

Abstract
One of the most popular experimental paradigms for functional neuroimaging studies of working memory has been the n‐back task, in which subjects are asked to monitor the identity or location of a series of verbal or nonverbal stimuli and to indicate when the currently presented stimulus is the same as the one presented n trials previously. We conducted a quantitative meta‐analysis of 668 sets of activation coordinates in Talairach space reported in 24 primary studies of n‐back task variants manipulating process (location vs. identity monitoring) and content (verbal or nonverbal) of working memory. We found the following cortical regions were activated robustly (voxelwise false discovery rate = 1%): lateral premotor cortex; dorsal cingulate and medial premotor cortex; dorsolateral and ventrolateral prefrontal cortex; frontal poles; and medial and lateral posterior parietal cortex. Subsidiary meta‐analyses based on appropriate subsets of the primary data demonstrated broadly similar activation patterns for identity monitoring of verbal stimuli and both location and identity monitoring of nonverbal stimuli. There was also some evidence for distinct frontoparietal activation patterns in response to different task variants. The functional specializations of each of the major cortical components in the generic large‐scale frontoparietal system are discussed. We conclude that quantitative meta‐analysis can be a powerful tool for combining results of multiple primary studies reported in Talairach space. Here, it provides evidence both for broadly consistent activation of frontal and parietal cortical regions by various versions of the n‐back working memory paradigm, and for process‐ and content‐specific frontoparietal activation by working memory. Hum Brain Mapp 25:46–59, 2005. © 2005 Wiley‐Liss, Inc.
Keywords: n‐back task, quantitative meta‐analysis, working memory, frontal cortex, parietal cortex
INTRODUCTION
Working memory has been described and discussed in various ways: as a cognitive system for the temporary storage and manipulation of remembered information [e.g., Baddeley,1986], as the type of memory that is active and only relevant for a short period [e.g., Fuster,1995; Goldman‐Rakic,1995], and, most specifically, as the process by which a remembered stimulus is held “on‐line” to guide behavior in the absence of external cues or prompts [Goldman‐Rakic,1996]. In part, this descriptive variability reflects the relative interests of those working with different primate species. Psychologists working mainly with humans often emphasize the organizational or higher‐order aspects of working memory tasks whereas those working with nonhuman primates tend to focus on those aspects of task performance related to the on‐line retention or short‐term storage of information. Nevertheless, although precise definitions vary, few disagree that working memory is a fundamental set of processes and an integral component of many cognitive operations, from complex decision making to selective attention [Baddeley,1986].
In recent years, variants of the “n‐back” procedure [Gevins and Cutillo,1993] have been employed in many human studies to investigate the neural basis of working memory processes. In the most typical variant of this task, the volunteer is required to monitor a series of stimuli and to respond whenever a stimulus is presented that is the same as the one presented n trials previously, where n is a prespecified integer, usually 1, 2, or 3. The task requires on‐line monitoring, updating, and manipulation of remembered information and is therefore assumed to place great demands on a number of key processes within working memory. Across studies, many different types of stimuli have been used via various input modalities (visual [including spatial], auditory, and olfactory) making demands on different processing systems. Load is often varied up to 3‐back, although some authors have questioned the validity of results when the ability to successfully perform the task decreases [Callicott et al.,1999]. Parametric designs, comparing n = 1, n = 2, and n = 3 trials are often employed, although in some studies a 0‐back control condition, which requires participants to respond whenever a prespecified stimulus is presented, has been used. This condition does not require the manipulation of information within working memory.
Quantitative meta‐analysis techniques, such as activation likelihood estimation (ALE) [Turkeltaub et al.,2002], can provide unique insights about anatomical consistency of results between a group of related studies within the existing literature. Synthesis and comparison of results is clearly important where variability exists between subjects and between studies and can assist the formation of theories based upon complex and diverse data sets. Because of its widespread usage in a number of domains, the n‐back task is an ideal candidate for such an approach and a wealth of relevant data already exists in the published literature. In this study, a quantitative meta‐analysis was carried out on the n‐back task using the ALE method.
After consideration of the available literature, comprising all published n‐back studies that have included detailed reports of activation coordinates in standard stereotaxic space, two clear divisions between studies emerged. The first concerned the manner in which stimuli were presented; approximately half of all studies employed verbal stimuli (e.g., letters and words), whereas the remaining studies employed nonverbal stimuli (including shapes, faces, and pictures). The second division concerned the type of monitoring that was required during the n‐back task. In some studies, it was the identity of the stimulus that had to be monitored (e.g., is this the same face as the one presented n‐back?), whereas in others it was the location of the stimulus (e.g., is this face in the same location as the one presented n‐back?).
It was hypothesized that a meta‐analysis of the available data from the n‐back task would show concordance between studies in several activation regions. The dorsolateral frontal cortex (approximate Brodmann areas [BA] 9/46) has been implicated in numerous cognitive functions that are relevant to the n‐back task, including holding spatial information on‐line [Courtney et al.,1998; Goldman‐Rakic,1994; Jonides et al.,1993], monitoring and manipulation within working memory [Owen,1997; Petrides,1994], response selection [Rowe et al.,2000], implementation of strategies to facilitate memory [Bor et al.,2003,2004], organization of material before encoding [Fletcher et al.,1998], and verification and evaluation of representations that have been retrieved from long‐term memory [Dobbins et al.,2002, Rugg et al.,1998]. The mid‐ventrolateral frontal cortex (BA 45,47), has been specifically implicated in a similarly diverse but distinct set of cognitive processes that may be relevant to the n‐back task, including the “selection, comparison and judgment of stimuli held in short‐term and long‐term memory” [Petrides,1994], holding nonspatial information on‐line [Courtney et al.,1997; Goldman‐Rakic,1994], stimulus selection [Rushworth et al.,1997], the specification of retrieval cues [Dobbins et al.,2002], and the “elaboration encoding” of information into episodic memory [Henson et al.,1999; Wagner et al.,1998]. The parietal cortex has been shown to be involved in a wide variety of cognitive tasks and in the context of these experiments it is often difficult to untangle its precise function from that of the prefrontal cortex. Typically, this region has been thought of as involved in the implementation of stimulus response mapping [Andersen and Buneo,2003; Corbetta and Shulman,2002; Dreher and Grafman,2003; Kimberg et al.,2000; Miller and Cohen,2001; Rushworth et al.,2001], although it has also been described as a “buffer for perceptual attributes” [Callicott et al.,1999] and is thought to be involved in storage of working memory contents [e.g., Jonides et al.,1997]. Activity in the anterior cingulate cortex is often described in relation to increased effort, complexity, or attention [Callicott et al.,1999; for review see Duncan and Owen,2000] and this region also seems to play a role in error detection and response correction [e.g., Rama et al.,2001].
MATERIALS AND METHODS
Multiple literature searches were conducted using Medline to find all functional neuroimaging studies using a variant of the n‐back working memory paradigm that had reported data on multiple healthy subjects. The reference lists of these articles were additionally checked for relevant studies not identified by computerized literature searching. Studies could only be included in meta‐analysis using the ALE [Turkeltaub et al.,2002] method if the primary data were reported as stereotactic coordinates in Talairach (or Montreal Neurological Institute [MNI]) space. In addition, studies reporting only data on n = 1 versions of the task were excluded, as were those linking working memory with reward or calculation.
Several studies reported coordinates of activation defined by multiple task versus control condition comparisons; each set of coordinates was considered individually for inclusion in the ALE meta‐analysis. Studies reporting results of conjunction analyses, which identified activations common to multiple contrasts, were also included. Table I lists all of the contrasts included in the meta‐analysis by first author and year of publication. Codes assigned to the tables, number of coordinates taken from a given source, the value of n‐back used, and stimulus type are also shown. Because the Talairach system is defined such that left is negative, coordinates based on the radiological convention were corrected. In addition, the spatial normalization template was determined for each study and all foci reported in MNI space were converted to Talairach space [Brett et al.,2001].
Table I.
Primary studies of the n‐back paradigm included in meta‐analysis
| Publication | Contrast | Foci | N‐back | Stimulus |
|---|---|---|---|---|
| All n‐back WM studiesa | ||||
| Awh,1996 | 2‐back–search control | 12 | 0, 2 | Letters |
| Awh,1996 | 2‐back–rehearsal control | 9 | 0,2 | Letters |
| Braver,1997 | Monotonic increases as function of load | 12 | 0,1,2,3 | Letters |
| Braver,2001 | Working memory (2‐back) | 4 | 2 | Words, faces |
| Callicott,1999 | Function of load | 18 | 0,1,2,3 | Numbers |
| Carlson,1998 | 2‐back vs. 0‐back visuospatial | 28 | 2,0 | Shapes |
| Carlson,1998 | 1‐back vs. 0‐back visuospatial | 17 | 1,0 | Shapes |
| Carlson,1998 | 2‐back vs. 1‐back visuospatial | 26 | 2,1 | Shapes |
| Casey,1998 | Boston: memory–motor | 13 | 0,2 | Shapes |
| Casey,1998 | Madison memory–motor | 6 | 0,2 | Shapes |
| Casey,1998 | Minnesota memory–motor | 22 | 0,2 | Shapes |
| Casey,1998 | Pittsburg memory–motor | 21 | 0,2 | Shapes |
| Cohen,1994 | Areas of activation | 9 | 0,2 | Letters |
| Cohen,1997 | Task‐related activity | 27 | 0,1,2,3 | Letters |
| Dade,2001 | WM‐sensorimotor for odors | 18 | 0,2 | Odors |
| Dade,2001 | WM‐sensorimotor for faces | 24 | 0,2 | Faces |
| Druzgal,2001 | Linear increases | 12 | 0,1,2 | Faces |
| Hautzel,2002 | Conjunction of verbal, object, shape, spatial | 16 | 0,2 | Letters, shapes, pictures |
| Hautzel,2002 | Real object vs. spatial | 3 | 0,2 | Pictures, shapes |
| Hautzel,2002 | Spatial vs. real object | 6 | 0,2 | Shapes, pictures |
| Hautzel,2002 | Spatial vs. shape | 6 | 0,2 | Shapes |
| Hautzel,2002 | Spatial vs. verbal | 4 | 0,2 | Shapes, letters |
| Hautzel, 2000 | Verbal vs. spatial | 1 | 0,2 | Letters, shapes |
| Honey,2000 | Generic brain activation | 10 | 0,2 | Letters |
| Jonides,1997 | 3‐back‐control | 24 | 3 | Letters |
| Jonides,1997 | 2‐back‐control | 22 | 2 | Letters |
| Jonides,1997 | 1‐back‐control | 3 | 1 | Letters |
| Jonides,1997 | 0‐back‐control | 2 | 0 | Letters |
| Kim,2002 | Simple pictures | 7 | 0,2 | Pictures |
| Kim,2002 | English words | 9 | 0,2 | Words |
| Kim,2002 | Korean words | 6 | 0,2 | Words |
| Kim,2003 | Healthy comparison subjects | 8 | 0,2 | Words, Pictures |
| Martinkauppi,2000 | 3‐back vs. 1‐back | 32 | 3,1 | Auditory tones, shapes |
| Martinkauppi,2000 | 2‐back vs. 1‐back | 30 | 2,1 | Auditory tones, shapes |
| Nystrom,2000 | Shapes > letters | 4 | 0,1,2,3 | Letters, shapes |
| Nystrom,2000 | Letters > shapes | 2 | 0,1,2,3 | Shapes, letters |
| Nystrom,2000 | Interactions (letters vs. shapes) | 2 | 0,1,2,3 | Letters, shapes |
| Nystrom,2000 | Locations > letters | 5 | 0,3 | Letters |
| Nystrom,2000 | Interactions (letters vs. locations) | 3 | 0,3 | Letters |
| Nystrom,2000 | Locations > shapes | 2 | 0,2 | Shapes |
| Nystrom,2000 | Shapes > locations | 1 | 0,2 | Shapes |
| Nystrom,2000 | Interactions (shapes vs. locations) | 4 | 0,2 | Shapes |
| Owen,1999 | Spatial manipulation–visuomotor control | 4 | 0,2 | Shapes |
| Owen,1999 | Spatial manipulation–spatial span | 2 | 0,2 | Shapes |
| Ragland,2002 | 1‐back vs. 0‐back letters | 6 | 0,1 | Letters |
| Ragland,2002 | 1‐back vs. 0‐back fractals | 5 | 0,1 | Fractals |
| Ragland,2002 | 2‐back vs. 0‐back letters | 7 | 0,2 | Letters |
| Ragland,2002 | 2‐back vs. 0‐back fractals | 9 | 0,2 | Fractals |
| Ragland,2002 | 2‐back vs. 1‐back letters | 10 | 2,1 | Letters |
| Ragland,2002 | 2‐back vs. 1‐back fractals | 6 | 2,1 | Fractals |
| Rama,2001 | 2‐back vs. 0‐back vocal expression | 32 | 2,0 | Emotional words |
| Rama,2001 | 1‐back vs. 0‐back vocal expression | 24 | 1,0 | Emotional words |
| Schumacher,1996 | Memory‐control for visual input | 12 | 0,3 | Letters |
| Schumacher,1996 | Memory‐control for auditory input | 13 | 0,3 | Letters |
| Smith,1996 | Experiment 2 verbal | 6 | 3,0 | Letters |
| Smith,1996 | Experiment 2 spatial memory | 9 | 3,0 | Shapes |
| Smith,1996 | Experiment 3 verbal memory | 14 | 2,0 | Letters |
| Veltman,2003 | Load‐related increases in activity for n‐back | 11 | 0,1,2,3 | Letters |
| Zurowski,2002 | Main effect of WM | 8 | 0,2 | Letters |
| Identity monitoring: verbal stimulib | ||||
| Awh,1996 | 2‐back‐search control | 12 | 0,2 | Letters |
| Awh,1996 | 2‐back‐rehearsal control | 9 | 0,2 | Letters |
| Braver,1997 | Monotonic increases as function of load | 12 | 0,1,2,3 | Letters |
| Cohen,1994 | Areas of activation | 9 | 0,2 | Letters |
| Cohen,1997 | Task‐related activity | 27 | 0,1,2,3 | Letters |
| Honey,2000 | Generic brain activation | 10 | 0,2 | Letters |
| Jonides,1997 | 3‐back‐control | 24 | 3 | Letters |
| Jonides,1997 | 2‐back‐control | 22 | 2 | Letters |
| Jonides,1997 | 1‐back‐control | 3 | 1 | Letters |
| Jonides,1997 | 0‐back‐control | 2 | 0 | Letters |
| Kim,2002 | English words | 9 | 0,2 | Words |
| Kim,2002 | Korean words | 6 | 0,2 | Words |
| Nystrom,2000 | Letters > shapes | 2 | 0,1,2,3 | Letters |
| Ragland,2002 | 1‐back vs. 0‐back letters | 6 | 0,1 | Letters |
| Ragland,2002 | 2‐back vs. 0‐back letters | 7 | 0,2 | Letters |
| Ragland,2002 | 2‐back vs. 1‐back letters | 10 | 2,1 | Letters |
| Schumacher,1996 | Memory‐control for visual input | 12 | 0,3 | Letters |
| Schumacher,1996 | Memory–control for auditory input | 13 | 0,3 | Letters |
| Smith,1996 | Experiment 2 verbal | 6 | 3,0 | Letters |
| Smith,1996 | Experiment 3 verbal memory | 14 | 2,0 | Letters |
| Veltman,2003 | Load‐related increases in activity for n‐back | 11 | 0,1,2,3 | Letters |
| Identity monitoring: nonverbal stimulic | ||||
| Dade,2001 | WM‐sensorimotor for faces | 24 | 0,2 | Faces |
| Druzgal,2001 | Linear increases | 12 | 0,1,2 | Faces |
| Kim,2002 | Simple pictures | 7 | 0,2 | Pictures |
| Kim,2003 | Healthy comparison subjects | 8 | 0,2 | Words, pictures |
| Nystrom,2000 | Shapes > letters | 4 | 0,1,2,3 | Letters, shapes |
| Nystrom,2000 | Shapes > locations | 1 | 0,2 | Shapes |
| Ragland,2002 | 1‐back vs. 0‐back fractals | 5 | 0,1 | Fractals |
| Ragland,2002 | 2‐back vs. 0‐back fractals | 9 | 0,2 | Fractals |
| Ragland,2002 | 2‐back vs. 1‐back fractals | 6 | 2,1 | Fractals |
| Location monitoring: nonverbal stimulid | ||||
| Carlson,1998 | 2‐back vs. 0‐back visuospatial | 28 | 2,0 | Shapes |
| Carlson,1998 | 1‐back vs. 0‐back visuospatial | 17 | 1,0 | Shapes |
| Carlson,1998 | 2‐back vs. 1‐back visuospatial | 26 | 2,1 | Shapes |
| Casey,1998 | Pittsburg memory‐motor | 21 | 0,2 | Shapes |
| Casey,1998 | Boston memory‐motor | 13 | 0,2 | Shapes |
| Casey,1998 | Madison memory‐motor | 6 | 0,2 | Shapes |
| Casey,1998 | Minnesota memory‐motor | 22 | 0,2 | Shapes |
| Nystrom,2000 | Locations > shapes | 2 | 0,2 | Shapes |
| Owen,1999 | Spatial manipulation‐visuomotor control | 4 | 0,2 | Shapes |
| Owen,1999 | Spatial manipulation‐spatial span | 2 | 0,2 | Shapes |
| Smith,1996 | Experiment 2 spatial memory | 9 | 3,0 | Shapes |
For each variant of the n‐back task studied, levels of task difficulty and the type of stimulus used are listed. Lists of studies included in the meta‐analysis are shown for all working memory (WM) n‐back data, identity monitoring of verbal stimuli, identity monitoring of nonverbal stimuli, and location monitoring of nonverbal stimuli.
668 coordinates, 24 papers.
226 coordinates, 12 papers.
76 coordinates, 6 papers.
150 coordinates, 5 papers.
The main meta‐analysis comprised all coordinates of n‐back task related activation reported in the primary literature, regardless of task variation or stimuli type. Then coordinates for the meta‐analysis were broken into three groups: identity task with verbal stimuli (verbal + identity), identity task with nonverbal stimuli (nonverbal + identity), and location task with nonverbal stimuli (nonverbal + location). There was not a sufficient number of coordinates reported to support a meta‐analysis of location tasks with verbal stimuli. This parsing of the studies allowed for a comparison of the identity and location task within the larger group of nonverbal articles as well as a nonverbal/verbal comparison in the context of monitoring stimulus identity.
The equally weighted coordinates were used to form estimates of the activation likelihood for each voxel in the brain as described by Turkeltaub et al. [2002]. In brief, to allow for error in spatial localization related to inter‐subject variation in functional anatomy and inter‐study differences in data smoothing and registration, the reported loci of maximal activation were modeled as the peaks of 3‐D Gaussian probability density functions (PDFs) with full‐width half‐maximum (FWHM) = 10 mm. The probabilities of each voxel in standard space representing each primary locus of activation were combined to form a map of the ALE score at each voxel. ALE maps were thresholded by a permutation test controlling the false discovery rate (FDR) at P < 0.01 and clusters of suprathreshold voxels exceeding 200 mm3 in volume were defined as loci of brain activation in common across all studies included in the meta‐analysis.
Meta‐analytic comparisons between groups of studies using different variants of the n‐back paradigm were carried out using the permutation test described in more detail by Laird et al. [2005]. We noted that studies on identity monitoring of nonverbal stimuli were the least frequent (76 coordinates reported for nine contrasts in six studies), compared to studies on location monitoring of nonverbal stimuli (150 coordinates for 11 contrasts in five studies) and studies of identity monitoring of verbal stimuli (226 coordinates for 21 contrasts in 12 studies). To control comparisons between these n‐back paradigm variants for the different number of contrasts reported in the primary literature, we randomly sampled a subset of nine contrasts from the studies on verbal identity monitoring and nonverbal location monitoring before comparing these subsampled data to the observed data on nonverbal identity monitoring. Contrast‐based sampling of the larger groups appropriately preserves dependencies between coordinates reported for the same contrast in a given study. Maps of differential activation by n‐back task variants were thresholded with P < 0.01 (FDR corrected).
RESULTS
N‐Back Working Memory: All Studies
The meta‐analysis of all n‐back studies included 668 foci from 24 studies (Fig. 1). Six cortical regions were defined as consistently activated across all studies: (1) bilateral and medial posterior parietal cortex, including precuneus and inferior parietal lobules (approximate BA7,40); (2) bilateral premotor cortex (BA6,8); (3) dorsal cingulate/medial premotor cortex, including supplementary motor area (SMA; BA32,6); (4) bilateral rostral prefrontal cortex or frontal pole (BA10); (5) bilateral dorsolateral prefrontal cortex (BA9,46); and (6) bilateral mid‐ventrolateral prefrontal cortex or frontal operculum (BA45,47). Additionally, there was a focus of activation in medial cerebellum. The volume of each activation locus, its maximum ALE score, and Talairach coordinates of the peak score are summarized in Table II.
Figure 1.
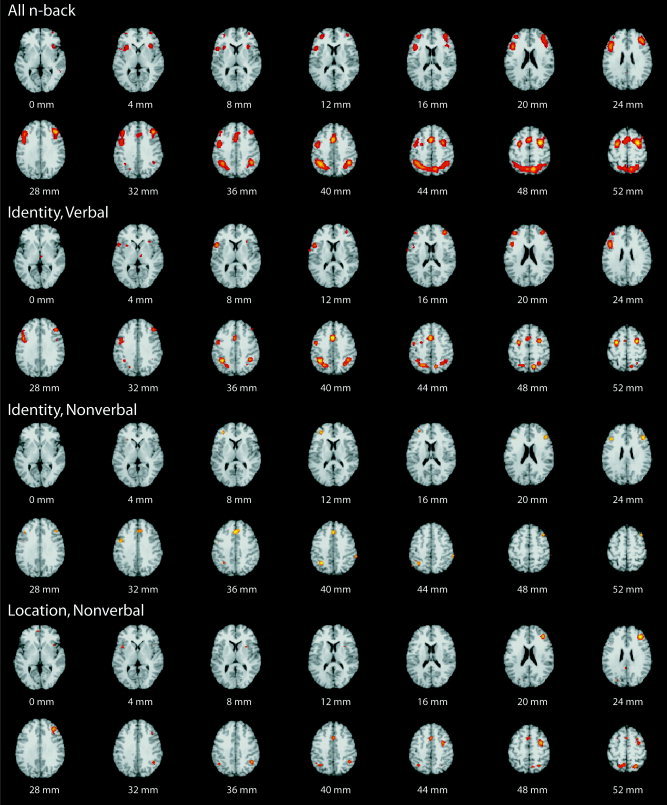
Meta‐analytic activation maps for all n‐back studies (top panel) and for three n‐back variants: verbal identity monitoring, nonverbal identity monitoring, and nonverbal location monitoring. Regions consistently activated across studies are color‐coded according to the probability of false discovery (voxelwise P < 0.01; FDR corrected). The right side of each section represents the right side of the brain; the z‐coordinate in Talairach space is indicated below each section. Regions of activation highlighted by these selected slices include lateral and medial premotor cortex (z = 52, 40), lateral and medial posterior parietal cortex (z = 52, 40), dorsolateral (z = 28) and ventrolateral (z = 4) prefrontal cortex and frontal poles (z = 4). For more anatomical details see Table II.
Table II.
Main regional loci of brain activation by meta‐analysis of n‐back working memory task variants in functional neuroimaging
| Region (BA) | x | y | z | ALE | Volume (mm3) |
|---|---|---|---|---|---|
| All n‐back working memory dataa | |||||
| Lateral premotor (6) | 28 | 4 | 50 | 0.0684 | 16,608 |
| −26 | 0 | 52 | 0.0488 | 3,016 | |
| −44 | −2 | 38 | 0.0361 | ? | |
| Dorsal cingulate/medial premotor (SMA) (32,6) | −2 | 12 | 42 | 0.0600 | |
| Dorsolateral prefrontal (46,9) | 40 | 32 | 30 | 0.0515 | 8,120 |
| −36 | 44 | 20 | 0.0267 | 992 | |
| −44 | 18 | 22 | 0.0422 | 1,2024 | |
| Ventrolateral prefrontal (45,47) | −30 | 18 | 6 | 0.0244 | |
| 32 | 20 | 4 | 0.0267 | 992 | |
| Frontal pole (10) | −36 | 44 | 20 | 0.0267 | 1,744 |
| Medial posterior parietal (7) | 10 | −66 | 48 | 0.0555 | 13,776 |
| Inferior parietal lobule (40) | −36 | −50 | 40 | 0.0551 | |
| 40 | −48 | 38 | 0.0535 | 8,096 | |
| Medial cerebellum | 2 | −64 | −24 | 0.0199 | 104 |
| Verbal stimuli, identity monitoring datab | |||||
| Lateral premotor (6) | 28 | 0 | 52 | 0.0334 | 2,096 |
| −26 | 2 | 52 | 0.0342 | 1,888 | |
| Dorsal cingulate/medial premotor (SMA) (32,6) | −2 | 12 | 42 | 0.0367 | 3,040 |
| Dorsolateral prefrontal (46,9) | 42 | 32 | 30 | 0.0220 | 1,144 |
| Ventrolateral prefrontal (44) | −50 | 12 | 8 | 0.0246 | 5,968 |
| −62 | 0 | 14 | 0.0138 | 216 | |
| Frontal pole (10) | −38 | 44 | 20 | 0.0204 | 896 |
| 36 | 46 | 18 | 0.0214 | 1,240 | |
| Medial posterior parietal (7) | 12 | −64 | 48 | 0.0302 | 1,312 |
| Inferior parietal lobule (40) | 30 | −58 | 42 | 0.0197 | 1,144 |
| 38 | −46 | 38 | 0.0249 | 1,928 | |
| −34 | −48 | 38 | 0.0379 | 5,136 | |
| Medial and lateral cerebellum | 2 | −64 | −24 | 0.0200 | 792 |
| 34 | −56 | −24 | 0.0180 | 768 | |
| 24 | −60 | −44 | 0.0170 | 712 | |
| −26 | −66 | −50 | 0.0163 | 328 | |
| Thalamus | 8 | −14 | 4 | 0.0121 | 208 |
| Nonverbal stimuli, identity monitoring datac | |||||
| Lateral premotor (6/8) | 38 | 20 | 50 | 0.0132 | 416 |
| Dorsal cingulate (32) | 0 | 26 | 36 | 0.0150 | 1,456 |
| Dorsolateral prefrontal (46,9) | 42 | 30 | 24 | 0.0141 | 1,064 |
| −44 | 4 | 32 | 0.0120 | 464 | |
| −40 | 26 | 24 | 0.0133 | 432 | |
| Frontal pole (10) | −32 | 42 | 10 | 0.0124 | 632 |
| −28 | 62 | −4 | 0.0125 | 280 | |
| Inferior parietal lobule (40) | −30 | −54 | 40 | 0.0129 | 792 |
| 58 | −36 | 44 | 0.0119 | 336 | |
| Nonverbal stimuli, location monitoring datad | |||||
| Lateral premotor (6) | 30 | 0 | 48 | 0.0204 | 1568 |
| 16 | 4 | 56 | 0.0124 | 352 | |
| Dorsal cingulate/medial premotor (SMA) (32,6) | 0 | 12 | 42 | 0.0166 | 1,144 |
| Dorsolateral prefrontal (46,9) | 36 | 36 | 24 | 0.0278 | 2,320 |
| Medial posterior parietal (7) | 10 | −58 | 54 | 0.0160 | 1,568 |
| 24 | −60 | 52 | 0.0172 | 824 | |
| 10 | −48 | 64 | 0.0115 | 200 | |
| Inferior parietal lobule (40) | −34 | −58 | 42 | 0.0136 | 704 |
| 42 | −50 | 36 | 0.0192 | 848 |
668 coordinates, 24 papers.
226 coordinates, 12 papers.
76 coordinates, 6 papers.
150 coordinates, 5 papers.
BA, approximate Brodmann area; ALE, activation likelihood estimation.
Verbal and Nonverbal Identity‐Monitoring Variants of the N‐Back Task
We then considered separately only those 226 coordinates reported in 12 studies of n‐back task variants demanding identity monitoring of verbal stimuli (Fig. 1; Table II). Broadly speaking, the same six cortical regions were identified by this meta‐analysis, namely lateral premotor cortex, dorsal cingulate and medial premotor cortex, dorsolateral and ventrolateral prefrontal cortex, frontal pole, and bilateral and medial posterior parietal cortex. There was more subcortical activation in medial and lateral cerebellum and thalamus. Many coordinates for peak ALE scores in these regions were within a few millimeters of the corresponding coordinates identified by the meta‐analysis of all studies. It was notable that maximum ALE scores and volumes of activation foci were generally reduced compared to the size and salience of activated loci identified by meta‐analysis of the larger set of coordinates drawn from all n‐back studies (Table II).
An analogous, separate analysis of the 76 coordinates reported by six studies of identity monitoring of nonverbal stimuli demonstrated again a broadly similar system of prefrontal, premotor, dorsal cingulate and posterior parietal activation (Fig. 1; Table II). The activation pattern was overall less salient and extensive than it was for the more numerous studies entailing identity monitoring of verbal stimuli; in particular, there was no left ventrolateral prefrontal cortical focus in the nonverbal data.
Nonverbal Location‐Monitoring Variants of the N‐Back Task
We also conducted a separate meta‐analysis of the 150 coordinates reported in five studies entailing location monitoring of nonverbal stimuli (Fig. 1; Table II). The resulting activation pattern was broadly similar to those obtained from the previous analyses, implicating prefrontal, premotor, and posterior parietal cortex.
Differential Activation Maps
Formal comparison between verbal and nonverbal identity‐monitoring studies demonstrated relatively enhanced activation by verbal tasks in left ventrolateral prefrontal cortex, medial and bilateral premotor cortex, bilateral medial posterior parietal cortex, and thalamus, as well as some small foci of enhanced activation by nonverbal tasks in frontal pole and dorsal cingulate regions (Fig. 2).
Figure 2.
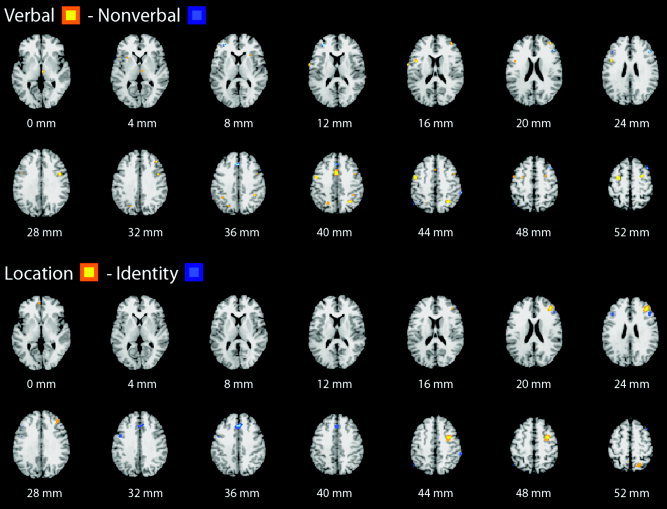
Meta‐analytic differential activation maps comparing verbal vs. nonverbal identity‐monitoring studies (top panel) and nonverbal location monitoring vs. nonverbal identity monitoring (bottom panel). Regions differentially activated between groups of studies are color‐coded according to the probability of false discovery (voxelwise P < 0.01; FDR corrected). The right side of each section represents the right side of the brain; the z‐coordinate in Talairach space is indicated below each section.
The comparison between location and identity monitoring of nonverbal stimuli demonstrated relatively enhanced activation by location monitoring in right dorsolateral prefrontal and lateral premotor cortex and right medial posterior parietal cortex; and relatively enhanced activation by identity monitoring in dorsal cingulate/medial premotor cortex (Fig. 2).
DISCUSSION
Although the importance of frontal and parietal regions in working memory is largely undisputed, no consensus has yet been reached regarding the fractionation of functions across these regions. We first review the results of the main meta‐analysis that comprised all loci of activation reported by primary studies using the n‐back paradigm, and discuss the likely functional roles of each main brain region that was shown in this way to be activated consistently across studies. We then discuss the functional implications of the subsidiary meta‐analyses focusing on differential activation by verbal and nonverbal stimuli, and differential activation by identity and location monitoring of nonverbal stimuli.
Dorsolateral Prefrontal Cortex
Perhaps the most widely and passionately debated issue to have emerged in this field in recent years concerns the functional relationship between the dorsal and ventral regions of the lateral frontal cortex and, more specifically, how the executive processes assumed to be dependent upon these regions might contribute to aspects of working memory. The region of dorsolateral prefrontal cortex identified by the main meta‐analysis of these n‐back data has been implicated frequently in many previous studies of working memory [for reviews see D'Esposito et al.,1998; Owen,1997,2000]. In fact, the coordinates of maximum ALE score (−44, 18, 22) are almost identical to those reported recently in a study investigating how this region contributes to the strategic reorganization and control of working memory contents [Bor et al.,2003]. In that study, a region of increased signal intensity was observed at (−46, 13, 21) in a spatial working memory task that required the reorganization of remembered material into higher‐level groups or “chunks” when compared to an unstructured condition in which grouping did not occur. An opportunity to reorganize materials into familiar or regular structures can increase working memory capacity, sometimes very substantially [Ericcson et al.,1980], and is a component of many working memory tasks, including the n‐back task. Similar results have been reported recently in the verbal domain [Bor et al.,2004] when memory for structured digit sequences (e.g., 2, 4, 6, 8, 9, 7, 5, 3) was compared to memory for unstructured sequences (e.g., 4, 7, 2, 5, 9, 6, 1, 8). Although structured sequences were easier to remember, they produced increased activation within the lateral frontal cortex relative to unstructured sequences, demonstrating that even when memory demand decreases, organization of working memory contents into higher‐level chunks is associated with increased prefrontal activity. Based on these results [Bor et al.,2003,2004], it was suggested that the lateral frontal cortex plays an essential role in increasing task performance, by selecting appropriate high‐level organizational chunks that then serve to facilitate memory by reducing the overall cognitive load. Neuropsychological data also support the possibility that this region of the prefrontal cortex contributes to the strategic control of working memory processing. Patients with frontal‐lobe damage are impaired on some but not all working memory tests, and in some cases deficits have been shown to relate to the inefficient use of organizational strategies that improve performance in healthy controls [Owen et al.,1996b; Petrides and Milner,1982]. Importantly, if such tasks are modified such that no obvious strategy exists to facilitate performance, frontal‐lobe patients can perform normally despite the fact that task difficulty may be increased substantially [Owen et al.,1996b]. N‐back tasks by their nature are very demanding and require volunteers to continually adjust the information held in working memory to incorporate the most recently presented stimulus while simultaneously rejecting or ignoring more temporally distant stimuli. Mnemonic strategies, which allow volunteers to simultaneously monitor such a series of stimuli and to make comparisons between each new stimulus and stimuli presented earlier in the sequence, are undoubtedly useful in such tasks and are therefore likely candidate processes for the cluster of activity observed in the dorsolateral frontal cortex in the current meta‐analysis.
Ventrolateral Prefrontal Cortex
A second prefrontal region identified by the main meta‐analysis was located just anterior to the insula within the mid‐ventrolateral frontal operculum. In humans, the mid‐ventrolateral frontal cortex lies below the inferior frontal sulcus and includes BA45 and 47. Like the mid‐dorsolateral frontal cortex, this mid‐ventrolateral frontal region has been implicated in many previous studies of working memory processes [for reviews see Owen,1997,2000]. In fact, the coordinates of maximum ALE score in this region (32, 20, 4) are close to the mean coordinates that have been reported previously (34, 19, −2) based on a systematic review of spatial and nonspatial working memory studies published before 1997 [Owen,1997]. Early functional neuroimaging studies that activated this region of the mid‐ventrolateral frontal cortex in humans tended to emphasize the explicit retrieval of one or a few pieces of information and the sequencing of responses based directly on that stored information. For example, in one PET study [Jonides et al.,1993; also see Smith et al.,1996], healthy volunteers were required to remember the location of three simultaneously presented stimuli and then to decide whether or not a probe circle occupied one of those same three locations after a 3‐s delay. Activation was observed in the mid‐ventrolateral frontal cortex at coordinates almost identical to that of the pooled results reported in the current analysis, but not in more dorsal regions of the frontal lobe. More recently, activation has been reported in this same region during all sorts of cognitive tasks that make varying demands on mnemonic and attentional processes. For example, the mid‐ventrolateral frontal cortex has been activated in tasks that require selection, comparison, and judgment of stimuli held in short‐ and long‐term memory [Petrides,1994], when spatial and nonspatial information is held on‐line [Courtney et al.,1997; Goldman‐Rakic,1994], during task switching [Dove et al.,2000], reversal learning [Cools et al.,2002], and stimulus selection [Rushworth et al.,1997], when the specification of retrieval cues is required [Dobbins et al.,2002], during the elaboration encoding of information into episodic memory [Henson et al.,1999; Wagner et al.,1998], and when judgments of word meaning are required [Kapur et al.,1994]. Although few of these studies set out specifically to investigate the functions of the ventrolateral frontal cortex, the combined results suggest that this region responds in a modality‐independent manner to a variety of explicit task demands.
In one recent study, event‐related fMRI was employed to test explicitly the hypothesis that activity in the mid‐ventrolateral frontal cortex is related specifically to the extent to which a volunteer explicitly intends to remember or retrieve a given stimulus, and the changes in attentional control that may be consequent upon such an intention [Dove et al.,2001]. In that study, conditions requiring volunteers to simply look at pictures of abstract art were compared to conditions in which they were instructed explicitly to remember similar stimuli for later retrieval. Looking, with no explicit instruction to remember, was associated with significant increases in signal intensity in the medial temporal lobe, but not the mid‐ventrolateral prefrontal cortex. When the task instructions were changed subtly to encourage the volunteers to remember the stimuli, significant increases in signal intensity were observed bilaterally, in the mid‐ventrolateral frontal cortex at coordinates very similar to those identified in the current meta‐analysis (38, 20, 0 and −34, 18, 2), but there was no concomitant increase within the medial temporal lobe region. Moreover, this cue‐related change in the mid‐ventrolateral frontal cortex, which was observed during both encoding and retrieval, was associated with significantly improved delayed recognition. These findings suggest that the implementation of an intended act or plan to remember or recall may be the common factor that underlies activation of the mid‐ventrolateral frontal cortex during previous neuroimaging studies of memory [e.g., Courtney et al.,1997; Henson et al.,1999; Jonides et al.,1993; Owen et al.,1996a,1999; Owen,2000; Smith et al.,1996; Wagner et al.,1998]. In the case of working memory tasks, such as the n‐back tasks included in the current meta‐analysis, this is likely to correspond to the relatively straightforward mapping of stimuli to responses that occurs whenever a specific target or nontarget stimulus is presented for inspection and comparison with previous stimuli. This notion also concurs well with other human brain imaging studies that have emphasized a role for the right ventrolateral prefrontal cortex in behavioral inhibition (or intentional stopping) using, for example, go‐no go tasks [Garavan et al.,1999; Konishi et al.,1999].
Lesion studies in the monkey also suggest that one role for the mid‐ventrolateral frontal cortex may be in the initiation and execution of many types of intended action. In the macaque, the mid‐ventrolateral frontal cortex lies below the principal sulcus on the inferior convexity and comprises areas BA12 or BA47/12 and BA45 [Carmichael and Price,1994]. Lesions of the ventrolateral frontal cortex, but not the more dorsal cortex surrounding the principal sulcus, cause impairments in nonspatial delayed‐matching‐to‐sample for single items [Mishkin and Manning,1978; Passingham,1975], spatial and nonspatial delayed alternation, the learning of arbitrary stimulus‐response associations [Gaffan et al.,1993; Petrides,1994], switching attention to behaviorally relevant aspects of the world [Dias et al.,1996], and even object matching when the sample and the match are simultaneously present and there is no delay component [Rushworth et al.,1997].
Rostral Prefrontal Cortex or Frontal Pole
Another frontal region for which few if any coherent theoretical accounts of function exist is BA10, known also as the frontal pole or rostral frontal cortex, that comprises the most anterior part of the frontal lobe. The main meta‐analysis identified a region of activation within BA10 (maximum ALE score at −36, 44, 20). The same region has been activated in numerous functional neuroimaging studies during tasks that are thought to tap functions as diverse as “processing of internal states” [Christoff and Gabrieli,2000], “retrieval mode,” “success monitoring” (or retrieval verification), source memory [Cabeza et al.,1997; Duzel et al.,1999; Lepage et al.,2000, Ranganath et al.,2000], prospective memory [Burgess et al.,2001], “branching” or “sub‐goal processing” [Koechlin et al.,1999], and reallocation of attentional resources and relational integration [Christoff et al.,2001; Kroger et al.,2002]. Many of these studies attempt to describe the functions of BA10 in terms of one particular type of task (e.g., source memory), and therefore have limited explanatory value outside of the immediate cognitive domain of that task. One recent analysis of the relevant literature has attempted to combine the core features of many models described above, but focuses on the common processes implied rather than the specific tasks employed [Ramnani and Owen,2004]. According to this view, frontopolar cortex is engaged when problems involve more than one discrete cognitive process, i.e., when the application of one cognitive operation (such as a rule) on its own is not sufficient to solve the problem as a whole and the integration of the results of two or more separate cognitive operations is required to fulfill the higher behavioral goal. The n‐back task is a perfect example of such a procedure, requiring simultaneous monitoring of a series of stimuli, ongoing adjustment of that information to incorporate recently presented items and reject temporally distant stimuli, and the drawing of comparisons (and the consequent switching of attention) between various items in the series. Multiple related cognitive operations can only be carried out successfully if they are coordinated, and it is suggested that the coordination of information processing and information transfer between multiple operations across supramodal cortex is an important aspect of BA10 function.
Bilateral and Medial Premotor Cortex
In the current meta‐analysis, a bilateral region between frontal areas BA6 and BA8 was identified in the meta‐analysis as being significantly concordant. Although somewhat medial in the left hemisphere, similar regions have again been reported frequently in many studies of working memory [e.g., Jonides et al.,1993; Owen et al.,1996a], but also in studies of spatial attention [Corbetta et al.,1993; Nobre et al.,1997]. On this basis, it has been suggested that activity in this region is related to the maintenance of visuospatial attention during working memory, a process that is likely to be particularly important where delays are imposed between a stimulus and a response to that stimulus [Owen,2000]. Such delays are by definition characteristic of the n‐back tasks whereby a response is determined not by the presence of a particular stimulus alone, but by the presence of a stimulus that is identical in some predefined respect to one that has been presented n trials previously.
Bilateral and Medial Posterior Parietal Cortex
In addition to signal changes in the prefrontal cortex, working memory studies frequently report concomitant activations in parietal lobe regions, particularly BA7 and BA40 [e.g., Awh et al.,1996; Jonides et al.,1993; Owen et al.,1996a]. In contrast, these areas are not often activated during studies of long‐term memory [Cabeza et al.,1997]. It is, therefore, not surprising that the current analysis of n‐back studies revealed a large focus of activation in these regions of parietal cortex. Smith and Jonides [1998] drew an important distinction between working memory storage and rehearsal mechanisms, suggesting that the left posterior parietal cortex is activated whenever short‐term storage of verbal memory is required. This involvement may be as part of a circuit mediating shifts of attention crucial for verbal rehearsal or may be part of a circuit involving storage itself [Jonides et al.,1998]. Analogous evidence from the same group suggests that spatial working memory may also be considered in terms of separable storage and rehearsal components with right‐hemisphere posterior parietal cortex being one component of a network that subserves spatial rehearsal [Smith and Jonides,1998]. Evidence from patients with language disorders also supports the claim that the left posterior parietal cortex mediates a storage function in verbal working memory. Lesions to this region can cause conduction aphasia and a common symptom is an inability to repeat back verbal material, even when the delay is minimal. In addition, conduction aphasics perform relatively normally on spatial working memory tasks [for discussion see Smith and Jonides,1998]. In contrast, patients with lesions of the right inferior parietal cortex are often impaired selectively in spatial processing and spatial memory, further supporting the possibility that this region plays a significant role in spatial rehearsal.
More recent studies have considered the possibility that there are two functionally distinct regions within the posterior parietal cortex that are relevant to studies of working memory [Ravizza et al.,2004]. Activation in the left dorsal aspect of the inferior parietal cortex has been observed frequently in contrasts of working memory load (e.g., delay and number of items) where the demands on attention are high. In contrast, activation in more ventral parietal areas has been found in relation to verbal versus nonverbal comparisons, with this region often seeming selectively active for verbal materials regardless of memory load. Ravizza et al. [2004] found support for this functional dissociation and concluded that whereas the dorsal inferior parietal cortex may support domain‐independent executive processes, the ventral inferior parietal cortex may support phonological encoding–recoding processes that are central to a variety of language tasks. Specifically, they suggest that the dorsal inferior parietal cortex may be “important for retaining temporal information” (in verbal, spatial, and visual working memory tasks), “for attentionally reactivating sources of information in neural regions,” “for rapid switching of attention,” or “for preparing for a given task.” In this context, perhaps it is unsurprising that this region emerged as a major cluster in the current analyses of n‐back studies. Ravizza et al. [2004] concluded that the ventral inferior parietal cortex may be involved in more basic speech processes, such as phonological discrimination and identification tasks, reading, “the mediation between auditory and articulatory representations,” and auditory imagery. This region thus may be more sensitive to the amount of phonological encoding or recoding that is required by a given condition rather than how much needs to be recalled [Ravizza et al.,2004]. In this context, it is surprising to note that none of the clusters within the current study fell within the inferior‐posterior limits described by Ravizza et al. [2004] for ventral inferior parietal cortex (z = 10–30), even in the subsidiary analysis of studies requiring the processing of verbal identity.
Functional Specialization of Frontoparietal Working Memory Systems
It is clear from the subsidiary activation maps for each major n‐back task variant (Fig. 1) and the differential activation maps (Fig. 2) that although all n‐back versions broadly activate the same frontal and parietal cortical regions, experimental differences in working memory process or content are associated with differential activation at a subregional level or in terms of hemispheric lateralization. For example, verbal identity monitoring (relative to nonverbal identity monitoring) was associated with enhanced activation in left ventrolateral prefrontal cortex, a region known to be important for (inner) speech. Nonverbal location monitoring (also relative to nonverbal identity monitoring) was associated with enhanced activation in right dorsolateral prefrontal, lateral premotor and posterior parietal cortex, a set of regions that have been described as a spatial attention network [Mesulam,2000].
On a more technical note, comparisons involving unequally sized groups of primary studies of different n‐back task variants are potentially confounded by the greater power to detect activation within a larger group of studies, as evidenced by the meta‐analytic activation statistics shown in Table II, which are consistently greater for the larger groups of studies. To address this issue, we have used a permutation test of the difference between two equally sized groups of statistical contrasts drawn from the primary literature [Laird et al.,2005] to test the hypothesis of working memory process‐ and content‐related specialization of frontal and parietal systems. This is methodologically novel and motivates further development of meta‐analytic permutation tests for more complex and sensitive comparisons between groups of primary studies.
CONCLUSIONS
Quantitative, comparative meta‐analysis of multiple functional neuroimaging studies can powerfully assist the research community to find commonalities and differences in brain activation elicited by a family of related experimental designs. We have found evidence for broadly consistent activation of six key frontal and parietal cortical regions by three species of the n‐back working memory paradigm. We have also found evidence for subregional and lateralized differences in activation compatible with reconfiguration of a generic frontoparietal network in response to experimental manipulations of working memory process and content.
REFERENCES
- Andersen RA, Buneo CA (2003): Sensorimotor integration in posterior parietal cortex. Adv Neurol 93: 159–177. [PubMed] [Google Scholar]
- Awh E, Jonides J, Smith EE, Schumacher EH, Koeppe RA, Katz S (1996): Dissociation of storage and rehearsal in verbal working memory: evidence from positron emission tomography. Physiological Science 7: 25–31. [Google Scholar]
- Baddeley AD (1986): Working memory. New York: Oxford University Press. [Google Scholar]
- Bor D, Cumming N, Scott CEM, Owen AM (2004): Prefrontal cortical involvement in encoding strategies, independent of stimulus modality. Eur J Neurosci 19: 3365–3370. [DOI] [PubMed] [Google Scholar]
- Bor D, Duncan J, Wiseman RJ, Owen AM (2003): Encoding strategies dissociate prefrontal activity from working memory demand. Neuron 37: 361–367. [DOI] [PubMed] [Google Scholar]
- Braver TS, Cohen JD, Nystrom LE, Jonides J, Smith EE, Noll DC (1997): A parametric study of prefrontal cortex involvement in human working memory. Neuroimage 5: 49–62. [DOI] [PubMed] [Google Scholar]
- Braver TS, Barch DM, Kelley WM, Buckner RL, Cohen NJ, Miezin FM, Snyder AZ, Ollinger JM, Akbudak E, Conturo TE, Petersen SE (2001): Direct comparison of prefrontal cortex regions engaged by working memory and long‐term memory tasks. Neuroimage 14: 48–59. [DOI] [PubMed] [Google Scholar]
- Brett M, Lancaster J, Christoff K (2001): Using the MNI brain with the Talairach atlas. Neuroimage 13(Suppl): 85. [Google Scholar]
- Burgess PW, Quayle A, Frith CD (2001): Brain regions involved in prospective memory as determined by positron emission tomography. Neuropsychologia 39: 545–555. [DOI] [PubMed] [Google Scholar]
- Cabeza R, Mangels J, Nyberg L, Habib R, Houle S, McIntosh AR, Tulving E (1997): Brain regions differentially involved in remembering what and when: a PET study. Neuron 19: 863–870. [DOI] [PubMed] [Google Scholar]
- Callicott JH, Mattay VS, Bertolino A, Finn K, Coppola R, Frank JA, Goldberg TE, Weinberger DR (1999): Physiological characteristics of capacity constraints in working memory as revealed by functional MRI. Cereb Cortex 9: 20–26. [DOI] [PubMed] [Google Scholar]
- Carlson S, Martinkauppi S, Rama P, Salli E, Korvenoja A, Aronen HJ (1998): Distribution of cortical activation during visuospatial n‐back tasks as revealed by functional magnetic resonance imaging. Cereb Cortex 8: 743–752. [DOI] [PubMed] [Google Scholar]
- Carmichael ST, Price JL (1994): Architectonic subdivision of the orbital and medial prefrontal cortex in the macaque monkey. J Comp Neurol 346: 366–402. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Cohen JD, O'Craven K, Davidson RJ, Irwin W, Nelson CA, Noll DC, Hu X, Lowe MJ, Rosen BR, Truwitt CL, Turski PA (1998): Reproducibility of fMRI results across four institutions using a spatial working memory task. Neuroimage 8: 249–261. [DOI] [PubMed] [Google Scholar]
- Christoff K, Gabrieli JDE (2000): The frontopolar cortex and human cognition: evidence for a rostrocaudal hierarchical organisation within the human prefrontal cortex. Psychobiology 28: 168–186. [Google Scholar]
- Christoff K, Prabhakaran V, Dorfman J, Zhao Z, Kroger JK, Holyoak KJ, Gabrieli JD (2001): Rostrolateral prefrontal cortex involvement in relational integration during reasoning. Neuroimage 14: 1136–1149. [DOI] [PubMed] [Google Scholar]
- Cohen JD, Forman SD, Braver TS, Casey BJ, Servan‐Schreiber D, Noll DC (1994): Activation of the prefrontal cortex in a nonspatial working memory task with functional MRI. Hum Brain Mapp 1: 293–304. [DOI] [PubMed] [Google Scholar]
- Cohen JD, Perlstein WM, Braver TS, Nystrom LE, Noll DC, Jonides J, Smith EE (1997): Temporal dynamics of brain activation during a working memory task. Nature 386: 604–608. [DOI] [PubMed] [Google Scholar]
- Cools R, Clark L, Owen AM, Robbins TW (2002): Defining the neural mechanisms of probabilistic reversal learning using event‐related functional magnetic resonance imaging. J Neurosci 22: 4563–4567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbetta M, Miezin FM, Shulman GL, Petersen SE (1993): A PET study of visuospatial attention. J Neurosci 13: 1202–1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbetta M, Shulman GL (2002): Control of goal‐directed and stimulus‐driven attention in the brain. Nat Rev Neurosci 3: 215–229. [DOI] [PubMed] [Google Scholar]
- Courtney SM, Petit L, Haxby JV, Ungerleider LG (1998): The role of prefrontal cortex in working memory: examining the contents of consciousness. Philos Trans R Soc Lond B Biol Sci 353: 1819–1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courtney SM, Ungerleider LG, Keil K, Haxby JV (1997): Transient and sustained activity in a distributed neural system for human working memory. Nature 386: 608–611. [DOI] [PubMed] [Google Scholar]
- Dade LA, Zatorre RJ, Evans AC, Jones‐Gotman M (2001): Working memory in another dimension: functional imaging of human olfactory working memory. Neuroimage 14: 650–660. [DOI] [PubMed] [Google Scholar]
- D'Esposito M, Aguirre GK, Zarahn E, Ballard D, Shin RK, Lease J (1998): Functional MRI studies of spatial and nonspatial working memory. Brain Res Cogn Brain Res 7: 1–13. [DOI] [PubMed] [Google Scholar]
- Dias R, Robbins TW, Roberts AC (1996): Dissociation in prefrontal cortex of affective and attentional shifts. Nature 380: 69–72. [DOI] [PubMed] [Google Scholar]
- Dobbins IG, Foley H, Schacter DL, Wagner AD (2002): Executive control during episodic retrieval: multiple prefrontal processes subserve source memory. Neuron 35: 989–996. [DOI] [PubMed] [Google Scholar]
- Dove A, Pollmann S, Schubert T, Wiggins CJ, von Cramon DY (2000): Prefrontal cortex activation in task switching: an event‐related fMRI study. Brain Res Cogn Brain Res 9: 103–109. [DOI] [PubMed] [Google Scholar]
- Dove A, Rowe JB, Brett M, Owen AM (2001): Neural correlates of passive and active encoding and retrieval: a 3T fMRI study. Neuroimage 13(Suppl): 660. [Google Scholar]
- Dreher JC, Grafman J (2003): Dissociating the roles of the rostral anterior cingulate and the lateral prefrontal cortices in performing two tasks simultaneously or successively. Cereb Cortex 13: 329–339. [DOI] [PubMed] [Google Scholar]
- Druzgal TJ, D'Esposito M (2001): Activity in fusiform face area modulated as a function of working memory load. Brain Res Cogn Brain Res 10: 355–364. [DOI] [PubMed] [Google Scholar]
- Duncan J, Owen AM (2000): Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends Neurosci 23: 475–483. [DOI] [PubMed] [Google Scholar]
- Duzel E, Cabeza R, Picton TW, Yonelinas AP, Scheich H, Heinze HJ, Tulving E (1999): Task‐related and item‐related brain processes of memory retrieval. Proc Natl Acad Sci USA 96: 1794–1799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ericcson KA, Chase WG, Falloon S (1980): Acquisition of a memory skill. Science 208: 1181–1182. [DOI] [PubMed] [Google Scholar]
- Fletcher PC, Shallice T, Dolan RJ (1998): The functional roles of prefrontal cortex in episodic memory. I. Encoding. Brain 121: 1239–1248. [DOI] [PubMed] [Google Scholar]
- Fuster JM (1995): Memory in the cerebral cortex: an empirical approach to neural networks in the human and nonhuman primate brain. Cambridge, MA: MIT Press; 372 p. [Google Scholar]
- Gaffan D, Murray EA, Fabre‐Thorpe M (1993): Interaction of the amygdala with the frontal lobe in reward memory. Eur J Neurosci 5: 968–975. [DOI] [PubMed] [Google Scholar]
- Garavan H, Ross TJ, Stein EA (1999): Right hemispheric dominance of inhibitory control: an event‐related functional MRI study. Proc Natl Acad Sci USA 96: 8301–8306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gevins AS, Cutillo BC (1993): Neuroelectric evidence for distributed processing in human working memory. Electroencephalogr Clin Neurophysiol 87: 128–143. [DOI] [PubMed] [Google Scholar]
- Goldman‐Rakic PS (1994): Working memory dysfunction in schizophrenia. J Neuropsychiatry Clin Neurosci 6: 348–357. [DOI] [PubMed] [Google Scholar]
- Goldman‐Rakic PS (1995): Architecture of the prefrontal cortex and the central executive. Ann N Y Acad Sci 769: 71–83. [DOI] [PubMed] [Google Scholar]
- Goldman‐Rakic PS (1996): The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive. Philos Trans R Soc Lond B Biol Sci 351: 1445–1453. [DOI] [PubMed] [Google Scholar]
- Hautzel H, Mottaghy FM, Schmidt D, Zemb M, Shah NJ, Muller‐Gartner HW, Krause BJ (2002): Topographic segregation and convergence of verbal, object, shape and spatial working memory in humans. Neurosci Lett 323: 156–160. [DOI] [PubMed] [Google Scholar]
- Henson RN, Shallice T, Dolan, RJ (1999): Right prefrontal cortex and episodic memory retrieval: a functional MRI test of the monitoring hypothesis. Brain 122: 1367–1381. [DOI] [PubMed] [Google Scholar]
- Honey GD, Bullmore ET, Sharma T (2000): Prolonged reaction time to a verbal working memory task predicts increased power of posterior parietal cortical activation. Neuroimage 12: 495–503. [DOI] [PubMed] [Google Scholar]
- Jonides J, Schumacher EH, Smith EE, Lauber EJ, Awh E, Minoshima S, Koeppe RA (1997): Verbal working memory load affects regional brain activation as measured by PET. J Cogn Neurosci 9: 462–475. [DOI] [PubMed] [Google Scholar]
- Jonides J, Schumacher EH, Smith EE, Koeppe RA, Awh E, Reuter‐Lorenz PA, Marshuetz C, Willis CR (1998): The role of parietal cortex in verbal working memory. J Neurosci 18: 5026–5034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonides J, Smith EE, Koeppe RA, Awh E, Minoshima S, Mintun MA (1993): Spatial working memory in humans as revealed by PET. Nature 363: 623–625. [DOI] [PubMed] [Google Scholar]
- Kapur S, Rose R, Liddle PF, Zipursky RB, Brown GM, Stuss D, Houle S, Tulving E (1994): The role of the left prefrontal cortex in verbal processing: semantic processing or willed action? Neuroreport 5: 2193–2196. [DOI] [PubMed] [Google Scholar]
- Kim JJ, Kim MS, Lee JS, Lee DS, Lee MC, Kwon JS (2002): Dissociation of working memory processing associated with native and second languages: PET investigation. Neuroimage 15: 879–891. [DOI] [PubMed] [Google Scholar]
- Kim JJ, Kwon JS, Park HJ, Youn T, Kang DH, Kim MS, Lee DS, Lee MC (2003): Functional disconnection between the prefrontal and parietal cortices during working memory processing in schizophrenia: a [15O]H2O PET study. Am J Psychiatry 160: 919–923. [DOI] [PubMed] [Google Scholar]
- Kimberg DY, Aguirre GK, D'Esposito M (2000): Modulation of task‐related neural activity in task‐switching: an fMRI study. Brain Res Cogn Brain Res 10: 189–196. [DOI] [PubMed] [Google Scholar]
- Koechlin E, Basso G, Pietrini P, Panzer S, Grafman J (1999): The role of the anterior prefrontal cortex in human cognition. Nature 399: 148–151. [DOI] [PubMed] [Google Scholar]
- Konishi S, Kawazu M, Uchida I, Kikyo H, Asakura I, Miyashita Y (1999): Contribution of working memory to transient activation in human inferior prefrontal cortex during performance of the Wisconsin Card Sorting Test. Cereb Cortex 9: 745–753. [DOI] [PubMed] [Google Scholar]
- Kroger JK, Sabb FW, Fales CL, Bookheimer SY, Cohen MS, Holyoak KJ (2002): Recruitment of anterior dorsolateral prefrontal cortex in human reasoning: a parametric study of relational complexity. Cereb Cortex 12: 477–485. [DOI] [PubMed] [Google Scholar]
- Laird AR, Fox PM, Price CJ, Glahn DC, Uecker AM, Lancaster JL, Turkeltaub PE, Kochunov P, Fox PT (2005): ALE meta‐analysis: controlling the false discovery rate and performing statistical contrasts. Hum Brain Mapp 25: 155–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lepage M, Ghaffar, Nyberg L, Tulving E (2000): Prefrontal cortex and episodic memory retrieval mode. Proc Natl Acad Sci USA 97: 506–511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinkauppi S, Rama P, Aronen HJ, Korvenoja A, Carlson S (2000): Working memory of auditory localization. Cereb Cortex 10: 889–898. [DOI] [PubMed] [Google Scholar]
- Mesulam M‐M (2000): Attentional networks, confusional states, and neglect syndromes In: Mesulam M‐M., editor. Principles of behavioral and cognitive neurology. New York: Oxford University Press; p 174–256. [Google Scholar]
- Miller EK, Cohen JD (2001): An integrative theory of prefrontal cortex function. Annu Rev Neurosci 24: 167–202. [DOI] [PubMed] [Google Scholar]
- Mishkin M, Manning FJ (1978): Nonspatial memory after selective prefrontal lesions in monkeys. Brain Res 143: 313–323. [DOI] [PubMed] [Google Scholar]
- Nobre AC, Sebestyen GN, Gitelman DR, Mesulam MM, Frackowiak RSJ, Frith CD (1997): Functional localization of the system for visuospatial attention using positron emission tomography. Brain 120: 515–533. [DOI] [PubMed] [Google Scholar]
- Nystrom LE, Braver TS, Sabb FW, Delgado MR, Noll DC, Cohen JD (2000): Working memory for letters, shapes and locations: fMRI evidence against stimulus‐based regional organization in human prefrontal cortex. Neuroimage 11: 424–446. [DOI] [PubMed] [Google Scholar]
- Owen AM (1997): The functional organization of working memory processes within human lateral frontal cortex: the contribution of functional neuroimaging. Eur J Neurosci 9: 1329–1339. [DOI] [PubMed] [Google Scholar]
- Owen AM (2000): The role of the lateral frontal cortex in mnemonic processing: the contribution of functional neuroimaging. Exp Brain Res 133: 3304. [DOI] [PubMed] [Google Scholar]
- Owen AM, Evans AC, Petrides M (1996a): Evidence for a two‐stage model of spatial working memory processing within the lateral frontal cortex: a positron emission tomography study. Cereb Cortex 6: 31–38. [DOI] [PubMed] [Google Scholar]
- Owen AM, Herrod NJ, Menon DK, Clark JC, Downey SP, Carpenter TA, Minhas PS, Turkheimer FE, Williams EJ, Robbins TW, Sahakian BJ, Petrides M, Pickard JD (1999): Redefining the functional organization of working memory processes within human lateral prefrontal cortex. Eur J Neurosci 11: 567–574. [DOI] [PubMed] [Google Scholar]
- Owen AM, Morris RG, Sahakian BJ, Polkey CE, Robbins TW (1996b): Double dissociations of memory and executive functions in working memory tasks following frontal lobe excisions, temporal lobe excisions or amygdalo‐hippocampectomy in man. Brain 119: 1597–1615. [DOI] [PubMed] [Google Scholar]
- Passingham R (1975): Delayed matching after selective prefrontal lesions in monkeys (Macaca mulata). Brain Res 92: 89–102. [DOI] [PubMed] [Google Scholar]
- Petrides M (1994): Frontal lobes and behaviour. Curr Opin Neurobiol 4: 207–211. [DOI] [PubMed] [Google Scholar]
- Petrides M, Milner B (1982): Deficits on subject‐ordered tasks after frontal‐ and temporal‐lobe lesions in man. Neuropsychologia 20: 249–262. [DOI] [PubMed] [Google Scholar]
- Ragland JD, Turetsky BI, Gur RC, Gunning‐Dixon F, Turner T, Schroeder L, Chan R, Gur RE (2002): Working memory for complex figures: an fMRI comparison of letter and fractal n‐back tasks. Neuropsychology 16: 370–379. [PMC free article] [PubMed] [Google Scholar]
- Rama P, Martinkauppi S, Linnankoski I, Koivisto J, Aronen H, Carlson S (2001): Working memory of identification of emotional vocal expressions: an fMRI study. Neuroimage 13: 1090–1101. [DOI] [PubMed] [Google Scholar]
- Ramnani N, Owen AM (2004): Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nat Rev Neurosci 5: 184–194. [DOI] [PubMed] [Google Scholar]
- Ranganath C, Johnson MK, D'Esposito M (2000): Left anterior prefrontal activation increases with demands to recall specific perceptual information. J Neurosci 20: 108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravizza SM, Delgado MR, Chein JM, Becker JT, Fiez JA (2004): Functional dissociations within the inferior prefrontal cortex in verbal working memory. Neuroimage 22: 562–573. [DOI] [PubMed] [Google Scholar]
- Rowe JB, Toni I, Josephs O, Frackowiak RS, Passingham RE (2000): The prefrontal cortex: response selection or maintenance within working memory? Science 288: 1656–1660. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Fletcher PC, Allan K, Frith CD, Frackowiak RS, Dolan RJ (1998): Neural correlates of memory retrieval during recognition memory and cued recall. Neuroimage 8: 262–273. [DOI] [PubMed] [Google Scholar]
- Rushworth MF, Nixon PD, Eacott MJ, Passingham RE (1997): Ventral prefrontal cortex is not essential for working memory. J Neurosci 17: 4829–4838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rushworth MF, Paus T, Sipila PK (2001): Attention systems and the organization of the human parietal cortex. J Neurosci 21: 5262–5271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schumacher EH, Lauber E, Awh E, Jonides J, Smith EE, Koeppe RA (1996): PET evidence for an amodal verbal working memory system. Neuroimage 3: 79–88. [DOI] [PubMed] [Google Scholar]
- Smith EE, Jonides J, Koeppe RA (1996): Dissociating verbal and spatial working memory using PET. Cereb Cortex 6: 11–20. [DOI] [PubMed] [Google Scholar]
- Smith EE, Jonides J (1998): Neuroimaging analyses of human working memory. Proc Natl Acad Sci USA 95: 12061–12068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA (2002): Meta‐analysis of the functional neuroanatomy of single‐word reading: method and validation. Neuroimage 16: 765–780. [DOI] [PubMed] [Google Scholar]
- Veltman DJ, Rombouts SA, Dolan RJ (2003): Maintenance versus manipulation in verbal working memory revisited: an fMRI study. Neuroimage 18: 247–256. [DOI] [PubMed] [Google Scholar]
- Wagner AD, Schacter DL, Rotte M, Koutstaal W, Maril A, Dale AM, Rosen BR, Buckner RL (1998): Building memories: remembering and forgetting of verbal experiences as predicted by brain activity. Science 281: 1188–1191. [DOI] [PubMed] [Google Scholar]
- Zurowski B, Gostomzyk J, Gron G, Weller R, Schirmeister H, Neumeier B, Spitzer M, Reske SN, Walter H (2002): Dissociating a common working memory network from different neural substrates of phonological and spatial stimulus processing. Neuroimage 15: 45–57. [DOI] [PubMed] [Google Scholar]