

Abstract
Manufacturing influenza virus vaccines using a mammalian cell line rather than embryonated chicken eggs may carry certain advantages. A quadrivalent inactivated influenza virus vaccine produced using the Madin Darby canine kidney cell line has been approved in the EU (Flucelvax® Tetra) and USA (Flucelvax Quadrivalent®; QIVc hereafter) for the prevention of influenza in adults and children. The clinical development of QIVc has built upon that of a cell-based trivalent influenza virus vaccine (TIVc) manufactured using the same processes; the additional influenza B strain contained in QIVc reduces the risk of the strain in the vaccine not matching that in circulation. Pivotal phase III clinical trials in adult and paediatric participants have demonstrated the immunogenicity of QIVc to be noninferior to that of TIVc formulations against shared strains and superior against the influenza B strain absent from each TIVc formulation. Protective efficacy data for TIVc is considered foundational for QIVc and, in a phase III clinical trial, TIVc was effective in protecting adults against antigenically matched influenza strains. Large real-world studies from the 2017/2018 US influenza season further support the prophylactic effectiveness of QIVc, with possible benefits over egg-based vaccines. QIVc was generally well tolerated in clinical trials. In adult and paediatric QIVc recipients, the most common solicited adverse reactions were injection site pain and headache. Reactogenicity was comparable to that of TIVc; no safety signals unique to QIVc emerged. Through circumventing concerns around egg adaptation, QIVc has the potential to be more effective than currently available egg-based quadrivalent vaccines.
Electronic supplementary material
The online version of this article (10.1007/s40265-019-01176-z) contains supplementary material, which is available to authorized users.
QIVc: clinical considerations in the prevention of influenza virus infections
| First quadrivalent influenza virus vaccine produced in mammalian cell cultures |
| Improves upon TIVc; protects against an additional influenza B strain and now entirely cell-derived |
| Immunogenicity not detrimentally impacted by additional strain |
| Effectively protects against influenza, as indicated by a phase III TIVc trial and real-world QIVc data |
| Generally well tolerated; comparable reactogenicity to TIVc |
Introduction
Influenza is associated with considerable morbidity and mortality worldwide, with 3–5 million cases of severe illness and an estimated 290,000–650,000 respiratory deaths resulting from seasonal influenza virus infections each year [1, 2]. While not providing complete protection, vaccination does substantially reduce the burden of influenza and thus represents a major public health initiative [3, 4]. As influenza viruses rapidly mutate and the prophylactic effects of vaccination wane over time, vaccine formulations are updated each year to antigenically match circulating strains and annual vaccination is recommended for optimal protection (and is of particular import in groups at high risk of influenza complications) [1, 4–6].
Trivalent vaccines comprising two influenza A strains and one influenza B strain have been used to protect against influenza since the late 1970s [4]. However, there are two distinct lineages of influenza B virus that now frequently co-circulate [the B/Yamagata/16/88-like (Yamagata) and B/Victoria/2/87-like (Victoria) lineages] [4, 7]. Effectiveness of trivalent vaccines is dependent on the extent to which the influenza B strain in the vaccine matches the strains in circulation, and it can be difficult to predict which lineage will predominate in a given season [4, 7]. Within the last decade, a number of quadrivalent vaccines containing a second influenza B strain have become available to mitigate the issue of mismatch [4].
Most contemporary influenza virus vaccines are produced from viruses grown in embryonated chicken eggs and chemically inactivated [3, 6]. While this production strategy is inexpensive and supported by vast infrastructure, it does have a number of limitations associated with it [6]. An adequate supply of eggs is necessary and production must start long before the influenza season commences, limiting the capacity for sudden changes in circulating strains to be reflected in egg-based vaccines [3, 6]. Importantly, mutations accumulate in the haemagglutinin proteins of influenza viruses in response to selective pressures in the eggs [3, 6, 8]. These mutations can alter antigenicity and may detrimentally impact the prophylactic effectiveness of the vaccine [3, 6, 8]; emerging evidence suggests that such mutations have affected antigenicity against H3N2 viruses over a number of influenza seasons [9].
Utilising mammalian cells instead of chicken eggs to grow influenza viruses may reduce the chance of haemagglutinin mutations arising during the production process, potentially improving vaccine effectiveness [3, 6, 8]. Cell-based vaccine manufacturing also provides a more flexible production timeline once the necessary infrastructure is established [6], and the cells that are used are stored frozen in cell banks to ensure adequate supply [3]. Containing no egg protein, cell-based vaccines eliminate the risk of allergic reactions to this in individuals with egg allergies [3, 6].
A cell-based quadrivalent inactivated influenza virus vaccine (referred to as QIVc hereafter) has been approved in the EU (Flucelvax® Tetra) [10] and USA (Flucelvax Quadrivalent®) [11] as an intramuscular injection for the prophylaxis of influenza in adults and children. Rather than being produced in chicken eggs, QIVc is manufactured from virus propagated in Madin Darby Canine Kidney (MDCK) cells [10–12]. Candidate vaccine viruses (CVVs) are isolated and amplified in the MDCK cell line, then pass through a number of steps validated for MDCK cell removal and are inactivated with β-propiolactone for use in the vaccine [11–13]. These processes reduce levels of viruses or bacteria and other adventitious agents to effectively zero [12, 14]. The active substance in QIVc is a suspension consisting predominantly of purified haemagglutinin and neuraminidase surface antigens; one 0.5 mL dose of QIVc contains ≈ 15 μg of haemagglutinin from each of four influenza strains (A/H1N1, A/H3N2 and B strains from the Yamagata and Victoria lineages [13]; 60 μg in total), against which it provides active immunisation [10, 11]. As with other influenza virus vaccines, administration of QIVc results in the production of humoral antibodies against haemagglutinins, which can neutralise influenza viruses [10, 11].
QIVc is standardised annually in compliance with World Health Organisation (WHO) and Committee for Medicinal Products for Human Use (CHMP) recommendations (in the EU) [10, 13], or United States Public Health Service requirements (in the USA) [11], in order to provide protection against the strains expected to circulate in the upcoming influenza season. While QIVc was initially produced using egg-derived CVVs (with mutations associated with egg adaptation still likely to be present in the vaccine antigens) [6, 8], a cell culture-derived CVV was used for the A/H3N2 strain in the formulation for the 2017/2018 Northern Hemisphere influenza season [8] and for both B strains (in addition to the A/H3N2 strain) in the formulation for the 2018/2019 season [5]. In the formulation for the imminent 2019/2020 season, the use of a cell culture-derived CVV for the A/H1N1 strain will complete the transition to a purely cell-based product [15].
This article focuses on the immunogenicity, protective efficacy and reactogenicity of QIVc. Where relevant, discussion of an established cell-based trivalent inactivated influenza virus vaccine [TIVc; Flucelvax® (USA)/Optaflu® (EU; registration expired)] is also included. TIVc provides immunisation against two influenza A strains (A/H1N1 and A/H3N2) and one influenza B strain (either of the Yamagata or Victoria lineage), with ≈ 15 μg of haemagglutinin from each strain in one 0.5 mL dose [13]. Developed as an upgrade of TIVc, QIVc is manufactured using processes consistent with those used for TIVc and the active drug substance and excipients are the same; the additional antigen is not expected to modify the pharmacology of the vaccine [13].
Immunogenicity
The immunogenicity of QIVc was compared with that of TIVc in two pivotal, double-blind, randomized, multicenter phase III noninferiority trials that enrolled adults aged ≥ 18 years (V130_01) [16] and children and adolescents aged 4 to < 18 years (V130_03) [17] in the USA. Key exclusion criteria included a body temperature ≥ 38 °C within 3 days prior to the vaccination, documented influenza or influenza virus vaccination within the prior 6 months, a history of known or suspected immunodeficiency (congenital or acquired) or receipt of immunosuppressants (within the prior 6 months [16]), and being potentially pregnant, pregnant or breast-feeding [16, 17].
Participants were assigned to either QIVc or one of two TIVc formulations [16, 17]. The TIVc formulations differed only with respect to influenza B strain lineage; TIV1c contained a strain of the Yamagata (B1) lineage while TIV2c contained a strain of the Victoria (B2) lineage. QIVc and TIV1c comprised the influenza strains recommended by the WHO for inclusion in quadrivalent and trivalent vaccines, respectively, in the 2013/2014 Northern Hemisphere influenza season, while the B/Victoria strain in TIV2c was recommended for inclusion in quadrivalent (but not trivalent) vaccines. Randomization was stratified by age (18 to < 65 years vs. ≥ 65 years in V130_01, and 4 to < 9 years vs. 9 to < 18 years in VI30_03) and, in the youngest cohort only, whether or not they had previously been vaccinated against seasonal influenza. Participants received a single 0.5 mL dose of their assigned study vaccine, with the exception of not previously vaccinated children aged 4 to < 9 years who received a second 0.5 mL dose (with the vaccine being administered on days 1 and 29). Vaccines were intramuscularly administered in the deltoid muscle, preferentially that of the non-dominant arm [16, 17].
The co-primary objectives in both trials were to demonstrate the noninferiority of QIVc to a TIVc comparator with respect to haemagglutinin inhibition (HI) geometric mean antibody titre (GMT) ratios (TIVc to QIVc) and differences in seroconversion rate (SCR) [TIVc minus QIVc] for each of the four influenza strains [13, 16, 17]. HI GMT and SCR were assessed at day 22 (in participants receiving one dose) or day 50 (in those receiving two doses) post-vaccination [13, 16, 17]. The comparator TIVc was TIV1c for A/H1N1, A/H3N2 and B1 (the Yamagata strain), and TIV2c for B2 (the Victoria strain) [10, 11, 13]. In V130_01, a key secondary objective was to evaluate antibody responses to all four influenza virus vaccine strains, based on Centre for Biologics Evaluation and Research (CBER) criteria, in adults aged 18 to < 65 years and ≥ 65 years [13]. Noninferiority was assessed in the per-protocol set (PPS), while the full analysis set (FAS) was used for all secondary immunogenicity outcomes [16, 17].
At baseline, demographics and other characteristics were generally well balanced across the vaccine groups in each trial [16, 17]. Participants enrolled in V130_01 and V130_03 had mean ages of ≈ 57 and 9.5 years, respectively, and roughly half were female (57% and 48%). The majority of participants were white (77% in V130_01 and 53% in V130_03) or black (13% and 22%) [16, 17]. In the trial in adults (V130_01), ≈ 25% of each vaccine group had received an influenza virus vaccination within 6–12 months prior to participation; across the four vaccine strains, 84–96% of participants had HI titres ≥ 1:10 [16]. Of the 2680 participants randomized in V130_01, 98% were included in the FAS and 94% were included in the PPS [16]; of the 2333 participants randomized in V130_03, the respective rates were 96% and 87% [17].
In both trials, QIVc was noninferior to the TIVc comparator in terms of HI GMT ratios and SCR differences (Table 1). Where stated (V130_01), analyses of these outcomes in the FAS yielded results that were consistent with those of the primary analyses [13]. Overall, QIVc performed well against the immunogenicity criteria specified by the CBER and CHMP [13, 16, 17].
Table 1.
Immunogenicity of cell-based quadrivalent inactivated influenza virus vaccine in adult and paediatric participants
| Vaccine (no. of participantsa) | A/H1N1 | A/H3N2 | B1 (B/Yamagata/16/88-like) | B2 (B/Victoria/2/87-like) | ||||
|---|---|---|---|---|---|---|---|---|
| GMT (95% CI) |
SCRb (%) (95% CI) |
GMT (95% CI) |
SCRb (%) (95% CI) |
GMT (95% CI) |
SCRb (%) (95% CI) |
GMT (95% CI) |
SCRb (%) (95% CI) |
|
| Adult participants aged ≥ 18 years (V130_01) [10, 11, 16] | ||||||||
| QIVc (n = 1250) |
302.8 (281.8–325.5) |
49.2 (46.4–52.0) |
372.3 (349.2–396.9) |
38.3 (35.6–41.1) |
133.2 (125.3–141.7) |
36.6 (33.9–39.3) |
177.2 (167.6–187.5) |
39.8 (37.0–42.5) |
| TIV1c/TIV2c (n = 635/n = 639) |
298.9 (270.3–330.5) |
48.7 (44.7–52.6) |
378.4 (345.1–414.8) |
35.6 (31.9–39.5) |
115.6 (106.4–125.6) |
34.8 (31.1–38.7) |
164.0 (151.4–177.7) |
35.4 (31.7–39.2) |
| Group ratio (95% CI)c |
1.0 (0.9 – 1.1) |
1.0 (0.9–1.1) |
0.9 (0.8–1.0) |
0.9 (0.9–1.0) |
||||
| Group difference (95% CI)c |
− 0.5 (− 5.3 to 4.2) |
− 2.7 (− 7.2 to 1.9) |
− 1.8 (− 6.2 to 2.8) |
− 4.4 (− 8.9 to 0.2) |
||||
| Paediatric participants aged 4 to < 18 years (V130_03) [13] | ||||||||
| QIVc (n = 1009–1014) |
1090 (1027–1157) |
72 (69–75) |
738 (703–774) |
47 (44–50) |
155 (146–165) |
66 (63–69) |
185 (171–200) |
73 (70–76) |
| TIV1c/TIV2c (n = 501–510) |
1125 (1034–1224) |
75 (70–78) |
776 (725–831) |
51 (46–55) |
154 (141–168) |
66 (62–70) |
185 (166–207) |
71 (67–75) |
| Group ratio (95% CI)c |
1.03 (0.93–1.14) |
1.05 (0.97–1.14) |
0.99 (0.89–1.1) |
1 (0.87–1.14) |
||||
| Group difference (95% CI)c |
2 (− 2.5 to 6.9) |
4 (− 1.4 to 9.2) |
0 (− 5.5 to 4.5) |
− 2 (− 6.5 to 3.2) |
||||
| Paediatric participants aged 9 to < 18 years (V130_03) [10, 17] | ||||||||
| QIVc (n = 545–547) |
1139 (1045–1242) |
70 (66–74) |
719 (673–767) |
42 (38–47) |
200 (185–218) |
63 (58–67) |
212 (192–235) |
72 (68–75) |
| TIV1c/TIV2c (n = 265–272) |
1138 (1007–1286) |
72 (67–78) |
762 (694–836) |
53 (46–59) |
200 (178–224) |
63 (57–69) |
203 (175–234) |
68 (62–74) |
GMT geometric mean titre,QIVc cell-based quadrivalent inactivated influenza virus vaccine, SCR seroconversion rate, TIV1 cell-based trivalent inactivated influenza virus vaccine (influenza B strain from the Yamagata lineage), TIV2c cell-based trivalent inactivated influenza virus vaccine (influenza B strain from the Victoria lineage)
aPer protocol set (defined as all participants who correctly received the assigned vaccine and were not excluded due to reasons defined prior to unblinding/analysis); noninferiority analyses were conducted in this population
bPercentage of participants with either a pre-vaccination H1 titre < 1:10 and post-vaccination H1 titre ≥ 1:40 or a pre-vaccination H1 titre ≥ 1:10 and ≥ 4-fold increase in post-vaccination H1 antibody titre
cCo-primary endpoints; noninferiority established if upper limit of the two-sided 95% CI for ratio of GMTs for HI antibody response (TIV1c or TIV2c divided by QIVc) was < 1.5 and if the upper limit of the two-sided 95% CI for SCR difference (TIV1c or TIV2c minus QIVc) was < 10%
CBER criteria require that, for each vaccine strain, the lower limit of the two-sided 95% CI for the percentage of participants achieving seroconversion for HI antibody is ≥ 40% (≥ 30% in patients aged ≥ 65 years) and that for an HI antibody titre ≥ 1:40 is ≥ 70% (≥ 60% in patients aged ≥ 65 years) [16, 17]. In adults aged 18 to < 65 years (at day 22) and paediatric participants aged 4 to < 18 years (at day 22 or 50), QIVc met both of these immunogenicity criteria for all four vaccine strains, as did TIV1c/TIV2c [16, 17]. In adults aged ≥ 65 years at day 22, QIVc and TIV1c/TIV2c met both CBER criteria for A/H1N1 and the HI antibody titre ≥ 1:40 criterion (but not the seroconversion criterion) for A/H3N2, B1 and B2 [16].
CHMP immunogenicity criteria for adults aged 18 to ≤ 60 years and ≥ 61 years are met if, for each vaccine strain, the point estimate for the geometric mean ratio (GMR; day 22 or 50/day 1) is > 2.5 and > 2.0, respectively; SCR is > 40% and > 30%; and the percentage of patients achieving an HI titre ≥ 1:40 is > 70% and > 60% [16, 17]. The criteria for adults aged 18 to ≤ 60 years was also used in the paediatric study, given the absence of specific CHMP criteria for this age group [17]. QIVc and TIV1c/TIV2c met all CHMP immunogenicity criteria for all four strains in paediatric participants aged 4 to < 18 years and adult participants aged 18 to ≤ 60 years [16, 17]. In older adults (≥ 61 years of age), QIVc and TIV1c/TIV2c met all CHMP criteria for the A/H1N1 strain and two criteria (GMR and HI titre ≥ 1:40, but not seroconversion) for the A/H3N2 and B2 strains [16]. QIVc met two criteria (GMR and HI titre ≥ 1:40, but not seroconversion) for the B1 strain, while TIV1c/TIV2c only met the HI titre ≥ 1:40 criterion [16].
QIVc demonstrated superior immunogenicity to TIV1c and TIV2c with respect to the influenza B strains not included in each trivalent formulation (i.e. B2 for TIV1c and B1 for TIV2c) [16, 17]. In both trials, GMTs and SCRs for the unmatched B strains were higher in QIVc recipients than in TIV1c and TIV2c recipients at 3 weeks post-vaccination, with upper limits of the two-sided 95% CIs for the ratio of GMTs for HI antibody response (GMT TIV1c or TIV2c divided by GMT QIVc) not exceeding the superiority margin of 1 and upper limits of the two-sided 95% CIs for difference in SCR (SCR TIV1c or TIV2c minus SCR QIVc) not exceeding the margin of 0 points [16, 17].
Post-hoc analyses of data from V130_01 and V130_03 generally showed no substantial differences between QIVc and TIV1c/TIV2c within subgroups based on variables such as age, sex, race/ethnicity and baseline immune status [13, 16]. In V130_01, noninferiority criteria were met in subgroups based on age (18 to < 65 years of age vs. ≥ 65 years of age) for all four strains [10, 13]. When stratified by baseline HI serostatus, SCRs were significantly (based on non-overlapping 95% CIs) higher for each strain in participants with baseline HI < 1:10 than in those with baseline HI ≥ 1:10 in both QIVc (71–87% vs. 35–43%) and TIVc (80–88% vs. 32–42%) groups [16]. In both paediatric participants who were seronegative at baseline and those with baseline titres ≥ 1:10, immune responses were comparable between age groups (4 to < 9 years and 9 to < 18 years) for all four strains following vaccination with either QIVc or TIV1c/TIV2c [13].
Results from V130_01 and V130_03 were consistent with the robust immunogenicity of TIVc demonstrated in other large clinical trials in adults aged ≥ 18 years [18–20] and children aged 4 to < 18 years [21]. These include studies that have demonstrated the noninferiority of TIVc to egg-based comparators for all strains in adult participants (based on CHMP immunogenicity criteria) [V58P4] [20] and for the A/H1NI and B strains in paediatric participants (based on ratios of GMTs and differences in SCR, as determined using a cell-derived antigen assay; for A/H3N2, noninferiority was shown for difference in SCR but not GMT ratio) [V58P12] [21]. Persistence of antibodies has also been demonstrated with TIVc; GMTs remained ≥ 3-fold higher than baseline 6 months after vaccination with TIVc in participants aged 18–60 years in V58P9 [18] and GMTs were ≈ 1.4–3-fold higher than baseline 1 year after vaccination with TIVc in adults aged 18 to < 61 years and ≥ 61 years in V58P4E1 [13]. In V58P4EI and V58P4E2, immunogenicity of TIVc in adult and elderly participants was not affected by concomitant vaccination with a pneumococcal vaccine nor by type of influenza virus vaccine received in the previous season [22].
Protective Efficacy
While the protective efficacy of QIVc has not been evaluated in clinical trials, the protective efficacy of TIVc is considered relevant due to the trivalent and quadrivalent vaccines being manufactured in the same manner and having overlapping compositions (Sect. 1) [10, 11]. This section will thus focus on the protective efficacy of TIVc in adults during the 2007–2008 influenza season, as evaluated in a multinational, randomised, observer-blind, placebo-controlled phase III trial (V58P13) [19]. Supportive evidence comes from real-world studies of the effectiveness of QIVc in adults and children during the 2017–2018 influenza season [23–27].
In a Phase III Trial
V58P13 enrolled healthy adults aged 18 to < 50 years in the USA, Finland and Poland [19]. Key exclusion criteria included a body temperature ≥ 37.8 °C and/or acute illness within 3 days of enrolment, laboratory-confirmed influenza or influenza virus vaccination within the 6 months prior to enrolment, a health condition for which an inactivated vaccine is recommended, and being pregnant or breast-feeding. Participants were randomized to receive either a 0.5 mL dose of TIVc (n = 3828), an egg-based trivalent inactivated influenza virus vaccine (TIVe; n = 3676) or placebo (n = 3900), administered in the deltoid muscle of their non-dominant arm. Each dose of TIVc or TIVe contained 15 μg haemagglutinin from each of the recommended virus strains for the 2007–2008 Northern Hemisphere influenza season [19].
The primary objective was to demonstrate the protective efficacy of each vaccine against laboratory-confirmed influenza illness caused by virus strains antigenically similar to those of the vaccines, compared with that of placebo [19]. Influenza surveillance commenced 21 days after vaccine administration, with participants reporting influenza-like illness (ILI) symptoms [defined as fever (temperature ≥ 37.8 °C) plus sore throat or cough], and body aches, chills, headache and runny or stuffy nose; there was also active ILI surveillance via weekly telephone calls. Participants who reported ILI symptoms were clinically evaluated, with nasal and throat specimens collected within 120 h of ILI onset for laboratory confirmation of the influenza virus. Each participant was observed for either the 6-month study surveillance period or 6 months after vaccination (whichever of these was longer) [19].
Demographic and baseline characteristics were similar across the three vaccine groups [19]. In the overall enrolled study population, the mean age was 33 years and 55% of participants were female [11]. The majority of participants were white (84%) [11]. Relatively few participants had previously received influenza virus vaccinations (13–15%) [19]. Of all randomized participants, 11,257 (99%) were evaluable during their 6-month surveillance period and thus included in the efficacy PPS [19].
During the surveillance period, ILI symptoms were reported in 189 TIVc recipients (5%), 243 TIVe recipients (7%) and 353 placebo recipients (9%) in the PPS [19]. Specimen samples were collected from 92% of these participants, with a mean interval of ≈ 39 h from symptom onset to specimen collection. For the cases in which the specimen was collected within the specified 120-h window (97% of cases), influenza virus was confirmed in 25% (42/168), 22% (49/218) and 44% (140/318) of TIVc, TIVe and placebo recipients, respectively [19]. While protective efficacy data pertaining to TIVe is presented in Table 2 for completeness, discussion focusses on TIVc.
Table 2.
Protective efficacy of cell- and egg-based trivalent inactivated influenza virus vaccines in adults
| Strains | TIVc (n = 3776a) | TIVe (n = 3638a) | Placebo (n = 3843a) | Vaccine efficacyb | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TIVc vs. placebo | TIVe vs. placebo | |||||||||
| No. of cases | Attack rate (%) | No. of cases | Attack rate (%) | No. of cases | Attack rate (%) | % | Lower limit of 97.5% CIc | % | Lower limit of 97.5% CIc | |
| Antigenically matchedd(i.e. vaccine-like) strains | ||||||||||
| Overalle | 7 | 0.19 | 9 | 0.25 | 44 | 1.14 | 83.8** | 61.0 | 78.4* | 52.1 |
| A/H1N1 | 5 | 0.13 | 8 | 0.22 | 43 | 1.12 | 88.2** | 67.4 | 80.3* | 54.7 |
| A/H3N2f | 2 | 0.05 | 1 | 0.03 | 0 | 0 | ||||
| Bf | 0 | 0 | 0 | 0 | 1 | 0.03 | ||||
| Non-antigenically matchedg(i.e. non-vaccine-like) strains | ||||||||||
| Overall | 30 | 0.79 | 29 | 0.80 | 74 | 1.93 | 58.7 | 33.5 | 58.6 | 32.9 |
| A/H1N1 | 1 | 0.03 | 0 | 0 | 8 | 0.21 | 87.3 | 4.6 | 100 | 33.9 |
| A/H3N2 | 0 | 0 | 2 | 0.05 | 8 | 0.21 | 100 | 36.3 | 73.6 | − 30.1 |
| B | 29 | 0.77 | 27 | 0.74 | 59 | 1.54 | 50.0 | 17.5 | 51.7 | 19.4 |
| All culture-confirmed influenza | ||||||||||
| Overall | 42 | 1.11 | 49 | 1.35 | 140 | 3.64 | 69.5** | 55.0 | 63.0* | 46.7 |
| A/H1N1 | 6 | 0.16 | 10 | 0.27 | 57 | 1.48 | 89.3** | 73.0 | 81.5** | 60.9 |
| A/H3N2 | 6 | 0.16 | 12 | 0.33 | 25 | 0.65 | 75.6 | 35.1 | 49.3 | − 9.0 |
| B | 30 | 0.79 | 27 | 0.74 | 61 | 1.59 | 49.9 | 18.2 | 53.2 | 22.2 |
Results of V58P13 (NCT00630331) [19]. Efficacies of TIVc and TIVe exceeded the CBER efficacy criteria
CBER Center for Biologics Evaluation and Research, TIVc cell-based trivalent inactivated influenza virus vaccine, TIVe egg-based trivalent inactivated influenza virus vaccine
*p < 0.01, **p < 0.001 (adjusted p-values; p < 0.025 indicates a vaccine efficacy significantly larger than 40%)
aEfficacy per protocol population (participants who were evaluable during their individual 6-month surveillance period)
b(1 − relative risk) × 100
cSimultaneous one-sided 97.5% CIs for vaccine efficacy of TIVc or TIVe relative to placebo; efficacy success criterion was a lower limit > 40% (prespecified for overall analyses but not for individual strains [11])
dIsolates were considered matched if there was a ≤ 4-fold difference in titre of the isolate and the vaccine strain against a reference antiserum
ePrimary endpoint (overall vaccine efficacy of each vaccine relative to placebo) achieved for each vaccine [13, 19]; see success criterion above
fToo few cases to adequately assess vaccine efficacy [10, 11]
gIsolates were considered non-vaccine-like if the haemagglutination inhibition antibody titre was ≥ 1:8 against specific reference strain antisera
TIVc provided effective prophylaxis against antigenically matched strains overall (primary endpoint) and against antigenically matched A/H1N1 virus, with vaccine efficacy relative to placebo being significantly higher than the prespecified criterion of 40% (Table 2) [19]. Overall, the attack rate for antigenically matched strains in participants who received TIVc was about one-sixth of that seen in placebo recipients (0.19 vs. 1.14; Table 2). There were too few cases of influenza caused by antigenically matched A/H3N2 and B strains to adequately assess vaccine efficacy (Table 2).
When looking at all circulating strains (matched and non-matched) responsible for cases of influenza, the vaccine efficacy of TIVc relative to placebo was also significantly higher than 40% (Table 2). The attack rate with TIVc was less than one-third of that seen with placebo (1.11 vs. 3.64; Table 2). With respect to individual virus strains, the vaccine efficacy of TIVc relative to placebo was significantly higher than 40% for A/H1N1 but not A/H3N2 or B (Table 2). When considering only non-matched strains, the vaccine efficacy did not exceed the 40% threshold for any individual strain nor overall (Table 2). Most cases of non-matched influenza were caused by influenza B strain viruses; across the groups, non-matched strains were responsible for 115/116 cases of culture-confirmed influenza B [19].
In Real-World Studies (2017/2018 Season)
The real-world protective effectiveness of QIVc during the 2017/2018 influenza season in the USA has been investigated in retrospective cohort studies [23, 25–27] and test negative case–control studies [24, 25, 28], the results of which indicate that, like egg-based vaccines, cell-based influenza virus vaccines are effective in preventing influenza. This section focuses on the three largest of these studies (retrospective cohort studies each in n > 90,000 QIVc recipients) [23, 25, 26]. Relative vaccine effectiveness [RVE; calculated as 100 × (1 − odds ratio) [23] or 100 × (1 − rate ratio) [26], with models adjusted for covariates [23, 25, 26]] is reported.
In a study using data from electronic medical records of patients aged ≥ 4 years of age presenting to primary care in the USA between 1 August 2017 and 31 March 2018, QIVc (n = 92,192 recipients) was significantly more effective than an egg-based quadrivalent vaccine (QIVe; n = 1,255,983) in preventing ILI after adjusting for demographic confounds (RVE of QIVc vs. QIVe 36.2%; 95% CI 26.1–44.9; p < 0.001) [abstract] [23]. When specific age groups were examined, this effect was significant in adults aged 18 to < 65 years (RVE 26.8; 95% CI 14.1–37.6; p < 0.001) but not paediatric patients aged 4 to < 18 years (RVE 18.8%; 95% CI −53.9 to 57.2) or older adults aged ≥ 65 years (RVE −7.3; 95% CI −51.6 to 24.0) [propensity-score matched models; no other covariates adjusted for] [23].
Data from the Defense Medical Surveillance System (DMSS) was used to examine the effectiveness of QIVc (n = 392,116) versus that of QIVe (n = 371,394) in active component US service members, the majority of whom were aged 18 to < 40 years (91% and 90% in the respective vaccine groups) and male (84% and 82%) [abstract] [25]. QIVc was significantly more effective than QIVe in preventing influenza diagnosed during any medical encounter (2732 vs. 3360 cases; RVE 16%; 95% CI 11–20), but not influenza diagnosed during hospitalization only (11 vs. 20 cases; RVE 46%; 95% CI −18 to 76), after adjusting for age group, sex, month of vaccination and receipt of influenza virus vaccination during the prior season (statistical significance based on 95% CIs). With respect to ILI, there were no significant differences between QIVc and QIVe for rates of ILI diagnosed during any medical encounter (40,736 vs. 40,991 cases; adjusted RVE 2%; 95% CI 0–4) or hospitalization only (120 vs 139 cases; adjusted RVE 16%; 95% CI −9 to 35) [25].
QIVc conferred significant benefits over QIVe in a large retrospective cohort study of Medicare beneficiaries aged ≥ 65 years who received an influenza virus vaccination from 6 August 2017 through 31 January 2018 [26]. QIVc was administered in 5% (n = 659,249) of eligible vaccinees, while QIVe was administered in 14% (n = 1,863,654). After adjusting for baseline differences in covariates between the vaccine groups, QIVc was significantly more effective than QIVe in preventing influenza-related hospital encounters (defined as an inpatient hospitalization or emergency department visit; primary outcome) [RVE 10.0%; 95% CI 7.0–13.0]. Results from sensitivity analyses were consistent with this estimate. QIVc was also significantly more effective than QIVe in preventing influenza-related office visits (defined as community-based physician office visits or hospital outpatient visits in which a rapid influenza diagnostic test was performed and a therapeutic course of oseltamivir was prescribed within 2 days) [RVE 10.5%; 95% CI 6.8–14.0 (2-way comparison adjusted for covariates)]. In five-way comparisons that included data from eligible vaccinees who received egg-based standard-dose (n = 1,018,494), high-dose (n = 8,489,159) or MF59-adjuvanted (n = 1,473,536) trivalent vaccinations, QIVc was significantly more effective than QIVe and egg-based standard-dose (but not high-dose) and adjuvanted trivalent vaccines in preventing influenza-related hospital encounters and influenza-related inpatient stays, and significantly more effective than QIVe and egg-based high-dose (but not standard-dose) and adjuvanted trivalent vaccines in preventing influenza-related office visits (adjusted for covariates; p ≤ 0.05 for all comparisons) [26].
Reactogenicity and Safety
QIVc was generally well tolerated in the pivotal phase III trials in adult (V130_01) [16] and paediatric participants (V130_03) [17] (which will be the focus of this section). Within 7 days of QIVc administration in the pivotal trials, solicited adverse reactions occurred in 62% of adult participants aged 18 to < 65 years (vs. 57% and 60% with TIV1c and TIV2c) [16] and in 71% of paediatric participants aged 9 to < 18 years (vs. 68% and 61%) [17]. The most common solicited adverse reactions (incidence ≥ 10% with any vaccine) reported in these age groups were injection site pain and headache (Fig. 1) [11]. In older adults (aged ≥ 65 years; n = 1332 with 7 days of solicited adverse reaction data [11]), solicited adverse reactions were reported by 41% of QIVc recipients (vs. 39% and 43% of TIV1c and TIV2c recipients) [16] and the most common were injection site pain (21.6% vs. 18.8% and 18.5%), injection site erythema (11.9% vs. 10.6% and 10.4%) and headache (9.3% vs. 8.5% and 8.3%) [11]. The overall paediatric population (aged 4 to < 18; n = 2264) reported similar rates of local and systemic adverse reactions to those reported by participants aged 9 to < 18 years [10]. In both adult and paediatric participants, the majority of solicited adverse reactions were mild to moderate in severity [13, 16, 17]. Fever (body temperature ≥ 38.0 °C) occurred at low rates across the QIVc, TIV1c and TIV2c arms (0.5%, 0.7% and 0.5% of adults aged ≥ 18 years and 3%, 4% and 2% of children aged 4 to < 18 years [13]); no cases of body temperature ≥ 40.0 °C were reported in adults and one case was reported in a paediatric participant (a QIVc recipient aged 9 to < 18 years) [13, 16].
Fig. 1.
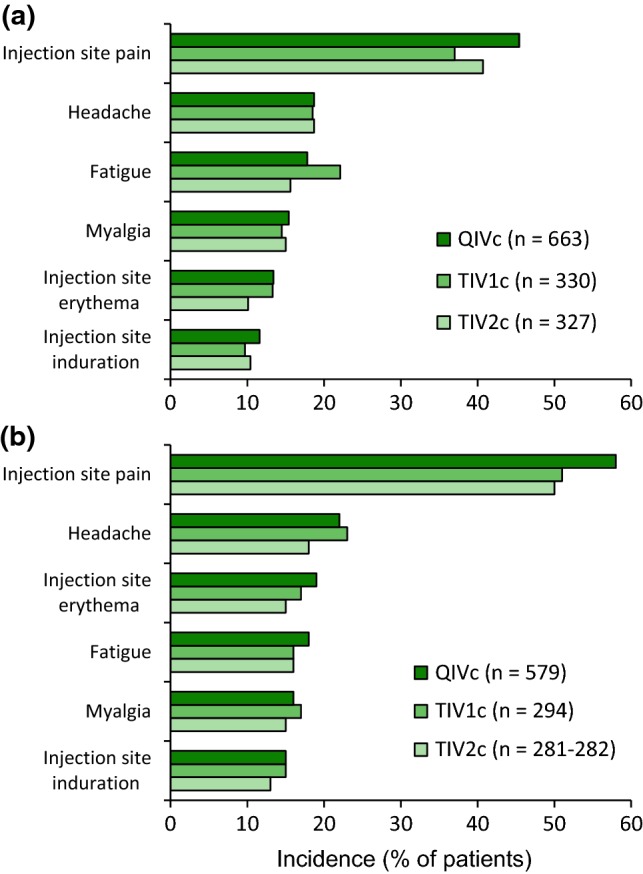
Most common any-grade solicited adverse reactions (incidence ≥ 10% in any treatment group) reported within 7 days of influenza virus vaccine administration in a adults aged 18 to < 65 years (data from V130_01) and b children aged 9 to < 18 years (data from V130_03) [11]. QIVc cell-based quadrivalent inactivated influenza virus vaccine,TIV1 cell-based trivalent inactivated influenza virus vaccine (influenza B strain from the Yamagata lineage), TIV2c cell-based trivalent inactivated influenza virus vaccine (influenza B strain from the Victoria lineage)
Severe solicited adverse reactions were infrequent (≥ 1%) of adults aged ≥ 18 years administered QIVc [13]; severe pain occurred in 0.2% and 0.1% of QIVc and TIV1c recipients [16]. In the overall paediatric population, there were low rates of severe injection site pain (1% of participants in each vaccine group), tenderness (2%, 1% and 2% of participants administered QIVc, TIV1c and TIV2c, respectively), erythema and induration (each < 1% of patients in each group); in participants aged 9 to < 18 years, no severe solicited local adverse reaction occurred at a rate of ≥ 1% in any vaccine group [13]. The only severe solicited systemic adverse reaction experienced by > 1% of paediatric QIVc recipients was irritability (2% with QIVc and TIV1c) [13].
Rates of unsolicited adverse events were comparable across vaccine groups in both adult (16%, 15% and 17% with QIVc, TIV1c and TIV2c, respectively) [13, 16] and paediatric participants (24%, 24% and 27%) [17]. Those considered to be at least possibly related to the study vaccine were infrequent (in 3–5% of adults aged 18 to < 64 years, 4–5% of adults aged ≥ 65 years [16] and 5–6% of paediatric participants [17]). In QIVc recipients aged ≥ 18 years, the most common possibly or probably vaccine-related unsolicited adverse event was injection site haemorrhage (0.8% vs. 0.4% and 0.6% with TIV1c and TIV2c) [16]. Medically attended adverse events also occurred at similar rates across vaccine groups (26%, 26% and 25% of adult participants receiving QIVc, TIV1c and TIV2c, respectively [16], and 27% of paediatric participants in each vaccine group [17]).
Serious adverse events (SAEs; collected ≤ 6 months after vaccination) were reported in 1.7%, 1.8% and 1.5% of QIVc, TIV1c and TIV2c recipients aged 18 to < 65 years, while the respective rates in adults aged ≥ 65 years were 6.2%, 4.7% and 4.7% [16]. New onset of chronic disease (NOCD) was reported in 3.6% of QIVc recipients aged 18 to < 65 years (vs. 3.0% and 3.7% of TIV1c and TIV2c recipients) and 5.8% of QIVc recipients aged ≥ 65 years (vs. 4.4% and 5.0%); these were most commonly metabolism and nutritional disorders (0.8% vs. 0.7% and 0.5%), cardiac disorders (0.8% vs. 0.6% and 0.3%), and musculoskeletal and connective tissue disorders (0.8% vs. 0.4% and 0.3%) [16]. In the paediatric trial, SAEs occurred at low rates (0.5%, 1.2% and 0.4% of QIVc, TIV1c and TIV2c recipients, respectively [13]), as did NOCD (2% of patients in each vaccine group) [17]. No SAE or case of NOCD was considered to be related to the study vaccine in either trial [13]. There were 12 deaths during the adult trial, none of which were considered to be related to the study vaccine and most of which occurred in adults ≥ 65 years of age (rates were 0.8%, 1.5% and 0.3% with QIVc, TIV1c and TIV2c, respectively, in this age group vs. 0%, 0% and 0.3% in adults aged 18 to < 65 years) [16]. No deaths were reported during the paediatric trial [17].
Safety data (pertaining to both QIVc and the TIVc comparators) from these trials were generally consistent with those from numerous clinical studies of TIVc in adults and children [13, 18–22, 29–33], with the additional strain not altering the safety profile of the QIVc vaccine; no new safety signals were identified [13]. The clinical development program for TIVc found it to have a safety profile comparable to that of an egg-based trivalent influenza virus vaccine [13, 18–22, 31, 32, 34]. Post-marketing safety data are available for TIVc, representing millions of adults and children vaccinated in Europe and the USA in the 2016/2017 influenza season, and have confirmed its established safety profile [13]; similarly, an earlier surveillance study in the USA (2013–2015) found no concerning patterns in the adverse events reported in TIVc recipients [35].
Dosage and Administration
QIVc is indicated for the prophylaxis of influenza in adults and children [10]. A single dose is 0.5 mL (which includes ≈ 15 μg of haemagglutinin from each influenza strain), administered as an intramuscular injection to the deltoid muscle of the upper arm [10, 11]. The minimum age of eligibility for the vaccine and the available presentations of the vaccine differ between the EU and USA (see Supplementary Table 1); local prescribing information should be consulted for specific details. Local prescribing information should also be consulted for details concerning administration, storage and handling, contraindications, warnings and precautions, and use in specific patient populations.
Place of Flucelvax® Tetra/Flucelvax Quadrivalent® in the Prevention of Influenza
Vaccination is currently the best strategy for reducing morbidity and mortality associated with influenza infection [1, 8], and the WHO recommends annual vaccination in individuals at high risk of influenza complications and those who live with or care for them [1]. While influenza virus vaccines have historically been produced in embryonated chicken eggs and the majority of contemporary quadrivalent and trivalent vaccines are egg-based, some manufacturers are now moving towards cell-based technology. Approved in the EU and USA (Sect. 5), QIVc is the most widely available cell-based quadrivalent vaccine. The most recent recommendations from the US Centers for Disease Control and Prevention (CDC) Advisory Committee on Immunization Practices (ACIP), developed for the 2018/2019 influenza season, state that vaccination providers may choose to administer any licensed, age-appropriate influenza virus vaccine; if more than one is appropriate, no preferential recommendation is made for use of any particular vaccine over another [5]. In individuals with a history of egg allergy of any severity, providers may also select any of a number of licensed, recommended and age-appropriate vaccines [5]. Benefits of quadrivalent versus trivalent formulations in any one season are dependent on the extent to which the influenza B strain in the trivalent vaccine matches those in circulation [4, 7] and, while acknowledging that quadrivalent vaccines are designed to provide broader influenza B protection, ACIP recommendations do not express a preference for either quadrivalent or trivalent inactivated influenza virus vaccines [5]. For the 2019/2020 influenza season in the UK, the Joint Committee on Vaccination and Immunisation has recommended the use of QIVc or QIVe over a trivalent vaccine in individuals < 65 years in at risk groups [36]. In adults ≥ 65 years of age, the use of QIVc, an adjuvanted trivalent vaccine or a high-dose trivalent vaccine is considered preferable to the use of QIVe [36].
The clinical development program of QIVc builds upon that of TIVc, which has demonstrated TIVc to have comparable immunogenicity (Sect. 2) and reactogenicity (Sect. 4) to licensed egg-based comparator vaccines [13]. Given that QIVc and TIVc are manufactured in the same manner aside from the additional influenza B strain in QIVc (Sect. 1), data from clinical trials of TIVc are considered relevant to QIVc [13].
In well-designed, phase III clinical trials in adult and paediatric participants, QIVc had noninferior immunogenicity to TIVc comparators based on ratios of GMTs and differences in SCRs for each matched influenza strain; the additional influenza B strain contained in QIVc did not interfere with immune responses to the other three strains (Sect. 2). Furthermore, QIVc was superior to the TIVc comparators with respect to the influenza B strain that each trivalent formulation did not contain (of Yamagata or Victoria lineage), indicating the potential for broader protection with the quadrivalent product. QIVc fulfilled all CBER and CHMP immunogenicity criteria, except in older adults (in whom the majority of the criteria were met) [Sect. 2]. These trials, whilst limited to the USA and excluding pregnant or breastfeeding women and individuals with impaired immunity, enrolled participants who were largely representative of the general population (including those with different underlying medical conditions) [13, 16].
While clinical trial data on the protective efficacy of QIVc are not available, TIVc provided effective prophylaxis in the phase III V58P13 trial with a vaccine efficacy of 84% (significantly > 40%) relative to placebo for antigenically matched, culture-confirmed influenza (Sect. 3). Based on immunogenicity results from V130_01 considered alongside immunogenicity and efficacy data from V58P13, QIVc is expected to induce comparable clinical protection in adults [13]. While there are no protective efficacy data pertaining to the use of TIVc in the paediatric age group, TIVc has demonstrated noninferior immunogenicity to an egg-based trivalent comparator with respect to certain influenza strains in a paediatric trial (Sect. 2) and the protective efficacy of this comparator has been established in children and adolescents [13]. Furthermore, the immunogenicity of TIVc in paediatric participants aged 9 to < 18 in the V58P12 study suggests that the protective efficacy shown in adults in V58P13 (which was conducted during the same influenza season) may be expected to apply to this younger age group [13]. Importantly, in addition to this indirect immunobridging, there is a phase III/IV trial (V130_12) currently evaluating the efficacy of QIVc in paediatric participants [37]. This trial was a post-approval requirement in the USA [13], and results are awaited with interest. Given the particular burden of influenza B infection in children and adolescents [4, 38], protection against both influenza B lineages is likely to be especially desirable in the paediatric population.
Manufacturing influenza virus vaccines using mammalian cell lines as opposed to embryonated chicken eggs appears to improve protective effectiveness. Replication of influenza viruses in MDCK cells rather than chicken eggs has been theorised to reduce the risk of haemagglutinin mutations arising [3, 6, 8]. Consequently, compared with the influenza strains contained in egg-based vaccines, the strains contained in cell-based influenza virus vaccines may be more closely matched to circulating strains [3, 6, 8]. Indeed, data from the 2011/2012 to 2017/2018 influenza seasons in the Northern Hemisphere has shown that, consistently, substantially higher proportions of circulating H3N2 viruses have matched MDCK-propagated reference viruses than have matched egg-propagated reference viruses [9]. The tendency for mismatch between circulating isolates and egg-propagated reference viruses may have contributed to historically low vaccine effectiveness against H3N2 strains [9]. QIVc has been gradually shifting from the use of egg-derived CVVs to cell culture-derived CVVs to ensure the insulation of the vaccine from concerns around egg adaptation (Sect. 1), and the upcoming 2019/2020 influenza season heralds the first purely cell-based formulation of QIVc [15]. As of yet, it is uncertain whether this shift to exclusively cell culture-derived CVVs will bestow improved vaccine effectiveness.
Large real-world studies (n > 90,000 QIVc recipients) using data from the 2017/2018 US influenza season suggest that, like egg-based alternatives, QIVc is effective in preventing influenza and may offer benefits over these products for influenza-related outcomes in some cohorts (Sect. 3.2). Medicare beneficiaries aged ≥ 65 years who received QIVc were 10% less likely than those who received QIVe to have an influenza-related hospital encounter, although this modest difference in vaccine effectiveness suggests that egg adaptation was not the sole cause for the low vaccine effectiveness reported during the A/H3N2-dominated 2017/2018 influenza season [26]. In the QIVc formulation produced for 2017/2018 influenza season, A/H3N2 was the only influenza strain for which a cell culture-derived CVV (as opposed to an egg-derived CVV) was used [8, 26]. Given the limitations of observational studies (e.g. the potential presence of residual confounds), a definitive comparative trial would be of use in determining the extent to which QIVc fully derived from and manufactured in a mammalian cell line improves vaccine effectiveness relative to egg-based quadrivalent products.
QIVc was generally well tolerated in both adult and paediatric participants in the pivotal phase III trials, with a safety profile consistent with that of TIVc; the additional strain did not markedly alter the reactogenicity of the vaccine and no safety signals unique to QIVc emerged (Sect. 4). In both adult and paediatric participants administered QIVc, the most common solicited local adverse reaction was injection site pain and the most common solicited systemic adverse reaction was headache. Solicited adverse reactions occurred at somewhat lower rates in older adults (aged ≥ 65 years of age) compared with younger participants. The majority of solicited adverse reactions were mild to moderate in severity. No SAE was considered to be related to the study vaccine. As there are currently limited safety data for QIVc in certain patient groups (e.g., those who are immunocompromised, have certain underlying diseases or are pregnant or breastfeeding), these populations are being followed post-authorization [13] and there is a pregnancy registry for QIVc and TIVc recipients [39]. In general, inactivated influenza virus vaccines such as QIVc are suitable for use in a wider population than live attenuated vaccines, which are not recommended in a number of patient groups [5].
As well as circumventing the issue of egg adaptation, cell-based manufacturing has other benefits over the traditional egg-based approach. Containing no egg protein, QIVc eliminates the risk of allergic reactions to this in individuals with egg allergies [3, 6]. Unlike egg-based vaccines, QIVc can be produced without need for antibiotics or preservatives [13]. Cell-based manufacturing is not dependent on egg supply and provides a more flexible production timeline where production can be promptly scaled up in the face of a pandemic and the vaccine can be altered at short notice to reflect late-emerging strains [6, 8]. However, the global-scale infrastructure necessary to produce QIVc in quantities comparable to egg-based vaccines has not yet been established [6]. QIVc is also more expensive than egg-based quadrivalent products (based on the CDC 2019/2020) price list [40], which may affect uptake of the product. Cost-effectiveness analyses comparing QIVc with various other available vaccines would be of interest.
In conclusion, QIVc is an immunogenic and generally well tolerated quadrivalent inactivated influenza virus vaccine for the prevention of influenza in adults and children. Immunogenicity and safety were not detrimentally impacted relative to that of TIVc by the inclusion of the additional influenza B strain, and QIVc has the capacity to offer broader influenza B protection than the trivalent formulation. Unlike the majority of quadrivalent vaccines, QIVc is cell-based and exemplifies a shift away from the use of egg-based technology in the manufacturing of influenza virus vaccines. Through circumventing concerns around egg adaptation, QIVc has the potential to be more effective than currently available egg-based quadrivalent vaccines.
Data Selection Flucelvax Tetra: 226 records identified
| Duplicates removed | 73 |
| Excluded during initial screening (e.g. press releases; news reports; not relevant drug/indication; preclinical study; reviews; case reports; not randomized trial) | 79 |
| Excluded during writing (e.g. reviews; duplicate data; small patient number; nonrandomized/phase I/II trials) | 34 |
| Cited efficacy/tolerability articles | 19 |
| Cited articles not efficacy/tolerability | 21 |
| Search Strategy: EMBASE, MEDLINE and PubMed from 1946 to present. Clinical trial registries/databases and websites were also searched for relevant data. Key words were Flucelvax TETRA, Flucelvax Quadrivalent, Optaflu, influenza vaccine. Records were limited to those in English language. Searches last updated 9 July 2019 | |
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
During the peer review process, the manufacturer of QIVc was also offered an opportunity to review this article. Changes resulting from comments received were made on the basis of scientific and editorial merit.
Compliance with Ethical Standards
Funding
The preparation of this review was not supported by any external funding.
Conflicts of interest
Yvette Lamb is a salaried employee of Adis International Ltd/Springer Nature, is responsible for the article content and declares no relevant conflicts of interest.
Footnotes
The original version of this article was revised due to a retrospective Open Access request.
The manuscript was reviewed by:G. Icardi, Department of Health Sciences, University of Genoa, Genoa, Italy; F. Krammer, Department of Microbiology, Icahn School of Medicine at Mount Sinai, New York, NY, USA; M. Petras, Preventive Medicine, Third Faculty of Medicine, Charles University, Prague, Czech Republic.
Additional information for this Adis Drug Evaluation can be found at 10.6084/m9.figshare.8830517.
Change history
11/25/2019
The article Cell-Based Quadrivalent Inactivated Influenza Virus Vaccine.
References
- 1.World Health Organization. Fact sheet: influenza (seasonal). 2018. http://www.who.int/. Accessed 9 Jul 2019.
- 2.Iuliano AD, Roguski KM, Chang HH, et al. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. Lancet. 2018;391(10127):1285–1300. doi: 10.1016/S0140-6736(17)33293-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Buhler S, Ramharter M. Flucelvax Tetra: a surface antigen, inactivated, influenza vaccine prepared in cell cultures. ESMO Open. 2019;4(1):e000481. doi: 10.1136/esmoopen-2018-000481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Barr IG, Jelley LL. The coming era of quadrivalent human influenza vaccines: who will benefit? Drugs. 2012;72(17):2177–2185. doi: 10.2165/11641110-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 5.Grohskopf LA, Sokolow LZ, Broder KR, et al. Prevention and control of seasonal influenza with vaccines: recommendations of the Advisory Commitee on Immunization Practices—United States, 2018–2019 influenza season. Morb Mortal Weekly Rep. 2018;67(3):1–20. doi: 10.15585/mmwr.rr6703a1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Harding AT, Heaton NS. Efforts to improve the seasonal influenza vaccine. Vaccines. 2018;6(19):E19. doi: 10.3390/vaccines6020019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tisa V, Barberis I, Faccio V, et al. Quadrivalent influenza vaccine: a new opportunity to reduce the influenza burden. J Prev Med Hyg. 2016;57(1):E28–E33. [PMC free article] [PubMed] [Google Scholar]
- 8.Barr IG, Donis RO, Katz JM, et al. Cell culture-derived influenza vaccines in the severe 2017–2018 epidemic season: a step towards improved influenza vaccine effectiveness. NPJ Vaccines. 2018;3:44. doi: 10.1038/s41541-018-0079-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rajaram S, van Boxmeer J, Leav B, et al. Retrospective evaluation of mismatch from egg-based isolation of influenza strains compared with cell-based isolation and the possible implications for vaccine effectiveness. Open Forum Infect Dis. 2018;5(Suppl. 1):S69. doi: 10.1093/ofid/ofy209.164. [DOI] [Google Scholar]
- 10.European Medicines Agency. Flucelvax Tetra: summary of product characteristics. 2018. http://www.ema.europa.eu/. Accessed 9 Jul 2019.
- 11.Seqirus Inc. Flucelvax Quadrivalent (influenza vaccine) suspension for intramuscular injection (2018-2019 formula): prescribing information. 2018. http://labeling.seqirus.com/. Accessed 9 Jul 2019.
- 12.Onions D, Egan W, Jarrett R, et al. Validation of the safety of MDCK cells as a substrate for the production of a cell-derived influenza vaccine. Biologicals. 2010;38(5):544–551. doi: 10.1016/j.biologicals.2010.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.European Medicines Agency. Flucelvax Tetra—public assessment report. 2018. http://www.ema.europa.eu/. Accessed 9 Jul 2019.
- 14.Gregersen JP. A quantitative risk assessment of exposure to adventitious agents in a cell culture-derived subunit influenza vaccine. Vaccine. 2008;26(26):3332–3340. doi: 10.1016/j.vaccine.2008.03.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Seqirus. Seqirus announces further advances in cell-based influenza vaccine technology 2019. http://www.prnewswire.com/. Accessed 9 Jul 2019.
- 16.Bart S, Cannon K, Herrington D, et al. Immunogenicity and safety of a cell culture-based quadrivalent influenza vaccine in adults: a phase III, double-blind, multicenter, randomized, non-inferiority study. Hum Vaccin Immunother. 2016;12(9):2278–2288. doi: 10.1080/21645515.2016.1182270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hartvickson R, Cruz M, Ervin J, et al. Non-inferiority of mammalian cell-derived quadrivalent subunit influenza virus vaccines compared to trivalent subunit influenza virus vaccines in healthy children: a phase III randomized, multicenter, double-blind clinical trial. Int J Infect Dis. 2015;41:65–72. doi: 10.1016/j.ijid.2015.11.004. [DOI] [PubMed] [Google Scholar]
- 18.Ambrozaitis A, Groth N, Bugarini R, et al. A novel mammalian cell-culture technique for consistent production of a well-tolerated and immunogenic trivalent subunit influenza vaccine. Vaccine. 2009;27(43):6022–6029. doi: 10.1016/j.vaccine.2009.07.083. [DOI] [PubMed] [Google Scholar]
- 19.Frey S, Vesikari T, Szymczakiewicz-Multanowska A, et al. Clinical efficacy of cell culture-derived and egg-derived inactivated subunit influenza vaccines in healthy adults. Clin Infect Dis. 2010;51(9):997–1004. doi: 10.1086/656578. [DOI] [PubMed] [Google Scholar]
- 20.Szymczakiewicz-Multanowska A, Groth N, Bugarini R, et al. Safety and immunogenicity of a novel influenza subunit vaccine produced in mammalian cell culture. J Infect Dis. 2009;200(6):841–848. doi: 10.1086/605505. [DOI] [PubMed] [Google Scholar]
- 21.Vesikari T, Block SL, Guerra F, et al. Immunogenicity, safety and reactogenicity of a mammalian cell-culture-derived influenza vaccine in healthy children and adolescents three to seventeen years of age. Pediatr Infect Dis J. 2012;31(5):494–500. doi: 10.1097/INF.0b013e31824bb179. [DOI] [PubMed] [Google Scholar]
- 22.Szymczakiewicz-Multanowska A, Lattanzi M, Izu A, et al. Safety assessment and immunogenicity of a cell-culture-derived influenza vaccine in adults and elderly subjects over three successive influenza seasons. Hum Vaccin Immunother. 2012;8(5):645–652. doi: 10.4161/hv.19493. [DOI] [PubMed] [Google Scholar]
- 23.Boikos C, Sylvester G, Sampalis J, et al. Effectiveness of the cell culture-based and egg-based, seasonal influenza vaccines during the 2017–2018 Northern Hemisphere influenza season [abstract]. In: Canadian Immunization Conference 2018. 2018.
- 24.Bruxvoort K, Luo Y, Ackerson B, et al. Comparison of vaccine effectiveness against influenza hospitalization of cell-based and egg-based influenza vaccines, 2017–2018 [abstract]. Annual Conference on Vaccinology Research (April 2019). 2019. [DOI] [PubMed]
- 25.Eick-Cost AA, Hu Z. Relative effectiveness of cell-based influenza vaccines compared to egg-based influenza vaccines, active component U.S. service members, 2017–18 season [abstract]. In: International Conference on Emerging Infectious Diseases. 2018.
- 26.Izurieta HS, Chillarige Y, Kelman J, et al. Relative effectiveness of cell-cultured and egg-based influenza vaccines among elderly persons in the United States, 2017–2018. J Infect Dis. 2018 doi: 10.1093/infdis/jiy716. [DOI] [PubMed] [Google Scholar]
- 27.Klein NP, Fireman B, Goddard K, et al. Vaccine effectiveness of Flucelvax relative to inactivated influenza vaccine during the 2017–18 influenza season in Northern California. Open Forum Infect Dis. 2018;5(Suppl. 1):S764. doi: 10.1093/ofid/ofy229.2189. [DOI] [Google Scholar]
- 28.Shoubaki L, Eick-Cost AA, Hawksworth AW, et al. Department of Defense midseason vaccine effectiveness estimates for the 2017–2018 influenza season. MSMR. 2018;25(6):26–28. [PubMed] [Google Scholar]
- 29.Groth N, Montomoli E, Gentile C, et al. Safety, tolerability and immunogenicity of a mammalian cell-culture-derived influenza vaccine: a sequential phase I and phase II clinical trial. Vaccine. 2009;27(5):786–791. doi: 10.1016/j.vaccine.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 30.Loebermann M, Voss U, Meyer S, et al. Clinical trial to evaluate the safety and immunogenicity of a trivalent surface antigen seasonal influenza vaccine produced in mammalian cell culture and administered to young and elderly adults with and without A(H1N1) pre-vaccination. PLoS ONE. 2013;8(8):e70866. doi: 10.1371/journal.pone.0070866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nolan T, Chotpitayasunondh T, Capeding MR, et al. Safety and tolerability of a cell culture derived trivalent subunit inactivated influenza vaccine administered to healthy children and adolescents: a phase III, randomized, multicenter, observer-blind study. Vaccine. 2016;34(2):230–236. doi: 10.1016/j.vaccine.2015.11.040. [DOI] [PubMed] [Google Scholar]
- 32.Reisinger KS, Block SL, Izu A, et al. Subunit influenza vaccines produced from cell culture or in embryonated chicken eggs: comparison of safety, reactogenicity, and immunogenicity. J Infect Dis. 2009;200(6):849–857. doi: 10.1086/605506. [DOI] [PubMed] [Google Scholar]
- 33.Vinnemeier CD, Fischer-Herr J, Meyer S, et al. Immunogenicity and safety of an inactivated 2012/2013 trivalent influenza vaccine produced in mammalian cell culture (Optaflu®): an open label, uncontrolled study. Hum Vaccin Immunother. 2014;10(2):441–448. doi: 10.4161/hv.27140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Diez-Domingo J, de Martino M, Lopez JG, et al. Safety and tolerability of cell culture-derived and egg-derived trivalent influenza vaccines in 3 to < 18-year-old children and adolescents at risk of influenza-related complications. Int J Infect Dis. 2016;49:171–178. doi: 10.1016/j.ijid.2016.06.018. [DOI] [PubMed] [Google Scholar]
- 35.Moro PL, Winiecki S, Lewis P, et al. Surveillance of adverse events after the first trivalent inactivated influenza vaccine produced in mammalian cell culture (Flucelvax®) reported to the Vaccine Adverse Event Reporting System (VAERS), United States, 2013-2015. Vaccine. 2015;33(48):6684–6688. doi: 10.1016/j.vaccine.2015.10.084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Joint Committee on Vaccination and Immunisation. Minutes (October 2018). 2018. http://www.gov.uk/. Accessed 9 Jul 2019.
- 37.Seqirus. Clinical study to evaluate the efficacy, safety and immunogenicity of QIVc in subjects ≥ 2 to < 18 years of age (NCT03165617). 2019.http://clinicaltrials.gov/. Accessed 9 Jul 2019.
- 38.Orsi A, Colomba GME, Pojero F, et al. Trends of influenza B during the 2010-2016 seasons in 2 regions of north and south Italy: the impact of vaccine mismatch on influenza immunisation strategy. Hum Vaccin Immunother. 2018;14(3):523–531. doi: 10.1080/21645515.2017.1342907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Seqirus. Flucelvax (TIVc or QIVc) pregnancy registry (NCT03438487). 2019. http://www.clinicaltrials.gov/. Accessed 9 Jul 2019.
- 40.Centers for Disease Control and Prevention. CDC vaccine price list. 2019. http://www.cdc.gov/. Accessed 9 Jul 2019.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.