

Abstract
We investigated neural substrates of global/local processing of bilateral hierarchical stimuli using functional magnetic resonance imaging (fMRI). Subjects were presented with two compound letters that were displayed simultaneously in the left and right visual fields, respectively. In a steady‐state, block‐design paradigm, hemodynamic responses were recorded while subjects detected infrequent global or local targets presented in one hemifield in separate epochs of trials. While behavioural responses were more accurate and faster to global than local targets, attention to the global level of bilateral visual inputs induced stronger activations in the left and right temporal cortex relative to attention to the local level. However, attention to the local level generated stronger activations in bilateral superior parietal cortex compared with attention to the global level. The results suggest that distinct neural substrates in the temporal and parietal cortices are preferentially engaged in the global and local processing of bilateral visual inputs, respectively. Hum. Brain Mapping 22:321–328, 2004. © 2004 Wiley‐Liss, Inc.
Keywords: compound stimulus, global/local processing, fMRI, temporal/parietal cortex
INTRODUCTION
The neural substrates underlying the processing of global and local properties of hierarchically organized stimuli have been studied extensively in the last two decades. Using Navon‐type compound stimuli that are large letters composed of small letters [Navon, 1977], neurological studies found that brain lesions of the left parieto‐temporal area result in deficits of memory of local features [Delis et al., 1986] or slow responses to the local aspects of compound letters [Lamb et al., 1989, 1990; Robertson et al., 1988]. In contrast, right hemisphere lesions in the corresponding area damage the memory of global features or delay global responses. The findings of the lesion studies suggest that the right and left hemispheres dominate the global and local processing of compound stimuli, respectively.
Neurophysiological studies of healthy subjects extend the findings of neurological research. For example, by recording event‐related brain potentials (ERPs), Han et al. [1997, 1999a, 2000] showed that an early positive component elicited by compound stimuli was enlarged when subjects attended to the local rather than global levels of the compound stimuli. This positive component has maximum amplitudes over the posterior occipital areas and peaks at about 100 msec after sensory stimulation (P1), which is supposed to originate from the extrastriate cortex [Clark and Hillyard, 1996; Heinze et al., 1994]. Identification of local targets also elicited an occipito‐temporal negativity peaking between 200 and 300 msec (the posterior N2), larger than that of global targets [Han et al., 1997, 2000; Heinze and Münte, 1993; Heinze et al., 1998]. The late P3 component may show shorter peak latencies and larger amplitudes in the global than in the local conditions when behavioral data were characterized with a typical global precedence effect [Han et al., 1997; Proverbio et al., 1998]. The ERP findings indicate that global/local processing may be differentiated in the visual cortex as well as in higher level brain structures.
This proposal has been strengthened by recent neuroimaging studies that measured regional cerebral blood flow or hemodynamic responses from human subjects. For instance, Sasaki et al. [2001] and Han et al. [2002b] found stronger activations in the medial occipital cortex in association with attention to the global than local targets presented at fixation, which may result from a larger attention window when identifying global relative to local targets [Robertson et al., 1993]. Central compound stimuli also induced activations in the right middle occipital cortex associated with global attention conditions but in the left inferior occipital cortex associated with local attention [Fink et al., 1996; Han et al., 2002b]. However, the asymmetric occipital activations related to global and local processing were weakened when compound stimuli were presented unilaterally [Han et al., 2002b]. The asymmetric activations related to global and local processing were also observed over the occipito‐temporal cortical junctions [Martinez et al., 1997], which are consistent with the results of neurological research. Taken together, the aforementioned studies indicate that neural mechanisms at multiple levels of brain structures are engaged in the processing of global and local properties and are modulated by a number of factors such as stimulus locations.
Surprisingly, presentation of a single compound stimulus on each trial was employed in most of the previous studies of neural substrates of global/local processing. Nevertheless, observers are usually confronted with multiple hierarchical stimuli in the visual environment. Competitions exist between global and local processing of a single compound stimulus [Han et al., 1999b; Han and Humphreys, 2002]. When multiple compound stimuli are presented simultaneously, however, competitions may occur between different compound stimuli and between global and local processing of each compound stimulus. Given the possible differences between the processing of a single and multiple compound stimuli, the neural mechanisms differentiating global and local perception of multiple simultaneously presented compound stimuli are poorly understood. Evans et al. [2000] observed that the P1 evoked by bilateral compound stimuli was enhanced over the left hemisphere in the local relative to the global condition and over the right hemisphere in the global relative to the local condition. However, as target stimuli were always presented in one hemifield in one block of trials, the ERP results of Evans et al. [2000] might be contaminated by effects of spatial attention to target location. The neural mechanisms underpinning multiple simultaneously presented compound stimuli are not defined given the limitation of the paradigms used in previous studies.
Our recent ERP study investigated neural substrates that differentiate global and local processing of two compound letters that were presented simultaneously in the left (LVF) and right (RVF) visual fields, respectively [Jiang and Han, unpublished data]. We recorded ERPs from subjects who detected infrequent global or local targets randomly presented in one hemifield and accompanied with a distractor compound letter in the opposite hemifield. We found that attention to the local level of compound stimuli elicited a positivity over lateral occipital sites at 80–120 msec (P1) with a larger amplitude than those in the global attention condition whereas global attention produced an enhanced positivity at 240–320 msec (P2) over lateral occipital sites relative to local attention. Nevertheless, because of the low spatial resolution of ERP measurement, the ERP results could not identify exactly which brain areas are engaged in the global and local perception of bilateral inputs. In particular, it is unknown whether activities of the visual cortex or high‐level brain structures are involved to differentiate global/local processing of bilateral inputs. The current work extended our previous findings by recording hemodynamic responses using fMRI and similar stimuli and procedure. Brain areas preferentially engaged in the global processing of bilateral visual inputs were identified by the increased fMRI signals in the global than the local attention conditions. Similarly, the neural substrates for local processing were localized by examining the increased activations in the local than the global attention conditions. This experimental design allowed us to examine neural substrates that differentiate global and local processes of multiple compound stimuli presented simultaneously.
SUBJECTS AND METHODS
Participants
Ten graduate and undergraduate students (2 men, 8 women; mean age 21.2 years; range 20–24 years) from Peking University participated in the experiment as paid volunteers. All participants were right‐handed, had normal or corrected‐to‐normal vision, and gave informed consent according to the guidelines of Department of Psychology, Peking University.
Stimuli and Procedure
The stimuli were presented through an LCD projector onto a rear‐projection screen located at the subject's feet. The screen was viewed with an angled mirror positioned on the head‐coil. The stimuli were global letters made up of local letters in a 7 × 7 matrix, as illustrated in Figure 1. Three capital letters (A, E, and S) were used to produce compound letters, resulting in six combinations (i.e., global/local: A/E, A/S, E/A, E/S, S/A, S/E). All letters served as target and nontarget stimuli in separate epochs of trials. In each epoch of trials, a designated target appeared only at the global or local level of one of the two compound letters that were presented simultaneously in the LVF and RVF, respectively. For example, if global letter A was defined as a target, in the target present condition, stimuli A/E or A/S could appear randomly in the LVF or the RVF accompanied with distractors (e.g., global/local: E/A, E/S, S/A, or S/E) in the opposite hemifield. In the target absent condition, two distractors appeared simultaneously in the LVF and RVF, respectively. Both the global and local letters in the two hemifields were different on all trials. At a viewing distance of 270 cm, global letters were 3.0° wide and 4.3° high and local letters were 0.32° wide and 0.50° high.
Figure 1.
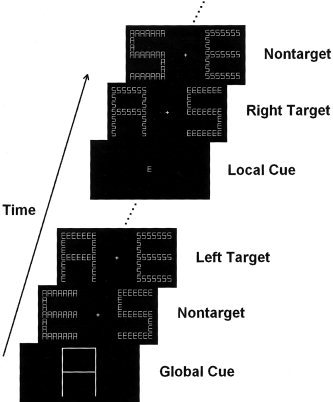
Illustrations of the stimuli and procedure used in the current study.
A fixation cross was continuously visible in the screen. Each trial began with the presentation of two compound letters simultaneously displayed to the left and right of the fixation, respectively. The distance between the fixation and inner edge of each compound stimulus was 2.5°. The stimulus displays were presented for 300 msec, and the interstimulus intervals (ISI) were randomized between 300–500 msec.
Subjects were asked to remain fixated on the fixation cross during the test. Each epoch of trials began with a large or a small letter made up of solid black lines, which was as large as the global or local letters of the compound stimuli and served as a cue to indicate target level (global or local) and target shape (A, E, or S). The cue remained on the screen for 1,000 msec. After 30 trials for practice, six scans of 64 sec were obtained from each subject. Each scan consisted of 2 epochs of 43 trials (30 sec for each epoch), alternating between global and local attention conditions. There were 14% target stimuli in each scan. Subjects were asked to respond to the global or local target letter by pressing a button with the right thumb. The order of global and local tasks was counterbalanced across subjects. Instructions emphasized both response speed and accuracy.
fMRI Image Acquisition and Analysis
Brain imaging was performed using a 1.5‐T GE Signa MR scanner with a custom head coil at Beijing Chaoyang Hospital. Fifteen axial slices of functional images that covered the whole cerebral cortex were acquired using echo‐planar imaging (64 × 64 × 15 matrix with 3.75 × 3.75 × 7‐mm spatial resolution, TR = 2,000 msec, TE = 40 msec, FOV = 240 mm, flip angle = 90°). Anatomical images were obtained with a standard 3‐D T1‐weighted sequence (resulting in a 256 × 256 × 84 matrix with 0.938 × 0.938 × 2.0‐mm spatial resolution, TR = 585 msec, TE = minimum). Subjects' heads were immobilized during the scanning sessions using pieces of foam.
SPM99 (the Wellcome Department of Cognitive Neurology, UK) was used for data processing and analysis. Following correction for differences in the timing of slice acquisition within a volume, the functional images were realigned to the first scan to correct for the head movement between scans. The structural image was coregistered with the mean functional image produced during the process of realignment. All images were normalized to a 2 × 2 × 2 mm3 Montreal Neurological Institute (MNI) template in Talairach space [Talairach and Tournoux, 1998] using bilinear interpolation. Functional images were spatially smoothed using a Gaussian filter with a full‐width at half maximum (FWHM) parameter set to 8 mm. The image data were modeled using a box‐car function. Contrasts were used to compare the effect of global/local processing. Regions preferentially engaged in global processing were defined as areas more activated by attention to global than local targets. Regions preferentially engaged in local processing were defined as areas more activated by attention to local than global targets. Random effect analyses were then conducted across the group of subjects based on statistical parameter maps from each individual subject to allow population inference. Areas of significant activation were identified at the cluster level for values exceeding an uncorrected P value of 0.001 for each individual subject and a corrected P value of 0.01 for the group analysis. The SPM coordinates for standard brain from Montreal Neurological Institute (MNI) were converted to Talaraich coordinates using a non‐linear transform method (available online at http://www.mrc-cbu.cam.ac.uk/Imaging/mnispace.html).
RESULTS
As our previous work showed that target locations (LVF vs. RVF) did not influence responses to global or local targets [Jiang and Han, unpublished data], the behavioral data corresponding to the LVF or RVF targets were collapsed. Paired t‐test showed that subjects hit more global than local targets (96.1 vs. 62.2%, t = 8.99, P < 0.001) and responses were faster to global than local targets (762 vs. 867 msec, t = 3.02, P < 0.014). False alarm rates were low and did not differ between global and local conditions (1.9 vs. 3.0%, t = 1.34, P > 0.2).
Brain areas preferentially activated by global and local attention are summarized in Table I. Relative to the local condition, global attention generated increased activations in the left middle and superior temporal cortices, corresponding to Brodmann areas 21 and 41 (Fig. 2a). Activations related to global attention were also observed in the right inferior temporal cortex, corresponding to Brodmann area 36 (Fig. 2b). Relative to the global condition, however, local attention elicited increased activations in the right and left superior parietal cortex, corresponding to Brodmann area 7 (Fig. 3).
Table I.
Brain areas activated by global/local processing of bilateral visual inputs*
| Condition/region | BA | x | y | z | Z value |
|---|---|---|---|---|---|
| Global | |||||
| Left middle temporal cortex | 21 | −44 | −2 | −18 | 4.71 |
| Left superior temporal cortex | 41 | −40 | −18 | 2 | 3.94 |
| 41 | −52 | −16 | 12 | 3.87 | |
| Right inferior temporal cortex | 36 | 32 | −24 | −14 | 3.99 |
| 36 | 24 | −40 | −8 | 3.34 | |
| 36 | 28 | −36 | −18 | 3.29 | |
| Local | |||||
| Left superior parietal cortex | 7 | −14 | −74 | 32 | 3.83 |
| 7 | −30 | −62 | 48 | 3.55 | |
| Right superior parietal cortex | 7 | 8 | −72 | 50 | 4.03 |
| 7 | 22 | −78 | 38 | 3.96 | |
| 7 | 24 | −64 | 58 | 4.04 |
Activations were reported if the significance was P < 0.01 corrected at cluster level. BA, Brodmann area.
Figure 2.
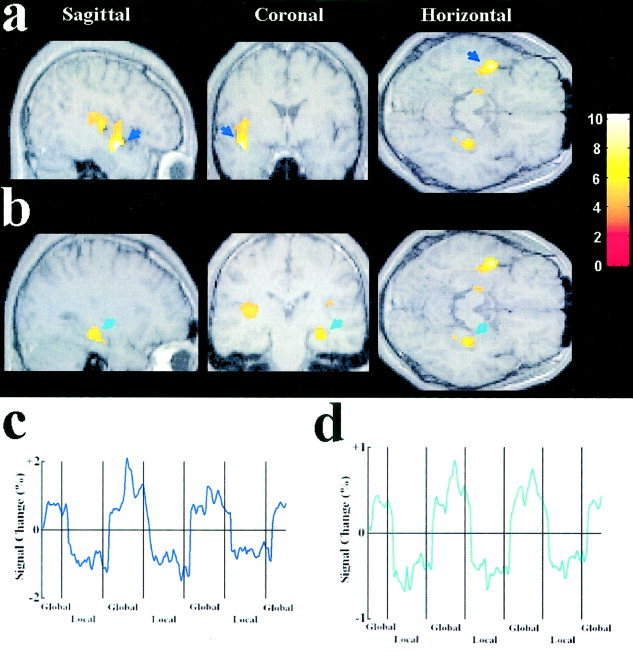
Brain areas activated by attention to the global level of bilateral compound stimuli. The results of the group analysis from 10 subjects were plotted on MR images of a representative subject. Threshold for activation of all clusters was P < 0.01 (corrected). The color bar indicates the scale of z values. a: Activations in the left temporal cortex (indicated by blue arrows). b: Activations in the right temporal cortex (indicated by green arrows). c: The time course of the signal change in the left superior temporal cortex as a function of global/local attention, averaged across the 10 subjects. The mean image values obtained from the average of the six scans were used as baseline. d: The time course of the signal change in the right inferior temporal cortex as a function of global/local attention, averaged across the 10 subjects. The x‐axis of the time course is for the whole session which contained 6 scans. Each local segment of the time course contains two 30‐sec epochs in two successive scans. The second and third global segments contain two 30‐sec epochs in two successive scans whereas the first and the fourth global segments contain one 30‐sec epoch in the first and the sixth scan, respectively. The time courses were averaged from raw fMRI signals.
Figure 3.
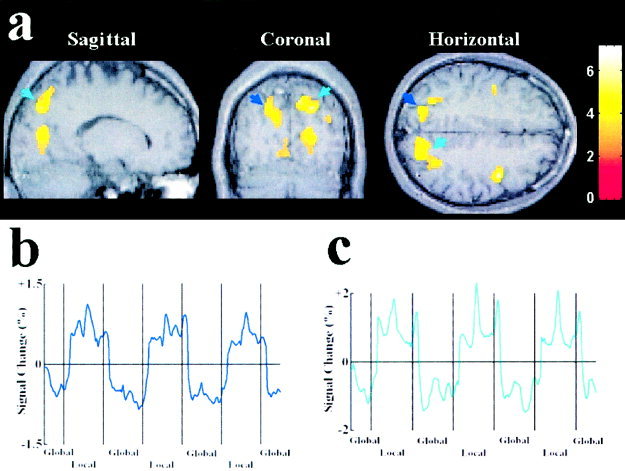
Brain areas activated by attention to the local level of bilateral compound stimuli. The results of the group analysis from 10 subjects were plotted on MR images of a representative subject. Threshold for activation of all clusters was P < 0.01 (corrected). The activations were observed in bilateral superior cortex. The color bar indicates the scale of z values. a: Activations in the left superior parietal cortex (indicated by blue arrows) and the right superior parietal cortex (indicated by green arrows). b: The time courses of the signal change in the left superior parietal cortex as a function of global/local attention, averaged across the 10 subjects. The mean image values obtained from the average of the six scans were used as baseline. c: The time courses of the signal change in the right superior parietal cortex as a function of global/local attention, averaged across the 10 subjects. The time courses were averaged from raw fMRI signals.
DISCUSSION
The objective of the present experiment was to identify neural substrates that are preferentially engaged in the global and local processing of two compound letters simultaneously presented in the two hemifields, respectively. The behavioral data showed a global RT advantage, consistent with previous studies in which compound stimuli were presented peripherally [Evans et al., 2000; Han et al., 1999a; Jiang and Han, unpublished data; Proverbio et al., 1998]. The fMRI data revealed separate brain systems that were involved in the attention to the global and local aspects of bilateral compound stimuli. Our data indicate that the left superior and middle temporal cortex and right inferior temporal cortex were activated when attention was directed to the global level of compound stimuli. When attention was focused on the local level, however, the left and right superior cortex was strongly activated. Because the compound stimuli presented in the global and local conditions were identical, the activations reflect per se the neural activities underlying global/local attention modulations rather than variation of stimulus attributes. Therefore, our current findings reinforce our previous ERP results that enhanced electrophysiological activities related to global/local processing of bilateral visual inputs had different time courses [Jiang and Han, unpublished data] and showed clear fMRI evidence for distinct neural substrates in the ventral and dorsal visual pathways related to the global and local processing of bilateral visual inputs, respectively.
The current fMRI results are different from those reported in previous research in which a single compound stimulus was displayed on a trial. For example, attention to the global level of compound stimuli displayed at fixation generated activations over the posterior occipital cortex [Fink et al., 1996; Han et al., 2002b; Sasaki et al., 2001] whereas no such activation was observed in the current experiment, indicating that neural mechanisms in the visual cortex did not play a critical role in differentiation of global/local perception of multiple compound stimuli. Moreover, previous work did not find activations in the temporal cortex associated with global attention but the current experiment did. The disparity between the experiments can not be simply attributed to the difference between foveal and peripheral vision because unilateral presentation of a single compound letter activated the posterior occipital cortex but not the temporal cortex in the global attention condition [Han et al., 2002b]. In addition, previous studies did not observe any activation related to local processing of unilateral compound letters [Han et al., 2002b] whereas the current work observed superior parietal activations associated with local attention. Thus, it appears that the neural substrates differentiating global/local processing are quite different between the conditions of bilateral presentation of two compound letters and unilateral presentation of a single compound letter. In particular, global/local processing of an individual compound stimulus can be differentiated as early as in the occipital visual cortex whereas higher order brain structures are involved in the differentiation of global/local processing of bilateral visual inputs.
Several mechanisms may contribute to the distinct patterns of brain activations associated with global and local processing of bilateral visual inputs. First, as researchers have demonstrated a close relationship between global/local processing and low/high spatial frequency (SFs) channels [Shulman et al., 1986; Shulman and Wilson, 1987], it has been suggested that selective attention to the global and local aspects of compound stimuli is mediated by selection of low and high SFs [Ivry and Robertson, 1998; Sergent, 1982]. Thus, it may be conjectured that the distinct activations in the temporal and parietal cortices observed here might reflect enhanced selection of low and high SFs, respectively. Nevertheless, recent ERP studies localized the early selection process of both high and low SFs in the dorsal extrastriate occipital areas whereas the late selection process of both high and low SFs in the ventral extrastriate cortex [Martinez et al., 2001]. The ERP study did not show any evidence for the involvement of the temporal and parietal cortex in the selection of low and high SFs, respectively. Furthermore, there has been no evidence that the contribution of SF selection to global/local processing varies as a function of the numbers of compound letters. Therefore, it is difficult to interpret the differential activations between the global and local conditions in the current experiment simply based on the SF account of global/local selection.
Second, our previous work identified that perceptual grouping of local elements is a key process that differentiates global from local processing [Han et al., 1999b; Han and Humphreys, 1999]. However, both ERP and fMRI studies found evidence for enhanced neural activities underlying perceptual grouping of local elements into global shapes in the posterior occipital cortex but not in the temporal cortex [Altmann et al., 2003; Han et al., 2001b; Han et al., 2002a; Kourtzi et al., 2003]. Thus, it is unlikely that the temporal activations observed here reflected the grouping operation in the global attention condition. Alternatively, as single unit studies have shown that neurons in the temporal cortex are sensitive to complex shapes and are organized into columns [for review, see Tanaka, 2003], it is possible that activities of a few columns of neurons are sufficient for identification local shapes. Global shape identification, however, may require coherent activities of multiple functional columns in the temporal cortex, which may be particularly important when multiple compound stimuli are fed into the visual system for recognition. This may account for the temporal activations associated with the global processing in the current experiment.
Finally, our previous work has shown that selection of a single local shape for identification is critical for local processing [Han and Humphreys, 2002; Han et al., 2001a; Han and He, 2003]. This selection process may become more important when the visual system is confronted with multiple, rather than one, global shapes that are made up of different local items. The increased activities in the superior parietal cortex in the local condition may reflect the enhanced local selection in the current work. This analysis is in agreement with previous fMRI results that the superior parietal cortex is selectively activated by orienting of spatial attention [Hopfinger et al., 2000]. The activations of the superior parietal cortex may also reflect stronger eye movement in the local than in the global conditions. However, activities associated with eye movement are distributed near intraparietal and postcentral sulcus as well as in the precentral gyrus [for review, see Corbetta, 1998]. These areas do not overlap with the parietal activations observed here. Moreover, our previous ERP studies did not show a significant difference in potentials indexing eye movement between global and local attention conditions [Jiang and Han, unpublished data]. Thus, it is unlikely that the superior parietal activities in the local condition reflect per se stronger eye movement in the local than global conditions.
In conclusion, the current experiment showed clear fMRI evidence that the ventral and dorsal visual pathways are engaged in the global and local processing of multiple compound stimuli, respectively. In comparison with the results of previous work, our findings indicate that the global/local processing of multiple compound stimuli may be essentially different from that of single compound stimulus that are presented either at fixation or in the periphery.
REFERENCES
- Altmann CF, Bülthoff HH, Kourtzi Z. (2003): Perceptual organization of local elements into global shapes in the human visual cortex. Curr Biol 13: 342–349. [DOI] [PubMed] [Google Scholar]
- Clark VP, Hillyard SA (1996): Spatial selective attention affects early extrastriate but not striate components of the visual evoked potential. J Cognit Neurosci 8: 387–402. [DOI] [PubMed] [Google Scholar]
- Corbetta M (1998): Frontaoparietal cortical networks for directing attention and the eye to visual locations: identifical, independent, or overlapping neural systems? Proc Natl Acad Sci USA 95: 831–838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delis D, Robertson LC, Efron R (1986): Hemispheric specialization of memory for visual hierarchical stimuli. Neuropsychologia 24: 205–214. [DOI] [PubMed] [Google Scholar]
- Evans MA, Shedden JM, Hevenor SJ, Hahn MC (2000): The effect of variability of unattended information on global and local processing: evidence for lateralization at early stages of processing. Neuropsychologia 38: 225–239. [DOI] [PubMed] [Google Scholar]
- Fink GR, Halligan PW, Marshall JC, Frith CD, Frackowiak RSJ, Dolan RJ (1996): Where in the brain does visual attention select the forest and the trees? Nature 382: 626–628. [DOI] [PubMed] [Google Scholar]
- Han S, He X (2003): Modulation of neural activities by enhanced local selection in the processing of compound stimuli. Hum Brain Mapp 19: 273–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han S, Humphreys GW (1999): Interactions between perceptual organization based on Gestalt laws and those based on hierarchical processing. Percept Psychophys 61: 1287–1298. [DOI] [PubMed] [Google Scholar]
- Han S, Humphreys GW (2002): Segmentation and selection contribute to local processing in hierarchical analysis. Q J Exp Psychol A 55: 5–21. [DOI] [PubMed] [Google Scholar]
- Han S, Fan S, Chen L, Zhuo Y (1997): On the different processing of wholes and parts: A psychophysiological study. J Cognit Neurosci 9: 686–697 [DOI] [PubMed] [Google Scholar]
- Han S, Fan S, Chen L, Zhuo Y (1999a): Modulation of brain activities by hierarchical processing: A high‐density ERP study. Brain Topogr 11: 171–183. [DOI] [PubMed] [Google Scholar]
- Han S, Humphreys GW, Chen L (1999b): Parallel and competitive processes in hierarchical analysis: perceptual grouping and encoding of closure. J Exp Psychol Hum Percept Perform 25: 1411–1432. [DOI] [PubMed] [Google Scholar]
- Han S, He X, Woods DL (2000): Hierarchical processing and level‐repetition effect as indexed by early brain potentials. Psychophysiology 37: 817–830. [PubMed] [Google Scholar]
- Han S, He X, Yund EW, Woods DL (2001a): Attentional selection in the processing of hierarchical patterns: An ERP study. Biol Psychol 56: 113–130. [DOI] [PubMed] [Google Scholar]
- Han S, Song Y, Ding Y, Yund EY, Woods DL (2001b): Neural substrates for visual perceptual grouping in human. Psychophysiology 38: 926–935. [DOI] [PubMed] [Google Scholar]
- Han S, Ding Y, Song Y (2002a): Neural mechanisms of perceptual grouping in humans as revealed by high density event related potentials. Neurosci Lett 319: 29–32. [DOI] [PubMed] [Google Scholar]
- Han S, Weaver J, Murray S, Yund EW, Woods DL (2002b): Hemispheric asymmetry in global/local processing: Effects of stimulus position and spatial frequency. NeuroImage 17: 1290–1299. [DOI] [PubMed] [Google Scholar]
- Heinze HJ, Münte TF (1993): Electrophysiological correlates of hierarchical stimulus processing: Dissociation between onset and later stages of global and local target processing. Neuropsychologia 31: 841–52. [DOI] [PubMed] [Google Scholar]
- Heinze HJ, Mangun GR, Burchert W, Hinrichs H, Scholz M, Munte TF, Gös A, Scherg M, Johannes S, Hundeshagen H, Gazzaniga MS, Hillyard SA. (1994): Combined spatial and temporal imaging of brain activity during visual selective attention in humans. Nature 372: 543–546. [DOI] [PubMed] [Google Scholar]
- Heinze HJ, Hinrichs H, Scholz M, Burchert W, Mangun GR (1998): Neural mechanisms of global and local processing. A combined PET and ERP study. J Cognit Neurosci 10: 485–498. [DOI] [PubMed] [Google Scholar]
- Hopfinger JB, Buonocore MH, Mangun GR (2000): The neural mechanisms of top‐down attentional control. Nature Neurosci 3: 284–291. [DOI] [PubMed] [Google Scholar]
- Ivry RB, Robertson LC (1999): Two sides of perception. Cambridge, MA: MIT Press. [Google Scholar]
- Jiang Y, Han S. Neural mechanisms underlying global/local processing of bilateral visual inputs (unpublished data).
- Kourtzi A, Tolias AS, Altmann CF, Augath M, Logothetis NK (2003): Integration of local features into global shapes: Monkey and human fMRI studies. Neuron 23: 333–346. [DOI] [PubMed] [Google Scholar]
- Lamb MR, Robertson LC, Knight RT (1990): Component mechanisms underlying the processing of hierarchically organized patterns: inferences from patients with unilateral cortical lesions. J Exp Psychol Learn Mem Cognit 16: 471–483. [DOI] [PubMed] [Google Scholar]
- Lamb MR, Robertson LC, Knight RT (1989): Attention and interference in the processing of global and local information: effects of unilateral temporal‐parietal junction lesions. Neuropsychologia 27: 471–483. [DOI] [PubMed] [Google Scholar]
- Martinez A, Moses P, Frank L, Buxton R, Wong E, Stiles J (1997): Hemispheric asymmetries in global and local processing: Evidence from fMRI. Neuroreport 8: 1685–1689. [DOI] [PubMed] [Google Scholar]
- Martinez A, Di Russo F, Anllo‐Vento L, Hillyard SA (2001): Electrophysiological analysis of cortical mechanisms of selective attention to high and low spatial frequencies. Clin Neurophysiol 112: 1980–1998 [DOI] [PubMed] [Google Scholar]
- Navon D (1977): Forest before trees: the precedence of global features in visual perception. Cognit Psychol 9: 353–383. [Google Scholar]
- Proverbio AM, Minniti A, Zani A (1998): Electrophysiological evidence of a perceptual precedence of global vs. local visual information. Cognit Brain Res 6: 321–334. [DOI] [PubMed] [Google Scholar]
- Robertson LC, Lamb MR, Knight RT (1988): Effects of lesions of temporal‐parietal junction on perceptual and attentional processing in humans. J Neurosci 8: 757–3769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson LC, Egly R, Lamb MR, Kerth L. (1993): Spatial attention and cuing to global and local levels of hierarchical structure. J Exp Psychol Hum Percept Perform 19: 471–487. [DOI] [PubMed] [Google Scholar]
- Sasaki Y, Hadjikhani N, Fischl B, Liu AK, Marret S, Dale AM, Tootell RB (2001): Local and global attention are mapped retinotopically in human occipital cortex. Proc Natl Acad Sci USA 98: 2077–2082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sergent J (1982): The cerebral balance of power: confrontation or cooperation? J Exp Psychol Hum Percep Perform 8: 253–272. [DOI] [PubMed] [Google Scholar]
- Shulman GL, Wilson J (1987): Spatial frequency and selective attention to local and global information. Perception 16: 89–101. [DOI] [PubMed] [Google Scholar]
- Shulman GL, Sullivan MA, Gish K, Sakoda WJ (1986): The role of spatial frequency channels in the perception of local and global structure. Perception 15: 259–279. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P (1998): Co‐planar stereotaxic atlas of the human brain. New York: Thieme. [Google Scholar]
- Tanaka K (2003): Columns for complex visual object features in the inferior temporal cortex: cluster of cells with similar but slightly different stimulus selectivities. Cereb Cortex 13: 90–99. [DOI] [PubMed] [Google Scholar]