

Abstract
This study used functional MRI (fMRI) to examine the neural effects of willfully allocating one's attention to one of two ongoing tasks. In a dual task paradigm, participants were instructed to focus either on auditory sentence comprehension, mental rotation, or both. One of the major findings is that the distribution of brain activation was amenable to strategic control, such that the amount of activation per task was systematically related to the attention‐dividing instructions. The activation in language processing regions was lower when attending to mental rotation than when attending to the sentences, and the activation in visuospatial processing regions was lower when attending to sentences than when attending to mental rotations. Additionally, the activation was found to be underadditive, with the dual‐task condition eliciting less activation than the sum of the attend sentence and attend rotation conditions. We also observed a laterality shift across conditions within language‐processing regions, with the attend sentence condition showing bilateral activation, while the dual task condition showed a left hemispheric dominance. This shift suggests multiple language‐processing modes and may explain the underadditivity in activation observed in the current and previous studies. Hum. Brain Mapp, 2007. © 2006 Wiley‐Liss, Inc.
Keywords: fMRI, dual task, prefrontal cortex
INTRODUCTION
To function effectively in a world with many simultaneous inputs, people must have a mechanism that allows for the selection of a subset of those inputs for further processing while ignoring others. One question of interest concerns the degree of control that we can exercise over what information is processed, and the degree to which it is processed. Demonstrations like the Stroop effect illustrate that some highly automatic processes (in the Stroop case, word encoding) cannot be entirely suppressed. Several neuroimaging studies have examined attentional control of visual inputs [e.g., Corbetta, 1998; Maunsell and Cook, 2002] as well as auditory inputs [e.g. Hugdahl et al., 2003; Thomsen et al., 2004a, b]. Such studies show that brain activation is indeed modulated by strategic intention. For example, Hugdahl et al. [ 2003] found that during an auditory selective attention task, activation within the temporal cortex was modulated (relative to passive listening) as a function of the aspect of the speech stream (the stream consisted on vowels, pseudowords, and words) that was selected for processing.
However, few neuroimaging studies have examined selective attention using two concurrent high‐level cognitive tasks in different modalities. The current study examines a dual task in which the two tasks were visual mental rotation and auditory sentence comprehension. Both task stimuli were continuously presented in a block design, creating two concurrent event streams. Participants were instructed to attend to the sentences only, attend to rotations only (selective attention conditions), or attend to both (the dual task condition). In this design, the neural fate of both the ignored and attended stimuli can be examined.
These auditory language and mental rotation tasks have been shown to involve primarily nonoverlapping neural systems. The neural system supporting sentence comprehension includes the classic language areas, the left superior/middle temporal cortex, the left inferior frontal gyrus, and, to a lesser extent, homologous areas on the right [Just et al., 1996; Schlosser et al., 1998]. The neural system supporting mental rotation includes the left and right parietal regions and, to some extent, the inferior temporal cortex [Carpenter et al., 1999; Cohen et al., 1996; Tagaris et al., 1997]. A previous dual task study using similar tasks demonstrated the separable nature of the underlying neural architecture that supports these two tasks [Just et al., 2001]. This study also found that during dual task performance there appeared to be interdependence between these topographically separable networks, such that the amount of activation in each system elicited in the dual task was less than that elicited when each task was performed alone. In other words, the sum of the activations observed in the two single tasks was much more than the activations in the dual task. Here we further explore this interdependence examining the effects of selective attention instructions in a dual task condition.
Prefrontal executive areas also come into play in dual task situations, presumably exercising some coordination or control function. For example, selective attention studies in which both tasks use the same modality stimuli (e.g., dichotic listening [Thomsen et al., 2004a, b] and visual selective attention [Corbetta, 1998; Maunsell and Cook, 2002]) have typically found a prefrontal/posterior cortical network involved (the posterior region being either temporal or parietal, depending on stimulus modality). A study examining selective and divided attention in which the two tasks use different input modalities (auditory and visual) reported activation of a prefrontal region during the divided attention condition, but failed to observe prefrontal activity during selective attention to either the visual or the auditory stimuli [Loose et al., 2003]. The prefrontal activity may have been necessary primarily when there was competition for sensory processing resources; namely, when the two tasks used the same modality.
The current study examines selective attention to either an auditory language comprehension task or a visuospatial mental rotation task. The primary goal of the study was to determine how selective attention instructions affect the brain activity associated with two concurrently presented tasks that use different modalities. For example, is the activation in language‐processing regions lower when attending to mental rotation than when attending to the sentences. A secondary goal was to further explore the interaction between two separable cortical networks during dual task processing compared to selective attention. A third question concerns the involvement of prefrontal regions during selective and divided attention in dual task performance.
PATIENTS AND METHODS
Participants
Fifteen right‐handed native English speakers (4 women), ages 18 to 26 who showed less than 40% errors in each condition (namely, 83% of all screened individuals), participated after signing a written consent form that had been approved by the University of Pittsburgh and Carnegie Mellon Institutional Review Boards. Data from 8 other participants were discarded because of excessive head motion (maximum head displacement > 0.4 voxels), technical problems during the scan, or because of inaccurate task performance.
Task and Stimulus Materials
There were three conditions, all of which entailed the concurrent presentation of visual mental rotation stimuli and auditory sentence comprehension stimuli. In the attend‐rotation condition, participants performed a mental rotation task involving drawings of complex, 3‐D figures originally used by Shepard and Metzler [ 1971]. The two figures were the same (with disparities of either 40° or 80°) in two thirds of the trials and different (mirror‐image isomorphs) in the other one third. Participants signaled their binary response using two handheld pushbuttons operated by their left thumb. Failure to respond within 5.5 s was treated as an error. The figures were presented in a sequence of four items, constituting an epoch. During each attend‐rotation epoch, sentences were concurrently presented in the auditory modality, but participants were instructed not to attend or respond to them. The duration of each epoch was 22.5 s.
In the attend‐sentence condition, participants listened to general knowledge sentences (e.g., “Botany is a biological science and it deals with the life structure and growth of plants”) that they verified as true or false while the Shepard‐Metzler rotation‐task figures were being displayed. Two‐thirds of the sentences were true and one‐third were false, and participants signaled their response using two handheld pushbuttons operated by their right thumb. Each sentence item took ∼6 s to present. A short tone sounded at the end of each sentence, and failure to respond within the next 3 s was treated as an error. The sentences were stored as digitized files and were presented using electrostatic headphones with a full frequency range MRI transducer. To ensure that the participants were exposed to both the rotation and the sentence stimuli during the attend sentences condition they were instructed to “keep eyes open and look at the rotation stimuli, just do not solve the problem.”
In the attend‐both condition, participants were told to pay equal attention to both tasks and to respond to both, using the appropriate hand (i.e., sentences with the right hand and rotations with the left).
The presentation of the stimuli was the same in the three conditions. The stimuli were presented in epochs consisting of three sentences and four rotations. Each sentence presentation of ∼6 s followed by a 1.5‐s blank interval consumed 7.5 s, such that the sequence of 3 sentences within each epoch consumed 22.5 s. It should be noted that participants responses could be recorded up to 1.5 s into the following sentence. Each rotation item was followed by a 0.5‐s blank interval and consumed 5.5 s, with the four rotation items in an epoch consuming 22 s. The asynchrony in the presentations of the items in the two tasks led to the responses to the two types of items generally occurring at different times, avoiding conflict in response mechanisms. The stimulus presentation intervals and the delay that followed were based on pilot behavioral data that indicated that the allotted time would accommodate the great majority of responses.
The study consisted of four epochs each of the three conditions: attend‐rotation, attend‐sentence, and attend‐both conditions. There were always two epochs of the same type presented in sequence, as an epoch pair. The order of the six epoch pairs was counterbalanced within a session. Across participants, three different orders were used (BRSSRB, SBRRBS and RSBBSR, where B = attend both, R = attend rotations, and S = attend sentences), so that each condition occurred equally often in each position. A 6‐s rest occurred between epochs. In addition, after every four epochs there was a 24‐s fixation condition in which the participant fixated a centered asterisk without performing any task; the fixation condition constituted a baseline measure of brain activation with which to compare experimental conditions. Prior to the start of each epoch, instructions were provided to inform the participants as to whether they were to attend to sentences alone, attend to rotations alone, or attend to both. Also, each participant underwent a practice session in which they were familiarized with the task prior to scanning.
Scanning Procedures
A 1.5 T GE Medical Systems (Milwaukee, WI) scanner using a GEMS quadrature birdcage head coil located at the MR Research Center of the University of Pittsburgh Medical Center was used to conduct this imaging study. Images were acquired in 14 adjacent oblique axial planes aligned to maximally cover the parietal and temporal lobes. The TR was 3000 ms, with TE = 50 ms, flip angle = 90°, a voxel size of 3.125 × 3.125 × 5 mm, field of view (FOV) = 400 × 200 mm, a 128 × 64 acquisition matrix, 5‐mm slice thickness, and a 1‐mm gap.
fMRI Data Analysis
The data were analyzed using statistical parametric mapping (SPM99 from the Wellcome Department of Cognitive Neurology, London, UK). Images were corrected for slice acquisition timing, motion‐corrected, normalized to the Montreal Neurological Institute (MNI) template, resampled to 2 × 2 × 2 mm voxels, and smoothed with an 8‐mm full‐width at half‐maximum (FWHM) Gaussian kernel to decrease spatial noise. At the individual level, statistical analysis was performed on each participant's data by using the General Linear Model and Gaussian random field theory as implemented in SPM99 [Friston et al., 1996]. For the random effects analysis on group data, one‐sample t‐tests were performed on contrast images obtained from each individual analysis. Activated brain areas surviving a threshold of P < 0.001 with a cluster size of k > 6 voxels (uncorrected for multiple comparisons) were rendered on a template brain in SPM. To compare the amount of activation in given regions across experimental conditions, a set of a priori, anatomically defined ROIs (regions of interest) in the MNI single‐subject dataset were defined [Tzourio‐Mazoyer et al., 2002], specifying locations in which there had previously been observed substantial clusters of activation in a similar dual task [Just et al., 2001]. Changes in mean signal intensity (relative to the fixation baseline) were computed from the averaged time‐course data extracted from each of these regions in each condition. These data were then compared across conditions using a within‐subjects ANOVA.
RESULTS
Behavioral Results
The behavioral measures indicated modest but significant increases in response times and errors when participants were attending to both tasks compared to when they were attending to only one. Nevertheless, participants performed well in all three conditions. The behavioral data were assessed using a one‐way analysis of variance (ANOVA), attend rotations (or sentences) vs. attend both. For mental rotation, the response times increased from 2835 ms to 3067 ms (F(1,14) = 7.24, P < 0.05), and the errors increased from 8.3% to 19.2% (F(1,14) = 14.70, P < 0.01) from the single (attend rotations) to the dual task condition. For sentence comprehension, the response times (measured from the end of the sentence) increased from 465 ms to 762 ms (F(1,14) = 30.46, P < 0.01), and errors increased from 4.6% to 12.8% (F(1,14) = 16.59, P < 0.01).
fMRI Results
Overview
One of the major findings of the current study is that the distribution of brain activation was amenable to strategic control, as determined by the attention‐dividing instructions. This strategic modulation of activation was particularly evident in the superior/middle temporal and occipito/parietal areas, which are central to the sentence and rotation tasks, respectively. Attending to either one of the tasks produced a high level of activation in that task and a much lower level of activation in the unattended task. Attend sentences produced much more activation in the superior/middle temporal area than did attend rotations, as shown in Figure 1. Analogously, attend rotations produced much more activation in the occipito/parietal cortex than did attend sentences, also shown in Figure 1. A second result was a laterality difference observed across the conditions, particularly in the language‐processing regions. The attend sentences condition elicited bilateral activation of temporal cortex, whereas attend both elicited only left hemisphere activation (as shown in Table I). Finally, the results replicated the previously observed decrease in activation from a single task condition to a dual‐task condition for tasks drawing on different neural substrates [Just et al., 2001]. If attend sentences and attend rotations are thought of as “single task” conditions, then one might expect that the activation in the dual task (attend both) might be the sum of the activations in the two “single task” conditions. However, the activation in attend both appears underadditive. In particular, the language‐related activation in the temporal regions was much lower during the attend both condition than during the attend sentences condition, as shown in Table II and Figure 2.
Figure 1.
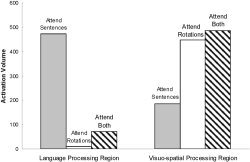
Sum of the activated voxels from a language‐processing region, the superior temporal cortex, bilaterally, and a visuospatial‐processing region, occipito/parietal cortex, bilaterally.
Table I.
Laterality ratios (volume in left hemisphere − volume in right hemisphere) / (volume in left + volume in right) for each cortical lobe
| Cortical lobe | Attend sentences | Attend rotations | Attend both |
|---|---|---|---|
| Frontal | 0.54 | −1 | 0.71 |
| Temporal | −0.18 | −1 | 0.75 |
| Parietal | 1 | 1 | n/a |
| Occipital | −1 | −0.84 | −0.87 |
Table II.
Experimental conditions compared to fixation
| Location of peak activation | Brodmann area | t(10) | Cluster size | MNI coordinates x, y, z | |
|---|---|---|---|---|---|
| Areas of activation observed for the attend sentences minus fixation | |||||
| Left | Middle frontal gyrus | BA 6 | 9.8 | 23 | −32, 2, 54 |
| Left | Precentral gyrus | BA 6 | 5 | 11 | −48, 2, 46 |
| Left | Inferior frontal gyrus | BA 9 | 5.15 | 34 | −54, 12, 32 |
| Left | Superior temporal gyrus | BA 38 | 8.1 | 113 | −48, 20, −14 |
| Left | Inferior frontal gyrus | BA 47 | 7.58 | −48, 30, −14 | |
| Left | Postcentral gyrus | BA 40 | 7.08 | 84 | −54, −32, 48 |
| Left | Inferior parietal lobule | BA 40 | 6.75 | −44, −38, 48 | |
| Left | Middle temporal gyrus | BA 21 | 6.26 | 194 | −60, −8, −8 |
| Left | Middle temporal gyrus | BA 21 | 6.17 | −54, −18, −6 | |
| Left | Middle temporal gyrus | BA 21 | 5.68 | −56, 4, −16 | |
| Right | Superior frontal gyrus | BA 8 | 4.87 | 32 | 2, 26, 50 |
| Right | Superior frontal gyrus | BA 8 | 4.56 | 11 | 2, 16, 48 |
| Right | Middle temporal gyrus | BA 21 | 8.96 | 259 | 54, −26, −6 |
| Right | Middle temporal gyrus | BA 21 | 6.04 | 60, −4, −12 | |
| Right | Middle temporal gyrus | BA 21 | 5.66 | 60, −14, −12 | |
| Right | Middle temporal gyrus | BA 22 | 4.91 | 14 | 68, −30, 4 |
| Right | Superior temporal gyrus | BA 38 | 4.8 | 6 | 48, 20, −20 |
| Right | Fusiform gyrus | BA 19 | 6.63 | 8 | 20, −62, −8 |
| Right | Middle occipital gyrus | BA 19 | 6.47 | 177 | 32, −90, 8 |
| Right | Middle occipital gyrus | BA 18 | 5.74 | 22, −100, 8 | |
| Right | Middle occipital gyrus | BA 18 | 4.69 | 32, −84, 14 | |
| Areas of activation observed for the attend rotations minus fixation | |||||
| Left | Inferior parietal lobule | BA 40 | 9.13 | 27 | −58, −30, 44 |
| Left | Middle occipital gyrus | BA 19 | 5.54 | 36 | −34, −84, 8 |
| Left | Middle occipital gyrus | BA 18 | 5.1 | −38, −90, 4 | |
| Right | Middle frontal gyrus | BA 9 | 5.36 | 81 | 56, 20, 32 |
| Right | Middle frontal gyrus | BA 8 | 4.67 | 54, 12, 40 | |
| Right | Middle occipital gyrus | BA 19 | 9.31 | 412 | 28, −86, 10 |
| Right | Superior occipital gyrus | BA 19 | 6.61 | 32, −84, 28 | |
| Right | Cuneus | BA 19 | 5.31 | 26, −94, 26 | |
| Right | Middle temporal gyrus | BA 21 | 6.54 | 9 | 60, 2, −18 |
| Areas of activation observed for the attend both minus fixation | |||||
| Left | Middle occipital gyrus | BA 19 | 7.28 | 31 | −52, −78, 0 |
| Left | Middle occipital gyrus | BA 18 | 5.64 | −42, −88, 0 | |
| Left | Middle occipital gyrus | BA 18 | 6.56 | 38 | −30, −98, 4 |
| Left | Middle frontal gyrus | BA 9 | 5.72 | 88 | −58, 10, 36 |
| Left | Inferior frontal gyrus | BA 9 | 5.2 | −58, 16, 30 | |
| Left | Middle frontal gyrus | BA 9 | 5.44 | 38 | −50, 26, 38 |
| Left | Middle frontal gyrus | BA 46 | 5.3 | −50, 40, 18 | |
| Left | Middle frontal gyrus | BA 46 | 4.91 | −52, 32, 26 | |
| Left | Middle temporal gyrus | BA 21 | 5.7 | 62 | −60, −16, −8 |
| Left | Superior temporal gyrus | BA 22 | 5.37 | −64, −18, 2 | |
| Right | Superior frontal gyrus | BA 10 | 5.3 | 8 | 38, 54, 28 |
| Right | Middle frontal gyrus | BA 10 | 4.92 | 13 | 36, 62, 14 |
| Right | Middle temporal gyrus | BA 21 | 4.71 | 9 | 52, −24, −10 |
| Right | Middle occipital gyrus | BA 19 | 4.7 | 15 | 50, −76, −2 |
| Right | Middle occipital gyrus | BA 18 | 11.28 | 440 | 30, −98, 10 |
| Right | Middle occipital gyrus | BA 18 | 8.56 | 24, −102, 6 | |
| Right | Middle occipital gyrus | BA 19 | 7.66 | 36, −92, 6 | |
Figure 2.
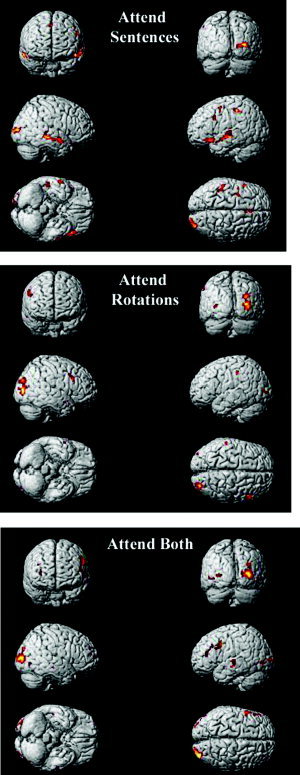
Average activation maps for each condition compared to a fixation baseline.
Attend Sentences
Attending to the auditory sentences elicited activation in a number of cortical regions often associated with auditory language processing, namely, the superior and middle temporal gyri, the left inferior frontal gyrus, and the left parietal cortex (Tables II, III; Fig. 3). The activation was approximately bilateral, with a slight right hemisphere bias (laterality ratio for temporal cortex was −0.18, Table I). We also compared across conditions the percent change (relative to fixation) in signal intensity in a key anatomical ROI (using Tzourio‐Mazoyer et al.'s [ 2002] ROI definition) that typically activates during auditory language comprehension, namely, the middle portion of the superior temporal gyrus. The percent change in signal intensity in this temporal ROI varied significantly across conditions, for both the left and right hemisphere ROI (left: F(1,10) = 9.23, P < 0.01; right: F(1,10) = 12.45, P < 0.01). Post‐hoc tests showed that the signal intensity during the attend sentences condition was significantly greater than during attend rotations (left: F(1,10) = 20.88, P < 0.01; right: F(1,10) = 20.08, P < 0.01) and attend both conditions (left: F(1,10) = 1.54, P >0.2; right: F(1,10) = 8.35, P < 0.05), as shown in Figure 3A. Attending to the auditory sentences also elicited activation within the left inferior frontal gyrus ROI (BA 47) extending into the anterior portion of temporal cortex. In addition to these auditory language‐processing regions, there was a cluster of activation in the right occipital lobe. The right occipital activation, related to visual processing, may be evidence that participants followed instructions and kept their eyes open during the attend sentences condition, and therefore viewed the rotation stimuli. In sum, the attention‐directing manipulation succeeded in modulating the amount of activation in key language areas, such that it was maximal in attend sentences, intermediate in attend both, and minimal in attend rotations.
Table III.
Selective attention tasks compared to dual task
| Location of peak activation | Brodmann area | t(10) | Cluster size | MNI coordinates x, y, z | |
|---|---|---|---|---|---|
| Areas of activation observed for attend sentences minus attend both | |||||
| Left | Insula | BA 13 | 4.7 | 6 | −46, 8, 16 |
| Left | Superior/middle temporal gyrus | BA 22/39 | 6.65 | 137 | −52, −54, 16 |
| 5.25 | −58, −58, 8 | ||||
| Left | Superior/middle temporal gyrus | BA 22 | 6.4 | 145 | −42, −30, −2 |
| 5.79 | −54, −10, −4 | ||||
| 5.51 | −48, −20, 0 | ||||
| Left | Superior temporal gyrus | BA 22 | 4.59 | 10 | −60, −24, 2 |
| Left | Superior frontal gyrus | BA 10 | 5.33 | 20 | −16, 64, 14 |
| Left | Superior frontal gyrus | BA 9 | 6.17 | 112 | −14, 48, 26 |
| Left | Inferior frontal gyrus | BA 45 | 6.03 | 27 | −56, 24, 4 |
| Left | Anterior cingulate | BA 24 | 4.2 | 6 | −8, 18, 22 |
| Left | Angular gyrus | BA 39 | 5.23 | 11 | −48, −70, 32 |
| Left | Postcentral gyrus | BA 43 | 4.85 | 16 | −56, −8, 22 |
| Right | Superior temporal gyrus | BA 13 | 7.04 | 41 | 44, −12, 16 |
| BA 13 | 5.18 | 46, −18, 8 | |||
| BA 13 | 5.06 | 38, −22, 16 | |||
| Right | Superior temporal gyrus | BA 39 | 4.56 | 9 | 48, −60, 30 |
| Right | Inferior temporal gyrus | BA 21 | 4.61 | 8 | 70, −18, −20 |
| Right | Middle temporal gyrus | BA 21 | 4.92 | 20 | 54, −28, 0 |
| Right | Parahippocampal gyrus | BA 30 | 4.71 | 20 | 20, −38, −2 |
| Right | Parahippocampal gyrus | BA 19 | 5.93 | 33 | 40, −40, 0 |
| Areas of activation observed for attend rotations minus attend both | |||||
| Left | Superior temporal gyrus | BA 22 | 6 | 16 | −64, 0, 8 |
| Right | Superior occipital gyrus | BA 19 | 7.11 | 32 | 40, −78, 26 |
| Right | Superior occipital gyrus | BA 19 | 5.1 | 10 | 42, −70, 20 |
| Right | Middle occipital gyrus | BA 37 | 5.02 | 59 | 38, −62, 10 |
| Right | Middle temporal gyrus | BA 19 | 4.81 | 36, −62, 18 | |
Figure 3.
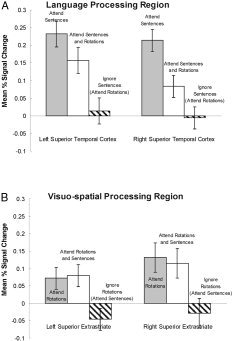
A: Signal change in a language‐processing region, the temporal cortex. B: Signal change in a visuospatial‐processing region, the occipito‐parietal cortex. The activation is amenable to strategic control, showing more activation when attending to a given stimulus and less when ignoring that stimulus.
Attend Rotations
Attending to the visual rotation stimuli activated the occipito/parietal cortex, as well as the posterior portion of the right middle frontal gyrus (Tables II, III; Fig. 2). The change in signal intensity within occipito/parietal cortex (i.e., in the superior extrastriate ROI) revealed a significant effect of condition in both hemispheres (left: F(1,10) = 4.97, P < 0.05; right: F(1,10) = 4.32, P < 0.05). Post‐hoc tests revealed that the change in signal intensity was similar in attend rotations and attend both (left: F < 1; right: F < 1), but was significantly lower in attend sentences (left: F(1,10) = 7.6, P < 0.05; right: F(1,10) = 10.39, P < 0.01), as shown in Figure 3B. In addition to occipito/parietal and right prefrontal activation, small clusters of activation were observed in the right superior temporal gyrus and the left inferior parietal cortex.
Attend Both
The divided attention condition elicited activation in both the language‐processing and visuospatial‐processing regions that were activated during the attend sentences and attend rotations conditions, respectively (Table III; Fig. 2). As in previous studies, the activation during the attend both condition appears to be underadditive, particularly in the language processing regions. As shown in Figures 1 and 2, there is significantly more activation in the temporal cortex for attend sentences than attend both. This difference indicates underadditivity.
DISCUSSION
One of the main goals of this study was to determine whether the distribution of brain activation between two tasks that use different input modalities and generally different neural substrates can be governed by attentional control, as it is with respect to the control of visual attention [Corbetta, 1998; Maunsell and Cook, 2002] and auditory attention [Hugdahl et al., 2003; Thomsen et al., 2004a, b]. The results show that the activation was in fact distributed in a way that corresponded to the attention‐dividing instructions. There were a number of additional findings related to performing two high‐level tasks concurrently.
Underadditivity of Activation in Dual Tasking
The dual task results presented here are similar to previous results in that they indicate an underadditivity, such that that the activation in the dual task is less than the sum of the activations in the two single tasks. Just et al. [ 2001] found that the activation volume related to performing the auditory sentence comprehension task and the mental rotation task alone was greater than when performing them together. That is, the amount of activation associated with one (or both) of the concurrently performed tasks is less than the activation in the corresponding single task. A similar effect was also found here when comparing selective attention to dual task performance, particularly for language processing, where the activation was considerably less in the attend both condition than in attend sentences. This new finding provides further support for the idea that there is an interdependence among cortical regions in how much activation they can sustain at a given time, probably because of the resource demands that they conjointly make during the performance of a cognitive task. In this particular study, one possible manifestation of the underadditivity, particularly during sentence processing, is that less elaborative processing occurred in the dual condition. For example, when resources are plentiful the language‐processing system may generate associations to enrich the meaning of a sentence, even though such elaborations may not be essential for adequate comprehension. However, when resources are limited, as is likely the case during the dual task condition, this elaborative processing may not occur. This idea is related to the “trimming” hypothesis that states that with practice neurons whose action is not essential for mapping the correct response onto the given stimulus become disengaged [Buchel et al., 1999; Gilbert et al., 2001; Rainer and Miller, 2000; Ramsey et al., 2004]. Here, trimming may be occurring as a function of the amount of available resources rather than the amount of practice.
Multitasking and Auditory Comprehension Lateralization
Auditory sentence processing has been found previously to elicit bilateral temporal activation [Michael et al., 2001], a finding that was replicated here in the attend sentences condition. However, during the attend both condition the activation became left lateralized by virtue of decreased right temporal activation. As mentioned above, this difference between conditions may be a result of a decrease in elective, elaborative processing during the attend both condition. This proposal is consistent with the right temporal area's known involvement in elaborative processes such as inference‐generation [Mason and Just, 2004]. So the temporal activation in auditory sentence comprehension is bilateral when the task is being performed by itself, or in a selective attention task that focuses on the comprehension (as in the attend sentences condition), but the activation becomes left‐dominant in attend both as the right temporal activation decreases.
Interestingly, no change in lateralization across conditions was observed in the visuospatial regions related to mental rotation; both attend rotations and attend both were highly right lateralized. Also, there is a smaller difference in the volume of activation between these two conditions, as shown in Figure 1, suggesting that the mental rotation processing is similar in both the attend rotations and attend both conditions. This stability of the lateralization pattern of the rotation‐related activation across conditions resembles the results of Just et al. [ 2001], where the rotation‐related activation was slightly right‐lateralized in both the single task and dual task conditions.
Dividing attention did not affect the activation in the rotation task very much (unlike the sentence task). The rotation‐related activation in the attend both condition resembled attend rotations but did not resemble the activation during attend sentences. It may be that the processing in auditory sentence processing is more adaptive to resource constraints, in the sense that there may be component processes that are executed only if resources permit. The processing associated with mental rotation, a much less familiar task, might be much less malleable. It appears that when resources are limited, as in the attend both condition, fewer resources are drawn from the language‐processing regions. Mental rotation, on the other hand, may be a more novel task in which there is only one processing strategy available that does not easily permit shortcuts.
An alternative, but related explanation for the differential activation patterns observed for the sentence and rotation tasks may be due to the automatic processing of auditory language stimuli. Because language processing is an overlearned, automatic process, it may be expected that it would be more difficult to ignore the sentence compared to the rotation stimuli. As a result, the sentences would interfere more with the rotation task during attend rotation than the rotation stimuli would interfere with the sentences during the attend sentence condition. The results presented here provide support for this hypothesis. The temporal cortex revealed a significant decrease in activation when comparing the attend sentence condition and the attend both condition, suggesting that differential processing was taking place during these two conditions—during attend sentences only sentences were processed, during attend both both sentences and rotations were processed. The spatial processing regions, on the other hand, revealed no difference when comparing attend rotations to attend both, suggesting that very similar processes were taking place—during attend rotations and attend both both rotations and sentences were being processed.
The Prefrontal Cortex
The prefrontal cortex has been considered central to the strategic control of attention [Frith, 2000; Jonides et al., 2002; Thomsen et al., 2004a, 2004b] and has been found to be activated in several dual task studies [D'Esposito et al., 1995; Dreher and Grafman, 2003; Loose et al., 2003; Szameitat et al., 2003; Thomsen et al., 2004a, 2004b]. However, its involvement during dual task studies has not been consistently observed [e.g., Adcock et al., 2000; Bunge et al., 2000; Goldberg et al., 1998; Just et al., 2001; Klingberg et al., 1998]. While there was a greater volume of activation in the left prefrontal region, particularly BA 6 and 9, for the dual condition (126 voxels) compared to both the attend sentences (68 voxels) and the attend rotations (0 voxels) conditions, the signal change was not greater and there were no reliable paired contrast results in this region. The failure to observe greater involvement of prefrontal cortex during the dual task condition is in conflict with some previous studies. The anterior cingulate, in particular, has been repeatedly activated in neuroimaging studies investigating attentional control and plays an essential role in theories of attention and cognitive control. However, a recent study has found that while the region is reliably activated, it does not play a necessary role in attention [Fellows and Farah, 2005]. A study of four patients with damage to the anterior cingulate showed normal performance on tests of cognitive control. Additionally, the current study failed to observe prefrontal activation, particularly anterior cingulate activation, related to selective attention. This absence of a prefrontal effect of selective attention is similar to that reported by Loose et al. [ 2003], suggesting that the prefrontal region may be involved in selective attention only when the same input modality is used for both the selected and the ignored stimuli.
The underadditivity and the laterality effects observed raise a number of questions for future study. One such question concerns the effect of automaticity on dual task performance. A sentence comprehension task would be less automatic, for example, if the sentences were presented in the weaker second language of bilingual participants. Would a less automatic task of this kind that nevertheless entails language comprehension evoke more frontal activation to coordinate the execution of the two tasks? The interest of such issues lies in exploring how automaticity and other factors might extend the outer limits of human performance during multitasking.
REFERENCES
- Adcock A, Constable RT, Gore JC, Goldman‐Rakic PS (2000): Functional neuroanatomy of executive processes involved in dual‐task performance. Proc Natl Acad Sci U S A 97: 3567–3572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchel C, Coull JT, Friston JJ (1999): The predictive value of changes in effective connectivity for human learning. Science 283: 1538–1541. [DOI] [PubMed] [Google Scholar]
- Bunge SA, Klingberg T, Jacobsen RB, Gabrieli JDE (2000): A resource model of the neural basis of executive working memory. Proc Natl Acad Sci U S A 97: 3573–3578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpenter PA, Just MA, Keller TA, Eddy WF, Thulborn KR (1999): Graded functional activation in the visuo‐spatial system with the amount of task demand. J Cogn Neurosci 11: 9–24. [DOI] [PubMed] [Google Scholar]
- Cohen MS, Kosslyn SM, Breiter HC, DiGirolamo GJ, Thompson WL, Anderson AK, Bookheimer SY, Rosen BR, Belliveau J (1996): Changes in cortical activity during mental rotation: a mapping study using functional MRI. Brain 119: 89–100. [DOI] [PubMed] [Google Scholar]
- Corbetta M (1998): Frontoparietal cortical networks for directing attention and the eye to visual locations: identical, independent, or overlapping neural systems? Proc Natl Acad Sci U S A 95: 831–838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Esposito M, Detre JA, Alsop DC, Shin RK, Atlas S, Grossman M (1995): The neural basis of the central executive system of working memory. Nature 378: 279–281. [DOI] [PubMed] [Google Scholar]
- Dreher J‐C, Grafman J (2003): Dissociating the roles of the rostral anterior cingulate and the lateral prefrontal cortices in performing two tasks simultaneously or successively. Cereb Cortex 13: 329–339. [DOI] [PubMed] [Google Scholar]
- Fellows LK, Farah MJ (2005): Is anterior cingulated cortex necessary for cognitive control? Brain 128: 788–796. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes A, Poline J‐B, Price CJ, Frith CD (1996): Detecting activations in PET and fMRI: levels of inference and power. Neuroimage 4: 223–235. [DOI] [PubMed] [Google Scholar]
- Frith CD (2000): The role of dorsolateral prefrontal cortex in the selection of action as revealed by functional imagine In: Monsell S, Driver J, editors. Control of cognitive processes, XVII ed. Cambridge, MA: MIT Press; p 549–566. [Google Scholar]
- Gilbert CD, Sigman M, Crist RE (2001): The neural basis of perceptual learning. Neuron 31: 681–697. [DOI] [PubMed] [Google Scholar]
- Goldberg TE, Berman KF, Fleming K, Ostrem J, Van Horn JD, Esposito G, Mattay VS, Gold JM, Weinberger DR (1998): Uncoupling cognitive workload and prefrontal cortical physiology: a PET rCBF study. Neuroimage 7: 296–303. [DOI] [PubMed] [Google Scholar]
- Hugdahl K, Thomsen T, Ersland L, Rimol LM, Niemi J (2003): The effects of attention on speech perception: an fMRI study. Brain Lang 85: 37–48. [DOI] [PubMed] [Google Scholar]
- Jonides J, Badre D, Curtis C, Thompson‐Schill SL, Smith EE (2002): Mechanisms of conflict resolution in prefrontal cortex In: Stuss DT, Knight RT, editors. Principles of frontal lobe function. New York: Oxford University Press; p 233–245. [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR (1996): Brain activation modulated by sentence comprehension. Science 274: 114–116. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Emery L, Zajac H, Thurlborn KR (2001): Interdependence of nonoverlapping cortical systems in dual cognitive tasks. Neuroimage 14: 417–426. [DOI] [PubMed] [Google Scholar]
- Klingberg T (1998): Concurrent performance of two working memory tasks: potential mechanisms of interference. Cereb Cortex 8: 593–601. [DOI] [PubMed] [Google Scholar]
- Loose R, Kaufmann C, Auer DP, Lange KW (2003): Human prefrontal and sensory cortical activity during divided attention tasks. Hum Brain Mapp 18: 249–259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason RA, Just MA (2004): How the brain processes causal inferences in text: a theoretical account of generation and integration component processes utilizing both cerebral hemispheres. Psychol Sci 15: 1–7. [DOI] [PubMed] [Google Scholar]
- Maunsell JHR, Cook EP (2002): The role of attention in visual processing. Philos Trans R Soc Lond B Biol Sci 357: 1063–1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michael EB, Keller TA, Carpenter PA, Just MA (2001): An fMRI investigation of sentence comprehension by eye and by ear: modality fingerprints on cognitive processes. Hum Brain Mapp 13: 239–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rainer G, Miller EK (2000): Effects of visual experience on the representation of objects in the prefrontal cortex. Neuron 27: 179–189. [DOI] [PubMed] [Google Scholar]
- Ramsey NF, Jansma JM, Jager G, Van Raalten T, Kahn RS (2004): Neurophysiological factors in human information processing capacity. Brain 127: 517–525. [DOI] [PubMed] [Google Scholar]
- Schlosser MJ, Aoyagi N, Fulbright RK, Gore JC, McCarthy G (1998): Functional MRI studies of auditory comprehension. Hum Brain Mapp 6: 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepard RN, Metzler J (1971): Mental rotation of three‐dimensional objects. Science 171: 701–703. [DOI] [PubMed] [Google Scholar]
- Szameitat AJ, Schubert T, Muller K, von Cramon DY (2002): Localization of executive functions in dual‐task performance with fMRI. J Cogn Neurosci 14: 1184–1199. [DOI] [PubMed] [Google Scholar]
- Tagaris GA, Kim S‐G, Strupp JP, Andersen P, Ugurbil K, Georgopoulos AP (1997): Mental rotation studied by functional magnetic resonance imaging at high field (4 Tesla): performance and cortical activation. J Cogn Neurosci 9: 419–432. [DOI] [PubMed] [Google Scholar]
- Thomsen T, Rimol LM, Ersland L, Hugdahl K (2004a): Dichotic listening reveals functional specificity in prefrontal cortex: an fMRI study. Neuroimage 21: 211–218. [DOI] [PubMed] [Google Scholar]
- Thomsen T, Specht K, Hammar A, Nyttingnes J, Ersland L, Hugdahl K (2004b): Brain localization of attentional control in different age groups by combining functional and structural MRI. Neuroimage 22: 912–919. [DOI] [PubMed] [Google Scholar]
- Tzourio‐Mazoyer N, Landeau B, Papthanassiou D, Crivello F, Etard O, Delcroix N, Mazoyer B, Joliot M (2002): Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single‐subject brain. Neuroimage 15: 273–289. [DOI] [PubMed] [Google Scholar]