

Abstract
Background
Prostaglandin F2α (PGF2α) may differentially affect viability of luteal cells by inducing either proliferation or cell death (via apoptosis or necroptosis). The diverse effects of PGF2α may depend on its local vs. systemic actions. In our study, we determined changes in expression of genes related to: (i) apoptosis: caspase (CASP) 3, CASP8, BCL2 associated X (BAX), B-cell lymphoma 2 (BCL2) and (ii) necroptosis: receptor-interacting protein kinase (RIPK) 1, RIPK3, cylindromatosis (CYLD), and mixed lineage kinase domain-like (MLKL) in the early and mid-stage corpus luteum (CL) that accompany local (intra-CL) vs. systemic (i.m.) analogue of PGF2α (aPGF2α) actions. Cows at day 4 (n = 24) or day 10 (n = 24) of the estrous cycle were treated by injections as follows: (1) systemic saline, (2) systemic aPGF2α (25 mg; Dinoprost), (3) local saline, (4) local aPGF2α (2.5 mg; Dinoprost). After 4 h, CLs were collected by ovariectomy. Expression levels of mRNA and protein were investigated by RT-q PCR, Western blotting and immunohistochemistry, respectively.
Results
We found that local and systemic administration of aPGF2α in the early-stage CL resulted in decreased expression of CASP3 (P < 0.01), but CASP8 mRNA expression was up-regulated (P < 0.05). However, the expression of CASP3 was up-regulated after local aPGF2α treatment in the middle-stage CL, whereas systemic aPGF2α administration increased both CASP3 and CASP8 expression (P < 0.01). Moreover, we observed that both local and systemic aPGF2α injections increased RIPK1, RIPK3 and MLKL expression in the middle-stage CL (P < 0.05) while CYLD expression was markedly higher after i.m. aPGF2α injections (P < 0.001). Moreover, we investigated the localization of necroptotic factors (RIPK1, RIPK3, CYLD and MLKL) in bovine CL tissue after local and systemic aPGF2α injections in the bovine CL.
Conclusion
Our results demonstrated for the first time that genes related to cell death pathways exhibit stage-specific responses to PGF2α administration depending on its local or systemic actions. Locally-acting PGF2α plays a luteoprotective role by inhibiting apoptosis and necroptosis in the early CL. Necroptosis is a potent mechanism responsible for structural CL regression during PGF2α-induced luteolysis in cattle.
Keywords: Necroptosis, RIPKs, Apoptosis, Prostaglandin F2α, Bovine CL
Background
The corpus luteum (CL) plays a crucial role in supporting pregnancy in cattle and other mammalian species, because of its production of progesterone (P4) [1, 2]. If pregnancy is not established, the bovine CL undergoes regression due to the action of uterine prostaglandin F2α (PGF2α) which is released in the late luteal phase of the estrous cycle [3].
The cascade of CL regression consists of: (i) functional luteolysis (interruption of steroidogenesis), and (ii) structural luteolysis (degradation/demise of CL tissue due to cell death) [4, 5]. Until now, a large number of reports have indicated that the caspase (CASP) – dependent apoptosis (type I programmed cell death) is the principal mechanism of CL cell death during structural luteolysis in cows [6, 7]. Several mediators are involved in the regulation and control of apoptosis in the CL, among them: B-cell lymphoma 2 (BCL2), and BCL2-associated X (BAX), which belong to the bcl-2 protein family (8), and caspases (CASP) [8, 9].
Recently, Hojo et al. [10, 11] proposed that necroptosis (CASP – independent cell death pathway) is an alternative luteolytic mechanism responsible for death of luteal steroidogenic cells (LSC) and luteal endothelial cells (LEC) and for their elimination from the bovine CL during luteolysis. This process is characterized by disrupted cellular membranes with leakage of their intracellular contents and tissue damage [12, 13]. In the clasical necroptosis pathway, receptor-interacting protein kinase 1 (RIPK1) is necessary for the activation of receptor-interacting protein kinase 3 (RIPK3) [14, 15]. Moreover, the deubiquitination of RIPK1 by cylindromatosis (CYLD), a K63-specific deubiquitinating enzyme (DUB), is crucial for initiation of necroptosis and mitochondrial complex II formation [16]. In the absence of CYLD, the generation of complex II is inhibited. The activation of RIPK3 and RIPK3 substrate-mixed lineage kinase domain-like (MLKL) by its phosphorylation [17] are key steps during the execution of necroptosis [18].
In farm animals, PGF2α and its analogues (aPGF2α) are widely used as pharmacological tools to induce luteolysis [19]. However, the newly formed CL is refractory to exogenous PGF2α before day 5 of the estrous cycle. Therefore, a single PGF2α treatment is ineffective for inducing luteolysis during the early luteal phase [19, 20]. Although the luteolytic action of PGF2α on the regression process has been widely studied [21–23], the mechanism of CL insensitivity, the acquisition of luteolytic capacity by the CL as well as mechanisms related to its stage-specific response to PGF2α all still need intensive studies. Previous studies [23–26] have suggested that the different actions of PGF2α on steroidogenesis pathways, immune functions and on pro- or anti-angiogenic factors may depend on the phase of the estrous cycle: the early-stage CL (PGF2α-resistant) vs. middle-stage CL (PGF2α-responsive). However, the effects of PGF2α on luteal steroidogenic cells may depend on its local, direct (autocrine/paracrine modes of action) effect or on indirect effects including several regulatory mechanisms within the female reproductive tract (e.g., endocrine action, blood flow regulation, contribution of the immune system, etc.) [27–29].
Prostaglandin F2α is essential for manipulate bovine reproduction because in dairy cattle farming using of hormonal treatments are very common procedures to influence the estrous cycle. General in this study, we demonstrated the effect PGF2α on new mechanism involved in the CL regression in cows (necroptosis), and that could provide new knowledge to optimize breeding methods of cows. Therefore, we intended to extend the understanding of the luteolytic process, and we hypothesised that PGF2α might induce various mechanisms of cell death (differences in luteal responses) in the bovine CL depending on its peripheral and local actions during the early and mid-luteal phase of the estrous cycle. The aim of the present study was to examine the differences in expression of genes related to: (i) apoptosis (CASP3, CASP8, BAX, BCL2) and (ii) necroptosis (RIPK1, RIPK3, CYLD, MLKL) in response to intra-CL (local) or i.m. (systemic) aPGF2α injections in the early- (day 4 of the estrous cycle) vs. middle-stage (day 10) bovine CL.
Results
Experiment 1. Changes in mRNA expression and protein concentration of CASP3, CASP8 and the ratio of BCL2 to BAX in the early- and midle-stage CL in response to local or systemic administration of aPGF2α
Figures 1 and 2 show the results for analysis of mRNA expression and protein concentration of CASP3 and CASP8 in the early and middle-stage bovine CL. An opposite effect of local and systemic aPGF2α action was observed in the early- versus middle-stage CL (P < 0.0001; Fig. 1A and 2A). Local and systemic administration of aPGF2α resulted in decreased mRNA expression (P = 0.0073, P = 0.0003, respectively; Fig. 1A) and protein concentration of CASP3 (P = 0.0021, P = 0.0038, respectively; Fig. 2A) in the early-stage CL, while both aPGF2α treatments increased CASP3 mRNA expression (P < 0.0001; Fig. 1A) and protein concentration (P < 0.0001; Fig. 2A) in the middle-stage CL. Hovewer, CASP8 mRNA expression was up-regulated by local and systemic aPGF2α injections in the early-stage CL (P = 0.0442, P = 0.0383, respectively; Fig. 1B), with no effect on CASP8 protein concentration (P = 0.4715, P = 0.9969, respectively; Fig. 2B). Additionally, only systemic aPGF2α injection increased CASP8 mRNA expression (P = 0.0129; Fig. 1B) and protein concentration (P = 0.0152; Fig. 2B) in the middle-stage CL.
Fig. 1.
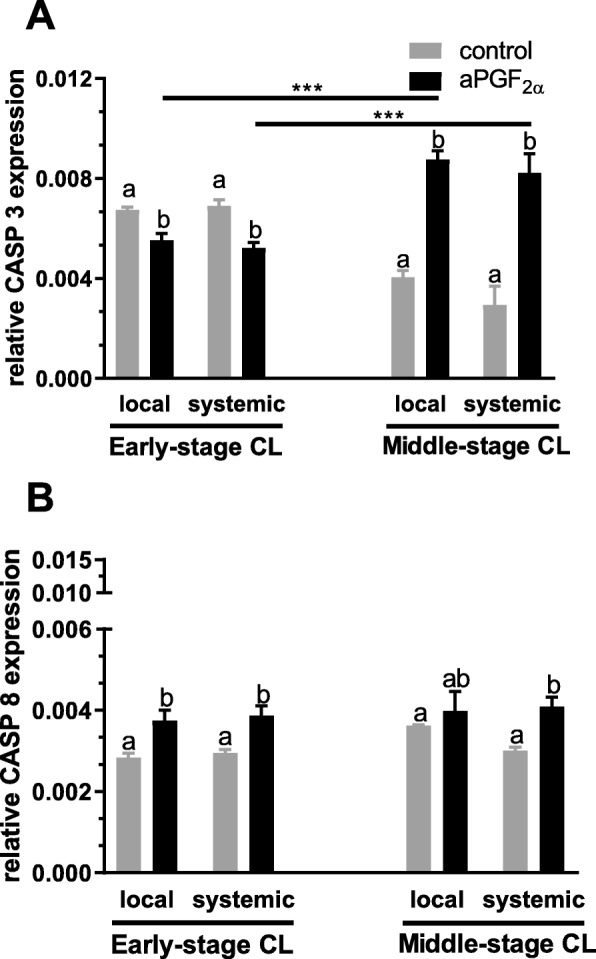
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the mRNA expression of (a) caspase 3 (CASP3), (b) caspase 8 (CASP8) in the early- and middle- stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Fig. 2.
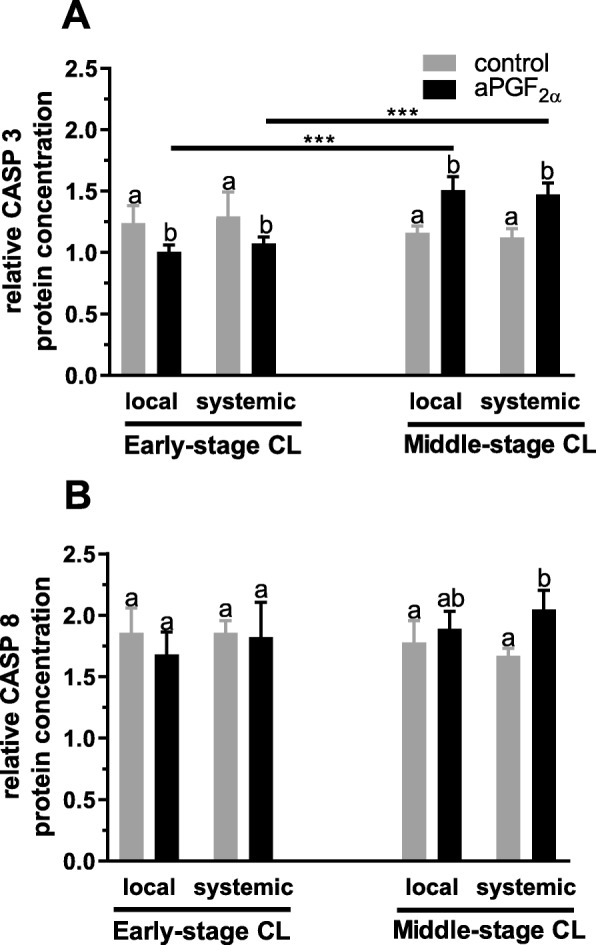
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the protein concentration of (a) caspase 3 (CASP3), (b) caspase 8 (CASP8) in the early- and middle- stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Figure 3 shows the results for analysis of mRNA expression and protein concentration of the ratio of BCL2 to BAX in the early- and middle-stage CL. An opposite effect of local and systemic aPGF2α action was observed in the early-stage CL versus middle-stage CL (P = 0.0003, P < 0.0001, respectively; Fig. 3A), namely local and systemic aPGF2α administration increased the ratio of BCL2 to BAX mRNA expression in the early-stage CL (P = 0.0164 and P < 0.0001, respectively; Fig. 3A), while both aPGF2α treatment decreased its mRNA expression in the middle-stage CL (P < 0.0001; Fig. 3A). Comparison of local to systemic administration of aPGF2α showed higher BCL2/BAX mRNA expression after systemic aPGF2α injection in the early-stage CL (P = 0.0099; Fig. 5A) while systemic aPGF2α action induced lower BCL2/BAX mRNA expression in the middle-stage CL (P < 0.0001; Fig. 3A). Moreover, both aPGF2α action enhanced the ratio of BCL2 to BAX protein concentration (P < 0.0001; Fig. 3B) in the early-stage CL, while their protein concentration was down-regulated by both local and systemic aPGF2α treatments in the middle-stage CL (P = 0.0084, P = 0.0244, respectively; Fig. 3B). Additionally, an opposite effect of local and systemic aPGF2α action on BCL2/BAX protein concentration was observed in early- versus middle-stage CL (P < 0.0001; Fig. 3B).
Fig. 3.
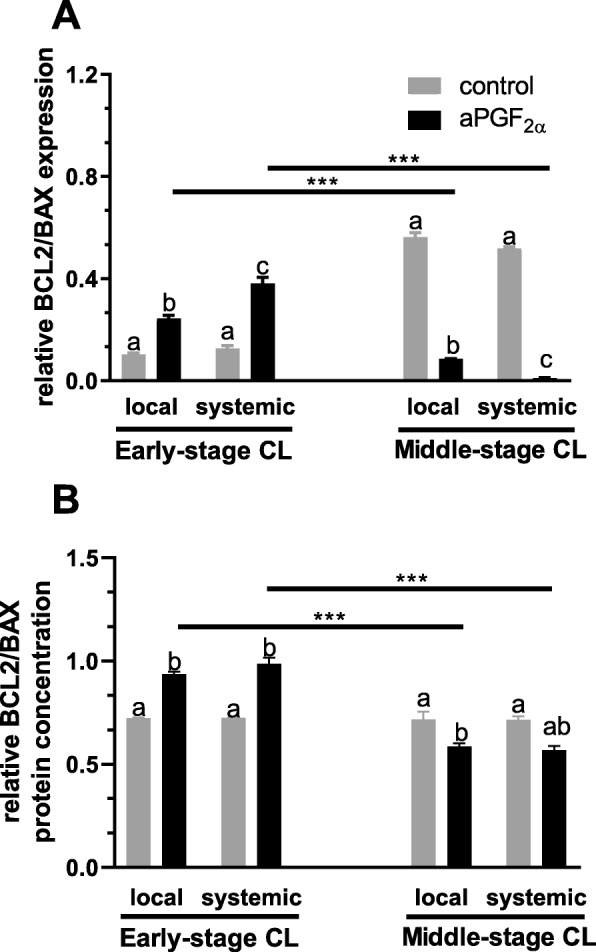
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the (a) the ratio of BCL2 to BAX mRNA expression and (b) the ratio of BCL2 to BAX protein concentration levels in the early- and middle- stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Fig. 5.
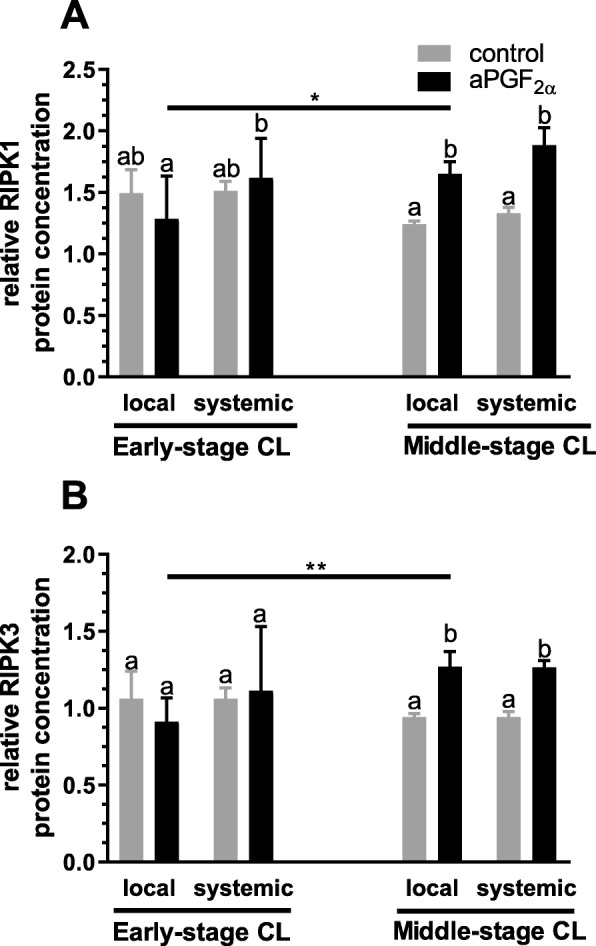
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the protein concentration of: (a) RIPK1, (b) RIPK3 in the early- and middle- stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Experiment 2. Changes in mRNA expression and protein concentration of RIPK1, RIPK3, CYLD, MLKL in the early- and middle-stage CL in response to local or systemic administration of aPGF2α
Figures 4 and 5 show the results for analysis of mRNA expression and protein concentration of RIPK1 and RIPK3 in the early- and middle-stage bovine CL. A local aPGF2α injection up-regulated RIPK1 mRNA expression (P = 0.0288; Fig. 4A) but down-regulated RIPK3 mRNA expression (P = 0.0130; Fig. 4B) in the early-stage CL. However, systemic aPGF2α administration resulted in a decrease in RIPK1 and RIPK3 mRNA expression in the early-stage CL (P = 0.0112, P = 0.0407; Fig. 4A and B). Furthermore, both local and systemic aPGF2α injections increased the mRNA expression of RIPK1(P = 0.0002, P < 0.0001, respectively; Fig. 4A) and RIPK3 (P = 0.0124, P < 0.0001, respectively; Fig. 4B) in the middle-stage CL. Additionally, we obserwed higher RIPK1 mRNA expression in the early-stage CL after local aPGF2α treatment compared with that in middle-stage CL(P < 0.0001; Fig. 4A). However, both routes of aPGF2α treatment had an opposite effect in RIPK3 mRNA expression in the early-stage CL compared with that action in the middle-stage CL (P < 0.0001; Fig. 4B). Moreover, both aPGF2α injections increased protein concentration of RIPK1(P = 0.0239, P < 0.0019, respectively; Fig. 5A) and RIPK3 (P = 0.0263, P = 0.0279, respectively; Fig. 5B) in the middle-stage CL. Only local aPGF2α action had an opposite effect on RIPK1 (P = 0.0128; Fig. 5A) and RIPK3 (P = 0.0068; Fig. 5B) protein concentration observed in the early- versus middle-stage CL. Additionally, comparison of local and systemic administration of aPGF2α showed differences in RIPK1 mRNA expression level in the early-stage CL (P < 0.0001; Fig. 5A). Moreover RIPK3 mRNA expression was greater in the middle-stage CL in response to systemic aPGF2α injection compared with the local treatment route (P = 0.0003; Fig. 5B).
Fig. 4.
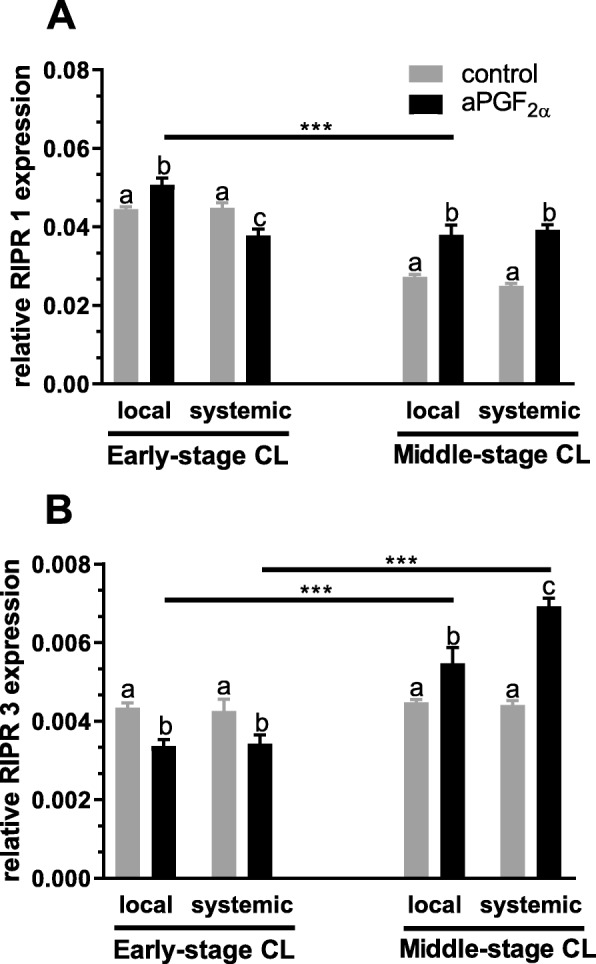
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the mRNA expression of: (a) RIPK1, (b) RIPK3 in the early and middle- stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b,c indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Figures 6 and 7 show the results for analysis of mRNA expression and protein concentration of CYLD and MLKL in the early- and middle-stage bovine CL. Only local aPGF2α administration up-regulated CYLD mRNA expression in the middle-stage CL (P = 0.0127; Fig. 6A). However, only systemic aPGF2α injection enhanced its protein concentration in the middle-stage CL (P < 0.0001; Fig. 7A). Furthermore, local aPGF2α injection induced down-regulation of MLKL protein concentration in the early-stage CL (P = 0.0002; Fig. 7B). On the other hand, local and systemic aPGF2α treatments increased its protein concentration in the middle-stage CL (P = 0.0033, P < 0.0001; Fig. 7B); however, this effect was greater when aPGF2α was injected systematically (P < 0.0001; Fig. 7B). Moreover, an opposite effect of local and systemic aPGF2α action on MLKL protein concentration was observed in the early- versus middle-stage CL (P < 0.0001; Fig. 7B).
Fig. 6.
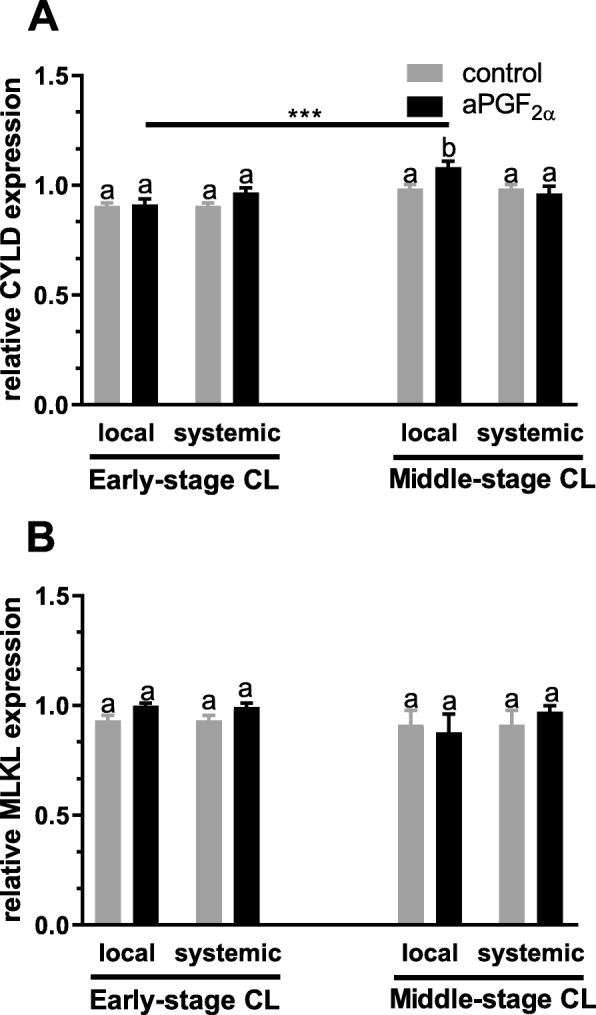
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the mRNA expression of: (a) CYLD, (b) MLKL in the early - and middle-stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Fig. 7.
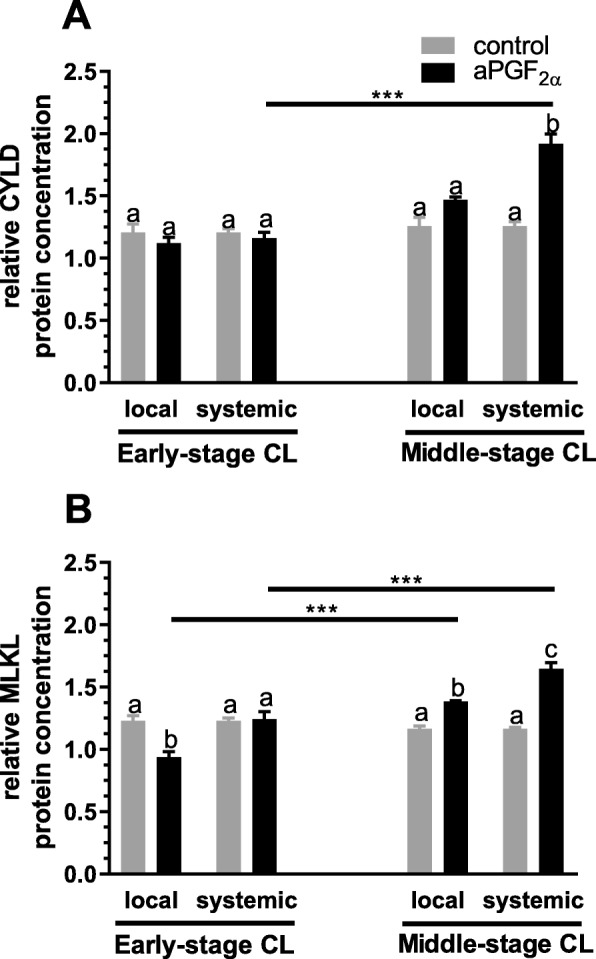
Effect of local or systemic PGF2α analogue (aPGF2α) administration on the protein concentration of: (a) CYLD, (b) MLKL in the early- and middle-stage corpora lutea (CL), respectively. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b,c indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early CL vs local/systemic aPGF2α injected middle-stage CL
Experiment 3. Immunohistochemistry localization and changes in intensities of RIPK1, RIPK3, CYLD, MLKL in the early- and middle-stage CL in response to local or systemic administration of aPGF2α
In another set of studies, we investigated the localization of RIPK1, RIPK3, CYLD and MLKL in bovine CL tissue after local and systemic aPGF2α treatment by immunohistochemistry. Representative sections of images are shown in Fig. 8 (early-stage CL) and Fig. 9 (middle-stage CL).
Fig. 8.

Representative section of images of localization of: (a, b, c) RIPK1, (d, e, f) RIPK3, (g, h, i) CYLD and (j, k, l) MLKL protein in the bovine early-stage corpora lutea (CL) at 4 h after local or systemic PGF2α analogue (aPGF2α) administration. Each small window shows a negative control stained with normal rabbit IgG instead of primary antibody. Positive immunohistochemistry staining was assessed as brown staining. Bar = 20 μm
Fig. 9.

Representative section of images of localization of: (a, b, c) RIPK1, (d, e, f) RIPK3, (g, h, i) CYLD and (j, k, l) MLKL protein in the bovine middle-stage corpora lutea (CL) at 4 h after local or systemic PGF2α analogue (aPGF2α) administration. Each small window shows a negative control stained with normal rabbit IgG instead of primary antibody. Positive immunohistochemistry staining was assessed as brown staining. Bar = 20 μm
Figure 10 shows the arithmetic means of intensities of RIPK1, RIPK3, CYLD and MLKL in the early- and middle-stage CL after local or systemic aPGF2α treatment. The intensity of RIPK1 was up-regulated after local aPGF2α injection in the early-stage CL (P < 0.0001; Fig. 10A). Moreover, local and systemic aPGF2α treatments increased its intensity in the middle-stage CL (P < 0.0001; Fig. 10A); however, this effect was greater when aPGF2α was injected systematically (P < 0.0001; Fig. 10A). Additionally, we observed higher intensity of RIPK1 in the middle-stage CL after systemic aPGF2α treatment compared with that in the early-stage CL (P = 0.0009; Fig. 10A). Furthermore, local aPGF2α injection increased the intensity of RIPK3 in the early- and middle-stage CL compared to the control group (P < 0.0001, P = 0.0030, respectively; Fig. 10B); while systemic aPGF2α treatment up-regulated the intensity of RIPK3 only in the middle-stage CL compared to the control group (P < 0.0001; Fig. 10B). Comparison of local to systemic administration of aPGF2α showed differences in RIPK3 intensity level in early- and middle-stage CL (P < 0.0001; Fig. 10B). Moreover, we observed higher RIPK3 intensity after local aPGF2α injection in the early-stage CL compare with that after local aPGF2α administration in the middle-stage CL (P < 0.0001; Fig. 10B) while RIPK3 intensity was higher after systemic aPGF2α treatment in the middle-stage CL compare to systemic injection in the early-stage CL (P < 0.0001; Fig. 10B). Systemic aPGF2α administration increased CYLD intensity (P = 0.0012, Fig. 10C). Comparison of systemic action of aPGF2α in both stages showed higher intensity of CYLD in the middle-stage CL (P = 0.0274; Fig. 10C). Moreover we obserwed higher CYLD intensity after systemic aPGF2α action compare to its intensity after local aPGF2α injection in the middle-stage CL (P = 0.0052; Fig. 10C). Futhermore, local and systemic aPGF2α treatments enhanced intensity of MLKL in the early- (P = 0.0048, P = 0.0081, respectively; Fig. 10D) and middle-stage CL (P < 0.0001; Fig. 10D). Comparison of local and systemic administration of aPGF2α showed differences in MLKL intensity in the middle-stage CL (P = 0.0016; Fig. 10D). Additionally, we observed higher MLKL intensity after local and systemic aPGF2α injection in the middle-stage CL compare with that after both aPGF2α administration in the early-stage CL (P < 0.0001; Fig. 10D).
Fig. 10.

The arithmetic means of intensities of: (a) RIPK1, (b) RIPK3, (c) CYLD and (d) MLKL in the bovine early- and middle-stage corpora lutea (CL) after local or systemic PGF2α analogue (aPGF2α) administration. The gray bars represent the control group, and the black bars represent local or systemic aPGF2α administered groups. Letters a,b,c indicate statistical differences between all experimental groups in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic aPGF2α injected early-stage CL vs local/systemic aPGF2α injected middle-stage CL
Discussion
Prostaglandin F2α is essential for manipulate bovine reproduction because in dairy cattle farming using of hormonal treatments are very common procedures to influence the estrous cycle. Until now, there have been no reports indicating a clear difference in PGF2α effects on cell-death mechanisms in the bovine CL with regard to its auto- or para-crine (local administration into the CL) vs. endocrine actions (systemic administration). Moreover, we confirmed in our previous in vivo and in vitro studies that during PGF2α-induced regression of the bovine CL, luteal cells are eliminated not only by apoptosis but also by programmed necrosis (RIPK-dependent necroptosis) [10, 11]. Futhermore, this is the first study describing details of a necroptotic pathway during PGF2α-induced luteal regression, showing up-regulation of CYLD and MLKL expression after PGF2α administration by both administration routes in the middle-stage CL. Therefore, we assume that better understanding of the effect of PGF2α on new mechanism involved in the CL regression in cows (necroptosis) may improve knowledge to optimize breeding methods of cows.
Prostaglandin F2α through its potent mediators plays a crucial role in regulation of the luteolytic cascade [21, 29], modulating numerous proteins associated with cell survival and cell death in different species [30]. It is well known that many factors are involved in PGF2α-induced luteolysis in cattle, including proinflammatory cytokines such as tumor necrosis factor α (TNF), interferon gamma (IFNG), Fas ligand (FASLG) [6, 31], endothelin 1 (EDN1) [32] and nitric oxide (NO) [28]. Moreover, communication between luteal and non-luteal cells is required for development and regression of the bovine CL [27, 33]. On the other hand, the lack of luteolytic effects of PGF2α in the bovine early CL may be associated with the absence of a well-established vascular system, despite the intensive angiogenesis occurring at this time [22, 34]. The early bovine CL is refractory to luteolytic actions of PGF2α in spite of the presence of PGF2α receptors [20]. However, the mechanism of insensitivity and acquisition to sensitivity of the CL to PGF2α is still not fully understood [23, 24, 26, 35].
The role of PGF2α in activating apoptotic signaling cascades in the CL during PGF2α- induced luteolysis has been previously examined [5]. The ratio of BCL2 to BAX expression levels is essential for cell survival or death [36–38]. In our study, we observed that the ratio of BCL2 to BAX mRNA expression levels was higher after systemic aPGF2α treatment compared to its local injection in the early-stage CL. In contrast, in the middle-stage CL this ratio of mRNA expression was markedly decreased after systemic PGF2α injection compared to its local administration. These findings suggest that PGF2α actions in the bovine CL depend on specific mediators participating in the progress of apoptosis by increasing BAX expression. Yadav et al. [39] found increased BAX and a constant mRNA and protein expressionof BCL2 in buffalo in the middle-stage CL 4 h after PGF2α treatment. In contrast, Kleim et al. [40] reported increased mRNA expression of BAX at 24 h after induced luteolysis, but in that study a different PGF2α analogue was used, which might explain the later increase in expression level.
In our study, we noticed that both PGF2α administration routes decreased CASP3 expression in the early CL, while increasing its expression in the middle-stage CL. Moreover, we showed that induction of CASP8 mRNA expression was responsive only to systemic PGF2α administration in the middle-stage CL. Additionally, there were no significant differences in CASP3 and CASP8 mRNA expression and protein concentration between local vs. systemic treatments with aPGF2α in both the early and middle-stage CL. These results correspond with the findings of other studies which reported that induction of CASP3 is an important factor in luteolysis, as well as the increase in CASP3 mRNA expression that occurs during PGF2α-induced luteolysis in the CL of different species [5, 8, 40].
Furthermore, one of the apoptosis initiators is CASP8 [41]. In the present study, we showed that induction of CASP8 mRNA expression and protein concentration was responsive only to systemic administration of PGF2α in the middle-stage CL. Therefore, we conclude that PGF2α is most potent as a luteolytic factor when it reaches the CL through the blood vasculature. It is well known that activated CASP8 promotes the apoptotic cascade by cleaving CASP3 [42]. Moreover, during the process of apoptosis accomplished via the mitochondrial pathway, active CASP8 regulates the binding of pro-apoptotic BAX to mitochondria and inhibits their connection with the antiapoptotic BCL2.
On the other hand, our present results indicate that either local or systemic action of PGF2α suppressed apoptotic signals via CASP3, and in parallel by affecting BCL2 and BAX expression in the early-stage CL. Therefore, PGF2α may exert an anti-apoptotic action on bovine luteal cells, thus playing a luteoprotective role in the early-stage CL. It is important to realise that CASP8 may be bound by some death effector domain (DED)-containing proteins such as cellular FLICE-like inhibitory protein (c-FLIP), which can inhibit apoptosis [43]. Our observations may explain why up-regulation of CASP8 mRNA expression in the early-stage CL was not followed by increased mRNA expression of effector CASP3. Importantly, the CASP8-cFLIP complex prevents not only apoptosis but also RIPK-induced necroptosis [44].
During the process of structural CL regression, not only apoptosis should be taken into consideration. Recently, Hojo et al. [10, 11] demonstrated that i.m. administration of PGF2α up-regulated both RIPK1 and RIPK3 expression in bovine CL cells and tissue in vivo and in vitro. In contrast to apoptosis, necroptosis occurs in the absence of CASPs activity (CASPs-independent programmed cell death). In our study, we investigated the regulatory mechanism of RIPK expression to clarify the mechanisms of necroptosis in the early and middle-stage CL in response to the local and systemic actions of PGF2α. We reported that expression of RIPK1 and RIPK3 mRNA and protein concentration was elevated after local and systemic PGF2α injections in the middle-stage CL. Moreover, it is clearly confirmed by immunohistochemistry analysis showing that intensities of RIPK1 and RIPK3 were up-regulated by both PGF2α treatments in the middle-stage CL. Therefore our results indicate that RIPK-dependent necroptosis is involved in aPGF2α-induced CL regression. Interestingly, we observed that systemic administration of PGF2α markedly up-regulated RIPK3 mRNA expression compared to its local action in the middle-stage CL. Based on the above results, we confirmed that PGF2α is a crucial luteolytic factor when administered systemically and the stimulatory effect of PGF2α on RIPKs expression may depend on different mediators or upon cell composition and cell contacts. Moreover, unlike apoptosis, necroptosis induces a more marked immune response that may function as a defensive mechanism [45]. A variety of cytokines produced by an increasing variety of local immune cells may be involved in the induction of luteal cell death processes in the bovine CL [3, 46].
In our study, systemic PGF2α injection may inhibit the necroptotic pathway by decreasing RIPK1 and RIPK3 mRNA expression in the early-stage CL. Additionally, we observed that local injection of PGF2α down-regulated RIPK3 mRNA expression, while oppositely affecting RIPK1 mRNA expression in the early-stage CL. Therefore, we suggest that formation of necrosomes does not occur in the early-stage CL. It has been reported that RIPK1 is a crucial mediator for RIPK3 activity and serves as a key mediator of cell death [18]. Therefore, we suspected that PGF2α through suppression of the death pathway in luteal cells may play a luteoprotective role in the early-stage CL. Bowolaksono et al. [47] suggested that PGF2α produced by bovine luteal cells inhibits apoptosis via stimulation of P4 in these cells, therefore luteal PGF2α is thought to be a luteoprotective factor [48].
To our knowledge, this is the first report showing the expression of CYLD and MLKL in the bovine CL. Interestingly, we observed higher protein concentration of CYLD and MLKL after systemic aPGF2α treatment compared to its local administration in the middle-stage CL. Additionally, immunohistochemistry analysis confirmed that intensity of CYLD was higher after systemic PGF2α treatment, while both administration of PGF2α affect intensity of MLKL in the middle-stage CL. It is known that CYLD is a key factor regulating cell survival and cell death, in a variety of ways including CASP8-mediated cell apoptosis and CASP8-independent cell necrosis [12, 49]. Moreover, MLKL is so far the most potent downstream effector of necroptosis that has been identified [50, 51]. The deubiquitination of RIPK1 by CYLD is critical for the activation of necroptosis and complex II formation [16]. In our study, we observed an increase in expression of the above-mentioned RIPKs and CYLD or MLKL, suggesting the activation of necroptosis and the formation of necrosomes during PGF2α-induced luteolysis. The above results are in agreement with previous studies carried out on different models based on selected human immune system cells [52] or embryonic fibroblasts from RIP3 knockout mice [53]. These authors emphasized the role of RIPK1, RIPK3 and MLKL as principal markers of TNF triggered necroptosis. Interestingly, in the present study mRNA expression of RIPK3 and the protein expression of CYLD and MLKL were higher after systemic administration of PGF2α compared to its local effect in the middle-stage CL. These findings may indicate that the systemic effect of PGF2α on the mechanism of cell death in the CL is more effective, depending on several auto/paracrine mediators activating luteolytic mechanisms, upon cell type and on cell-to-cell contact [33], and participation of the vascular system [22, 34, 54]. Therefore, we should take into consideration the fact that the distribution of capillaries is different during luteal development and regression [24, 54].
Conclusion
In conclusion, we have confirmed that PGF2α differentially modulates the expression of genes involved in apoptosis and necroptosis depending on the route of its administration (local vs. systemic), while local PGF2α plays a luteoprotective role by inhibiting necroptosis and apoptosis pathways in the early-stage CL. We confirmed that RIPK-dependent necroptosis is a potent mechanism involved in structural CL regression during PGF2α-induced luteolysis in cattle. Interestingly, the mechanism of the necroptotic pathway was evidently more affected by systemic PGF2α actions compared to its local impact during PGF2α-induced regression in the middle-stage CL, confirming that PGF2α influences CL function through auto/paracrine mediators.
Methods
Ethical authorization
The present authors ensured that their manuscript reported adheres to the arrive guidelines for the reporting of animal experiments. This statement address to their manuscript that these guidelines were followed: EU Directive of the European Parliament and the Council on the protection of animals used for scientific purposes (22 September 2010; no 2010/63/EU), the Polish Parliament Act on Animal Protection (21 August 1997, Dz.U. 1997 No 111 poz. 724) with further updates – the Polish Parliament Act on the protection of animals used for scientific or educational purposes (15 January 2015, Dz.U. 2015 pos. 266). All animal procedures were designed to avoid or minimize discomfort, distress and pain to the animals, moreover, were reviewed and accepted following the guidelines of the Local Ethics Committee for Experiments on Animals in Olsztyn, Poland (Approval no 23/2012/N).
Animals and treatments
For the present study, 48 healthy, cycling Polish Holstein-Friesian cows from a local commercial dairy farm were used. The history of the cows and the structure of the farms were investigated by a questionnaire for the owners. Written owner consent was available through farm manager. This study was conducted from May 2018 to December 2018. The cows were bred by artificial insemination with a standard, routine protocol. The farm was monitored by trained veterinary and nutrition consultants and was free of Bovine Herpesvirus Type 1 (BHV1), Bovine Viral Diarrhea-Mucosal Disease (BVD/MD virus), Enzootic bovine leukosis (EBL) and tuberculosis. The experiment was performed in a group of non-pregnant cows (n = 48; 612 ± 97 kg; 3 to 5 parities; aged 5–7 years) and that were considered for culling because of their low milk production. The experimental cows were housed in an indoor facility in free-stall barns, were milked on a 12 h cycle, and fed with a TMR to meet the nutritional requirements of milking cows (15–20 kg/day) with ad libitum access to water and a salt-based mineral supplement. Prior to the experiment, an experienced veterinarian confirmed the absence of reproductive tract disorders by an ultrasonographic visualization (USG) per rectum with a 7.5 MHz linear array transducer (MyLab 30VET Gold Colour Doppler Diagnostic Ultrasound System, ESOATE Pie Medica, Genoa, Italy). Moreover, all the experimental cows underwent a general clinical examination in which rectal temperature (38.0–39.2 °C), general attitude (healthy), respiratory rates (27–30 breaths per minute), heart rates (60–82 beats per minute) and BCS (3.0 ± 0.5) were determined. The estrous cycle was synchronized in all cows by two injections of aPGF2α (Dinoprost, 25 mg/5 ml; Dinolytic; Zoetis, Poland) with an 11-days interval, as reported previously [55]. Follicular development and structural changes of the CL during the entire estrous cycle were monitored using transrectal USG, and visible signs of estrus (i.e., vaginal mucus and standing behavior) were taken as its confirmation. The onset of estrus was considered as day 0 of the estrous cycle. Additionally, the stage of the estrous cycle was established by P4 concentrations in blood plasma samples collected from the coccygeal vessels using radioimmunoassay (RIA). The concentration of P4 was 0.38 ± 0.09 ng/ml (mean ± SEM) in blood samples collected during estrus (day 0 of the estrous cycle). After our in vivo study the experimental cows (n = 48) were slaughtered in local abbatoir due to farmer’s breeding and management program.
In vivo study design
The cows were divided into two groups depending on the phase of the estrous cycle: group I (early luteal phase, day 4 of the estrous cycle; n = 24) and group II (day 10, mid-luteal phase; n = 24). The concentration of P4 was 2.98 ± 0.46 ng/ml or 9.54 ± 0.28 ng/ml (mean ± SEM) in blood samples collected from cows in group I or II, respectively. Afterwards, the cows were treated as follows: (1) i.m. (systemic) sterile 0.9% saline solution injection (control; n = 6), (2) systemic aPGF2α injection (25 mg/5 ml Dinoprost; Dinolytic, Zoetis, Poland; n = 6), (3) intra-CL (local) saline injection (control; n = 6), and (4) local aPGF2α injection (2.5 mg/0.5 ml Dinoprost; n = 6). Figure 11 shows in vivo study design. The dose of aPGF2α for intra-CL injection was established in our previous study [56]. The time of injections of saline solution or aPGF2α was defined as hour ‘0’. Before intra-CL injections, the animals were premedicated with xylazine (i.m. 25–30 mg/animal; Xylavet 2%; ScanVet, Poland). Then, the cows were anesthetized via an epidural block using 4 ml of 2% procaine hydrochloride (Polocainum Hydrochloricum; Biowet Drwalew, Poland). Then, intra-CL injections were administered under ultrasound guidance through a sterile 1.25 × 50 mm (18 G × 2″). Ovum Pick-up disposable veterinary injection needle (BOVIVET, Poland). The transducer and needle guide were coated with a sterile lubricant (Medicum, Poland) and positioned within the vagina. We perform intra-CL injection with USG guided ovum pick-up system in cattle. The ovary bearing the CL was positioned rectally to visualize it. The needle was then passed through the vaginal wall, and the aPGF2α or saline was injected directly into the CL. Each disposable catheter was filled with 0,5 mL saline/ dinoprost. Moreover, the injected substance was observed by USG as a white shade on the monitor and it was seen to diffuse within the CL. Four hours after each treatment, the ovaries with CL were collected by colpotomy using a Hauptner’s efeminator (Hauptner & Herberholz GmbH & Co. KG, Solingen, Germany). Ovary collection was described previously by Piotrowska et al. [57]. To avoid or minimize discomfort, distress and pain to the animals during the in vivo study, all experimental cows were kept in a barn as the separate group. Moreover, during the experimental day, the cows were fed with grass hay and were given free access to water. After experimental procedures cows were put into 24 h observation and quarantine and return to farmer’s breeding and management program. After that cows were slaughtered in local abbatoir due to farmer’s breeding and management program.
Fig. 11.

Schematic diagram of the study design. Cows were synchronized via two injections of aPGF2α administered with an 11-days interval, starting on protocol day − 14. The onset of estrus was considered as day 0 of the estrous cycle. On day 4 or 10 respectively, the cows received an local injection of saline/ aPGF2α (2.5 mg Dinoprost/0.5 ml) or systemic injection of saline/ aPGF2α (25 mg Dinoprost/5 ml). At 4 h after treatment the cows were ovariectomized. CLs were collected for gene and protein expression
Each CL tissue was divided into three parts. The tissue was immediately placed into a 1.5 ml microcentrifuge tube containing either 1 ml RNALater (#R0901, Sigma Aldrich, Germany) or which was empty, immediately homogenized, and stored at − 80 °C. mRNA and protein expression of apoptosis - or necroptosis-related factors in CL tissues after local and systemic injections of saline or aPGF2α were examined by RT-qPCR and western blotting, respectively. For immunohistochemistry, the third part of the CL tissue was fixed in 4% (vol/vol) neutral formalin (pH 7.4) for 20–24 h and then embedded in paraffin wax.
RNA extraction and cDNA production
Total RNA was extracted from CL tissues (40 ± 5 mg) using the Total RNA Mini (#031–100, A&A Biotechnology, Poland) according to the manufacturer’s instructions. The content and purity of RNA was assessed on a NanoDrop 1000 (Thermo Fisher Scientific, ND-1000, Wilmington, DE, USA). The 260/280 absorbance ratio for all samples was approx. 2.0, and the 260/230 absorbance ratio ranged between 1.8–2.2. Then, 1 μg RNA was reverse-transcribed into cDNA using a QuantiTect Reverse Transcription Kit (#205311, Qiagen, Germany) according to the manufacturer’s instructions. The cDNA was stored at − 20 °C until Reverse transcriptional PCR or RT-q PCR was carried out.
RT-qPCR
RT-qPCR assays were performed in an ABI 7900 HT sequence detection system using SYBR Green PCR master mix (Applied Biosystems, Foster City, CA, USA). For the examined genes, samples (n = 48) were run in duplicates. Primer sequences used for determination of CASP3, CASP8, BAX, BCL2, RIPK1, RIPK3, CYLD, MLKL, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), actin beta (ACTB) and 18S ribosomal RNA (RN18S) mRNA expression are detailed in Table 1. All primers were designed using Primer-BLAST and synthesized by Sigma (Custom Oligos Sigma Aldrich). The stability of the reference genes was determined in the NormFinder program as previously described by Andersen et al. [58]. Gene expression data are expressed relative to the best combination of two housekeeping genes and are presented as arbitrary units. In our study gene expression is expressed as a ratio of target genes to ACTB/RN18S1. Total reaction volume was 10 μl containing: 3 μl cDNA (10 ng), 1 μl forward and reverse primers each (500 nM) and 5 μl SYBR Green PCR master-mix. RT-qPCR was carried out as follows: initial denaturation (10 min at 95 °C), followed by 45 cycles of denaturation (15 s at 95 °C) and annealing (1 min at 60 °C). After each PCR reaction, melting curves were obtained by stepwise increases in temperature from 60 °C to 95 °C to ensure single product amplification. Specificity of the product was confirmed by electrophoresis on 2% agarose gel. RT-qPCR results were analyzed using the method described by Zhao & Fernald [59].
Table 1.
Sequences for primers and accession numbers for genes
| Gene name | Sequence of nucleotide | GenBank Accession No. |
PCR products size |
|---|---|---|---|
| CASP3 |
5′ TGGTGCTGAGGATGACATGG ‘3 5′ GAGCCTGTGAGCGTGCTTTT’3 |
NM_001077840.1 | 163 bp |
| CASP8 |
5′ CTGAGAGAAGAGGCCCGTGA ‘3 5′ CCCGGCTTAGGAACTTGAGG ‘3 |
DQ319070.1 | 173 bp |
| BAX |
5′ GTGCCCGAGTTGATCAGGAC ‘3 5′ CCATGTGGGTGTCCCAAAGT ‘3 |
U92569.1 | 126 bp |
| BCL2 |
5′ GAGTTCGGAGGGGTCATGTG ‘3 5′ GCCTTCAGAGACAGCCAGGA ‘3 |
U92434.1 | 203 bp |
| RIPK1 |
5′ GCAATAGCTCCAAGCAGGTC ‘3 5′ TGTGCAGCAGGAAGTCATTC ‘3 |
NM_001035012 | 148 bp |
| RIPK3 |
5′ CCAGAGAGAGCAGGTTCCAC ‘3 5′ AATCAGGCGGTTGTTGTTTC ‘3 |
NM_001101884.2 | 219 bp |
| CYLD |
5′ GCAATGCTCAAGTCCACCT ‘3 5′ CGTGTCTCCAGTCCCAGTC ‘3 |
XM_015475764.2 | 96 bp |
| MLKL |
5′ ACTTCCATCAGCCGACAAAC ‘3 5′ CTCCCAGAGGACAATTCCAA ‘3 |
XM_024978879.1 | 144 bp |
| GAPDH |
5′ CACCCTCAAGATTGTCAGCA ‘3 5′ GGTCATAAGTCCCTCCACGA ‘3 |
BC102589 | 103 bp |
| ACTB |
5′ GAGGATCTTCATGAGGTAGTCTGTCAGG ‘3 5′ CAACTGGGACGACATGGAGAAGATCTGGCA ‘3 |
AY141970 | 349 bp |
| RN18S1 |
5′ AAGTCTTTGGGTTCCGGG ‘3 5’GGACATCTAAGGGCATCACA’3 |
AF176811 | 365 bp |
Western blotting
Protein expression levels for CASP3, CASP8, BAX, BCL2, RIPK1, RIPK3, CYLD and MLKL and ACTB in the CL tissues (n = 48, each sample weight 100 mg) were determined by Western blotting as previously described [10]. Specific antibodies are described in detail in Table 2. Protocols for overnight incubation were used following dilution of each antibody at 4 °C (Table 2). Subsequently, membranes were incubated with a 1:20,000 dilution of secondary polyclonal anti-rabbit IgG or anti-mouse IgG alkaline phosphatase-conjugated antibodies (#S3687, #S3562, Sigma Aldrich, Germany) for 1.5 h at room temperature (RT). Immune complexes were detected using the alkaline phosphatase visualization procedure. Each sample was checked to evaluate the intensity of immunological reactions by measuring the optical density in the defined area with computerized densitometry via NIH Image (National Institutes of Health, Bethesda, MD, USA). The protein concentration profiles are presented in arbitrary units as the ratio of the test proteins to the reference protein – ACTB. Representative western blot bands for CASP3, CASP8, BAX, BCL2, RIPK1, RIPK3, CYLD and MLKL and ACTB are shown in Additional files 1 and 2.
Table 2.
Specific antibodies used for Western immunoblotting and Immmunohistochemistry
| Antibodies | Clone | Biological source | Commercial source | Dilution for WB | Dilution for IHC |
|---|---|---|---|---|---|
| Anti-ACTB | monoclonal | mouse | Sigma, #A2228 | 1:10.000 | |
| Anti-CASP3 | polyclonal | rabbit | Abcam, ab44976 | 1:500 | |
| Anti-CASP8 | monoclonal | rabbit | Abcam, ab108333 | 1:1.000 | |
| Anti-BAX | polyclonal | rabbit | Abcam, ab53154 | 1:500 | |
| Anti-BCL2 | polyclonal | rabbit | Abcam, ab59348 | 1:500 | |
| Anti-RIPK1 | polyclonal | rabbit | Sigma, #SAB3500420 | 1:1.000 | 1:200 |
| Anti-RIPK3 | polyclonal | rabbit | Sigma, #SAB2102009 | 1:1.000 | 1:100 |
| Anti-CYLD | polyclonal | rabbit | Abcam, 137,524 | 1:500 | 1:100 |
| Anti-MLKL | monoclonal | rabbit | Abcam, ab187091 | 1:1.000 | 1:100 |
Sigma Aldrich, Germany
Abcam, United Kingdom
Immmunohistochemistry
After dewaxing and washing, paraffin-embedded sections, cut at 4-μm thickness, were incubated at RT with 0.3% hydrogen peroxide in methanol for 20 min to inactivate endogenous peroxidase. Then, the sections were washed in PBS and incubated with normal goat serum for 60 min at RT followed by RIPK1, RIPK3, CYLD or MLKL antibodies at 4 °C overnight. Specific antibodies and their dilutions were described in Table 2. After washing twice, the sections were incubated with biotinylated anti-rabbit IgG (1:500; #PK-6101, Vector Laboratories, CA, USA) for 60 min at RT. The reaction sites were visualized using 3,3 Diaminobenzidine tetrahydrochloride, TLC approx. 97% (DAB) (#D5637, Sigma Aldrich, Germany) for 5 min. The sections were counterstained for 2 min with hematoxylin. Positive immunohistochemistry staining was assessed as a characteristic brown staining using a light microscope (Zeiss Imager. Z1; Zeiss, Germany). For negative controls, primary antibodies were excluded and samples were incubated with rabbit IgG. All signals were visualized by Zeiss Axio Observer System (Carl Zeiss, Germany) using 25/0.8 NA or 63/1.3 NA immerse objectives. Images were converted to 16-bit, grayscale version and analyzed using by Zeiss Axio Observer System (Carl Zeiss, Germany) software. For quantification arithmetic means of all intensities of immunostaining were measured as optical desnity and evaluated in correlation to the signal of the control group. All values are expressed as an n-fold increase.
Statistical analysis
The statistical analyses of the results of mRNA expression (n = 48 samples) and protein concentration and intensity (n = 48 samples) were performed using two way ANOVA followed by Sidak multiple comparison test (GraphPad Prism ver. 8.2.1; Graph Pad Software, San Diego, CA, USA). All numerical data are shown as standard errors of the means (± SEM) for values obtained in our experiment (six samples/treatment), each performed in duplicates. Letters a,b,c indicate statistical differences between treatment in the early and middle- stage CL. Asteriks * indicate statistical differences between local/systemic PGF2α injected early CL vs local/systemic PGF2α injected middle-stage CL.
Supplementary information
Additional file 1. Representative western blots bands for (a) CASP3 (32 kDa), CASP8 (18 kDa), BCL2 (26 kDa), BAX (74 kDa) and ACTB (43 kDa) (b) RIPK1 (74 kDa), RIPK3 (57 kDa), CYLD (107 kDa), MLKL (54 kDa) and ACTB (43 kDa) in the early CL at 4 h after local or systemic PGF2α administration; C – control group, PGF2α − experimental group respectively local (2.5 mg Dinoprost intra-CL) or systemic PGF2α injection (25 mg Dinoprost i.m.)
Additional file 2. Representative western blots bands for (a) CASP3 (32 kDa), CASP8 (18 kDa), BCL2 (26 kDa), BAX (74 kDa) and ACTB (43 kDa) (b) RIPK1 (74 kDa), RIPK3 (57 kDa), CYLD (107 kDa), MLKL (54 kDa) and ACTB (43 kDa) in the mid-stage CL at 4 h after local or systemic PGF2α administrations; C – control group, PGF2α − experimental group respectively local (2.5 mg Dinoprost intra-CL) or systemic PGF2α injection (25 mg Dinoprost i.m.).
Acknowledgements
The authors are grateful Dr. M M Bah for his assistance and cooperation in providing animals for the study from the farm in Wolka Szlachecka. The authors wish to thank K Jankowska and W Krzywiec for technical support in the experiments. The authors wish to thank K Witek for immunohistochemistry pictures analysis.
Authors’information
AWJ – Ph.D. Student / Assistant Professors in the Department of Reproductive Immunology and Pathology in the Institute of Animal Reproduction and Food Research Polish Academy of Science. KKPT – DVM, Ph.D.; Assistant Professors in the Department of Reproductive Immunology and Pathology in the Institute of Animal Reproduction and Food Research Polish Academy of Science. DJS - DVM, Ph.D., Dr.Sci., Head of Department of Reproductive Immunology and Pathology in the Institute of Animal Reproduction and Food Research Polish Academy of Science.
Abbreviations
- aPGF2α
Prostaglandin F2α analogue
- ANOVA
Analysis of variance
- BAX
BCL2 associated X
- BCL2
B-cell lymphoma 2
- BCS
Body condition score
- BHV1
Bovine Herpesvirus Type 1
- BVD/MD
Bovine Viral Diarrhea-Mucosal Disease
- CASP3
Caspase 3
- CASP8
Caspase 8
- cDNA
Complementary DNA
- CL
Corpus luteum
- CYLD
cylindromatosis
- DAB
3,3 Diaminobenzidine tetrahydrochloride
- DUB
K63-specific deubiquitinating enzyme
- EBL
Enzootic bovine leukosis
- EDN1
Endothelin 1
- FASLG
Fas ligand
- i.m.
intra muscular
- IFNG
Interferon gamma
- LEC
Luteal endothelial cells
- LSC
Luteal steroidogenic cells
- MLKL
mixed lineage kinase domain-like
- mRNA
messenger RNA
- NO
Nitric oxide
- P4
Progesterone
- PBS
Phosphate-buffered saline
- PCR
Polymerase chain reaction
- PGF2α
Prostaglandin F2α
- RIA
Radioimmunoassay
- RIPK1
receptor-interacting protein kinase 1
- RIPK3
receptor-interacting protein kinase 3
- RT
Room temperature
- TMR
Total mix ration
- TNF
Tumor necrosis factor α
- USG
ultrasonographic visualization
Authors’ contributions
DJS and KKPT designed the experiments; AWJ, KKPT, DJS performed the experiments and analyses; AWJ, KKPT and DJS analyzed the results; AWJ and KKPT wrote the manuscript. This research is a part of a PhD thesis conducted by AWJ. All authors have read and approved the manuscript.
Funding
This research was supported by the National Science Centre projects 2017/X/NZ9/00363 (research on potential mechanisms of cell-death pathways in the mid CL, day 10 of the estrous cycle); 2018/29/B/NZ9/00391 (research on the comparison of mechanisms controlling luteal cell viability in the developing CL, day 4 of the estrous cycle). The Polish National Science Centre was the founder of the research and did not participate in the design of the study and collection, analysis, and interpretation of data as well as writing the manuscript.
Availability of data and materials
The datasets used and analysed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
All procedures performed were in accordance with the ethical standards of the institution and practice. All animal procedures were reviewed and accepted following the guidelines of the Local Ethics Committee for Experiments on Animals in Olsztyn, Poland (Approval no 23/2012/N). Moreover, written owner consent was available through farm manager.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information accompanies this paper at 10.1186/s12917-019-2167-3.
References
- 1.Niswender GD, Juengel JL, Silva PJ, Rollyson MK, McIntush EW. Mechanisms controlling the function and life span of the corpus luteum. Physiol Rev. 2000. 10.1152/physrev.2000.80.1.1. [DOI] [PubMed]
- 2.McCracken JA, Custer EE, Lamsa JC. Luteolysis: a neeuroendocrine-mediated event. Physiol Rev. 1999. 10.1152/physrev.1999.79.2.263. [DOI] [PubMed]
- 3.Miyamoto A, Shirasuna K, Shimizu T, Bollwein H, Schams D. Regulation of corpus luteum development and maintenance: specific roles of angiogenesis and action of prostaglandin F2alpha. Soc Reprod Fertil Suppl. 2010;67:289–304. doi: 10.7313/upo9781907284991.024. [DOI] [PubMed] [Google Scholar]
- 4.Juengel JL, Garverick HA, Johnson AL, Youngquist RS, Smith MF. Apoptosis during luteal regression in cattle. Endocrinology. 1993. 10.1210/endo.132.1.8419126. [DOI] [PubMed]
- 5.Yadav VK, Lakshmi G , Medhamurthy R. Prostaglandin F2α-mediated activation of apoptotic signaling cascades in the corpus luteum during apoptosis: involvement of caspase-activated DNase. J Biol Chem. 2005;doi: 10.1074/jbc. M409596200. [DOI] [PubMed]
- 6.Davis JS, Rueda BR. The corpus luteum: an ovarian structure with maternal instincts and suicidal tendencies. Front Biosci. 2002;7:1949–1978. doi: 10.2741/davis1. [DOI] [PubMed] [Google Scholar]
- 7.Hojo T, Oda A, Lee SH, Acosta TJ, Okuda K. Effects of tumor necrosis factor α and interferon γ on the viability and mRNA expression of TNF receptor type I in endothelial cells from the bovine corpus luteum. J Reprod Dev. 2010. 10.1262/jrd.10-056t. [DOI] [PubMed]
- 8.Pru JK, Tilly JL. Programmed cell death in the ovary: insights and future prospects using genetic technologies. Mol Endocrinol. 20014; doi:10.1210/mend.15.6.0646. [DOI] [PubMed]
- 9.Carambula SF, Matikainen T, Lynch MP, Flavell RA, Dias Gonçalves PB, Tilly JL, Rueda BR. Caspase-3 is a pivotal mediator of apoptosis during regression of the ovarian corpus luteum. Endocrinol. 2002. 10.1210/endo.143.4.8726. [DOI] [PubMed]
- 10.Hojo T, Siemieniuch MJ, Lukasik K, Piotrowska-Tomala KK, Jonczyk AW, Okuda K, Skarzynski DJ. Programmed necrosis - a new mechanism of steroidogenic luteal cell death and elimination during luteolysis in cows. Sci Rep. 2016. 10.1038/srep38211. [DOI] [PMC free article] [PubMed]
- 11.Hojo T, Piotrowska-Tomala KK, Jonczyk AW, Lukasik K, Jankowska K, Okuda K, Witek KJ, Skarzynski DJ. Receptor interacting protein kinases-dependent necroptosis as a new, potent mechanism for elimination of the endothelial cells during luteolysis in cow. Theriogenology. 2019. 10.1016/j.theriogenology.2019.01.035. [DOI] [PubMed]
- 12.Zhou W, Yuan J. Necroptosis in health and diseases. Semin Cell Dev Biol. 2014. 10.1016/j.semcdb.2014.07.013. [DOI] [PubMed]
- 13.Sun L, Wang X. A new kind of cel suicide: mechanisms and functions of programmed necrosis. Trends Biochem Sci. 2014. 10.1016/j.tibs.2014.10.003. [DOI] [PubMed]
- 14.Holler N, Zaru R, Micheau O, Thome M, Attinger A, Valitutti S, Bodmer JL, Schneider P, Seed B, Tschopp J. Fas triggers an alternative, caspase-8-independent cell death pathway using the kinase RIP as effector molecule. Nat Immunol. 2000. 10.1038/82732. [DOI] [PubMed]
- 15.Cho YS, Challa S, Moquin D, Genga R, Ray TD, Guildford M, Chan FK. Phosphorylation-driven assembly of the RIP1–RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell. 2009. 10.1016/j.cell.2009.05.037. [DOI] [PMC free article] [PubMed]
- 16.Moquin DM, McQuade T, Chan FK. CYLD Deubiquitinates RIP1 in the TNFα-induced Necrosome to facilitate kinase activation and programmed necrosis PLoS one. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhao J, Jitkaew S, Cai Z, Choksi S, Li Q, Luo J, Liu ZG. Mixed lineage kinase domain-like is a key receptor interacting protein 3 downstream component of TNF-induced necrosis. Proc Natl Acad Sci U S A. 2012. 10.1073/pnas.1200012109. [DOI] [PMC free article] [PubMed]
- 18.Zhang DW, Shao J, Lin J, Zhang N, Lu BJ, Lin SC. RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to necrosis. Science. 2009. 10.1126/science.1172308. [DOI] [PubMed]
- 19.Lauderdale JW. ASAS centennial paper: conttributions in the journal of animal science to the development of protocols for breeding management of cattle through synchronization of estrus and ovulation. J Anim Sci. 2009. 10.2527/jas.2008-1407. [DOI] [PubMed]
- 20.Levy N, Kobayashi S, Roth Z, Wolfenson D, Miyamoto A, Meidan R. Administration of prostaglandin F2α during the early bovine luteal phase does not alter the expression of ET-1 and of its type a receptor: a possible cause for corpus luteum refractoriness. Biol Reprod. 2000. 10.1095/biolreprod63.2.377. [DOI] [PubMed]
- 21.Skarzynski DJ, Jaroszewski JJ, Bah MM, Deptula KM, Barszczewska B, Gawronska B, Hansel W. Administration of a nitric oxide synthase inhibitor counteracts prostaglandin F2-induced luteolysis in cattle. Biol Reprod. 2003. 10.1095/biolreprod.102.008573. [DOI] [PubMed]
- 22.Skarzynski DJ, Piotrowska-Tomala KK, Lukasik K, Galvão A, Farberov S, Zalman Y, Meidan R. Growth and regression in bovine corpora lutea: regulation by local survival and death pathways. Reprod Domest Anim. 2013. 10.1111/rda.12203. [DOI] [PubMed]
- 23.Mondal M, Schilling B, Folger J, Steibel JP, Buchnick H, Zalman Y, Ireland JJ, Meidan R, Smith GW. 2011 deciphering the luteal transcriptome: potential mechanisms mediating stage-specific luteolytic response of the corpus luteum to prostaglandin F2α. Physiol Genomics. 2011. 10.1152/physiolgenomics.00155.2010. [DOI] [PubMed]
- 24.Shirasuna K, Sasahara K, Matsui M, Shimizu T, Miyamoto A. Prostaglandin F2alpha differentially affects mRNA expression relating to angiogenesis, vasoactivation and prostaglandins in the early and mid corpus luteum in the cow. J Reprod Dev. 2010. 10.1262/jrd.10-004o. [DOI] [PubMed]
- 25.Atli MO, Bender RW, Mehta V, Bastos MR, Luo WX, Vezina CM. WiltbankMC, Patterns of gene expression in the bovine corpus luteum following repeated intrauterine infusions of low doses of prostaglandin F2alpha. Biol Reprod. 2012. 10.1095/biolreprod.111.094870. [DOI] [PMC free article] [PubMed]
- 26.Zalman Y, Klipper E, Farberov S, Mondal M, Wee G, Folger JK, Smith GW, Meidan R. Regulation of angiogenesis-related prostaglandin f2alpha-induced genes in the bovine corpus luteum. Biol Reprod. 2012. 10.1095/biolreprod.111.095067. [DOI] [PubMed]
- 27.Pate JL. Involvement of immune cells in regulation of ovarian function. J Reprod Fertil Suppl. 1995;49:365–377. [PubMed] [Google Scholar]
- 28.Korzekwa A, Okuda K, Woclawek-Potocka I, Murakami S, Skarzynski DJ. Nitric oxide induces apoptosis in bovine luteal cells. J Reprod Dev. 2006. 10.1262/jrd.17092. [DOI] [PubMed]
- 29.Acosta TJ, Bah MM, Korzekwa A, Woclawek-Potocka I, Markiewicz W, Jaroszewski JJ, Okuda K, Skarzynski DJ. Acute changes in circulating concentrations of progesterone and nitric oxide and partial pressure of oxygen during PGF2α – induced luteolysis in cattle. J Reprod Dev. 2009. 10.1262/jrd.20133. [DOI] [PubMed]
- 30.Meidan R, Girsh E, Mamluk R, Levy N, Farberov S. The Life Cycle of the Corpus Luteum. Switzerland: Springer International Publishing; 2017. Luteolysis in Ruminants: Past Concepts, New Insights, and Persisting Challenges; pp. 159–182. [Google Scholar]
- 31.Okuda K, Miyamoto Y, Skarzynski DJ. Regulation of endometrial prostaglandin F (2alpha) synthesis during luteolysis and early pregnancy in cattle. Domest Anim Endocrinol. 2002;23(1–2):255–264. doi: 10.1016/S0739-7240(02)00161-3. [DOI] [PubMed] [Google Scholar]
- 32.Watanebe S, Shirasuna K, Matsui M, Yamamoto D, Berisha B, Schams D, Miyamoto A. Effect of intraluteal injection of endothelin type a receptor antagonist on PGF2α-induced luteolysis in the cow. J Reprod Dev. 2006. 10.1262/jrd.18018. [DOI] [PubMed]
- 33.Korzekwa AJ, Jaroszewski JJ, Woclawek-Potocka I, Bah MM, Skarzynski DJ. Luteolytic effect of prostaglandin F 2 alpha on bovine corpus luteum depends on cell composition and contact. Reprod Domest Anim. 2008. 10.1111/j.1439-0531.2007.00936.x. [DOI] [PubMed]
- 34.Acosta TJ, Hayashi KG, Ohtani M, Miyamoto A. Local changes in blood flow within the preovulatory follicle wall and early corpus luteum in cows. Reprod. 2003;125(5):759–767. doi: 10.1530/rep.0.1250759. [DOI] [PubMed] [Google Scholar]
- 35.Goravanahally MP, Salem M, Yao J, Inskeep EK, Flores JA. Differential gene expression in the bovine corpus luteum during transition from early phase to midphase and its potential role in acquisition of luteolytic sensitivity to prostaglandin F2alpha. Biol Reprod. 2009. 10.1095/biolreprod.108.069518. [DOI] [PubMed]
- 36.Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell. 1993. 10.1016/0092-8674(93)90509-o. [DOI] [PubMed]
- 37.Williams GT, Smith CA. Molecular regulation of apoptosis: genetic controls on cell death. Cell. 1993. 10.1016/0092-8674(93)90457-2. [DOI] [PubMed]
- 38.Antonsson B. Bax and other pro-apoptotic Bcl-2 family “killer-proteins” and their victim the mitochondrion. Cell Tissue Res. 2001. 10.1007/s00441-001-0472-0. [DOI] [PubMed]
- 39.Kleim H, Berisha B, Meyer HHD, Schams D. Regulatory changes of apoptotic factors in the bovine corpus luteum after induced luteolysis. Mol Reprod Dev. 2009. 10.1002/mrd.20946. [DOI] [PubMed]
- 40.Yadav VK, Sudhagar RR, Medhamurthy R. Apoptosis during spontaneous and prostaglandin F-2alpha-induced luteal regression in the Buffalo cow (Bubalus bubalis): involvement of mitogen-activated protein kinases. Biol Reprod. 2002. 10.1095/biolreprod.102.004077. [DOI] [PubMed]
- 41.Siegel RM. Caspases at the crossroads of immune-cell life and death. Nat Rev Immunol. 2006. 10.1038/nri1809. [DOI] [PubMed]
- 42.Budihardjo I, Oliver H, Lutter M, Luo X, Wang X. Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell Biol. 1999. 10.1146/annurev.cellbio.15.1.269. [DOI] [PubMed]
- 43.Budd RC, Yeh WC, Tschopp J. cFLIP regulation of lymphocyte activation and development. Nat Rev Immunol. 2006. 10.1038/nri1787. [DOI] [PubMed]
- 44.Djerbi M, Darreh-Shori T, Zhivotovsky B, Grandien A. A characterization of the human FLICE- inhibitory protein locus and comparison of the anti-apoptotic activity of four different flip forms. Scand J Immunol. 2001;54:180–189. doi: 10.1046/j.1365-3083.2001.00941.x. [DOI] [PubMed] [Google Scholar]
- 45.Chen D, Yu J, Zhang L. Necroptosis: an alternative cell death program defending against cancer. Biochim Biophys Acta. 2016. 10.1016/j.bbcan.2016.03.003. [DOI] [PMC free article] [PubMed]
- 46.Friedman A, Weiss S, Levy N, Meidan R. Role of tumor necrosis factor alpha and its type I receptor in luteal regression: induction of programmed cell death in bovine corpus luteum-derived endothelial cells. Biol Reprod. 2000. 10.1095/biolreprod63.6.1905. [DOI] [PubMed]
- 47.Bowolaksono A, Nishimura R, Hojo T, Sakumoto R, Acosta TJ, Okuda K. Anti-apoptotic roles of prostaglandin E2 and F2alpha in bovine luteal steroidogenic cells. Biol Reprod. 2008;79:310–317. doi: 10.1095/biolreprod.107.066084. [DOI] [PubMed] [Google Scholar]
- 48.Schams D. Berisha B. Regulation of corpus luteum function in cattle-an overview Reprod Domest Anim. 2004. 10.1111/j.1439-0531.2004.00509.x. [DOI] [PubMed]
- 49.Hitomi J, Christofferson DE, Ng A, Yao J, Degterev A, Xavier RJ. Identification of a molecular signaling network that regulates a cellular necrotic cell death pathway. Cell. 2008. 10.1016/j.cell.2008.10.044. [DOI] [PMC free article] [PubMed]
- 50.Wu J, Huang Z, Ren J, Zhang Z. He P, Li Y. Mlkl knockout mice demonstrate the indispensable role of MLKL in necroptosis Cell Res. 2013. 10.1038/cr.2013.91. [DOI] [PMC free article] [PubMed]
- 51.Jouan-Lanhouet S, Riquet F, Duprez L, Vanden Berghe T, Takahashi N, Vandenabeele P. Necroptosis, in vivo detection in experimental disease models. Semin Cell Dev Biol. 2014. 10.1016/j.semcdb.2014.08.010. [DOI] [PubMed]
- 52.Cai Z, Jitkaew S, Zhao J, Chiang HC. Choksi S, Liu J. Plasma membrane translocation of trimerized MLKL protein is required for TNF-induced necroptosis Nat Cell Biol. 2014. 10.1038/ncb2883. [DOI] [PMC free article] [PubMed]
- 53.He S, Huang S, Shen Z. Biomarkers for the detection of necroptosis. Cell Mol Life Sci. 2016. 10.1007/s00018-016-2192-3. [DOI] [PMC free article] [PubMed]
- 54.Acosta TJ, Yoshizawa N, Ohtani M, Miyamoto A. Local changes in blood flow within the early and midcycle corpus luteum after prostaglandin F (2 alpha) injection in the cow. Biol Reprod. 2002;66:651–658. doi: 10.1095/biolreprod66.3.651. [DOI] [PubMed] [Google Scholar]
- 55.Skarzynski DJ, Piotrowska KK, Bah MM, Korzekwa A, Woclawek-Potocka I, Sawai K, Okuda K. Effects of exogenous tumour necrosis factor-alpha on the secretory function of the bovine reproductive tract depend on tumour necrosis factor-alpha concentrations. Reprod Domest Anim. 2009. 10.1111/j.1439-0531.2007.01016.x. [DOI] [PubMed]
- 56.Jonczyk AW, Piotrowska-Tomala KK, Kordowitzki P, Skarzynski DJ. Effects of prostaglandin F2α on angiogenic and steroidogenic pathways in the bovine corpus luteum may depend on its route of administration. J Dairy Sci. 2019 Sep 11. pii: S0022–0302(19)30802–1; doi: 10.3168/jds.2019-16644. [Epub ahead of print]. [DOI] [PubMed]
- 57.Piotrowska KK, Wocławek-Potocka I, Bah MM, Piskula M, Pilawski W, Bober A, Skarzynski DJ. Phytoestrogens and their metabolites inhibit the sensitivity of bovine CL on luteotropic factors. J Reprod Dev. 2006. 10.1262/jrd.17054. [DOI] [PubMed]
- 58.Andersen CL, Jensen JL, Ørntoft TF. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004. 10.1158/0008-5472.CAN-04-0496. [DOI] [PubMed]
- 59.Zhao H, Fernald RD. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J Comput Biol. 2005. 10.1089/cmb.2005.12.1047. [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Representative western blots bands for (a) CASP3 (32 kDa), CASP8 (18 kDa), BCL2 (26 kDa), BAX (74 kDa) and ACTB (43 kDa) (b) RIPK1 (74 kDa), RIPK3 (57 kDa), CYLD (107 kDa), MLKL (54 kDa) and ACTB (43 kDa) in the early CL at 4 h after local or systemic PGF2α administration; C – control group, PGF2α − experimental group respectively local (2.5 mg Dinoprost intra-CL) or systemic PGF2α injection (25 mg Dinoprost i.m.)
Additional file 2. Representative western blots bands for (a) CASP3 (32 kDa), CASP8 (18 kDa), BCL2 (26 kDa), BAX (74 kDa) and ACTB (43 kDa) (b) RIPK1 (74 kDa), RIPK3 (57 kDa), CYLD (107 kDa), MLKL (54 kDa) and ACTB (43 kDa) in the mid-stage CL at 4 h after local or systemic PGF2α administrations; C – control group, PGF2α − experimental group respectively local (2.5 mg Dinoprost intra-CL) or systemic PGF2α injection (25 mg Dinoprost i.m.).
Data Availability Statement
The datasets used and analysed during the current study are available from the corresponding author on reasonable request.