

Abstract
Reading in a second language (L2) is a complex task that entails an interaction between L2 and the native language (L1). To study the underlying mechanisms, we used functional magnetic resonance imaging (fMRI) to visualize Chinese–English bilinguals' brain activity in phonological processing of logographic Chinese and alphabetic English, two written languages with a sharp contrast in phonology and orthography. In Experiment 1, we found that phonological processing of Chinese characters recruits a neural system involving left middle frontal and posterior parietal gyri, cortical regions that are known to contribute to spatial information representation, spatial working memory, and coordination of cognitive resources as a central executive system. We assume that the peak activation of this system is relevant to the unique feature of Chinese that a logographic character has a square configuration that maps onto a monosyllabic unit of speech. Equally important, when our bilingual subjects performed a phonological task on English words, this neural system was most active, whereas brain areas mediating English monolinguals' fine‐grained phonemic analysis, as demonstrated by Experiment 2, were only weakly activated. This suggests that our bilingual subjects were applying their L1 system to L2 reading and that the lack of letter‐to‐sound conversion rules in Chinese led Chinese readers to being less capable of processing English by recourse to an analytic reading system on which English monolinguals rely. Our brain imaging findings lend strongest support to the idea that language experience tunes the cortex. Hum. Brain Mapping 18:158–166, 2003. © 2003 Wiley‐Liss, Inc.
Keywords: fMRI, second language learning, bilingualism, reading in Chinese, neural bases for second language learning, brain mechanisms of Chinese reading
INTRODUCTION
For people who speak two languages, understanding tens of thousands of words in either of the two languages is an exceptional accomplishment of their cognitive and neural‐anatomical systems. Over the past half‐century, researchers have been attempting to elucidate the fundamental mechanisms that achieve this remarkable feat. Two prominent theories have been proposed. According to the universal language system theory, a general cognitive [Clarke, 1980; Goodman, 1973; Perfetti et al., 1992] and neuro‐anatomical network is engaged in reading across languages and the age that learners begin to acquire L2 does not matter [Chee et al., 1999; Hernandez et al., 2001; Klein et al., 1995]. Alternatively, it is hypothesized that reading involves language‐specific processing and representation systems [Fiez, 2000; Gottardo et al., 2001; Holm and Dodd, 1996; Hu and Catts, 1998; Neville et al., 1998; Siok and Fletcher, 2001; Vaid, 2002; Vaild and Hull, 2002; Wang and Geva, in press], especially for “late bilinguals” who begin to learn L2 in early adulthood [Kim et al., 1997; Weber‐Fox and Neville, 1996]. On this account, language similarity in orthography, phonology, and syntax determines the transfer of appropriate reading skills [Koda, 1994; Weinreich, 1953; Yip and Matthews, 2000].
To discover the cross‐linguistic nature of L2 literacy, we carried out one fMRI experiment with bilinguals who learned Chinese as L1 and English as L2. Chinese–English bilinguals were recruited because the two languages they have learned are most widely used in the world and possess the sharpest contrast in design principle. While English words map onto phonemes (minimal sound units as represented by letters) and abide by letter–sound conversion rules, written Chinese uses the character as the basic writing unit whose phonology is defined at the monosyllabic level, with no parts in the character corresponding to phonological segments such as phonemes. Although about 85% of present‐day Chinese characters are phonetic compounds containing a phonetic component that may give information about the pronunciation of the compound [Perfetti and Tan, 1999], estimates of the validity of this information reveal that only 38% of phonetic components are helpful for the pronunciation of whole characters. Moreover, it is never the case in Chinese that a phonetic component maps onto a subsyllabic phonological representation in the way that a letter maps onto a segment of a word's phonological form in an alphabetic system. For instance, in the English word beech, the b corresponds to /b/, and the latter is a segment of the word. For the Chinese phonetic compound 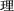 (pronounced /li3/, meaning reason; the numeral here refers to Chinese tone), the phonetic component
(pronounced /li3/, meaning reason; the numeral here refers to Chinese tone), the phonetic component  located on the right (also pronounced /li3/, meaning inside) does not correspond to a piece of the word's phonological form; it is the syllable that segmentally is exactly the word. There is no part within the written word that represents the segment /l/ or /i/ or the tone. Thus, Chinese writing does not allow a true segmental analysis that is fundamental to alphabetic systems [Mattingly, 1987], and quasi‐regular letter–sound conversion rules that exist in all alphabetic languages are impossible in Chinese. However, Chinese characters possess a number of strokes that are packed into a square shape; they map onto morphemes, often having their meaning suggested by visual configurations [Rayner and Pollatsek, 1989].
located on the right (also pronounced /li3/, meaning inside) does not correspond to a piece of the word's phonological form; it is the syllable that segmentally is exactly the word. There is no part within the written word that represents the segment /l/ or /i/ or the tone. Thus, Chinese writing does not allow a true segmental analysis that is fundamental to alphabetic systems [Mattingly, 1987], and quasi‐regular letter–sound conversion rules that exist in all alphabetic languages are impossible in Chinese. However, Chinese characters possess a number of strokes that are packed into a square shape; they map onto morphemes, often having their meaning suggested by visual configurations [Rayner and Pollatsek, 1989].
Two fMRI experiments are reported in this study. In Experiment 1, we utilized fMRI to localize the brain areas responsible for the phonological processing of the two languages by our bilingual subjects. Twelve right‐handed, neurologically normal male bilinguals who started to learn English after age 12 were scanned. Despite being “late” English learners, our assessment of their English proficiency indicated that they were moderately fluent L2 speakers. We used a rhyme judgment task [Booth et al., 2002; Lurito et al., 2000; Poldrack et al., 2001; Pugh et al., 1996; Xu et al., 2001], in which the subject decided whether a pair of synchronously exposed Chinese characters or English words rhymed with each other. Theoretically, rhyme judgments may involve phonological processes at several levels including syllable, rime, and phoneme [Poldrack et al., 1999; Zatorre et al., 1996]. For native English speakers, rhyme judgments of English words are assumed to provoke fine‐grained phonemic processes, because cognitive research has agreed that letter–sound conversions occur rapidly and obligatorily in identifying a written English word [Perfetti et al., 1988; Pollatsek et al., 1992; Van Orden, 1987]. In Chinese readers, rhyme judgments of Chinese characters may be more similar to syllable judgments, if subjects are not explicitly required to segment a syllable into subsyllabic units such as rime and onset. This is due to the dominance of syllable level processing over phonemic processing in reading Chinese. In the control scan, subjects decided whether two Chinese characters or English words had the same physical size (font size judgment). This task served as a baseline, which should control for activation due to the visuo‐orthographic and semantic processing of linguistic stimuli in the experimental task [Tan et al., 2001]. The two Chinese or English words exposed in each pair shared no visuo‐orthographic similarity so that subjects had to make a decision based on phonological rather than visual properties (Fig. 1).
Figure 1.

Examples of experimental stimuli used in study 1. a: The subject judged whether the two Chinese characters rhymed with each other. The character above the fixation crosshair is pronounced /nao/, with meanings of “noisy” and “stir up trouble.” The character below the fixation crosshair is pronounced /tao/, meaning “hitch up” and “sheath”. b: The subject judged whether the two Chinese characters had an identical physical size. The character above the fixation crosshair is pronounced /yin/, with a meaning of “because.” The character below the fixation crosshair is pronounced /lu/, meaning “road.” c,d: The subject also performed rhyme and font size judgments, respectively. With the above Chinese characters as examples, it can be seen that Chinese characters are composed of a number of strokes (the smallest writing unit of logographs), and are salient as holistic square‐shape units. Moreover, a character is pronounced monosyllabically, with no parts of the character corresponding to phonemes. For instance, in a, the character above the fixation crosshair is pronounced /nao/; but in this character, there is no component which is pronounced /n/ or /a/ or /ao/.
In order to compare results from our Chinese–English bilingual subjects directly with results from native English speakers, we carried out Experiment 2, wherein native English users were required to make the rhyme decision on the same set of English words used by Chinese subjects. The same MRI scanner was used.
SUBJECTS AND METHODS
Subjects
In Experiment 1, we scanned 12 male bilingual subjects ranging in age from 29 to 39 years. They gave informed consent in accordance with guidelines set by the University of Texas Health Science Center at San Antonio (UTHSCSA). All were native Chinese speakers from China who started to learn English as L2 after age 12. They were graduate students who were studying in a doctoral program at University of Texas.
All subjects were strongly right handed as judged by the handedness inventory devised by Snyder and Harris [ 1993]. In this inventory, we adopted nine items involving unimanual tasks (tasks that can be done by only one hand). A 5‐point Likert‐type scale was used, with 1 representing exclusive left‐hand use and 5 representing exclusive right‐hand use. The items were: writing a letter, drawing a picture, throwing a ball, holding chopsticks, hammering a nail, brushing teeth, cutting with scissors, striking a match, and opening a door. The scores on the 9 items were summed for each subject, with the lowest score (9) indicating exclusive left‐hand use for all tasks, and the highest score (45) indicating exclusive right‐hand use. All subjects had scores higher than 40.
A language experience questionnaire was devised to obtain measures of self‐reported current fluency in English for the bilingual subjects. On average, subjects began speaking their second language at 12 years and already received a minimum of 12 years of formal training in English throughout primary school, high school, and college in China. They all had a TOEFL score of 570 or higher at the time of admission to a graduate program. Their length of residence in the United States varied from 3 to 7 years. The questionnaire also contained a rating scale to assess subjects' language skills in speaking, reading, and understanding. The endpoints of the rating scale were 1 (not fluent) and 5 (very fluent). The average rating scores of fluency in subjects' English were 4.42 (speaking, SD = 0.669), 4.67 (reading, SD = 0.492), and 4.33 (understanding, SD = 0.651).
In Experiment 2, we scanned 12 male native English speakers ranging in age from 25 to 38. None of the native English speakers ever learned the Chinese language. They were strongly right handed as judged by the handedness inventory. They also gave informed consent in accordance with guidelines set by UTHSCSA.
Apparatus and Procedure
Experiments were performed on a 1.9 T GE/Elscint Prestige whole‐body MRI scanner (GE/Elscint Ltd., Haifa, Israel) at the Research Imaging Center at UTHSCSA. The subject's head was immobilized by a tightly fitting thermally molded plastic facial mask that extended from hairline to chin [Fox et al., 1985]. A T2*‐weighted gradient‐echo echo planar imaging (EPI) sequence was used for fMRI scans, with the slice thickness = 6 mm, in‐plane resolution = 2.9 mm × 2.9 mm, and TR/TE/θ = 2,000 msec/45 msec/90 degrees. The field of view was 372 × 210 mm, and the acquisition matrix was 128 × 72. Twenty contiguous axial slices were acquired to cover the whole brain. The anatomical MRI was acquired using a T1‐weighted, three‐dimensional, gradient‐echo pulse‐sequence. This sequence provided high resolution (1 × 1 × 1 mm) images of the entire brain.
Materials and Behavioral Performance
Twenty‐four pairs of rhyming Chinese characters and 24 pairs of rhyming English words were used for rhyme judgments. Another 24 pairs of Chinese characters and 24 pairs of English words were used for font size judgments. To minimize the visual similarity of stimuli, the two Chinese characters in each pair shared no components, while the two English words in each pair shared as few letters as possible. Both the Chinese characters and English words were commonly used in terms of their frequency (mean = 159 per million and SD = 276 for Chinese; mean = 184 and SD = 283 for English stimuli).
The stimuli were shown through a LED projector system. In the rhyme decision task, subjects judged whether the two viewed characters or English words rhymed with each other. In the font size decision task, subjects decided whether the two viewed items had a same physical size. In each trial, a pair of characters was exposed synchronously for 1,500 msec, one above and one below a fixation crosshair. After the presentation of the two items, a fixation crosshair was exposed for 500 msec. Subjects indicated a positive response by pressing the key corresponding to the index finger of their right hand and a negative response by pressing the key corresponding to the index finger of their left hand. Chinese (or English) items were randomized within 24‐sec blocks comprised of 6 pairs of rhyming items and 6 pairs of unrelated stimuli that served as fillers. In the control scan, 6 pairs of items had the same physical size and another 6 pairs a different physical size. Experiment 1 was conducted in a single run, consisting of 4 blocks for each of the 4 conditions (i.e., Chinese and English rhyme decision, and Chinese and English font size decision). Presentation of the four conditions was counterbalanced and randomized across subjects.
In Experiment 2, 12 native English speakers performed the rhyme and font‐size decision tasks on the same English words that were used for our bilingual subjects. The design and procedures were identical to those of Experiment 1, except that the native English speakers did not view Chinese stimuli.
Data Analysis
Matlab (Mathworks, Natick, MA) and in‐house software were used for image processing [Xiong et al., 1995], which included corrections for head motion and global MRI signal shift. Skull stripping of the 3D MRI T1‐weighted images was done using Alice software (Perceptive Systems, Boulder, CO). These images were then spatially normalized to the Talairach brain atlas [Talairach and Tournoux, 1988] using the Convex Hull algorithm [Lancaster et al., 1999].
Functional images were grouped into Chinese rhyme decision, Chinese font size decision, English rhyme decision, and English font size decision. (For native English speakers, functional images were grouped into English rhyme decision and English font size decision.) Images from the first 8 sec of each condition were excluded from further functional data processing to minimize the transit effects of hemodynamic responses. Activation maps were calculated by comparing images acquired during the rhyme task state with those acquired during the control state (font size decision), using a students' group t‐test. To suppress noise, fMRI images were filtered using a spatial Gaussian filter with full width at half maximum (FWHM) of three pixels. The activation maps were also spatially normalized into Talairach space using the Convex Hull algorithm. The averaged activation maps across the subjects with a t value threshold of 2.4 (P < 0.01) were then overlaid on the corresponding T1 images. For each condition, Talairach coordinates of the center‐of‐mass and volume (mm3) of the activation clusters were determined based on the averaged activation maps. A commonality analysis [Roskies et al., 2001] was performed to identify regions common to Chinese and English rhyme decisions for our bilinguals. Direct comparisons of activated brain regions in terms of activation volume and activation index [Gao et al., 1996] were conducted to see the difference between Chinese and English readers.
RESULTS
For Chinese–English bilingual subjects, activations related to rhyme decision contrasted with font size decision were remarkably similar for Chinese characters and English words (see Fig. 2). Peak activations occurred in the left middle frontal cortex at Brodmann Areas (BAs) 9 and 46. The left motor cortex and supplementary motor cortex were also strongly active. Other important activated areas included precuneus (BA 7) and inferior parietal lobule (BA 40) in the left hemisphere. Minor activations were seen in the left inferior prefrontal gyrus (BA 45/47). The commonality analysis identified the left middle frontal cortex and precuneus to be significant both for Chinese characters and English words. There were no differential activations between Chinese and English stimuli.
Figure 2.
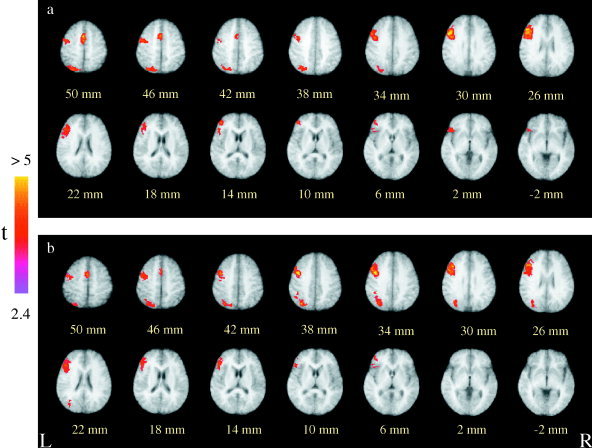
Averaged brain activations provoked by phonological analysis (rhyme decisions minus font size decisions) of Chinese characters (a) and English words (b) for late Chinese–English bilinguals. Normalized t‐maps (in color) pooled from 12 subjects are overlaid on the corresponding T1 images (in gray scale), demonstrating statistically significant activations (P < 0.01). Planes are axial sections, labeled with the height (mm) relative to the bicommissural line. a: Activations in reading Chinese peaked at the left middle frontal cortex (x = −44, y = 21, z = 26, t = 6.43) and also can be seen in the left precuneus (x = −28, y = −72, z = 43, t = 2.97), the left inferior parietal lobule (x = −45, y = −55, z = 54, t = 2.67) and supplementary motor area (SMA, x = −2, y = 4, z = 51, t = 3.39). b: Peak activation in reading English was seen in the left middle frontal cortex (x = −44, y = 18, z = 31, t = 6.28). Strong brain activations were also observed in the left precuneus (x = −31, y = −69, z = 43, t = 4.42), the left inferior parietal cortex (x = −43, y = −42, z = 36, t = 2.72) and SMA (x = −1, y = 4, z = 52, t = 3.27). According to a commonality analysis, activations that are common to Chinese and English phonological analyses were housed in the left middle frontal region (x = − 44, y = 20, z = 29) and precuneus (x = −29, y = −69, z = 44).
For native English speakers, as shown in Figure 3, phonological processing of English monolinguals was mediated by the peak activations of the left inferior frontal (BA 44/45) and superior temporal gyri (BA 22). Activation in the middle frontal cortex was weaker. This pattern of brain activations is in striking contrast to the neural networks responsible for reading Chinese and English by our bilingual subjects.
Figure 3.
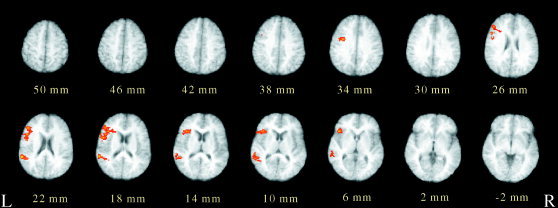
Averaged brain activations associated with phonological analysis (rhyme decisions minus font size decisions) for native English users. Normalized t maps (color) pooled from 12 subjects are overlaid on the corresponding T1 images (gray‐scale), demonstrating statistically significant activations (P < 0.01). Planes are axial sections, labeled with the height (mm) relative to the bicommissural line. Peak activations can be seen in the inferior prefrontal cortex (x = −49, y = 11, z = 20, t = 6.27; and x = −35, y = 20, z = 12, t = 4.61) and the superior temporal gyri (x = −57, y = −42, z = 21, t = 6.21) on the left. Weak brain activity was observed in the left middle frontal cortex (x = −49, y = 38, z = 24, t = 4.59).
Direct comparisons of the cortical activations (in volume) between Chinese and English readers further qualified the cultural difference (Fig. 4). Greater activations were obtained in the left middle frontal cortex for Chinese and English rhyme decisions in Chinese readers as compared with the rhyme decisions of native English readers. In contrast, native English readers showed greater activations in the left superior temporal gyrus and inferior frontal cortex than Chinese readers. Direct comparisons employing activation index, which uses activation volume × the fMRI signal changes averaged over the activation volume [Gao et al., 1996], showed the same pattern.
Figure 4.
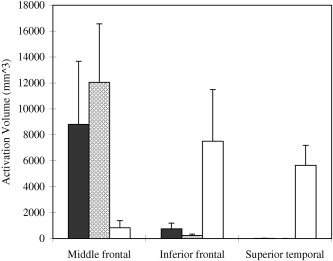
Direct comparison of activation volume (mm3) between native Chinese and English readers. In the left middle frontal cortex covering BAs 9 and 46, greater activation was found in the phonological processing of Chinese and English words by Chinese readers compared with native English readers (t(22) = 2.81, P < 0.01, and t(22) = 4.28, P < 0.0001, respectively). In the left superior temporal cortex, however, significantly stronger brain activity was seen for native English speakers than for Chinese (t(22) = 6.28, P < 0.0001 compared to Chinese readers' processing of Chinese characters, and t(22) = 6.29, P < 0.0001, compared to Chinese readers' processing of English words). Similarly, native English readers also showed greater activation in the left inferior prefrontal gyrus (t(22) = 2.92, P < 0.008 compared to Chinese readers' processing of Chinese characters, and t(22) = 3.17, P < 0.004, compared to Chinese readers' processing of English words). Activation difference between phonological processes of Chinese and English for our bilingual subjects was not statistically significant in the aforementioned brain regions. No other cortical areas showed a significant difference between Chinese and English readers. Black bars, Chinese rhyme decision; hatched bars, English rhyme decision by Chinese; open bars, English rhyme decision by native readers.
DISCUSSION
Previous brain mapping research has commonly implicated left mid‐superior temporal gyri [Booth et al., 2002; Petersen et al., 1988; Simos et al., 2000; Xu et al., 2001] and left posterior portion of inferior frontal cortex [Bookheimer et al., 1995; Fiez et al., 1999; Fiez and Petersen, 1998; Poldrack et al., 2001; Xu et al., 2001] in phonological processing of alphabetic words by English monolinguals, with left mid‐superior temporal regions more specifically identified as being responsible for fine‐grained phonemic analysis [Simos et al., 2000, 2002]. The results of our Experiment 2 with native English speakers provide a perfect replication of these findings. Our data revealed that phonemic processing of English words as mediated by letter‐to‐sound conversions occurs obligatorily and may dominate over syllabic level phonological processing in native speakers.
Our results with Chinese logographs suggest an important segregation of cortical representations of phonological information for Chinese and native English speakers. Phonological analysis in Chinese peaked in left middle frontal cortex, with a much weaker involvement of inferior frontal gyri. As left middle lateral frontal cortex is known to mediate spatial information storage and spatial working memory as a central executive system responsible for coordination of cognitive resources [e.g., Courtney et al., 1998; D'Esposito et al., 1995; Petrides et al., 1993; Wilson et al., 1993], we assume that the extremely strong activation seen in these areas in reading Chinese is associated with the coordination of the phonological processing of the Chinese logographs that was explicitly required by the experimental task and the elaborated analyses of the visual–spatial locations of the strokes in a logograph that was demanded by its unique square configuration. In other words, the cortical organization of phonological knowledge of written words reflects associations with visual features and associations with linguistic attributes (e.g., orthographic). This hypothesis of phonological organization is parallel to the sensory‐motor hypothesis of semantic systems [Allport, 1985; Chao et al., 1999; Damasio et al., 1996; Martin et al., 1996; see Bookheimer, 2002, for review]. Previous imaging data from semantic judgments and word generation of logographs also implicated these regions in coordination and integration of linguistic and visuo‐spatial analyses [Kuo et al., 2001; Tan et al., 2000, 2001] though these areas are not activated in passive viewing tasks [Chen et al., 2002; Fu et al., 2002].
The inactivity of left mid‐superior temporal cortex in phonological processing of Chinese is attributable to the monosyllabic nature of written characters that does not call for automatic phonemic analysis. Our present finding that phonological analysis was subserved by left inferior parietal cortex corroborated the role of visuo‐spatial processing in reading Chinese.
Equally important is our discovery of the high similarity of neural networks involved in phonological analysis of L1 and L2 words. One might expect that, as English words are linearly constructed and follow the letter‐to‐sound conversion rules that require elaborated phonemic processing, the left mid‐superior temporal gyrus would be active. Indeed, our second fMRI experiment with native English users did find the peak activity of left inferior frontal and superior temporal gyri in phonological processing. Our failure to show activation in the mid‐superior temporal gyrus when Chinese–English bilinguals processed English words phonologically is thus not due to inadequate power to detect the activation. Rather, it suggests that the processing of L1 phonology (where logographic characters are pronounced monosyllabically) carries over to L2 processing. Clearly, our Chinese subjects were applying the strategy of processing Chinese to processing English words. They did not automatically use the letter‐to‐sound conversion rules to pronounce English words. Collectively, our current fMRI experiments have produced the most compelling data in support of the hypothesis that language experience tunes the cortex [Fiez, 2000; Neville et al., 1998].
In conclusion, the nature of the Chinese writing system that logographs do not map onto phonemes is supposed to be responsible for the indolence of the letter–sound conversion route in Chinese–English bilinguals' computational processing of phonological information of English words. Although Chinese readers learn pinyin (an alphabetic system that is used to aid the learning of character pronunciation) for two months prior to learning characters in primary school in China, their exposure to the pinyin system is short and does not help the development of phonemic processing of English words [Siok and Fletcher, 2001]. Our brain mapping studies lend strong support to the prominent theory that reading involves language‐specific neurocognitive systems in which L2 reading is shaped by L1 for bilinguals.
REFERENCES
- Allport DA (1985): Distributed memory, modular subsystems and dysphasia In: Newman SK, Epstein E, editors. Current perspectives in dysphasia. Edinburgh: Churchill Livingstone; p 207–244. [Google Scholar]
- Bookheimer SY (2002): Functional MRI of language: new approaches to understanding the cortical organization of semantic processing. Annu Rev Neurosci 25: 151–188. [DOI] [PubMed] [Google Scholar]
- Bookheimer SY, Zeffiro T, Blaxton T, Gaillard W, Theodore W (1995): Regional cerebral blood flow during object naming and word reading. Hum Brain Mapp 3: 93–106. [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM (2002): Functional anatomy of intra‐and cross‐modal lexical tasks. Neuroimage 16: 7–22. [DOI] [PubMed] [Google Scholar]
- Chao LL, Haxby JV, Martin A (1999): Attribute‐based neural substrates in temporal cortex for perceiving and knowing about objects. Nat Neurosci 2: 913–919. [DOI] [PubMed] [Google Scholar]
- Chee M, Tan E, Thiel T (1999): Mandarin and English single word processing studied with functional magnetic resonance imaging. J Neurosci 19: 3050–3056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Fu S, Iversen SD, Smith SM, Matthews PM (2002): Testing for dual brain processing routes in reading: a direct contrast of Chinese character and pinyin reading using fMRI. J Cogn Neurosci 14: 1088–1098. [DOI] [PubMed] [Google Scholar]
- Clarke MA (1980): The short circuit hypothesis of ESL reading or when language competence interferes with reading performance. Mod Lang J 64: 203–209. [Google Scholar]
- Courtney SM, Petit L, Maisog JM, Ungerleider LG, Haxby JV (1998): An area specialized for spatial working memory in human frontal cortex. Science 279: 1347–1351. [DOI] [PubMed] [Google Scholar]
- D'Esposito M, Detre JA, Alsop DC, Shin RK (1995): The neural basis of the central executive system of working memory. Nature 378: 279–281. [DOI] [PubMed] [Google Scholar]
- Damasio H, Grabowski TJ, Tranel D, Hichwa RD, Antonio R (1996): A neural basis for lexical retrieval. Nature 380: 499–505. [DOI] [PubMed] [Google Scholar]
- Fiez JA (2000): Sound and meaning: how native language affects reading strategies. Nature Neurosci 3: 3–5. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Balota, DA , Raichle ME, Petersen SE (1999): Effects of lexicality, frequency, and spelling‐to‐sound consistency on the functional anatomy of reading. Neuron 24: 205–218. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Petersen SE (1998): Neuroimaging studies of word reading. Proc Natl Acad Sci USA 95: 914–921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox PT, Perlmutter JS, Raichle ME (1985): A stereotactic method of anatomical localization for positron emission tomography. J Comp Assist Tomogr 9: 141–153. [DOI] [PubMed] [Google Scholar]
- Gao JH, Parsons LM, Bower JM, Xiong J, Li J, Fox PT (1996): Cerebellum implicated in sensory acquisition and discrimination rather than motor control. Science 272: 545–547. [DOI] [PubMed] [Google Scholar]
- Goodman KS (1973): Psycholinguistic universals in the reading process In: Smith F. editor. Psycholinguistics and reading. New York: Holt, Rinehart and Winston, p 21–27. [Google Scholar]
- Gottardo A, Yan B, Siegel LS, Wade WL (2001): Factors related to English reading performance in children with Chinese as a first language: More evidence of cross‐language transfer of phonological processing. J Educ Psychol 93: 530–542. [Google Scholar]
- Hernandez AE, Dapretto M, Mazziotta J, Bookheimer S (2001): Language switching and language representation in Spanish–English bilinguals: an fMRI study. NeuroImage 14: 510–520. [DOI] [PubMed] [Google Scholar]
- Holm A, Dodd B (1996): The effect of first written language on the acquisition of English literacy. Cognition 59: 119–147. [DOI] [PubMed] [Google Scholar]
- Hu CF, Catts HW (1998): The role of phonological processing in early reading ability: what we can learn from Chinese. Scientific Studies of Reading 2: 55–79. [Google Scholar]
- Kim KS, Relkin NR, Lee KM, Hirsch J (1997): Distinct cortical areas associated with native and second languages. Nature 388: 171–174. [DOI] [PubMed] [Google Scholar]
- Klein D, Milner B, Zatorre RJ, Meyer E, Evans AC (1995): The neural substrates underlying word generation: a bilingual functional‐imaging study. Proc Natl Acad Sci USA 92: 2899–2903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koda K (1994): Second language reading research: problems and possibilities. Appl Psycholinguist 15: 1–28. [Google Scholar]
- Lancaster JL, Fox PT, Downs H, Nickerson DS, Hander TA, Mallah ME, Kochunov PV, Zamarripa K (1999): Global spatial normalization of human brain using convex halls. J Nucl Med 40: 942–955. [PubMed] [Google Scholar]
- Lurito JT, Kareken DA, Lowe MJ, Chen SA, Matthews VP (2000): Comparison of rhyming and word generation with FMRI. Hum Brain Mapp 10: 99–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin A, Wiggs CL, Ungerleider LG, Haxby JV (1996): Neural correlates of category‐specific knowledge. Nature 379: 649–652. [DOI] [PubMed] [Google Scholar]
- Mattingly IG (1987): Morphological structure and segmental awareness. Curr Psychol Cogn 7: 488–93. [Google Scholar]
- Neville HJ, Bavelier D, Corina D, Rauschecker J, Karni A, Lalwani A, Braun A, Clark V, Jezzard P, Turner R (1998): Cerebral organization for language in deaf and hearing subjects: biological constraints and effects of experience. Proc Natl Acad Sci USA 95: 922–929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perfetti CA, Tan LH (1999): The constituency model of Chinese word identification In: Wang J, Inhoff AW. editors. Reading Chinese script: a cognitive analysis. Mahwah, NJ: Erlbaum; p 115–134. [Google Scholar]
- Perfetti CA, Bell L, Delaney S (1988): Automatic phonetic activation in silent word reading: evidence from backward masking. J Mem Lang 27: 59–70. [Google Scholar]
- Perfetti CA, Zhang S, Berent I (1992): Reading in English and Chinese: evidence for a “universal” phonological principle In: Frost R, Katz L, editors. Orthography, phonology, morphology, and meaning. Amsterdam: North‐Holland; p 227–248. [Google Scholar]
- Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME (1988): Positron emission tomographic studies of cortical anatomy of single‐word processing. Nature 331: 585–589. [DOI] [PubMed] [Google Scholar]
- Petrides M., Alivasatos B., Meyer E, and Evans AC (1993): Functional activation of the human frontal cortex during the performance of verbal working memory tasks. Proc Natl Acad Sci USA 90: 878–882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poldrack RA, Wagner AD, Prull MW, Desmond JE, Glover GH, Gabrieli JD (1999): Functional Specialization for Semantic and Phonological Processing in the Left Inferior Prefrontal Cortex. Neuroimage 10: 15–35. [DOI] [PubMed] [Google Scholar]
- Poldrack RA, Temple E, Proropapas A, Nagarajan S, Tallal P, Merzenich M, Gabrieli JD (2001): Relations between the neural bases of dynamic auditoryprocessing and phonological processing: evidence from fMRI. J Cogn Neurosci 3: 687–697. [DOI] [PubMed] [Google Scholar]
- Pollatsek A, Lesch M, Morris RK, Rayner K (1992): Phonological codes are used in integrating information across saccades in word identification and reading. J Exp Psychol Human 18: 148–162. [DOI] [PubMed] [Google Scholar]
- Pugh KR, Shaywitz BA, Shaywitz SE, Constable RT, Skudlarski P, Fulbright RK, Bronen RA, Shankweiler DP, Katz L, Fletcher JM, Gore JC (1996): Cerebral organization of component processes in reading. Brain 119: 1221–1238. [DOI] [PubMed] [Google Scholar]
- Rayner K, Pollatsek A (1989): The psychology of reading. Englewood Cliffs, NJ: Prentice‐Hall. [Google Scholar]
- Roskies AL, Fiez JA, Balota DA, Raichle ME, Petersen SE (2001): Task‐dependent modulation of regions in the left inferior frontal cortex during semantic processing. J Cogn Neurosci 13: 829–843. [DOI] [PubMed] [Google Scholar]
- Simos PG, Breier JI, Wheless JW, Maggio WW, Fletcher JM, Castillo EM, Papanicolaou AC (2000): Brain mechanisms for reading: the role of the superior temporal gyrus in word and pseudowords naming. Neuroreport 11: 2443–2447. [DOI] [PubMed] [Google Scholar]
- Simos PG, Breier JI, Fletcher JM, Foorman BR, Castillo EM, Andrew C, Papanicolaou AC (2002): Brain mechanisms for reading words and pseudowords: an integrated approach. Cereb Cortex 12: 297–305. [DOI] [PubMed] [Google Scholar]
- Siok WT, Fletcher P (2001): The role of phonological awareness and visual‐orthographic skills in Chinese reading acquisition. Dev Psychol 37: 886–899 [PubMed] [Google Scholar]
- Snyder PJ, Harris LJ (1993): Handedness, sex, and familial sinistrality effects on spatial tasks. Cortex 29: 115–134. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P (1988): Co‐planar stereotactic atlas of the human brain. New York: Theime Medical Publishers, Inc; 122 p. [Google Scholar]
- Tan LH, Spinks JA, Gao JH, Liu A, Perfetti CA, Xiong J, Pu Y, Liu Y, Stofer KA, Fox PT (2000): Brain activation in the processing of Chinese characters and words: A functional MRI study. Hum Brain Mapp 10: 16–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH, Liu HL, Perfetti CA, Spinks JA, Fox PT, Gao JH (2001): The neural system underlying Chinese logograph reading. Neuroimage 13: 836–846. [DOI] [PubMed] [Google Scholar]
- Van Orden GC (1987): A ROWS is a ROSE: Spelling, sound and reading. Mem Cog 15: 181–198. [DOI] [PubMed] [Google Scholar]
- Vaid J (2002): Bilingualism In: Encyclopedia of the human brain. New York: Elsevier Science. [Google Scholar]
- Vaid J, Hull R (2002): Re‐envisioning the bilingual brain using functional neuroimaging: Methodological and interpretive issues In: Fabbro F, ed. Advances in the neurolinguistics of bilingualism. Udine: Forum, p 315–355. [Google Scholar]
- Wang M, Geva E. (in press). Spelling performance of Chinese ESL children: Lexical and visual‐orthographic processes. Appl Psycholing [Google Scholar]
- Weber‐Fox C, Neville HJ (1996): Maturational constraints on functional specializations for language processing: ERP and behavioral evidence in bilingual speakers. J Cogn Neurosci 8: 231–256. [DOI] [PubMed] [Google Scholar]
- Weinreich U (1953): Languages in contact: findings and problems. London: Mouton; 148 p. [Google Scholar]
- Wilson FA, Scalaidhe SP, Goldman‐Rakic PS (1993): Dissociation of object and spatial processing domains in primate prefrontal cortex. Science 260: 1955–1958. [DOI] [PubMed] [Google Scholar]
- Xiong J, Gao JH, Lancaster JL, Fox PT (1995): Clustered pixels analysis for functional MRI activation studies in the human brain. Hum Brain Mapp 3: 209–223. [Google Scholar]
- Xu B, Grafman J, Gaillard WD, Ishii K, Vega‐Bermudez F, Pietrini P, Reeves‐Tyer P, DiCamillo P, Theodore W (2001): Conjoint and extended neural networks for the computation of speech codes: the neural basis of selective impairment in reading words and pseudowords. Cereb Cortex 11: 267–277. [DOI] [PubMed] [Google Scholar]
- Yip V, Matthews S (2000): Syntactic transfer in a Cantonese‐English bilingual child. Bilingualism Lang Cogn 3: 193–208. [Google Scholar]
- Zatorre RJ, Meyer E, Gjedde A, Evans AC (1996): PET studies of phonetic processing of speech: review, replication, and reanalysis. Cereb Cortex 6: 21–30. [DOI] [PubMed] [Google Scholar]