but also with respect to stimulus length (which might in turn complicate interpretations of any neurophysiological task differences) we added two experimental tasks to allow for powerful neuroscientific contrasts. The following four tasks, each comprising eight test items, were presented in separate blocks. In the Alternative Uses (AU) task, participants had to think of unusual/original uses of conventional everyday objects such as a “tin” (example answers: “mirror”, “exhaust for a car”) or an “umbrella” (example answers: “boat for animals”, “epee”). Unlike this, in the Object Characteristics (OC) task participants had to think of typical characteristics of conventional everyday objects such as “shoes” (example answers: “leathery”, “matched”) or “coat hook” (example answers: “wooden”, “hanging”). In the Name Invention (NI) task pairs of letters representing fictional abbreviations (such as “K M”, “T S”) were given to the participants and they had to invent as original names as possible that might belong to the given abbreviation (e.g., K M: “Kissing Manual”, “Kaleidoscope Monster”, T S: “Tissue Spender”, “Time Saver”). And finally, in the Word Ends (WE) task, German suffixes (“‐ung”, “‐nis”) were presented that had to be completed by the participants. Items of all four tasks were presented alternately in a fixed sequence (AU1 ‐ OC1 ‐ NI1 ‐ WE1, AU2 ‐ OC2 ‐ …). The item was displayed in the centre of the screen, a short instruction of the respective task given in the top part of the screen.
Apparatus/EEG recording
The EEG was measured (BrainAmp amplifier) by means of gold electrodes (9 mm diameter) located in an electrode cap in 33 positions (according to the international 10–20 system with interspaced positions); a ground electrode was located on the forehead, the reference electrode was placed on the nose. To register eye movements, an electrooculogram (EOG) was recorded bipolarly between two gold electrodes diagonally placed above and below the inner respectively the outer canthus of the right eye. The EEG signals were filtered between 0.1 and 100 Hz; an additional 50‐Hz notch filter was applied to avoid power line contamination. Electrode impedances were kept below 5 kΩ for the EEG and below 10 kΩ for the EOG. All signals were sampled at a frequency of 500 Hz.
As depicted in Figure 1, each trial started with the presentation of a fixation cross (for a time period of 20 s) which served for the assessment of reference brain activity. Subsequently the test item was presented (again for a time period of 20 s) and the participants started with thinking of possible responses (activation interval). During this time, no overt response was required and participants were instructed not to speak. Afterwards the font of the item changed its colour from white to green which indicated that the participant now had to name his or her ideas (response interval = 8 s). In each trial, an 18‐s time period during the activation interval and an 18‐s time period during the reference interval were used for EEG analyses (the first and the last second of each time period were excluded from EEG analyses). Reference and activation blocks were carefully checked for artifacts by means of visual inspection, and artifactual epochs (because of eye blinks, body movements etc.) were excluded from further analyses.1 For both the reference and the activation intervals the EEG band power (μV2) was calculated within a lower (8–10 Hz) and an upper (10–12 Hz) alpha frequency band. For quantifying task‐related changes (TRP) in EEG alpha power [cf. Pfurtscheller, 1999], the (log‐transformed) power during the reference intervals (averaged over all blocks) was subtracted from the (log‐transformed) power during the activation intervals for each electrode i and trial according to the formula: TRPi = log (Powi activation) − log (Powi reference). Decreases in alpha band power from the reference to the activation interval are reflected in negative TRP values (task‐related desynchronisation), whereas task‐related increases (synchronisation of EEG activity) are expressed in positive values [cf. Pfurtscheller, 1999].
Figure 1.
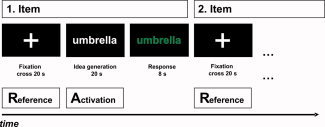
Schematic time course of experimental tasks. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
For statistical analyses, electrode positions were aggregated as following: anteriofrontal (AF) left (FP1, AF3), frontal (F) left (F3, F7), frontocentral (FC) left (FC1, FC5), centrotemporal (CT) left (C3, T3), centroparietal (CP) left (CP1, CP5), parietotemporal (PT) left (P3, T5), parietooccipital (PO) left (PO3, PO5, O1); analogously for the right hemisphere. The midline electrodes (FZ, CZ, PZ) were not included in the analyses as we were also interested in hemispheric differences.
Procedure
At the beginning of the EEG recording session, two 2‐min EEG sequences under resting conditions were recorded, the first with eyes closed, the second with eyes open. Then, after a thorough instruction (demonstration of tasks, type of responding etc.) the participants started to work on the experimental tasks described above. Another two 2‐min resting EEG sequences (with eyes closed and eyes open, respectively) were recorded at the end of the EEG session. In total the EEG recording session took about one hour.
Results
Behavioral results
A one‐way ANOVA with TASK as within‐subjects variable reveals that the OC and the WE task (which presumably involve more verbal ability‐related demands) were associated with a larger number of responses than the more “free‐associative” AU and NI task (34.00, 29.94, 20.79, 18.04 for the OC, WE, AU and NI task, respectively; F [2.65, 121.83] = 157.04, P < 0.01, η2 = 0.77). Performance (i.e., number of ideas) in all tasks was significantly correlated with verbal ability (as assessed by means of the verbal intelligence test scale of the BIS). Interestingly the correlations were somewhat higher for the OC (r = 0.43) and the WE (r = 0.46) than for the AU (r = 0.40) and the NI task (r = 0.32). Moreover, the AU and the NI task were significantly (P < 0.05) correlated with performance in the Unusual Uses scale of the BIS‐test (r = 0.31 and r = 0.41 for the AU and NI task, respectively) while the OC and WE task were not (r = 0.28 and r = 0.14 for the OC and WE task, respectively).
EEG results
EEG data were analyzed by means of two ANOVAs for repeated measures (separately for the lower alpha and the upper alpha band) considering the variables TASK (AU, OC, NI, WE), HEMISPHERE (left vs. right), and AREA (from anteriofrontal to parietooccipital) as within‐subjects variables. In case of violations of sphericity assumptions degrees of freedom were corrected by means of the most conservative Greenhouse Geisser procedure. The probability of a Type I error was maintained at 0.05.
Dealing first with the lower alpha band, we observed a significant main effect of AREA, F (1.50, 69.18) = 52.66, P < 0.01, η2 = 0.53, suggesting a stepwise decrease of lower alpha synchronisation from anteriofrontal to centrotemporal cortices while in centroparietal, parietotemporal and parietooccipital brain regions even a small desynchronisation of lower alpha activity was observed. Moreover, the right hemisphere exhibited a stronger synchronisation than the left hemisphere (main effect of HEMISPHERE, F [1, 46] = 6.01, P < 0.05, η2 = 0.12), which was—as an interaction between TASK and HEMISPHERE suggests, F (2.19, 100.69) = 8.93, P < 0.01, η2 = 0.16—most prominent during performance of the AU task. Performance of the experimental tasks was associated with different patterns of lower alpha (de‐)synchronisation as was also evident by a significant main effect of TASK, F (1.94, 89.20) = 29.09, P < 0.01, η2 = 0.39, a TASK by AREA interaction, F (3.86, 177.78) = 21.73, P < 0.01, η2 = 0.32, and an interaction between TASK, HEMISPHERE and AREA, F (7.29, 335.33) = 3.02, P < 0.01, η2 = 0.06. We found comparably strong right‐hemispheric lower alpha synchronisation during performance of the AU and OC task (which was somewhat more pronounced in the AU than in the OC task) and desynchronisation of lower alpha activity during performance of the NI and WE task (which was more pronounced in the NI than the WE task), particularly apparent in posterior regions of the brain (centroparietal to parietooccipital; cf. Fig. 2).
Figure 2.
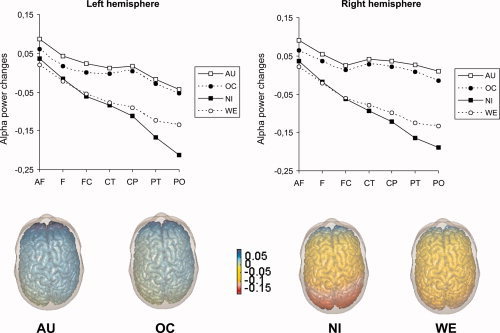
Task‐related changes in EEG alpha activity (upper alpha band, 10–12 Hz) during performance of experimental tasks. Blue regions indicate increases in alpha activity relative to rest, red regions decreases. AU: Alternative uses; OC: Object characteristics; NI: Name invention; WE: Word ends; AF: anteriofrontal; F: frontal; FC: frontocentral; CT: centrotemporal; CP: centroparietal; PT: parietotemporal; PO: parietooccipital. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Similar effects emerged in the upper alpha band. The repeated measurements ANOVA revealed a significant main effect of AREA (F [1.68, 77.21] = 53.14, P < 0.01, η2 = 0.54) indicating upper alpha synchronisation in anteriofrontal and frontal brain regions and a desynchronisation of upper alpha activity in the remaining cortices (monotone increase in desynchronisation from frontocentral to parietooccipital cortices). An interaction between AREA and HEMISPHERE (F [3.05, 140.16] = 2.78, P < 0.05, η2 = 0.06) yielded evidence that this effect was more pronounced in the left than in the right hemisphere. The main effect TASK (F [1.99, 91.64] = 45.50, P < 0.01, η2 = 0.50), as well as the interaction between TASK and AREA (F [3.47, 159.79] = 21.43, P < 0.01, η2 = 0.32), reached statistical significance, suggesting upper alpha synchronisation during performance of the AU and OC task and desynchronisation of upper alpha activity during performance of the NI and WE task, particularly apparent in posterior regions of the brain. The left hemisphere displayed a stronger upper alpha desynchronisation than the right hemisphere, (F [1, 46] = 6.11, P < 0.05, η2 = 0.12), but, as it was evident by an interaction between TASK and HEMISPHERE (F [2.51, 115.65] = 7.14, P < 0.01, η2 = 0.13) and an interaction between TASK, HEMISPHERE and AREA (F [6.44, 296.23] = 3.08, P < 0.01, η2 = 0.06), performance of the AU and OC task was associated with a comparatively strong synchronisation in the posterior cortices of the right hemisphere while in the NI and WE task only alpha desynchronisation was observed, comparably in size for both hemispheres (cf. Fig. 2).
Brain activity during the generation of unusual uses in lower vs. higher original participants
Based on the originality of ideas given during the performance of the AU task participants were divided into a lower original (n = 25, 14 females, 11 males) and a higher original group (n = 22, 8 females, 14 males). Originality of ideas was determined by three female and two male raters who were instructed to evaluate the given responses on a five‐point rating scale ranging from “1” (highly original) to “5” (not original at all). A repeated measures ANOVA was performed on the task‐related power changes in the upper alpha band including the variables HEMISPHERE and AREA as within‐subjects variables and ORIGINALITY group as between‐subjects variable. The ANOVA yielded a significant main effect of AREA, F (1.92, 86.59) = 7.78, P < 0.01, η2 = 0.15, with the largest amount of upper alpha synchronisation in anteriofrontal and the lowest synchronisation in parietooccipital brain regions. The right hemisphere exhibited more synchronisation of alpha activity than the left hemisphere (main effect of HEMISPHERE, F [1, 45] = 7.90, P < 0.01, η2 = 0.15), most prominent in posterior (from centroparietal to parietooccipital) cortices (interaction between HEMISPHERE and AREA, F [3.62, 162.82] = 6.58, P < 0.01, η2 = 0.13). Also, the ANOVA yielded effects related to the originality of ideas: A significant interaction between HEMISPHERE and ORIGINALITY group, F (1, 45) = 4.77, P < 0.05, η2 = 0.10, which was further moderated by topographical AREA, F (3.62, 162.82) = 4.72, P < 0.01, η2 = 0.09. The pattern of this interaction (see Fig. 3) suggests that those participants who produced more original ideas showed pronounced hemispheric differences in posterior regions of the brain (centroparietal to parietooccipital), with more upper alpha synchronisation in the right than in the left hemisphere, while in the lower originality group no hemispheric differences were observed.
Figure 3.
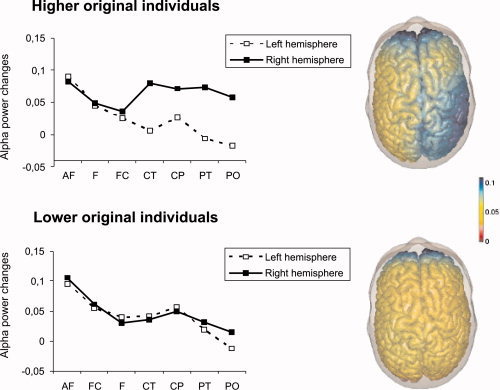
Task‐related changes in EEG alpha activity (upper alpha band, 10–12 Hz) during the generation of unusual uses in the AU task. Blue regions indicate increases in alpha activity relative to rest, red regions decreases. AF: anteriofrontal; F: frontal; FC: frontocentral; CT: centrotemporal; CP: centroparietal; PT: parietotemporal; PO: parietooccipital. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Interim discussion
Analyses of performance data reveal that the AU and the NI task were actually more creativity‐related than the OC and the WE task which were in turn somewhat more strongly correlated with verbal intelligence. Similar to our former EEG studies, the generation of unusual uses was associated with a comparatively strong synchronisation of alpha activity (both in the lower and in the upper alpha band), while in the WE task the lowest level of synchronisation or even desynchronisation was observed [cf. Fink et al., 2007). Analyses point to the particular role of frontal cortices during performance of the presented experimental tasks. In all tasks and particularly in the AU task synchronisation of alpha activity was highest in frontal regions of the brain (cf. Fig. 2). A particular role of posterior parietal brain regions in creative thinking (i.e., generating unusual uses) emerged when individual differences in task performance were taken into account: Those participants who displayed high originality during the generation of unusual uses exhibited a comparatively strong hemispheric asymmetry of upper alpha synchronisation (with more synchronisation in the right than in the left hemisphere) while in those individuals who produced less original responses no hemispheric differences were found.
Taken together, the findings of study 1 resemble the findings of EEG alpha synchronisation in response to creative thinking that were observed in previous studies of our laboratory [cf. Fink et al., 2006, 2007; Grabner et al., 2007]. In study 2 brain activity during performance of these tasks is investigated by means of fMRI.
STUDY 2
Method
Participants
For the fMRI study, another sample of 21 students (10 males) was selected from the pool of participants. This sample was matched with the EEG sample with respect to age (ranging from 20 to 32 years; M = 24.29, SD = 4.09) as well as verbal intelligence and creativity. All participants were healthy, right‐handed, gave written informed consent, and were paid for their participation in the fMRI test session.
Experimental tasks
The same experimental tasks which were used in the EEG study were administered in the fMRI test session, i.e., Alternative Uses (AU), Object Characteristics (OC), Name Invention (NI), and Word Ends (WE). Also task timing and instruction exactly corresponded to the EEG study (see Fig. 1). However, although participants were similarly instructed to articulate their ideas in the response phase following idea generation, their responses could not be recorded and analyzed due to the background noise of the gradient coils.
Apparatus/MR acquisition
Imaging was performed on a 3.0 T Tim Trio system (Siemens Medical Systems, Erlangen, Germany) using an eight‐channel head coil. To minimize head movement, subjects' heads were stabilized with foam cushions. Functional images were obtained with a single shot gradient echo EPI sequence sensitive to blood oxygen level‐dependent (BOLD) contrast (TR = 2,800 ms, TE = 30 ms, FA = 90°, matrix size = 64 × 64, pixel size = 3 × 3 mm). Thirty‐six 3.0‐mm‐thick transverse slices with a distance factor of 25% were acquired parallel to the AC‐PC line in descending acquisition order. In each session, 524 functional volumes were obtained. The first two volumes were discarded to account for T1 saturation effects. In addition to the functional volumes, structural images were obtained using a T1‐weighted 3D MPRAGE sequence (TR = 1,900 ms, TE = 2.2 ms) which provided 1 × 1 × 1 mm isotropic resolution. Stimulus presentation was accomplished with the Eloquence system (Invivo Corporation, Orlando, FL), containing an LCD display with full XGA solution, visible for the participant through a mirror mounted above the head coil. The paradigm was presented with the software package Presentation (Neurobehavioral Systems, Albany, CA).
Analysis of MRI data
Structural and functional imaging data analysis was performed using SPM5 software (Wellcome Department of Imaging Neuroscience, London, U.K.). The functional data of each participant were motion‐corrected, coregistered with the structural data, and then spatially normalized into the standard MNI space (Montreal Neurological Institute). Subsequently, the data were smoothed in the spatial domain using a Gaussian kernel of 8 mm FWHM. All statistical analyses were conducted by means of the general linear model. Model time courses for each experimental condition (i.e., idea generation without the response period) were generated on the basis of the hemodynamic response function implemented in SPM5. A high‐pass filter with a cut‐off frequency of 1/256 Hz was employed to remove low frequency drifts.
The analysis for the entire group was performed by computing linear t‐contrasts (experimental conditions vs. fixation period and between experimental conditions) for each subject individually which were then entered into random effects one‐sample t‐tests. Specifically, each experimental condition (AU, OC, NI, WE) was contrasted with the fixation period to assess activation patterns elicited by the task demands. In addition, t‐contrasts between the experimental tasks sharing the same stimulus material (words in AU and OC; letters in NI and WE) were calculated for revealing demand‐related activation differences. All task‐related effects are reported at P < 0.05 corrected for multiple comparisons by means of the conservative FWE (family wise error) procedure implemented in SPM5. Only activation clusters exceeding a spatial extent threshold of 80 voxels (2 × 2 × 2 mm) are reported.
Procedure
MR imaging started with the acquisition of the structural scans, followed by the experimental paradigm (functional scans). Before MR imaging was performed, the tasks were demonstrated and practiced outside the scanner. The total time of the fMRI test session was about 45 min.
Results
Contrasts against fixation
The results of the contrasts between experimental tasks and fixation are presented in Table I and Figure 4a. As depicted in Figure 4a, all four experimental tasks elicited similar brain activity comprising frontal, parietal, temporal, and occipital cortices as well as the cerebellum. Except for the largely bilateral occipital and cerebellar activation, most of the remaining brain areas displayed a predominantly left‐hemispheric activation pattern. The strongest and most widespread activation cluster in all tasks was found in the left frontal lobe where the activation extends from the inferior frontal gyrus dorsally to the supplementary motor area (SMA) and the precentral gyrus, as well as medially to the anterior cingulate. In addition to this, all tasks elicited significant activation clusters in the left inferior and superior parietal gyrus as well as in the inferior temporal gyrus. Additional activation clusters were found in the hippocampus (bilaterally in the AU und OC task), the left thalamus (in the AU and WE task), and the right middle frontal gyrus (in the NI task).
Table I.
Overview of significant activation clusters (voxelwise P < 0.05 corrected) for the contrasts of the experimental tasks against fixation and between the experimental tasks
| Brain area | MNI peak coordinate | k | t | P corr |
|---|---|---|---|---|
| AU > Fixation | ||||
| L ant cingulate, L inf frontal G, L precentral G | −20 32 22 | 11415 | 15.54 | <0.001 |
| R cerebellum | 36 −66 −26 | 2786 | 13.38 | <0.001 |
| R lingual G, R inf occipital G | 22 −94 −8 | 315 | 11.88 | <0.001 |
| L inf temporal G | −50 −48 −18 | 863 | 11.75 | <0.001 |
| R hippocampus | 36 −40 0 | 303 | 11.68 | <0.001 |
| L lingual G, L calcarine G, L cerebellum | −20 −94 −12 | 860 | 11.57 | <0.001 |
| L sup parietal G | −24 −68 46 | 152 | 11.26 | <0.001 |
| L inf parietal G | −42 −38 44 | 434 | 9.37 | 0.001 |
| R calcarine G | 28 −70 4 | 115 | 9.08 | 0.001 |
| L thalamus, L hippocampus | −10 −22 20 | 182 | 8.86 | 0.002 |
| OC > Fixation | ||||
| L SMA, L inf frontal G | −4 16 46 | 9217 | 16.39 | <0.001 |
| R inf occipital G, R cerebellum | 24 −94 −8 | 2293 | 15.01 | <0.001 |
| L lingual G, L cerebellum, L inf temporal G | −16 −94 −10 | 1879 | 14.48 | <0.001 |
| L inf parietal G | −26 −66 44 | 176 | 11.40 | <0.001 |
| R hippocampus | 34 −36 2 | 195 | 9.01 | 0.001 |
| L hippocampus | −34 −30 −4 | 152 | 8.65 | 0.002 |
| NI > Fixation | ||||
| L inf frontal G, R caudate, L SMA | −46 16 24 | 16588 | 20.99 | <0.001 |
| R cerebellum | 34 −64 −26 | 2545 | 15.31 | <0.001 |
| R inf occipital G | 26 −92 −8 | 349 | 12.78 | <0.001 |
| L mid occipital G, L inf temporal G, L lingual G | −38 −88 −6 | 2229 | 12.58 | <0.001 |
| R mid occipital G | 26 −54 34 | 221 | 10.56 | <0.001 |
| L inf parietal G, L sup parietal G | −26 −66 46 | 336 | 10.33 | <0.001 |
| L inf parietal G | −44 −40 44 | 177 | 9.25 | 0.001 |
| R mid frontal G | 32 44 30 | 170 | 8.33 | 0.004 |
| WE > Fixation | ||||
| L SMA, L mid cingulate, L ant cingulate | −6 16 46 | 3430 | 16.78 | <0.001 |
| L inf frontal G | −52 14 12 | 5024 | 13.81 | <0.001 |
| R postcentral G | 50 −8 34 | 248 | 10.52 | <0.001 |
| R cerebellum | 40 −66 −26 | 1022 | 10.47 | <0.001 |
| L sup parietal G | −28 −64 46 | 230 | 10.46 | <0.001 |
| L inf occipital G, L lingual G | −42 −88 −8 | 1064 | 10.30 | <0.001 |
| L hippocampus, L thalamus | −28 −34 8 | 121 | 8.89 | 0.001 |
| R inf occipital G | 40 −88 −12 | 193 | 8.18 | 0.004 |
| L inf parietal G | −46 −40 44 | 128 | 8.13 | 0.005 |
| R sup occipital G | 28 −56 36 | 112 | 8.04 | 0.005 |
| L precentral G | −36 −4 60 | 90 | 7.94 | 0.006 |
| AU > OC | ||||
| L angular G | −48 −74 30 | 108 | 9.18 | 0.001 |
| OC > AU | ||||
| R angular G, R supramarginal G | 36 −58 48 | 544 | 12.56 | <0.001 |
| NI > WE | ||||
| R inf occipital G | 50 −74 −4 | 118 | 10.35 | <0.001 |
| WE > NI | ||||
| — | — | — | — | n.s. |
Note. Coordinates are reported in MNI space as given by SPM5 and correspond only approximately to Talairach and Tournoux space [Talairach and Tournoux, 1988]. Anatomical labels are based on the AAL (automated anatomical labeling) atlas [Tzourio‐Mazoyer et al., 2002]. The first label represents the location of the peak activation, additional labels denote submaxima if located in a different brain region. Abbreviations: L = left hemisphere, R = right hemisphere, G = Gyrus, inf = inferior, sup = superior, mid = middle.
Figure 4.
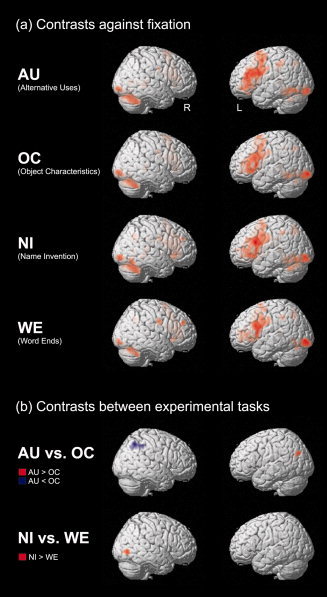
Overview of significant activation clusters in the fMRI t‐contrasts (a) of the experimental tasks against fixation and (b) between the experimental tasks. Activation clusters are depicted on the standard single‐subject volume‐rendered brain implemented in SPM5 (sagittal views; all effects at voxelwise P < 0.05 corrected for multiple comparisons). [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
Comparisons between experimental tasks
In contrasting the AU and OC task it was observed that the left angular gyrus was more strongly activated in the AU task whereas the right angular gyrus displayed higher activity in the OC task (see Table I and Fig. 4b). The latter activation cluster is also larger and additionally covers parts of the supramarginal gyrus.
The contrasts between the NI and WE task only revealed a significantly higher activation in the right inferior occipital gyrus extending into the inferior temporal lobe in the NI task (see Fig. 4b).
Interim discussion
The fMRI analysis revealed that a widespread and very similar neural network is involved in all four experimental tasks. This network is predominantly left‐hemispheric and comprises activation clusters in all four lobes. The largest (and also strongest) activation was observed in the left inferior frontal gyrus extending to the SMA and anterior cingulate. The left inferior frontal gyrus is a key region of language processing and has been reported to be engaged in a wide range of cognitive tasks demanding verbal information processing [for a review, cf. Gernsbacher and Kaschak, 2003]. Beyond its involvement in general phonological and semantic processes, the inferior frontal gyrus has recently been emphasized to play an important role in semantic selection during creativity‐related demands [Jung‐Beeman, 2005]. In particular, it is assumed that it supports processes such as sorting out and inhibiting competing activated concepts for action or for consciousness. Semantic selection does not only appear to loom large in creative idea generation per se, but may have been particularly involved in the administered experimental tasks. Due to the short response period of 8 s, participants had to memorize and select the most original ideas for response that came into their mind during the idea generation period. Directly related to this is the finding of anterior cingulate and SMA activation in the left hemisphere. Both regions are involved in verbal working memory (in particular in rehearsal processes) as well as selective attention processes such as inhibiting the processing of interfering information in working memory [cf. Baddeley, 2003; Smith and Jonides, 1999].
Further left‐hemispheric activation in all experimental tasks was observed in areas of the inferior and superior parietal lobe. The inferior part has also been associated with verbal working memory processes, in particular with the phonological store [Baddeley, 2003]. While inferior frontal regions are supposed to subserve the maintenance of verbal information in working memory through vocal or subvocal rehearsal, the inferior parietal gyrus is regarded as the store where the verbal material is (temporarily) phonologically represented [Baddeley and Hitch, 1974]. The superior part of the parietal lobe, in contrast, is typically associated with attention processes [Posner and Dehaene, 1994].
Another cortical area that has been found to be significantly activated in all tasks and that is linked with verbal information processing is the left posterior inferior temporal region. In contrast to the mostly bilateral occipital brain areas engaged in all tasks which may reflect the more intense visual information processing of the task material as compared with the fixation cross, this brain region may have been particularly engaged during linguistic stimuli processing, both at letter and word level [cf. Gernsbacher and Kaschak, 2003].
Although the general activation patterns elicited by the four experimental tasks are highly similar and share several language‐related brain regions, significant task‐related differences emerged. In contrasting the AU and OC task, in which single words (labelling objects) were presented with a different instruction, it was found that the left angular gyrus is more active in the AU task while the right angular gyrus is more strongly engaged in the OC task. In addition, the activation cluster in the contrast OC > AU was considerably larger and also comprised parts of the right supramarginal gyrus. Since the AU task represents a traditional creativity task requiring divergent, free‐associative thinking, in contrast to the OC task which was designed as a rather convergent control task, the present results suggest that creative thinking is accompanied by stronger activation in the left but weaker activation in the right angular gyrus. On the one hand, this finding is in line with previous studies demonstrating that left temporoparietal regions are specifically involved in verbal creativity tasks [Pavlova and Romanenko, 1988; Ref. given in Bechtereva et al., 2004]. Bechtereva et al. [2004], for instance, required participants to create stories from lists of different words while their rCBF was measured by means of PET. They found stronger activation in left temporoparietal brain regions including the angular gyrus and supramarginal gyrus for word lists requiring more (as compared with less) creative thinking and concluded that these brain regions play “a crucial role in solving verbal creative tasks” [p. 19; for similar findings, see Petsche, 1996 and Petsche et al., 1997]. On the other hand, the finding of lower right angular and supramarginal activity in the AU task stands in contrast to studies emphasizing the role of right‐hemispheric cortices in creative information processing [e.g., Jung‐Beeman et al., 2004; Razoumnikova, 2000]. Besides the general assumption that the right hemisphere works in a more parallel, holistic, free‐associative processing mode than the rather logic‐analytical left hemisphere [e.g., Martindale et al., 1984], Jung‐Beeman [2005] has recently proposed a hemispheric‐related specialisation of temporoparietal brain areas with regard to semantic information processing (Bilateral‐Activation‐Integration‐Selection theory). He argues that angular and supramarginal gyri are specifically involved in semantic activation during verbal tasks. This activated semantic field is assumed to be strongly focused (on the dominant or relevant meaning of a word) in the left but relatively diffuse (including distant and contextually irrelevant meanings) in the right hemisphere. Such a diffuse or coarse semantic coding could be particularly useful in the context of creative idea generation tasks where distant and unusual associations between different semantic concepts are required, similar to the concept of flat associative hierarchies put forward by Mednick [1962]. However, to our knowledge there is no direct evidence that it is especially the right‐hemispheric temporo‐parietal regions that need to be strongly activated during creative idea generation. As outlined above, rather the contrary seems to hold true. Also, in the recent fMRI study by Howard‐Jones et al. [2005] no evidence of increased right‐hemispheric temporo‐parietal activity during creative thinking was observed. They instructed participants to generate either creative or uncreative stories and found that creative (as compared to uncreative) story generation was associated with stronger bilateral frontal and right middle occipital activation but lower activity in the right inferior parietal lobe near the angular gyrus.
The comparison between the NI and WE task revealed a significant activation difference in the right‐hemisphere: Specifically, the NI task elicited a stronger activation in the right inferior occipital gyrus extending to the inferior temporal gyrus than the WE task. These brain regions are part of the ventral stream of the visual cortex and are discussed to subserve object processing [Ungerleider and Haxby, 1994], mental imagery [Mellet et al., 1998], and visual working memory [Ungerleider et al., 1998]. In the context of creative thinking, occipital activation (as measured by means of PET or fMRI) has only been reported by Howard‐Jones et al. [2005] who discussed this result in terms of mental imagery processes. The present finding might tentatively be interpreted in a similar vein. It appears well plausible that inventing original names to fictional abbreviations might demand mental imagery processes to a greater extent than to complete German suffixes (since the WE task requires merely the retrieval of well‐known verbal material, not involving any kind of creative recombination of objects or concepts). Alternatively, the NI task might have also placed stronger demands to visual working memory than the WE task.
Taken together, the fMRI study has revealed a widespread and highly similar network of brain areas that is engaged in all four employed experimental tasks. Significant differences between more creativity‐related and more intelligence‐related verbal tasks were observed in parietotemporal areas for the AU vs. OC comparison and in occipitotemporal areas in the NI vs. WE comparison. Previous investigations and the present fMRI study suggest that parietotemporal cortices including angular and supramarginal gyri are critically involved in creative thinking. Occipitotemporal brain areas might additionally support creative idea generation by mental imagery or visual working memory. How these fMRI results can be related to the findings from Study I and to previous EEG studies is addressed in the following.
GENERAL DISCUSSION
The research presented in this article was designed to investigate possible brain correlates underlying the generation of novel, original ideas. We measured brain activity by means of EEG and fMRI during the generation of unusual uses which is commonly known as a good measure of creativity. Along with this classic creativity task brain activity was also measured in response to a name invention task and during two control tasks (name object characteristics and completing suffixes, respectively) which were constructed to allow for powerful neurophysiological contrasts between experimental tasks.
Similar to our previous EEG studies on creative cognition [cf. Fink et al., 2007], study 1 reveals that thinking about unusual uses of common, everyday objects is accompanied by a comparatively strong synchronisation of EEG alpha activity (both in the lower and in the upper alpha band), particularly in frontal regions of the brain. Unlike this, alpha synchronisation was lower when typical attributes of everyday objects had to be named. In the name invention and in the word ends task, where the participants were required to operate with letters, they showed a somewhat weaker alpha synchronisation in frontal cortices, and even a desynchronisation in posterior parietal regions of the brain. With respect to fMRI all tasks exhibited similar widespread activation (relative to rest) with the largest clusters in frontal regions of the brain (cf. Fig. 4).
As outlined in the introduction, frontal alpha synchronisation during creative cognition can be interpreted in at least two different ways. In viewing EEG alpha synchronisation as a functional correlate of a reduced state of active information processing or cortical idling in the underlying cortical networks [Pfurtscheller et al., 1996], the observed alpha synchronisation in frontal brain regions would conform to the interpretation that the executive frontal brain temporarily needs to disengage when novel or original information processing is required. In a metaphorical sense, the comparatively strong level of frontal alpha synchronisation in the unusual uses tests could then suggest that the participants temporarily “switch‐off” or reduce control functions of the frontal lobe (e.g., some kind of reduced “censorship”) to allow unique or original ideas to enter into conscious awareness. However, this interpretation is complicated by the fact that performance of the remaining tasks exhibited, though to a lesser extent, frontal alpha synchronisation as well. Thus, interpreting the observed frontal alpha synchronisation during the generation of unusual uses simply in terms of frontal lobe deactivation seems difficult to maintain. Also, this interpretation would be difficult to reconcile with the fMRI findings observed in study 2. As evident in Figure 4, we observed relatively clear‐cut evidence that task performance was associated with activation particularly in frontal regions of the brain. This finding is in agreement with recent PET, NIRS and fMRI studies which report converging evidence that creative cognition is associated with frontal brain activation, as opposed to the performance of tasks with lower creativity demands [e.g., Carlsson et al., 2000; Folley and Park, 2005; Goel and Vartanian, 2005]. Hence, frontal alpha synchronisation during creative thinking more likely reflects an active cognitive process rather than mere deactivation of the frontal cortex.
Von Stein and Sarnthein [2000] proposed that alpha activity reflects the absence of stimulus‐driven, external bottom up stimulation and, thus, a form of top‐down activity which “is maximal in situations where cortical processes … are driven by free floating associations, mental imagery, planning etc.” (p. 311). More specifically, on the basis of an integrated overview of relevant research literature, Klimesch et al. [2007] proposed that synchronisation of alpha activity can be interpreted as a functional correlate of inhibition or top‐down control which is especially relevant during internal processing demands. Accordingly, alpha synchronisation is expected to occur over cortical sites that are under or exert top‐down control. For instance, Sauseng et al. [2005] observed alpha power increases in prefrontal brain regions during performance of a working memory task which requires active maintenance and manipulation of information. They interpreted this finding as reflecting selective top‐down inhibition in such a way that frontal alpha synchronisation could protect information processing in frontal brain regions against interfering cognitive processes. In that sense frontal brain regions remain “immune” against concurrent cognitive processes as long as on‐going information processing (i.e., working memory processing) takes places. Consistent with this idea of selective top‐down inhibition, the observed frontal alpha synchronisation could suggest that during the generation of novel, original ideas [which presumably also requires top‐down processing; cf. Dietrich, 2004] frontal brain regions must not be distracted by interfering cognitive processes during on‐going idea generation [cf. Klimesch et al., 2007; Sauseng et al., 2005].
The idea of active inhibition of brain areas (rather than cortical idling) would be also in agreement with the fMRI findings observed in study 2. In all tasks, brain activation (relative to rest) was strongest in the left inferior frontal gyrus. This brain region is believed to be involved in the selection of semantic representations and in inhibiting competing activated concepts for action or for consciousness [Jung‐Beeman, 2005]. The generation of novel, original ideas certainly necessitates some kind of semantic selection [cf. Jung‐Beeman, 2005] which is presumably mediated by the inferior frontal gyrus. Quite similarly, Heilman et al. [2003] argue that the frontal lobe could support creativity by selectively activating remote conceptual or semantic networks of the brain and by inhibiting brain circuits that store similar semantic information. Of course, frontal alpha synchronisation was observed in all experimental tasks and, therefore, the same interpretation would apply to the other tasks as well. However, in this particular regard it is noteworthy that when the significance threshold in the fMRI contrasts between the experimental tasks was lowered (P < 0.00001 uncorrected), a stronger left‐hemispheric frontal activation in the AU as compared with the OC task emerged (peak coordinates: x = −14, y = 66, z = 2; left superior medial frontal gyrus). There was no additional frontal activation cluster in the reverse contrast.
Along with synchronisation of alpha in frontal brain areas, the generation of unusual uses was also associated with a diffuse and widespread synchronisation over posterior regions of the brain. Specifically, the strongest amount of posterior parietal alpha synchronisation was observed in the AU task, followed by the OC task, and finally by the WE and NI task in which even alpha desynchronisation was found. Particularly the NI task exhibited comparatively strong alpha desynchronisation in parietotemporal and parietooccipital brain regions which could reflect the heightened demands on visual attention or visual working memory [Ungerleider et al., 1998] during performance of the fictional abbreviations given in this task [cf. also Howard‐Jones et al., 2005]. In line with this, in study 2 the NI task elicited stronger hemodynamic responses in the right inferior occipital gyrus than the WE task (cf. Fig. 4).
Synchronisation of posterior parietal alpha activity was also reported to emerge in more original as opposed to less original, conventional ideas [Fink and Neubauer, 2006; Grabner et al., 2007]. Similarly, in the present EEG study higher original individuals exhibited a hemispheric asymmetry in alpha synchronisation with more (upper) alpha activity in the right than in the left hemisphere. In contrast, those individuals who produced less original ideas showed no hemispheric differences with respect to alpha band power. Enhanced alpha activity was also found during insightful problem solving, as recently reported by Jung‐Beeman et al. [2004]. Specifically, the authors had their participants work on remote associate problems (finding a compound to three given stimulus words) and compared brain activity during solutions that were accompanied by subjective experience of “AHA!” (as determined by self‐report) with solutions that were solved without insight. Interestingly, those solutions that were solved with insight were associated with more alpha band power in the right posterior parietal cortex than those that were solved without insight. The authors presume that enhanced EEG alpha activity during subjective experience of insight could “attenuate bottom‐up activation or other neural activity not related to solution” (p. 507), thereby allowing “…processing about more distant associations between the problem words” (ibid.).
The fMRI study revealed that the AU task was associated with stronger brain activation than the OC task in the angular gyrus of the left hemisphere but a lower activation of the respective region in the right hemisphere extending to parts of the supramarginal gyrus. [cf. Bechtereva et al., 2004; Howard‐Jones et al., 2005; Petsche et al., 1997]. This corresponds to a diffuse and widespread pattern of alpha synchronisation in bilateral (but particularly right‐hemispheric) parietal brain regions in the EEG data. In general, the pronounced synchronisation of the posterior parietal cortex during the generation of novel ideas could possibly indicate that the underlying cortical networks are in a state of “internal attention” [cf. Knyazev et al., 2006; see also Knyazev, 2007] or in a state of internal top‐down activity [von Stein and Sarnthein, 2000] that is less likely disturbed by interfering cognitive processes [cf. Sauseng et al., 2005]. The finding of a diffuse and widespread alpha synchronisation during the generation of original ideas is also in accordance with von Stein and Sarnthein's [2000] work which suggests that long range frontoparietal interactions during mental imagery are particularly reflected in the alpha frequency range. Along with a comparatively weak fMRI activation of regions of the right‐hemispheric posterior cortex the diffuse und widespread alpha synchronisation may facilitate the (re‐)combination of semantic information that is normally distantly related [cf. Jung‐Beeman, 2005].
However, the hemispheric fMRI activation differences between the AU and OC task are difficult to reconcile with the EEG data with respect to their interpretation as cortical activation or deactivation. Specifically, a diffuse and widespread pattern of right‐hemispheric alpha synchronisation during performance of the AU task—which was particularly apparent in higher original individuals—goes along with comparatively low fMRI brain activation (relative to the OC task) in the right‐hemispheric angular and supramarginal gyri. This finding would suggest that alpha synchronisation in posterior brain regions would reflect cortical deactivation or idling which, in turn, would be in conflict with our interpretation of alpha band synchronisation as a sign of active cognitive processes. Moreover, this finding appears to stand in contrast with findings of simultaneous EEG‐fMRI studies which suggest a negative relationship between alpha band power and brain activation as measured by the hemodynamic BOLD response [e.g., Feige et al., 2005; Goldman et al., 2002; Laufs et al., 2003; Moosmann et al., 2003]. However, the majority of these studies investigated spontaneous alpha activity during rest with eyes closed and found negative correlations primarily over occipital brain areas. Therefore, it is questionable whether these findings can be generalized to task‐related alpha power changes during higher‐order cognitive demands [see also Klimesch et al., 2007].
Though highly speculative, differences in neural microcircuitry between the right and the left hemisphere could also account for the finding that in parietal brain regions EEG alpha synchronisation goes along with lower fMRI activation. Jung‐Beeman [2005] refers to data suggesting a higher interconnectivity in the right than in the left hemisphere. Specifically, he argues that the right hemisphere has a greater proportion of white matter (or connections between neurons, respectively) than the left hemisphere, a higher correlation of activity across different cortical regions, or more diffuse electrophysiological responses [Jung‐Beeman, 2005, p. 513]. This could—at least partly—also explain our finding of a diffuse and widespread (rather than localized) pattern of EEG alpha synchronisation in the right hemisphere, while fMRI (possibly unaffected by right‐hemispheric interconnectivity due to its high spatial accuracy) revealed highly localized hemodynamic responses. Future studies which perform EEG and fMRI measurements simultaneously are needed to clarify this issue.
The research presented in this article has shown that creative thinking or creative idea generation appears to comprise a variety of “ordinary” cognitive processes. Creative thinking involves, among others, cognitive flexibility or the ability to develop alternative concepts or strategies [Heilman et al., 2003]. Moreover, the generation of novel, original ideas by combining already stored information [cf. Dietrich, 2004] presumably also involves working memory or inhibitory top‐down control [Klimesch et al., 2007; Sauseng et al., 2005]. In this particular context it could be objected that the employed AU task was rather a memory than a creativity task, inducing alpha band synchronisation simply because the task required participants to temporarily maintain information in mind (for later recall), as it is typically the case in classic working memory tasks. However, this would hardly explain why frontal alpha synchronisation was higher in the AU than in the OC task, particularly in view of the fact that in the latter more responses were given [which should result in a larger memory load and, consequently, in a stronger synchronisation of alpha; cf. Jensen et al., 2002]. Also, we observed alpha synchronisation in the posterior parietal cortex in employing another experimental procedure as well. For instance, in Fink and Neubauer [2006] brain activity was measured prior to the production of a single idea. Regardless whether the idea was conventional or original, in both cases a single idea had to be temporarily maintained in mind. However, synchronisation over posterior brain regions was significantly stronger in more original than conventional ideas.
Though the reported findings may uncover some brain correlates underlying creative thinking, some important issues are still unresolved. First, more scrutiny is needed with respect to the interpretation of neurophysiological findings in creative cognition. In the present research we administered exactly the same tasks and experimental design in both EEG and fMRI to explore the functional significance of the well‐established finding of increased alpha synchronisation during creative idea generation. Even though the results from study 1 and study 2 provide a plausible picture of potential brain correlates of creative thinking, some findings do not entirely correspond. This holds true, for instance, for the differential frontal alpha synchronisation of the experimental tasks in the EEG while there was no significant task‐related difference in fMRI. Moreover, right‐hemispheric alpha synchronisation in posterior brain regions goes along with lower fMRI brain activation in the right hemisphere. Such discrepancies may also be due to the fact that different (though matched) samples were investigated separately in EEG and fMRI. As aforementioned, the simultaneous measurement of EEG and fMRI in future studies on creative thinking might resolve some of these open issues.
Second, the employed creative idea generation tasks of our laboratory are comparatively simple or basic types of tasks that had to be modified (or simplified, respectively) to be reasonably applicable in EEG and fMRI measurements. Thus, the employed tasks can only be indicative of basic aspects of creative thinking and performance in these tasks cannot be generalized to “real‐life” creative achievements. The difficulty of operationalizing creative thinking in neuroscientific studies of creative cognition is additionally complicated by the fact that participants (unlike to their natural environment) are required to be creative while they are mounted with an electrode cap sitting in a shielded EEG cabin or lying supine in the fMRI scanner. Thus, future neuroscientific research on creativity is also challenged by the investigation of brain activity in response to more complex, “real‐life” creativity tasks.
Acknowledgements
The valuable contributions of Anna Kanape, Christine Kragl, Nadja Kozel and Klaus Feichtinger to this research project are gratefully acknowledged.
Footnotes
Discarded samples (in %; averaged over trials and participants) in the activation period: AU task: 19.17%; OC task: 19.08%; NE task: 19.28%; WE task: 20.79%. Discarded samples (in %) in the reference period: 17.62%. A discarding rate of for instance 20% means that on overage 20% of the samples of a trial (or 3.60 seconds out of 18 s, respectively) had to be excluded from further analyses.
REFERENCES
- Baddeley A( 2003): Working memory: Looking back and looking forward. Nat Rev Neurosc 4: 829–839. [DOI] [PubMed] [Google Scholar]
- Baddeley AD,Hitch GJ ( 1974): Working memory In: Bower GA, editor. Recent Advances in Learning and Motivation, Vol. 8 New York: Academic Press; pp. 47–90. [Google Scholar]
- Bechtereva NP,Korotkov AD,Pakhomov SV,Roudas MS,Starchenko MG,Medvedev SV ( 2004): PET study of brain maintenance of verbal creative activity. Int J Psychophysiol 53: 11–20. [DOI] [PubMed] [Google Scholar]
- Bhattacharya J,Petsche H ( 2005): Drawing on mind's canvas: Differences in cortical integration patterns between artists and non‐artists. Hum Brain Mapp 26: 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borkenau P,Ostendorf F ( 1993): NEO‐Fünf‐Faktoren Inventar (NEO‐FFI) nach Costa und McCrae. Göttingen: Hogrefe. [Google Scholar]
- Bowden EM,Jung‐Beeman M ( 2007): Methods for investigating the neural components of insight. Methods 42: 87–99. [DOI] [PubMed] [Google Scholar]
- Bowden EM,Jung‐Beeman M,Fleck J,Kounios J ( 2005): New approaches to demystifying insight. Trends Cogn Sci 9: 322–328. [DOI] [PubMed] [Google Scholar]
- Carlsson I,Wendt PE,Risberg J ( 2000): On the neurobiology of creativity. Differences in frontal activity between high and low creative subjects. Neuropsychologia 38: 873–885. [DOI] [PubMed] [Google Scholar]
- Cooper NR,Croft RJ,Dominey SJJ,Burgess AP,Gruzelier JH ( 2003): Paradox lost? Exploring the role of alpha oscillations during externally vs. internally directed attention and the implications for idling and inhibition hypotheses Int J Psychophysiol 47: 65–74. [DOI] [PubMed] [Google Scholar]
- Dietrich A ( 2004): The cognitive neuroscience of creativity. Psychon B Rev 11: 1011–1026. [DOI] [PubMed] [Google Scholar]
- Feige B,Scheffler K,Esposito F,Di Salle F,Hennig J,Seifritz E ( 2005): Cortical and subcortical correlates of electroencephalographic alpha rhythm modulation. J Neurophysiol 93: 2864–2872. [DOI] [PubMed] [Google Scholar]
- Fink A,Neubauer AC ( 2006): EEG alpha oscillations during the performance of verbal creativity tasks: Differential effects of sex and verbal intelligence. Int J Psychophysiol 62: 46–53. [DOI] [PubMed] [Google Scholar]
- Fink A,Neubauer AC ( 2008): Eysenck meets Martindale: The relationship between extraversion and originality from the neuroscientific perspective. Pers Indiv Differ 44: 299–310. [Google Scholar]
- Fink A,Grabner RH,Benedek M,Neubauer AC ( 2006): Divergent thinking training is related to frontal electroencephalogram alpha synchronization. Eur J Neurosci 23: 2241–2246. [DOI] [PubMed] [Google Scholar]
- Fink A,Benedek M,Grabner RH,Staudt B,Neubauer AC ( 2007): Creativity meets neuroscience: Experimental tasks for the neuroscientific study of creative thinking. Methods 42: 68–76. [DOI] [PubMed] [Google Scholar]
- Folley BS,Park S ( 2005): Verbal creativity and schizotypal personality in relation to prefrontal hemispheric laterality: A behavioral and near‐infrared optical imaging study. Schizophr Res 80: 271–282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gernsbacher MA,Kaschak MP ( 2003): Neuroimaging studies of language production and comprehension. Annu Rev Psychol 54: 91–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goel V,Vartanian O ( 2005): Dissociating the roles of right ventral lateral and dorsal lateral prefrontal cortex in generation and maintenance of hypotheses in set‐shift problems. Cereb Cortex 15: 1170–1177. [DOI] [PubMed] [Google Scholar]
- Goldman RI,Stern JM,Engel J,Cohen MS ( 2002): Simultaneous EEG and fMRI of the alpha rhythm. Neuroreport 13: 2487–2492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabner RH,Fink A,Neubauer AC ( 2007): Brain correlates of self‐rated originality of ideas: Evidence from event‐related power and phase‐locking changes in the EEG. Behav Neurosci 121: 224–230. [DOI] [PubMed] [Google Scholar]
- Guilford JP ( 1950): Creativity. Am Psychol 5: 444–454. [DOI] [PubMed] [Google Scholar]
- Heilman KM,Nadeau SE,Beversdorf DO ( 2003): Creative innovation: possible brain mechanisms. Neurocase 9: 369–379. [DOI] [PubMed] [Google Scholar]
- Howard‐Jones PA,Blakemore S‐J,Samuel EA,Summers IR,Claxton G ( 2005): Semantic divergence and creative story generation: An fMRI investigation. Cogn Brain Res 25: 240–250. [DOI] [PubMed] [Google Scholar]
- Jäger AO,Süß H‐M,Beauducel A ( 1997): Berliner Intelligenzstruktur‐Test. Göttingen: Hogrefe. [Google Scholar]
- Jaušovec N ( 2000): Differences in cognitive processes between gifted, intelligent, creative, and average individuals while solving complex problems: An EEG Study. Intelligence 28: 213–237. [Google Scholar]
- Jaušovec N,Jaušovec K ( 2000a): Differences in resting EEG related to ability. Brain Topogr 12: 229–240. [DOI] [PubMed] [Google Scholar]
- Jaušovec N,Jaušovec K ( 2000b): EEG activity during the performance of complex mental problems. Int J Psychophysiol 36: 73–88. [DOI] [PubMed] [Google Scholar]
- Jensen O,Gelfand J,Kounios J,Lisman JE ( 2002): Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short‐term memory task. Cereb Cortex 12: 877–882. [DOI] [PubMed] [Google Scholar]
- Jung‐Beeman M ( 2005): Bilateral brain processes for comprehending natural language. Trends Cogn Sci 9: 512–518. [DOI] [PubMed] [Google Scholar]
- Jung‐Beeman M,Bowden EM,Haberman J,Frymiare JL,Arambel‐Liu S,Greenblatt R,Reber PJ,Kounios J ( 2004): Neural activity when people solve verbal problems with insight. PLOS Biol 2: 500–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimesch W,Doppelmayr M,Schwaiger J,Auinger P,Winkler T ( 1999): “Paradoxical” alpha synchronization in a memory task. Cogn Brain Res 7: 493–501. [DOI] [PubMed] [Google Scholar]
- Klimesch W,Sauseng P,Hanslmayr S ( 2007): EEG alpha oscillations: The inhibition‐timing hypothesis. Brain Res Rev 53: 63–88. [DOI] [PubMed] [Google Scholar]
- Knyazev GG ( 2007): Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci Biobehav Rev 31: 377–395. [DOI] [PubMed] [Google Scholar]
- Knyazev GG,Savostyanov AN,Levin EA ( 2006): Alpha synchronization and anxiety: Implications for inhibition vs. alertness hypotheses. Int J Psychophysiol 59: 151–158. [DOI] [PubMed] [Google Scholar]
- Kounios J,Frymiare JL,Bowden EM,Fleck JI,Subramaniam K,Parrish TB,Jung‐Beeman M ( 2006): The prepared mind. Neural activity prior to problem presentation predicts subsequent solution by sudden insight. Psychol Sci 17: 882–890. [DOI] [PubMed] [Google Scholar]
- Kris E ( 1952): Psychoanalytic Explorations in Art. New York: International Universities Press. [Google Scholar]
- Laufs H,Kleinschmidt A,Beyerle A,Eger E,Salek‐Haddadi A,Preibisch C,Krakow K ( 2003): EEG‐correlated fMRI of human alpha activity. Neuroimage 19: 1463–1476. [DOI] [PubMed] [Google Scholar]
- Laux L,Glanzmann P,Schaffner P,Spielberger CD ( 1981): State‐Trait‐Angstinventar STAI. Weinheim: Beltz. [Google Scholar]
- Martindale C ( 1999): Biological bases of creativity In Sternberg R, editor. Handbook of Creativity. Cambridge: University Press; pp. 137–152. [Google Scholar]
- Martindale C,Hasenfus N ( 1978): EEG differences as a function of creativity, stage of the creative process, and effort to be original. Biol Psychol 6: 157–167. [DOI] [PubMed] [Google Scholar]
- Martindale C,Hines D ( 1975): Creativity and cortical activation during creative, intellectual, and EEG feedback tasks. Biol Psychol 3: 71–80. [DOI] [PubMed] [Google Scholar]
- Martindale C,Hines D,Mitchell L,Covello E ( 1984): EEG alpha asymmetry and creativity. Pers Indiv Differ 5: 77–86. [Google Scholar]
- Mednick SA ( 1962): The associative basis of the creative process. Psychol Rev 69: 220–232. [DOI] [PubMed] [Google Scholar]
- Mellet E,Tzourio N,Denis M,Mazoyer B ( 1998): Cortical anatomy of mental imagery of concrete nouns based on their dictionary definition. Neuroreport 9: 803–809. [DOI] [PubMed] [Google Scholar]
- Mendelsohn GA ( 1976): Associative and attentional processes in creative performance. J Pers 44: 341–369. [Google Scholar]
- Miller R ( 2007): Theory of the normal waking EEG: From single neurons to waveforms in the alpha, beta and gamma frequency ranges. Int J Psychophysiol 64: 18–23. [DOI] [PubMed] [Google Scholar]
- Mölle M,Marshall L,Wolf B,Fehm HL,Born J ( 1999): EEG complexity and performance measures of creative thinking. Psychophysiol 36: 95–104. [DOI] [PubMed] [Google Scholar]
- Moosmann M,Ritter P,Krastel I,Brink A,Thees S,Blankenburg F,Taskin B,Obrig H,Villringer A ( 2003): Correlates of alpha rhythm in functional magnetic resonance imaging and near infrared spectroscopy. Neuroimage 20: 145–158. [DOI] [PubMed] [Google Scholar]
- Neubauer AC,Fink A,Grabner RH ( 2006): Sensitivity of alpha band ERD/ERS to individual differences in cognition In: Neuper C,Klimesch W, editors. Event‐Related Dynamics of Brain Oscillations—Progress in Brain Research, Vol. 159 Amsterdam: Elsevier; pp. 167–178. [DOI] [PubMed] [Google Scholar]
- Neuper C,Klimesch W ( 2006): Event‐related dynamics of brain oscillations. Series: Progress in Brain Research, Vol. 159 Amsterdam: Elsevier. [Google Scholar]
- Petsche H,Kaplan S,von Stein A,Filz O ( 1997): The possible meaning of the upper and lower alpha frequency ranges for cognitive and creative tasks. Int J Psychophysiol 26: 77–97. [DOI] [PubMed] [Google Scholar]
- Petsche H ( 1996): Approaches to verbal, visual and musical creativity by EEG coherence analysis. Int J Psychophysiol 24: 145–159. [DOI] [PubMed] [Google Scholar]
- Pfurtscheller G ( 1999): Quantification of ERD and ERS in the time domain In: Pfurtscheller G,Lopes da Silva FH, editors. Event‐Related Desynchronization. Handbook of Electroencephalography and Clinical Neurophysiology, rev. edition, Vol. 6 Amsterdam: Elsevier; pp. 89–105. [Google Scholar]
- Pfurtscheller G,Lopes da Silva FH ( 2005): EEG event‐related desynchronization (ERD) and event‐related synchronization (ERS) In: Niedermeyer E,Lopes da Silva FH, editors. Electroencephalography: Basic Principles, Clinical Applications, and Related Fields, 5th ed Philadelphia: Lippincott; pp. 1003–1016. [Google Scholar]
- Pfurtscheller G,Stancak A Jr,Neuper C ( 1996): Event‐related synchronization (ERS) in the alpha band—An electrophysiological correlate of cortical idling: A review. Int J Psychophysiol 24: 39–46. [DOI] [PubMed] [Google Scholar]
- Plucker JA,Renzulli JS ( 1999): Psychometric approaches to the study of human creativity In: Sternberg RJ,editor. Handbook of Creativity Cambridge: University Press; pp. 35–61. [Google Scholar]
- Posner MI,Dehaene S ( 1994): Attentional networks. Trends Neurosci 17: 75–79. [DOI] [PubMed] [Google Scholar]
- Ray WJ,Cole HW ( 1985): EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 228: 750–752. [DOI] [PubMed] [Google Scholar]
- Razumnikova OM ( 2000): Functional organization of different brain areas during convergent and divergent thinking: an EEG investigation. Cogn Brain Res 10: 11–18. [DOI] [PubMed] [Google Scholar]
- Rihs TA,Michel CM,Thut G ( 2007): Mechanisms of selective inhibition in visual spatial attention are indexed by α‐band EEG synchronization. Eur J Neurosci 25: 603–610. [DOI] [PubMed] [Google Scholar]
- Sauseng P,Klimesch W,Doppelmayr M,Pecherstorfer T,Freunberger R,Hanslmayr S ( 2005): EEG alpha synchronization and functional coupling during top‐down processing in a working memory task. Hum Brain Mapp 26: 148–155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawyer RK ( 2006): Educating for innovation. Think Skills Creativ 1: 41–48. [Google Scholar]
- Schoppe K ( 1975): Verbaler Kreativitäts‐Test (V‐K‐T). Göttingen: Hogrefe. [Google Scholar]
- Smith EE,Jonides J ( 1999): Storage and executive processes in the frontal lobes. Science 283: 1657–1661. [DOI] [PubMed] [Google Scholar]
- Sternberg RJ,Lubart TI ( 1996): Investing in creativity. Am Psychol, 7: 677–688. [Google Scholar]
- Talairach J,Tournoux P ( 1988): Co‐planar Stereotaxic Atlas of the Human Brain. New York,NY: Thieme Medical. [Google Scholar]
- Torrance EP ( 1966): Torrance Tests of Creative Thinking. Bensenville, IL: Scholastic Testing Service. [Google Scholar]
- Tzourio‐Mazoyer N,Landeau B,Papathanassiou D,Crivello F,Etard O,Delcroix N,Mazoyer B,Joliot M ( 2002): Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single‐subject brain. Neuroimage 15: 273–289. [DOI] [PubMed] [Google Scholar]
- Ungerleider LG,Haxby JV ( 1994): ‘What’ and ‘where’ in the human brain. Curr Opin Neurobiol 4: 157–165. [DOI] [PubMed] [Google Scholar]
- Ungerleider LG,Courtney SM,Haxby JV ( 1998): A neural system for human visual working memory. Proc Natl Acad Sci USA 95: 883–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Von Stein A,Sarnthein J ( 2000): Different frequencies for different scales of cortical integration: From local gamma to long range alpha/theta synchronization. Int J Psychophysiol 38: 301–313. [DOI] [PubMed] [Google Scholar]
- Ward TB ( 2007): Creative cognition as a window on creativity. Methods 42: 28–37. [DOI] [PubMed] [Google Scholar]