

Abstract
Event‐related brain potentials were recorded to study whether verbs and nouns activate topographically distinct cortical generators. Fifteen subjects performed a primed lexical decision task with verb/verb and noun/noun pairs. The relatedness between prime and target items was varied in three steps (unrelated, moderately, and strongly related) and the EEG was recorded from 124 scalp electrodes. The topography of cortical sources of the N400 effect was evaluated by standardized differences scores and by cortical current source estimates which were constrained by the individual MRI‐determined cortex anatomy. A behavioral priming effect and a substantial N400 effect was found for both word categories. However, the topography of the grand average N400 effect of verbs and nouns did not differ, neither for raw nor for standardized amplitudes. Cortical current source estimates of the N400 effect revealed a very broad and scattered distribution of active locations with pronounced interindividual differences. Cortical current source estimates obtained with the L1‐norm and L2‐norm model, respectively, differed in the distribution of sources over the cortex but converged on the same “hot spots.” The data give no indication that the N400 effect is generated by word category‐specific networks which have a different topography. The marked individual differences are discussed with respect to the involved processes and the current source estimation procedures. Hum. Brain Mapping 11:178–192, 2000. © 2000 Wiley‐Liss, Inc.
Keywords: ERP, N400, cortical current source estimation, verbs and nouns, word processing, topography
INTRODUCTION
Neuropsychological findings provide evidence that nouns and verbs are processed in specialized and distinct cortical networks [Caramazza and Hillis, 1991; Damasio and Tranel, 1993; Daniele et al., 1994]. Damasio and Tranel [1993], for example, reported data of three patients: Two had deficits in object naming, i.e. in a noun production task but were unimpaired in verb production, while the third patient showed the reversed pattern of deficits, i.e. correct noun production but impaired verb retrieval. Based on this double dissociation the authors suggested a preponderance of left frontal areas for verb processing and of left temporal areas for noun processing. Converging evidence for a dissociation of verb and noun processing also comes from brain imaging studies. For example, Fiez et al. [1996] studied activation patterns within the left temporal cortex during language tasks and reported a higher activation of midtemporal areas during verb generation and a higher activation of temperoparietal areas during passive listening of content words belonging to different syntactical categories, i.e. also an anterior‐posterior dissociation between verb and nonverb processing. A similar dissociation for action and color words (i.e. verbs vs. adjectives) was reported by Martin et al. [1995]. Further, domain‐specific lexical regions have been described, among others, by Spitzer et al. [1998] for different semantic categories (animals, furniture) using functional magnetic resonance imaging in a naming task and by Beauregard et al. [1997] for abstract, concrete, and emotional words on the basis of a PET study.
Attempts have also been made to prove a word‐category‐specific dissociation of noun and verb processing by means of EEG recordings. Preissl et al. [1995] recorded event‐related potentials of the EEG while subjects performed a lexical decision task, i.e. they had to distinguish verbs and nouns from pseudowords. The authors reported a significant interaction of ANOVA factors “word class” and “electrode position,” and demonstrated in a difference map that nouns showed a stronger positivity than verbs around 200 ms after stimulus onset over fronto‐central areas. They concluded that ”nouns and verbs are processed differently in the intact brain during lexical decision tasks.“ This conclusion, however, is at variance with the findings of Gomes et al. [1997]. These authors presented noun/noun or noun/verb pairs in a lexical decision task. The two words of a pair could either be semantically related or unrelated and thus it was possible to analyze not only the raw potentials of each word category but also the semantic N400 effect, i.e. the negativity around 400 ms after stimulus onset which distinguishes unprimed from primed words [Anderson and Holcomb, 1995; Bentin et al., 1985; Holcomb, 1993]. Although Gomes et al. [1997] observed modality‐specific topographies for auditorily and visually presented stimuli, they did not find any topographic differences between verb and noun stimuli in the time window of the N400, neither in the raw potentials nor in the net N400 effect measured in the difference potential between primed and unprimed words.
Thus, the two studies provide contradictory evidence for the claim that average ERPs can reflect the topography of word‐category‐specific cortical cell assemblies. However, both studies suffer from some methodological deficits that limit the final conclusions. First, in both studies the topography was evaluated by a limited number of recording sites only (29 electrodes in Preissl et al. [1995], 30 in Gomes et al. [1997]). Such a small number of electrodes can provide a very crude picture of distinct cortical activation patterns only and small differences may have remained undetected. Moreover, measurements with EEG do not localize sources when only voltages are recorded. Second, the topography was analyzed with grand average data only. Thus, individual differences of the functional anatomy were not considered, despite the fact that the location of language sensitive areas in the cortex seems to vary considerably [Ojemann, 1983; Ojemann et al., 1989, see also Spitzer et al., 1998]. Third, in Preissl et al. [1995] the topographic difference between noun and verb processing was derived from unstandardized potential amplitudes. In that case a difference must not necessarily result from a genuine location difference of the generators but, rather, can be due to amplitude differences of one and the same generator [see McCarthy and Wood, 1985]. Finally, in both studies the experimental manipulations and the intended psychological processing differences were suboptimal. In the study of Preissl et al. [1995], no behavioral data were analyzed at all. Therefore, it cannot be excluded that there existed differences in response time which indirectly may have affected the ERP. With different response times the response‐related motor potential has a differently timed overlap with the word evoked ERP and this can result in amplitude differences. If the N400 net effect is analyzed by means of the difference potential, as in Gomes et al. [1997], such an unspecific influence can be excluded. However, the study of Gomes et al. suffers from another problem: The priming effect was weak with visual stimuli and in that case not significant for the noun/noun condition. Moreover, the priming conditions were not equivalent for different word categories because target nouns were primed by elements of the same and verbs by elements of a different word category. This may have obscured topographic differences because the “noun”‐specific centers were activated in both conditions.
In summary, the available evidence with respect to the question whether word category‐specific cell assemblies can be delineated by means of ERPs is not yet decisive. The objective of the present study, therefore, was to shed some more light on this issue by changing the measurement procedures, the methods of analysis, and the experimental manipulations. (1) To improve the measurement procedures we used a high‐density recording array of 124 electrodes. (2) Topographies were analyzed not only for grand averages but also by means of an individual estimation of cortical sources. Moreover, these estimates were constrained by means of the individual cortex anatomy derived from magnetic resonance images (MRI). (3) With respect to the psychology of the ERP‐evoking conditions we also used a primed lexical decision task. This allows a study of the N400 effect, i.e. the net activation pattern of the one or the other word category uncontaminated by effects that are not directly related to lexical activation or semantic integration processes. However, unlike Gomes et al. [1997], we varied the relatedness between prime and target in three steps (unrelated, moderately related, and closely related), and used elements of the same word category as primes and as targets, i.e. we presented either verb/verb or noun/noun pairs. This should restrict the lexical‐specific activation to the representations of the one or the other category only.
Given the above mentioned neuropsychological observations of an anterior‐posterior dissociation between verb and noun processing, and considering the semantic fact that most verbs are representing actions while concrete nouns are representing perceptual entities, we held the hypothesis that verbs should activate responses more anteriorly and nouns more posteriorly because their respective cell assemblies are located either closer to motor or to sensory projection areas.
The major variable of interest in our study, therefore, is the location of the cortical sources of the semantic N400 effect. Functionally, the semantic N400 effect seems to be a manifestation of processing steps that are invoked if the system encounters a word that is not primed by the preceding context. Its amplitude increases with increasing semantic and associative distance between a target word and the preceding context [Rösler and Hahne, 1992; Van Petten and Kutas, 1991]. Up until now, it is not completely clear if the N400 effect reflects primarily an automatic process of lexical access and/or more controlled processes of semantic integration [Anderson and Holcomb, 1995; Brown and Hagoort, 1993; Chwilla, Hagoort, and Brown, 1998; Holcomb, 1993]. For the present question this theoretical issue seems to be of secondary importance, because a topographic difference between noun‐ and verb‐evoked N400 effect can be expected for both processing steps. Automatic spread of activation should take place within the mental lexicon, and if verb and noun entries in this lexicon are located at anatomically distinct areas the topography of the N400 effect should be distinct, too. The controlled process of semantic integration that relates a word to the currently available semantic context involves an activation of lexical entries as well. If the meaning associations of verbs and nouns are stored at different locations, then their activation during semantic integration should result in a distinct topographical pattern of the N400 effect as well. Supporting evidence for the hypothesis that the topography of the N400 effect can vary with the type of activated representations comes from a study in which the N400 effect was either evoked by inappropriate words in sentences or by inappropriate numbers in mental calculation problems [Niedeggen et al., 1999]. In this study the descending part of the N400 effect showed a clear topographic difference, i.e. a left posterior maximum for the arithmetic and a centro‐parietal maximum for the semantic priming condition.
Estimation of cortical sources from ERP distributions
Estimating the cortical generators of surface‐measured electric or magnetic fields is an ill‐posed inverse problem that can only be solved by using additional constraints to limit the degrees of freedom [Nunez, 1981]. In some cases compelling a priori assumptions exist about the number and the location of sources. For example, the early components of the somatosensory‐evoked response can be well explained by a moving equivalent dipole solution, which locates two distinct generators in the primary and the secondary somatosensory cortex of each hemisphere [Elbert et al., 1995a, 1995b]. Comparable low‐dimensional dipole solutions have been described for the middle and late auditory evoked potential [Scherg and von Cramon, 1986]. In contrast, with the so‐called “cognitive” event‐related potentials, e.g., the N400 effect, the situation is much less specified. First, there is no plausible a priori reason to assume that such an effect is caused by a very small number of generators which can be modeled by few equivalent dipoles. Rather, an activation of lexical entries and semantic integration may involve widely distributed cortical networks. Therefore, a model is needed that restricts the location of sources to the cortex but leaves the number of contributing generators open. Second, mapping language functions by intracranial stimulation [Ojemann et al., 1989] and by fMRI [Spitzer et al., 1998] suggests that there exist substantial interindividual differences in the location of cell assemblies which are activated during word access in naming tasks. Therefore, possible interindividual differences in the location of estimated generators have to be taken into account as well.
Both conditions are met by recently developed current density reconstruction algorithms provided by the CURRY‐software® [Fuchs et al., 1994; Wagner et al., 1996]. These models are restricted by the individual topography of the cortical surface. The individual cortex anatomy is reconstructed from a structural magnetic resonance image and represented by a triangulated surface comprising some thousand elements. For each of these elements current sources are estimated from the measured individual potential distribution. Depending on further constraints a solution results which has either a rather broad distribution of many sources (L2 norm or minimum norm least squares (MNLS; see Method) or a distribution of a limited number and more focused sources (L1 norm). With the present set of data we applied both models.
MATERIALS AND METHODS
Participants
In total, 15 right‐handed subjects (11 women and 4 men) were recruited from the student population of the Philipps‐University of Marburg. Mean age was 23 years (range 19–32 years). All subjects were native speakers of German and had normal or cor‐ rected‐to‐normal vision. Data from two subjects had to be discarded because of too many artifacts during data collection. Thus, the final sample comprised 13 subjects. Subjects received a monetary compensation.
Material
A total set of 1,200 prime‐target pairs was constructed from German verbs and nouns. Of these, 600 had a verb and the other 600 a noun as prime. The target following the prime was selected from the same word category and was either strongly, moderately, or not associated with the prime, or it was a pseudoword. Pseudowords were derived from legal verbs or nouns by replacing at least one letter such that a pronounceable letter string resulted that did not match with an existing German word. Each event category with a word as target comprised 100 trials and each event category with a pseudoword as target 300 trials. The material had been selected on the basis of a pilot study in which association norms had been determined with an independent group of subjects [Rösler et al., 2000]. Moreover, the material was matched with respect to word frequency [CELEX database, 1995] and word length. Strong, moderate, nonassociated, and pseudoword targets in the group of noun‐stimuli were, for example: MEER‐OZEAN (approximate English translation: sea‐ocean), MEER‐TEICH (sea‐pond), MEER‐KALENDER (sea‐calendar), MEER‐*BLUBE (sea‐*floner), and in the group of verb stimuli KEHREN‐FEGEN (sweep‐dust), KEHREN‐WISCHEN (sweep‐wipe), KEHREN‐DRUCKEN (sweep‐print), KEHREN‐GEMEN (sweep‐*wank). Note that normally, verbs and nouns can be discriminated in German by the initial letter, which is lowercase for verbs and uppercase for nouns. To avoid any influence of orthography all words were presented with capital letters only. In the pilot study, reliable priming effects were observed with this material for both word categories, verbs and nouns [Rösler et al., 2000].
Procedure
The subject was seated in a comfortable chair in a dimly illuminated, sound attenuating, electrically shielded room. The stimuli were presented on a monitor (ATARI SM124, refresh rate 70 Hz) that was positioned at eye level 0.7 m in front of the participant. The font of the stimuli was small to prevent large saccades during reading. The height of the letters was 0.5 cm and the width of a word did not exceed 3 cm (visual angle 2.5°). Primes and targets were presented for 200 ms each, separated by a blank screen of 50 ms length, i.e., stimulus‐onset‐asynchrony (SOA) was 250 ms. All stimuli (prime‐target pairs) were presented in random order with the restriction that trials using the same prime were separated by at least 50 trials. Subjects were instructed to make a lexical decision with respect to the status of the target (word‐nonword). The response was given by moving either the index or the middle finger of the right hand out of a cavity equipped with a light gate. The mapping of the responding fingers to the response category was permuted across the participants. The intertrial interval varied randomly between 3 and 3.5 sec. Trials were presented in blocks of 40, which were separated by a short break of about 20 sec.
Subjects first performed a short training session in which another set of 40 prime‐target word pairs were presented. These stimuli showed no duplication with the stimuli of the main experiment. The total session lasted for approximately 1 hour.
EEG recording, artifact handling, and signal extraction
The EEG was recorded from 124 electrodes using a cap in which AgAgCl inserts were fixed by individual electrode supports (System Falk Minow, Munich). Positions of the 124 scalp electrodes were arranged in a triangular pattern and covered the upper part of the head. The central electrode row matched the Fpz‐Inion meridian of the 10–20 system (see Fig. 1). The cap was positioned on the head with reference to the nasion, inion, and the preauricular notches so that the vertex electrode was positioned correctly. The positioning of each electrode was then determined by means of a 3D coordinate tracking system (Polhemus fastrack) and stored on disk.
Figure 1.
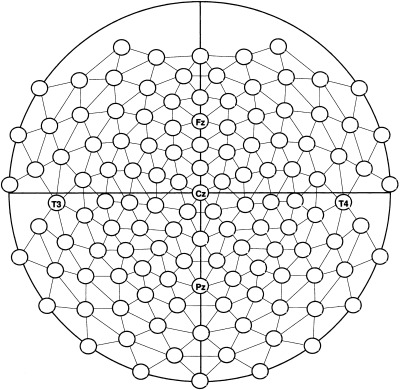
Layout of the 124 channel electrode array. Some of the prominent 10–20 electrode system positions are marked.
All scalp electrodes were referenced to the tip of the nose. Eye movements and blink artifacts were monitored by means of two bipolar channels. Two electrodes placed at the outer canthi of the two eyes provided the horizontal EOG and two electrodes at the sub‐ and supra‐orbital ridges of the left eye the vertical EOG. Electrode impedance was kept below 8 kΩ. The left mastoid served as ground. Recording bandpass was set from DC to 40 Hz and the digitization rate was 250 Hz. Trials in which the vertical EOG exceeded ±125 μV or in which the range between the highest and lowest signal exceeded 350 μV at one of the scalp electrodes were rejected. Furthermore, drift artifacts were corrected according to the method suggested by [Hennighausen et al., 1993]. Only trials with correct responses were used for the average. For further analysis, stimulus‐locked averages were computed for all targets separately for each word category and association level. Averages comprised between 43 and 96 trials with a median of 71 trials.
Structural MRI and anatomy reconstruction
Individual anatomical features were determined by a 1.5. Tesla T2‐weighted MRI scan (3D fast‐spoiled gradient echo, TE = 1.5 ms, TR = 11 ms).
Data analysis
ERPs
Difference potentials were computed for each word category by subtracting point by point the potential evoked by strongly related targets from that evoked by unrelated or moderately related targets. These difference potentials reflect the net N400 effect. The expression of the N400 effect was measured by the average amplitudes within a time window of 52 ms centered around the global field power maximum. The latency of the global field power maximum coincides by and large with the latency where the N400 effect has its maximum [Niedeggen et al., 1999]. This measurement procedure guarantees that the amplitudes are not contaminated by activity that contributes with equal strength to both raw potentials—for example, sensory or motor components. Moreover, because the measurements are adjusted to the individual‘s effect maximum for each condition, the results are not biased by the fact that the latencies of the verb and the noun priming effect are different. These amplitude measures were submitted to an ANOVA with the factors of word category (noun, verb), associative strength (strong, moderate), and electrode. A nonadditive repeated measures ANOVA model was used which explicitly considered possible interactions between subjects and conditions in the model terms [Winer, 1971]. The degrees of freedom of the F tests were adjusted using the procedure described by [Geisser and Greenhouse, 1958].
Source estimation
Cortical current densities were estimated as current per local surface area (unit: μA/mm2). Current densities were individually estimated on the surface of the cortex of each subject. The individual gray matter, the cerebral spinal fluid, the skull, and the skin of each participant were determined from T2‐weighted nMR images by gray‐value analysis as described by [Wagner, 1989]. The compartments of the cortical surface are then represented by a net of triangles. We restricted the number of surface triangles of the gray matter to ≈30,000. The adjustment is good enough to represent the shape of each cortex as a smooth surface while the number of unknown variables in the model equation are not unnecessarily increased.
The estimation approach is summarized in equation (1). The aim is to find the vector j of n cortical current source parameters that minimizes Δ2, i.e. the difference between the measurement vector m with s = 124 amplitude values and the forward solution Lj restricted by the necessary normalization matrix W and the regularization parameter λ.
| (1) |
The Lead‐Field matrix L represents the current flow attenuation from its origin through the compartments cerebral fluid, skull, and skin to the measurement sensors, i.e. the 124 electrodes. Here ‖•‖ symbolizes the squared Euclidean norm, which in our case is the squared sum of all the elements between the double bars (sum over all s electrodes and n current source parameters, with n = r*c, i.e. r cortex locations and c source components per location). The regularization term in equation (1) is necessary because the number of unknowns, i.e. the strength (and direction) of currents at each of the ≈30,000 triangles exceeds by far the number of knowns, i.e. the potentials measured at each of the 124 sensors at one point in time. Therefore, the regularization term is defined to pick one solution out of the manifold of solutions with equal deviation Δ2. The regularization parameter λ defines to what extent the model term ‖Wj‖2 is weighted relative to the data term ‖m‐Lj‖2. We adjusted λ so that the deviation Δ2 corresponds to the expectancy value of a χ2 distribution. In that case the explained variance corresponds to 1‐1/SNR (SNR = signal‐to‐noise ratio of the data). This is highly recommended to avoid a solution that explains more variance than can be attributed to the signal in the data. Clearly, the shape of the solution depends on the choice of the regularization matrix W. We chose a simple model with the matrix W being the unit matrix. This implies that the sum of all squared currents has to be a minimum. This model with an Euclidean metric is called minimum norm least squares’ (MNLS) and its solution is characterized by a rather broad distribution of mainly weak‐loaded current densities [Wagner et al., 1996].
Changing the metric from an Euclidean to a Minkowski L1 norm, also called city block metric, results in equation (2).
| (2) |
This L1‐norm model provides solutions in which a limited number of strong current densities describe the measurement. The solutions look similar to those obtained by fitting equivalent dipoles, but in the L1‐norm model the number of dipoles is not restricted by a priori assumptions. Rather, all ≈30,000 source triangles can contribute to the result. However, most of them achieve current strengths which are equal or close to zero.
Both models restrict the currents to be perpendicular to the local surface triangles representing the gray matter of each individual cortex. This constraint is physiologically realistic because potentials measured at scalp electrodes are most likely to be generated by pyramidal cells [Nunez, 1981], and these are arranged perpendicular to the gray matter surface. This constraint reduces the number of unknowns to one per triangle, since only the strength of the current at each of the local cortex triangles has to be estimated. Without this constraint two more unknowns, i.e. the angles of each of the currents, would have to be estimated, too. While the individual cortex was represented as realistic as possible by the triangulated surface, the other compartments between generator and sensor (cerebral‐spinal fluid, skull, skin) were defined by a spherical shell model. This semirealistic head model reduces computation time in comparison to a full realistic head model without distorting the results. Simulation studies and tests with real data did not show differences greater than the accuracy we expect to locate the sources in this study. Sources were estimated at the same time points as described for the ERP data, i.e. the average amplitudes of a 52 ms wide window arranged symmetrically around the global field power maximum were used as measurements m in equations (1) and (2). Cortical current source models were fitted to the difference potentials (unrelated‐ strongly related, and moderately related‐strongly related) of each word category and subject.
Comparable locations of “hot spots” in different subjects or conditions can only be detected by means of a statistical analysis of the locations of estimated sources. To take into account the different shapes and sizes of individual cortices the results were projected onto a “normalized brain” by means of a stepwise linear transformation as described by Talairach and Tournoux [1988]. For each individual brain, the anterior commissure was identified in the central saggital MRI slice and taken as the origin of a coordinate system with the x‐axis defined by the vector from the left to the right preauricular point, the y‐axis as the vector from inion to nasion, and the z‐axis perpendicular to x and y. Along these coordinates a 3D lattice was constructed by dividing the maximum distance of each axis into equally spaced steps. The result is a bricklike structure that encapsulates the whole brain (see Fig. 2). The sum of surface elements with a significant current source was then determined for each lattice element, separately for each subject and condition. These data are directly comparable across subjects and were investigated with an ANOVA with factors “lattice elements” and “word category.” This analysis tests the null hypothesis that the estimated cortical currents are equally distributed across all lattice elements (main effect) and distributed in like manner across all lattice elements for both word categories (interaction “word category * lattice element”).
Figure 2.
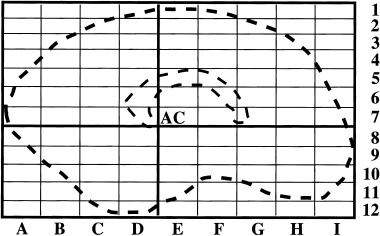
“Normalized brain voxels” constructed by means of a stepwise linear transformation. The anterior commissure (AC) is taken as the origin of a coordinate system with the x‐axis defined by the vector from the left to the right preauricular point, the y‐axis as the vector from inion to nasion, and the z‐axis perpendicular to x and y. Along these coordinates a 3D grid is constructed by dividing the maximum distance of each axis into equally spaced steps.
RESULTS
Performance data
Response times and errors were analyzed for the total set of 15 subjects. The response times revealed substantial priming effects for both word categories (see Table I). This is confirmed by an ANOVA with factors “word category” and “associative relatedness.” It provides significant main effects “word category” (F(1,14) = 140.19, P = 0. 0001), “relatedness” (F(2,28) = 68.09, P = 0. 0001, ϵ(GG) = 0. 7990), and a significant interaction (F(2,28) = 9.14, P = 0. 0025, ϵ(GG) = 0. 7757). Post‐hoc Scheffé tests revealed that the differences between unrelated and strongly related items, compared with unrelated and moderately related items, respectively, were significant (P < .01) for both word categories. In both word categories the priming effect is about twice as large for strongly related than for moderately related items, but the interaction indicates that the priming effects were somewhat more pronounced in the noun than in the verb condition. The main effect word category substantiates the observations that verb targets are, in general, processed slower than nouns (mean diff RT = 32 ms). Error rates show the same pattern of main effects (“word category” (F(1,14) = 43.26, P = 0. 0001), “relatedness” (F(2,28) = 30.55, P = 0. 0001, ϵ(GG) = 0. 7377)) but no significant interaction (F(2,28) = 0.39). The similarity of the effect patterns of both variables excludes that there was a speed‐accuracy trade‐off.
Table 1.
Reaction time, size of the priming effect, and error rate for the different priming conditions (Mean values in ms and %, ±standard deviation)
Grand Average ERPs and Surface Topographies
In Figure 3 the grand average ERPs elicited by noun and verb targets are shown for frontal, central, and parietal electrode positions. Superimposed are the ERPs of the three levels of “associative relatedness.” An N400 effect is present for both word categories, but it is more prominent and more systematic for nouns. For both categories, the unrelated targets are much more negative around 500 ms poststimulus than strongly related targets. In contrast, moderately related targets differ from strongly related targets only if they are nouns. This is more clearly revealed by the difference potentials which were computed by subtracting the potential of strongly related targets from that of unrelated targets and that of moderately related targets, respectively (see Fig. 4). The full topography of the N400 effect is shown in Figure 5. These topographic maps are computed from the average amplitude of a 50 ms wide window centered symmetrically around the maximum of the global field power. This individually adjusted measurement procedure minimizes the effect of individual time jitter and eliminates latency effects of the responses to nouns and verbs. An ANOVA of the average amplitudes measured ±25 ms around the maximum of the global field power provided the following main effects “word class” F(1,12) = 4.13, P = 0.065, “associative strength” F(1,12) = 17.45, P = 0. 0013, “electrodes” F(123,1476) = 6.20, P = 0. 0001, and the interactions “electrode * word class” F(123,1476) = 1.40, P = 0. 0038, ϵ(GG) = 0. 0528, “electrode * associative strength” F(123,1476) = 3.04, P = 0. 0001, ϵ (GG) = 0. 0506).
Figure 3.
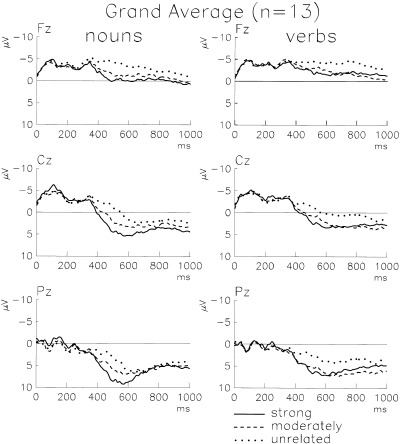
Grand average ERPs elicited by noun (left) and verb (right) targets following either strongly, moderately or unrelated primes (n = 13). Shown are the three midline electrodes Fz, Cz, and Pz. ERPs are aligned with respect to a 100 ms long baseline preceding word onset. Negativity is up.
Figure 4.
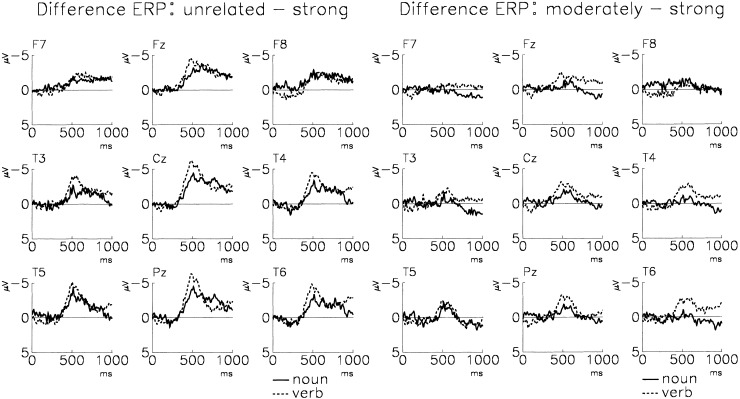
Shown are the grand average difference potentials which result from subtracting the potential of strongly related targets from that of unrelated targets and from that of moderately related targets, respectively, for both word categories. Superimposed are the difference potentials of the noun and verb condition, separately for the unrelated (left) and moderately related targets (right). Negativity is up.
Figure 5.
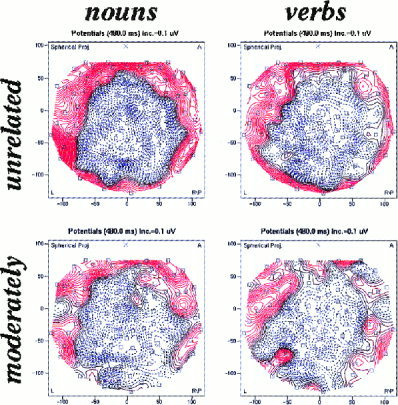
Topography of the difference ERPs to target words preceded by unrelated words minus those preceded by strongly related words for nouns (left) and verbs (right) (n = 13). The maps are based on an average amplitude of a 50 ms epoch symmetrically adjusted to the maximum of the global field power.
The main effect word class is due to the fact that, on average, the N400 amplitude is larger for nouns than for verbs and the main effect associative strength confirms that unrelated targets evoke a larger N400 effect than moderately related targets. The interaction “electrodes * word class” is weak, which suggests that there is not much of a topographic difference between the two conditions. Moreover, the question whether the average generator has a different location for the two word categories can only be answered by another ANOVA in which the amplitude measures are standardized, i.e. after amplitude and variance differences between the conditions are eliminated. This analysis detects genuine topographic differences which cannot be due to amplitude differences alone. In that case the interaction “electrodes * word class” should be significant. In our analysis this test provided F(123, 1476) = 1.36, P = 0. 0060, ϵ (GG) = 0. 0693, which is not significant after GG correction.
Single subject topographies
The grand average difference potentials showed a prominent N400 effect for unrelated minus strongly related targets and for both word categories. According to the group statistics the topography of this N400 effect did not differ between nouns and verbs. Thus, one should expect that there is not much variability of the topography across subjects. However, scanning the individual topographies proved the contrary. We exemplarily show the topographies of the difference potentials of three subjects in Figure 6. Maps of the N400 effect were computed as for the grand average above, i.e. based on the average amplitude of a 50 ms wide window arranged symmetrically around the maximum global field power. The left side of Figure 6 shows the unstandardized difference maps for nouns, the right side for verbs. These maps suggest that there might exist topographic differences between nouns and verbs for individual subjects. For example, subject 6 has a right hemisphere dominance of the N400 effect for nouns but a more central and posterior peak for verbs. Subject 8 shows a more posterior peak for nouns and a more anterior peak for verbs, while subject 16 has a clear, posteriorly located and symmetrically distributed N400 effect for both nouns and verbs. These striking individual differences motivated us to individually estimate the cortical current sources. As a possibility, one could expect that the measured potentials are caused by some sources which are common to all subjects and conditions (reflected by the grand average) and by some individual sources.
Figure 6.
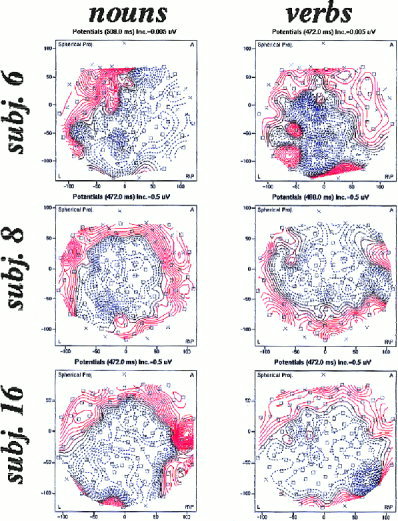
Single subject topographies of the N400 effect for nouns (left) and verbs (right). The difference potentials were computed by subtracting point by point the potential of strongly related items from that of unrelated items. An average amplitude was then computed for a 50 ms‐wide window extending symmetrically around the individual maximum of the global field power.
Cortical current density estimation
Cortical generators were estimated for the data of each individual subject. In the first step, we restricted our analyses to the difference potentials “unrelated‐strongly related” of both word categories. Since the N400 effect was weak for moderately related target verbs an estimation of sources of this effect makes less sense. However, to see how reliable our estimates were, we also estimated the cortical current sources for the difference potential of “moderately‐strongly related nouns.”
In Figure 7, as an example, we show the source estimates obtained with the L1‐norm model for three subjects. These are the same as in Figure 6. For each of the three subjects the currents of noun processing (red) and verb processing (green) are projected on the individual cortices, shown from the left and right side. The explained variance for these L1‐norm solutions for the three subjects 6, 8, and 16 amount to 88%, 90%, 91% for nouns, and 78%, 90%, 86% for verbs, respectively. The estimated sources of subjects 6 and 8 are more pronounced on the left hemisphere, those of subject 16, especially for nouns, are more pronounced on the right hemisphere. The figure reveals that the locations of estimated current sources are simliar for nouns and verbs but also that there is not a complete overlap. Moreover, a comparison of the three subjects reveals a large variability of the locations. The same was valid for the other 11 subjects. Figure 8 summarizes the locations of all estimated sources. In this figure each significant source of each subject and condition is projected on a sphere where it is represented as a dot: red for noun sources and green for verb sources. In this presentation of the results no prominent clusters pop out. To substantiate this impression statistically we transformed each individual result to the Talairach‐based coordinate system and summed the amount of each local current in each of the 864 lattice elements, separately for each subject and each word category. An ANOVA with factors “word category” and “lattice‐element” provided a significant main effect “word category” (F(1,12) = 9.89, P = 0. 0084) but an interaction which is not siginificant after GG correction (F(863,10356) = 1.24, P = 0.001, P (after GG) = 0. 2866). This implies that the estimated cortical current dipoles have the same distribution for both word categories and no prominent cluster across subjects.
Figure 7.
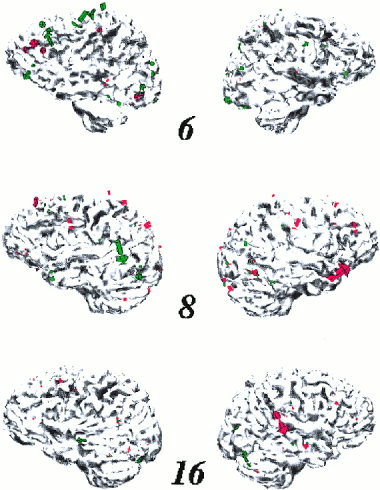
Cortical current sources of the N400 effect estimated with the L1‐norm model for three individual subjects. Sources are represented by arrows that are projected onto the individual cortices. The direction of each arrow is perpendicular to the local surface and its length represents the dipole moment in a local volume. Red arrows represent sources of nouns and green arrows sources of verbs on the left and right hemisphere.
Figure 8.

Estimated sources of the N400 effect of all subjects using the L1‐norm. Each dipole moment of each subject is represented by a dot, sources for nouns are red, sources for verbs are green. The 3D distribution is presented as a projection to a coronal sphere (view from top, nose is pointing upward).
As mentioned in the Methods section there is no a priori reason to assume that the L1‐norm model, which provides a limited number of focal sources, is the best choice. Therefore, we estimated the cortical current sources also by using the L2 norm (MNLS). One MNLS result is presented in Figure 9, it is the solution for nouns of subject 6. Thus the solution is the direct counterpart to the red arrows of subject 6 in Figure 7. As expected, the distribution of cortical current sources is much broader with the L2‐norm model. This solution explains 87% of total variance.
Figure 9.
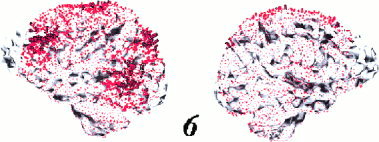
Estimated sources of the N400 effect of one subject (6) evoked by nouns using the L2‐norm. Sources are represented by arrows that are projected onto the individual reconstructed cortex. The direction of each arrow is perpendicular to the local surface triangle and its length represents the dipole moment in a local volume.
We again transformed each individual result to the Talairach‐based coordinate system and performed an ANOVA the same way as described of the L1‐norm result. With this analysis we observed a similar pattern of results, i.e. the interaction “word class * lattice element” was not significant after GG correction (F(863,10356) = 1.247, P = 0. 0001, P(GG) = 0. 2860). Again, the result is in agreement with the result obtained for the N400 amplitudes. It indicates that the location of sources estimated for the noun and verb evoked N400 effect does not differ systematically across subjects.
Moreover, both the L1‐ and the L2‐norm solution substantiate the observation of large interindividual differences in the location and strength of estimated sources. If the Talairach standardized analyses are run with the generalized least squares model, it is possible to estimate the contribution of the factor “subjects.” For the L1 norm this term provides F(12, 20724) = 24.98, P ≪ 0. 0001, and for the L2 norm F(12, 20724) = 206.24, P ≪ 0. 0001. In both cases, interindividual differences explain a much larger part of variance than the experimentally induced effect “word category.” Nevertheless, the individual solutions were reliable. To estimate the reliability we compared the results of the unrelated and the moderately related stimulus condition by correlating the dipole moments of the L1 norm of the two conditions across the Talairach coordinates. The average correlation, retransformed from Fisher′s z‐scores, amounted to 0.79 with a variance of 0.29.
DISCUSSION
The aim of the present study was to substantiate possible differences in the location of cortical generators related to verb and noun processing. To this end we used a priming task with verb/verb and noun/noun pairs and we analyzed the N400 effect in the EEG, i.e. the amplitude differences, which result, if the ERPs to items strongly related to a prime are subtracted from those evoked by items, which are either unrelated or moderately related to the prime. This difference reflects the net priming effect due to semantic relatedness. In brief, we observed the following results:
-
1
Response times indicated a significant priming effect for both word categories and both levels of associatedness; this effect was more pronounced for nouns than for verbs.
-
2
For unrelated items and both word categories the raw ERPs revealed a substantial and significant N400 effect. For moderately related items the N400 effect was present for nouns only.
-
3
The topography of the grand average N400 effect of verbs and nouns did not differ, neither for the raw amplitudes nor for the z‐score standardized amplitudes.
-
4
Cortical current source estimates of the N400 effect which were constrained by the individual cortex anatomy revealed a very broad distribution of active locations and pronounced interindividual differences. Within each subject the solutions for verbs and nouns showed substantial overlap but across subjects no systematic clustering was observed.
-
5
Cortical current source estimates obtained with the L1‐norm and L2‐norm model, respectively, differed in the distribution of sources over the cortex. The solutions were more circumscribed for the L1 norm and rather broadly distributed for the L2 norm. However, for the data of each subject the location of the “hot spots” found with the one or the other model was very similar.
-
6
Cortical current source estimates show a satisfactory reliability as estimated by the correlation of the source moments across Talairach lattice elements for solutions of moderately and unrelated targets.
Distinct priming effects for verbs and nouns?
The RT and the grand average ERP results replicate and extend previous findings and they prove that our experimental manipulations had been successful, i.e. substantial priming effects were induced with both word categories and both levels of relatedness. In former ERP studies semantic priming effects had been investigated mostly with nouns or with stimulus material not exactly specified with respect to the involved word categories (e.g., Anderson and Holcomb [1995] and Chwilla et al. [1998]). In the present study, however, this factor was explicitly controlled and we could provide unequivocal evidence that not only nouns can prime associated nouns but also that verbs can prime associated verbs. This is reflected by the behavioral data (the RT differences) as well as by the electrophysiological measures (the N400 effect). The pattern of effects observed for nouns and verbs proved as very similar for both levels of priming and both dependent variables. Nevertheless, in case of moderately related targets the N400 effect was larger and more systematic for nouns than for verbs. Our results extend the findings of [Gomes et al., 1997], because we used items from the same word category in both conditions, i.e. verb‐verb and noun‐noun pairs, while Gomes et al. [1997] had only used nouns as primes. Moreover, the RT priming effects observed in our study with visual stimuli were much larger than those observed by Gomes et al. Thus, our data show that a verb can prime significantly another semantically related verb even if the SOA is rather short. From a psycholinguistic perspective this is an interesting result, because the possibility of verb‐verb priming has been questioned for short SOAs [see Rösler et al., 2000].
Contrary to our expectations, however, we did not find reliable topographic differences of the grand average N400 effects for nouns and verbs. In each of the four combinations of word category and item relatedness the topography of the N400 amplitude was by and large the same. The maximum was always located at parieto‐central leads (Fig. 5). There are some differences at more peripheral leads (over the posterior temporal part of both hemispheres), but these differences were not reliable. This is all the more surprising, as we had used a rather dense electrode array of 124 recording sites. If there had been systematic topographic differences, even rather weak ones, we should have had a good chance to detect them. However, this was not the case. There are two explanations for this nil result, one concerns the involved processes, and the other the limitations of a statistical analysis based on group data.
Ad processes
Given that the grand average distributions of the N400 effect of the two‐word categories are indistinguishable, one has to conclude that the additional processes which are evoked by an unrelated target and which are reflected by the N400 effect [Rösler and Hahne, 1992; Van Petten and Kutas, 1991] take place in the very same cortical networks. This can imply that either the lexica of the two domains of words are represented by the same neuronal networks (given that N400 reflects an automatic priming effect), or that the semantic integration processes which are triggered by unrelated or moderately related words make use of the same set of cell assemblies. For the time being we do not know exactly which of the two alternatives is the more likely one. There is evidence supporting the idea that the N400 effect is functionally linked to controlled processes of semantic integration [Chwilla et al., 1998], but for the time being it cannot be excluded that at least some variation of the N400 effect may also be related to an automatic activation of memory representations, in particular if the SOA is short [Niedeggen and Rösler, 1999; Niedeggen et al., 1999].
Speculating about the possible processes which are involved in the priming task and which may become manifest in the N400 effect another explanation of the identical topography has to be considered: The verb‐verb priming effects may have been mediated, at least in part, by nouns, too. I.e. a verb prime may have triggered associated nouns and these, in turn, may have preactivated some verb related representations, or, as another possibility, the processing of the second verb may have profited from backward priming effects which triggered the same mediating noun associations as the prime (see, e.g., Zwitserlood et al. 1996). In that case it would not be too surprising that verb targets evoked a similar topography as noun targets, because an activation of noun entries would have taken place in both cases. However, even if verbs had activated mediating nouns, the verbs themselves should have activated at least some verb‐specific representations which are distinct from the noun representations. So, the neuronal networks activated in the noun‐noun and the verb‐verb priming condition should have been at least partially different. These differences could have been small and thus could have led to small topographic differences only, but if so, it would nevertheless be surprising if we did not detect them with a spatially very sensitive array of 124 electrodes. Moreover, the strongly related verb targets showed considerable semantic overlap with the primes, i.e. they mostly denoted the very same action (KEHREN‐FEGEN, sweep ‐ dust), and therefore, it seems likely that the verb prime triggered directly the related verb entry. All these arguments suggest that nouns and verbs in our task must have triggered representations which are at least partially distinct. However, these lexically‐specific processes seem not to become apparent in the grand average N400 effect. Rather, it looks as if the N400 effect reflects a process which is unspecific with respect to the lexical category of the triggering word.
It has to be added that we also looked for possible topographic differences in the original ERPs evoked by the primes and the targets (see Rösler et al., 2000, for a detailed report of these results). These analyses showed that the ERPs evoked by the primes were virtually identical for both word categories. There was just a small amplitude difference, but an analysis with standardized z‐scores revealed absolutely no genuine topographic difference between verb‐ and noun‐primes. The z‐score analysis of the ERPs evoked by the targets, on the other hand, indicated a slight topographic difference between nouns and verbs. However, this topographic difference is difficult to interpret, because nouns were processed faster than verbs (see Table I). A difference in processing time can result in a different overlap of ERP compontents related to lexical access and semantic interpretation on the one side and of ERP compontents related to lexical decision and response preparation on the other. Thus, a topographic difference between noun and verb targets could be due to the fact that response‐related brain areas are activated with a different time course in the two conditions, while the processes related to lexical activation are nevertheless identical in timing and topography. This possibility also applies to the results of Preissl et al. [1995]. Therefore, we give more weight to the effects observed with the primes and the response‐uncontaminated difference potentials (N400‐effects). Both sets of data gave no clue that ERPs evoked by verbs and nouns are topographically distinct. This clearly invalidates the hypothesis that verbs and nouns are representated and activated within distinct cortical areas (see also Brown et al., 1999, for further evidence of, by and large, identical ERPs for nouns, verbs, and adjectives in a normal reading situation).
Ad limitations of statistical analysis
Nevertheless, it would be premature to draw a final conclusion about the involved processes from the present findings. The raw ERPs, i.e. the difference potentials and the topographies derived from them, provided evidence for very pronounced interindividual differences. The maximum of the N400 effect was different for each subject and so was the complete topography. Moreover, the topographies for verbs and nouns were also different within each subject. These inter‐ and intraindividual differences were reflected by the current source estimates. Given these differences, it could be that different processes are involved during noun and verb processing within each subject, but that their neural localization was too idiosyncratic to be detected by any group statistic. This would be in line with observations from cortical stimulation studies and fMRI studies [Ojemann, 1983; Ojemann et al., 1989; Spitzer et al., 1998] which showed considerable variation of language‐specific cell assemblies over subjects. Thus, the grand average N400 effect might pick up the common activation pattern only, while minor and possibly functionally relevant individual differences are lost and treated as unsystematic error variance in the statistical analyses.
Cortical current source estimates
In the present study we used a new method to delineate the cortical sources of scalp‐recorded ERP effects. This method utilizes the information of a large number of recording sites and it constrains the estimates by the individual cortex anatomy. This approach guarantees a much more predetermined and realistically constrained solution than fitting equivalent dipoles. Equivalent dipole solutions must be constrained by a priori hypotheses about possible “hot spots.” These are easily available in case of sensory evoked potentials whose generators are most likely located in the primary and secondary projection areas. For components related to cognitive processes, as the N400 effect, similar compelling a priori hypotheses, however, are not available. The neuronal counterparts of these processes are hardly known yet and, therefore, any solution must be arbitrary.
The advantage of the present method is that it needs no hypothesis about a specific generator location. Rather, the only constraint is that the sources are located within the cortex, which is individually determined as a triangulated surface from the structural MRI. The algorithm will then fit the most likely solution with respect to the measured potential distribution. Depending on the model (L1 or L2 norm), one gets either a more localized or a more distributed pattern of current sources.
As any method that derives scores from a model fitting algorithm, the source estimation procedure “cleans” the data from “error variance” in the sense that unsystematic variations are not “used” by the model. With PCA factor scores this usually has the consequence that experimental effects are more clearly represented by the derived (factor) scores than by the original data (i.e. the effect variance becomes larger in relation to the remaining error variance, see Rösler and Manzey, 1981). In the present experiment such an effect could not be observed when the distribution of the derived scores (the current strengths) were analyzed with group statistics. This gives further evidence to the conclusion that there were indeed no differences in the location of verb‐ and noun‐specific cell assemblies.
The method is free in distributing the current sources over the triangulated cortex, and our results show that they indeed cover more or less the entire upper hemisphere of the head when they are accumulated over all subjects (Fig. 8). This broad distribution is compatible with the very broad extension of the N400 effect in the grand average. However, the individual analysis of source locations suggests that the broad potential distribution in the grand average might not be caused by a homogeneous source but rather by a very patchy arrangement of idiosyncratic sources which vary considerably across subjects. This nourishes some scepticism about what a grand average effect might actually reveal about the functioning of the brain.
Here again, however, one has to avoid premature conclusions. Although it is very likely that most of the variance observed for the estimated source locations is due to genuine intraindividual variance of the generators, it must not be the case completely. The goodness of fit of the estimated models is dependent on the quality of the data. We derived conservative estimates of the S/N‐ratio for the data to which the models were fitted and found them in the size of 5, i.e. there still was about 20% of noise in the average ERPs of each subject. Consequently, the individually fitted current sources explain 80% of the data variance. This is not too bad a value, but it leaves room for random variations of the estimated sources. And it is, of course, less than what can be achieved with sensory evoked potentials. There, several hundred or even some thousand trials can be easily repeated and the average may have an S/N‐ratio of about 40. With such a good S/N‐ratio the current source estimates are much more stable and they seem to converge to the correct ocations1. With cognitive components such a good fit will never be achieved because the S/N‐ratio cannot be reduced below a certain limit. This restriction follows from the experimental conditions and the number of replications which can be realized. In the present study, for example, the number of word pairs which can be constructed by controlling word frequency, word length, and relatedness, is limited to about 100. Considering that some trials will always be lost because of artifacts, the remaining sample size can hardly be larger than about 70 to 80. A frequent repetition of the conditions is also not possible because then the psychological processes will be changed (subjects will habituate, the nature of priming will change, etc.). Considering these limitations the cortical current source estimation procedures are not yet satisfactory if cognitive components are to be modeled. We are currently investigating the reliability of the interindividual differences which we observed in the source estimates. This should provide an answer to what extent the observed differences in the locations of verb‐ and noun‐specific current sources reflect true variance being functionally significant or only noise.
Footnotes
A pilot study in which weak electrical currents were applied to the fingertips of the right hand revealed very plausible results. Stimuli were applied in a random sequence to the finger tips with an ISI of 1000 ms ±500 ms. Stimulus strength had been individually adjusted and was well below pain threshold but clearly above absolute threshold. After artifact correction about 700 trials were averaged for each of the five digits. Cortical current sources were estimated with the L1‐norm model for the first prominent component of the SEP, i.e. a peak with an average latency of about 54 ms. The choice of the L1 norm as the appropriate model was motivated by expecting rather focal activation in the somatosensory area of the contralateral hemisphere. The results showed that the sources were indeed all located in the somatosensory area along the sulcus centralis.
REFERENCES
- Anderson JE, Holcomb PJ (1995): Auditory and visual semantic priming using different stimulus onset asynchronies: an event‐related brain potential study. Psychophysiology 32: 177–190. [DOI] [PubMed] [Google Scholar]
- Beauregard M, Chertkow H, Bub D, Murtha S, Dixon R, Evans A (1997): The neural substrate for concrete, abstract and emotional word lexica: a positron emission tomography study. J Cogn Neurosci 9: 441–461. [DOI] [PubMed] [Google Scholar]
- Bentin S, McCarthy G, Wood CC (1985): Event‐related potentials, lexical decisions and semantic priming. Electroencephalogr Clin Neurophysiol 60: 343–355. [DOI] [PubMed] [Google Scholar]
- Brown CM, Hagoort P (1993): The processing nature of the N400: evidence from masked priming. J Cogn Neurosci 5: 34–44. [DOI] [PubMed] [Google Scholar]
- Brown CM, Hagoort P, Ter Keurs M (1999): Electrophysiological signatures of visual lexical processing: open‐ and closed‐class words. J Cogn Neurosci 1: 261–281. [DOI] [PubMed] [Google Scholar]
- Caramazza A, Hillis AE (1991): Lexical organization of nouns and verbs in the brain. Nature 349: 788–790. [DOI] [PubMed] [Google Scholar]
- Chwilla DJ, Hagoort P, Brown CM (1998): The mechanism underlying backward priming in a lexical decision task: spreading activation versus semantic matching. Q J Exp Psychol Hum Exp Psychol 51A: 531–560. [Google Scholar]
- Damasio AR, Tranel D (1993): Nouns and verbs are retrieved with differently distributed neural systems. Proc Natl Acad Sci U S A 90: 4957–4960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniele A, Giustolisi M, Silveri C, Colosimo C, Gainotti G (1994): Evidence for a possible neuroanatomical basis for lexical processing of nouns and verbs. Neuropsychologia 32: 1325–1341. [DOI] [PubMed] [Google Scholar]
- Elbert T, Junghöfer M, Scholz B, Schneider S (1995a): Separation of overlapping neuronal sources in the first and second somatosensory cortices. Brain Topogr 7: 275–282. [DOI] [PubMed] [Google Scholar]
- Elbert T, Pantev C, Wienbruch C, Rockstroh B, Taub E (1995b): Increased cortical representation of the fingers of the left hand in string players. Science 270: 305–307. [DOI] [PubMed] [Google Scholar]
- Fiez J, Raichle ME, Balota DA, Tallal P, Petersen SE (1996): PET activation of posterior temporal regions during auditory word presentation and verb generation. Cereb Cortex 6: 1–10. [DOI] [PubMed] [Google Scholar]
- Fuchs M, Wagner M, Wischmann H‐A, Ottenberg K, Dössel O (1994): Possibilities of functional brain imaging using a combination of MEG and MRT In: Pantev C, Elbert T, Lütkenhöner B, editors. Oscillatory event related brain dynamics. London, New York: Plenum Press, p 435–457. [Google Scholar]
- Geisser S, Greenhouse SW (1958): An extension of Box's results on the use of the F distribution in multivariate analysis. Ann Math Stat 29: 885–891. [Google Scholar]
- Gomes H, Ritter W, Tartter VC, Vaughan HGJ, Rosen JR (1997): Lexical processing of visually and auditorily presented nouns and verbs: evidence from reaction time and N400 priming data. Cogn Brain Res 6: 121–134. [DOI] [PubMed] [Google Scholar]
- Hennighausen E, Heil M, Rösler F (1993): A correction method for DC drift artifacts. Electroencephalogr Clin Neurophysiol 86: 199–204. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ (1993): Semantic priming and stimulus degradation: Implications for the role of the N400 in language processing. Psychophysiology 30: 47–61. [DOI] [PubMed] [Google Scholar]
- Martin A, Haxby JV, Lalonde FM, Wiggs CL, Ungerleider LG (1995): Discrete cortical regions associated with knowledge of color and knowledge of action. Science 270: 102–105. [DOI] [PubMed] [Google Scholar]
- McCarthy G, Wood CC (1985): Scalp distributions of event‐related potentials: an ambiguity associated with analysis of variance models. Electroencephalogr Clin Neurophysiol 62: 203–208. [DOI] [PubMed] [Google Scholar]
- Niedeggen M, Rösler F (1999): N400‐effects reflect activation spread during arithmetic fact retrieval. Psychol Sci 10: 271–276. [Google Scholar]
- Niedeggen M, Rösler F, Jost K (1999): Processing of incongruous mental calculation problems: evidence for an arithmetic N400‐effect. Psychophysiology 36: 307–324. [DOI] [PubMed] [Google Scholar]
- Nunez PL (1981): Electrical fields of the brain. New York: Oxford University Press. [Google Scholar]
- Ojemann G (1983): Brain organization for language from the perspective of electrical stimulation mapping. Behav Brain Sci 6: 189–206. [Google Scholar]
- Ojemann G, Ojemann J, Lettich E, Berger M (1989): Cortical language localization in left, dominant hemisphere. J Neurosurg 71: 316–326. [DOI] [PubMed] [Google Scholar]
- Preissl H, Pulvermüller F, Lutzenberger W, Birbaumer N (1995): Evoked potentials distinguish between nouns and verbs. Neurosci Lett 197: 81–83. [DOI] [PubMed] [Google Scholar]
- Rösler F, Hahne A (1992): Hirnelektrische Korrelate des Sprachverstehens: zur psycholinguistischen Bedeutung der N400‐Komponente im EEG. Sprache und Kognition 11: 149–161. [Google Scholar]
- Rösler F, Manzey D (1981): Principal components and VARIMAX‐rotated components in event‐related potential research: some remarks on their interpretation. Biol Psych 13: 3–26. [DOI] [PubMed] [Google Scholar]
- Rösler F, Streb J, Haan H (2000): Word category specific priming? Event‐related brain potentials evoked by verbs and nouns in a lexical decision task. submitted. [PubMed]
- Scherg M, von Cramon D (1986): Evoked dipole source potentials of the human auditory cortex. Electroencephalogr Clin Neurophysiol 65: 344–360. [DOI] [PubMed] [Google Scholar]
- Spitzer M, Kischka U, Gückel F, Bellemann ME, Kammer T, Seyyedi S, Weisbrod M, Schwartz A, Brix G (1998): Functional magnetic resonance imaging of category‐specific cortical activation: evidence for semantic maps. Cogn Brain Res 6: 309–319. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P (1988): Co‐planar sterotaxic atlas of the human brain. Stuttgart: Thieme. [Google Scholar]
- Van Petten C, Kutas M (1991): Electrophysiological evidence for the flexibility of lexical prosessing In: Simpson GB, editor. Understanding word and sentence. Amsterdam: Elsevier, p 129–174 [Google Scholar]
- Wagner M (1989): Rekonstruktion neuronaler Ströme aus biomagnetischen Messungen auf der aus MR‐Bildern segmentierten Hirnrinde. Aachen: Shaker Verlag. [Google Scholar]
- Wagner M, Wischmann H‐A, Fuchs M, Köhler T, Drenckhahn R (1996): Current density reconstructions using the L1 norm. In: Proceedings of the Biomag 96.
- Winer BJ (1971): Statistical principles in experimental design (2nd ed). New York: McGraw‐Hill. [Google Scholar]
- Zwitserlood P, Drews E, Bolwiender A, Neuwinger E (1996): Kann man Geschenke umbringen? Assoziative Bahnungsexperimente zur Bedeutungsheterogenität von Verben In: Habel C, editor. Perspektiven der kognitiven Linguistik: Modelle und Methoden. Opladen: Westdeutscher Verlag, p 211–232. [Google Scholar]