

Abstract
Hydrogen sulfide (H2S) is the latest member of the gasotransmitter family and known to play essential roles in cancer pathophysiology. H2S is produced endogenously and can be administered exogenously. Recent studies showed that H2S in cancers has both pro- and antitumor roles. Understanding the difference in the expression and localization of tissue-specific H2S-producing enzymes in healthy and cancer tissues allows us to develop tools for cancer diagnosis and treatment. Urological malignancies are some of the most common cancers in both men and women, and their early detection is vital since advanced cancers are recurrent, metastatic, and often resistant to treatment. This review summarizes the roles of H2S in cancer and looks at current studies investigating H2S activity and expression of H2S-producing enzymes in urinary cancers. We specifically focused on urothelial carcinoma, renal cell carcinoma, and prostate cancer, as they form the majority of newly diagnosed urinary cancers. Recent studies show that besides the physiological activity of H2S in cancer cells, there are patterns between the development and prognosis of urinary cancers and the expression of H2S-producing enzymes and indirectly the H2S levels. Though controversial and not completely understood, studying the expression of H2S-producing enzymes in cancer tissue may represent an avenue for novel diagnostic and therapeutic strategies for addressing urological malignancies.
1. Hydrogen Sulfide
For several centuries, hydrogen sulfide (H2S) was known as a pollutant, but now its physiological and pathophysiological processes are well known. H2S is widely recognized as the third endogenous gasotransmitter after carbon monoxide (CO) and nitric oxide (NO) in mammals and some other species, with similar pathophysiological characteristics [1, 2]. H2S is synthesized endogenously by reverse transsulfidation and oxidation of cysteine [3–6], by three tissue-specific enzymes: cystathionine β-synthase (CBS), cystathionine γ-lyase (CSE), and 3-mercaptopyruvate sulfurtransferase (3-MPST) [3, 4, 7–11]. All of them are cytosolic [12–14], but 3-MPST is also localized in the mitochondria [3, 12, 15]. Upon synthesis in different cell compartments such as in the mitochondria, a free form of H2S can be released into the cytoplasm or be stored inside the cell as bound sulfane sulfur for subsequent release of H2S (Figure 1) [16, 17].
Figure 1.
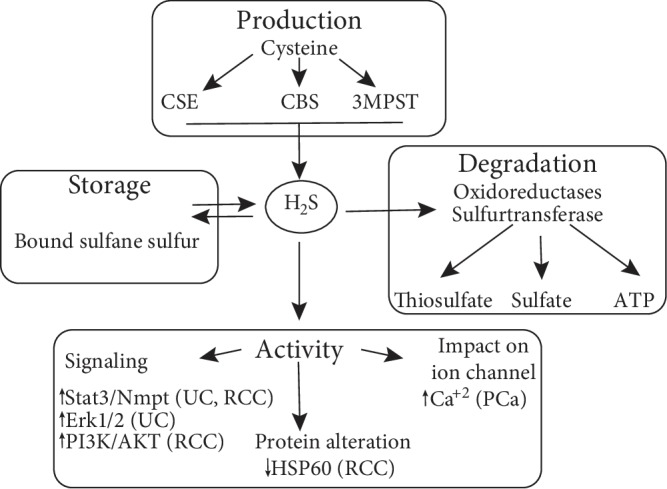
Synthesis, storage, degradation, and activity of H2S, especially in urinary cancers. H2S has roles in different pathways of urinary cancers such as signaling or ion channel. Abbreviations: CBS: cystathionine β-synthase, CSE: cystathionine γ-lyase, 3-MPST: 3-mercaptopyruvate sulfurtransferase, Stat3/Nmpt: signal transducer and activator of transcription 3/nicotinamide phosphoribosyltransferase, HSP60: heat shock protein 60, PI3K/AKT: phosphatidylinositol 3-kinase, UC: urothelial carcinoma, RCC: renal cell cancer, PCa: prostate cancer.
Endogenous H2S is a key signaling molecule in humans and other mammals. It has been detected in many organs, and it is involved in the various physiological and pathophysiological processes [12, 18–20]. H2S is known to play a role in redox homeostasis and antioxidant responses [21–23], angiogenesis [24–30], vasodilation [31], regulation of synaptic transmission [32], inflammatory responses [33], glucose metabolism [34, 35], ATP production [36], and apoptosis and cell proliferation [23, 31, 37–42]. The role that H2S plays in these processes appears to be concentration dependent. The concentration of free H2S in plasma could not be measured in a proper way because it is affected by environmental factors such as pH [43], but an initial study utilizing the methylene blue method reported to be between 50 and160 μM in human and rat serum [44]. However, the recent studies are suggesting that the endogenous concentration of H2S is much less and is between 10 nM and 3 μM [45, 46]. As H2S has a dual effect, at lower concentrations, it has a physiological function in different tissues, whereas at higher concentrations, H2S exerts its toxic effects by reversibly blocking of cytochrome C oxidase and inhibiting the electron transport chain in the mitochondria [47–49]. The catabolism of H2S occurs mainly in the mitochondria by enzymatic pathways such as oxidoreductases and sulfurtransferase that break it into thiosulfate and sulfate. Moreover, oxidation of H2S reduces the FAD prosthetic group, which uses ubiquinone (Q) as an electron acceptor, in electron transport chain which has a role in ATP production (Figure 1) [15, 36, 50–52]. However, under hypoxic conditions, oxidation of H2S in the mitochondria reduces, allowing H2S to accumulate and function as an oxygen sensor [53, 54]. H2S accumulation during hypoxia helps to maintain cell function by upregulating anaerobic metabolic pathways like glycolysis [55] and other cytoprotective pathways [56]. H2S also promotes restoration of the tissue oxygen supply by relaxation of vascular smooth muscles (vasodilation) and also stimulation of endothelial cell proliferation and migration (angiogenesis) [24, 57]. Beside the mitochondrial sulfide oxidation [58], H2S can be oxidized and catabolized by two other minor pathways [9, 59]. The first pathway is the methylation of H2S by thiol S-methyltransferase in the cytosol [60], and the second pathway is an interaction between H2S and methemoglobin that leads to the production of sulfhemoglobin and polysulfides, which can be used as a biomarker for plasma H2S levels [61, 62].
2. H2S in Cancer
Several studies have shown that H2S and its synthases are associated with the pathophysiology of tumors [20, 49, 63–66]. It has been shown that H2S can modulate oxidative stress, interact with free radicals, and activate tumorigenic pathways [39, 61]. Several studies investigated the role and presence of H2S in tumors. The expression of H2S-producing enzymes (CBS, CSE, and 3-MPST) has been studied in various cancers including liver, colon, ovarian, breast, gastric, lung, oral squamous cell carcinoma, and melanoma [42, 49, 67–74]. However, the role and effect of H2S on tumor biology, development, and progression are controversial [75–78]. Previous reviews have adequately summarized that H2S can have pro- or anticancerous effects based on the type of tumor and the involved organ [23, 67]. It is reported that endogenous H2S can have procancerous effects and help the survival of tumors by stimulating angiogenesis along with promoting cell proliferation, metastasis, and drug resistance [32, 49, 67, 79–81]. The anticancerous effects of exogenous H2S administration have been reported for several human cancers [82, 83]. Endogenous H2S can be employed as a biomarker for cancer imaging in mice and for differentiating cancer cells [84, 85]. Several pathways, such as inhibition of proliferation, induction of apoptosis, reduction of NF-κB levels, DNA damage, and modification of the cell cycle, are involved in the anticancer activity of H2S [27, 29, 82, 86].
Similar to endogenous H2S, the effect of exogenous H2S treatment also shows a biphasic dose-dependent response on cancer cells as it does in healthy tissues whereby low concentrations of H2S exhibit a procancerous effect and high concentrations exert an anticancerous effect [65, 67, 82, 83, 87]. The hypoxic environment of solid tumors [88] leads to a higher level of endogenous H2S synthesis [89, 90] and reduces the sulfide detoxification ability of the mitochondria [54, 91], which makes tumors more susceptible to H2S toxicity. However, Malagrinò et al. showed that in hypoxic conditions, the activity of the mitochondrial sulfide-oxidizing pathway of quinone oxidoreductase (SQR) adaptively increased and improved the H2S detoxification of mitochondria [92].
The direct quantification of H2S in tissue samples is a challenge since it has a very short half-life [93]; one study used live fluorescent imaging techniques to visualize the H2S in live cells directly [90]. However, in general, the expression level of H2S-producing enzymes can be used to indirectly show the correlation between H2S and its effects on healthy tissues and tumors [49, 80]. Increased levels of H2S and the upregulation of one or more H2S-synthesizing enzymes in comparison to healthy tissues have been reported in several tumors [49, 71, 72, 80, 94, 95]. It is also quite interesting that these three enzymes are expressed differently according to the type of cancer [67] and hence lend themselves as potential new targets for therapy.
3. H2S in Urinary Cancers
Urinary cancers specifically kidney, urothelial, and prostate are relatively common in developed countries. Prostate cancer [96] is the second most commonly diagnosed cancer in men, and urothelial carcinomas (UCs) [97] are the fourth most common tumors both in men and in women. Kidney cancers are highly lethal, and their incidence is increasing incidentally by the common use of diagnostic tools. It is estimated that more than 300,000 new cases of urinary cancers and 33,429 deaths (excluding prostate cancer) will occur in 2019 in the United States [97]. As such, the role of H2S and the differential expression of H2S-producing enzymes in urinary cancers are of interest, and this review is aimed at summarizing recent evidence on this subject in the context of three common urinary cancers: urothelial cancer, renal cell carcinoma, and prostate cancer.
3.1. Urothelial Cancer
Urothelial carcinoma can be located in the lower (bladder and urethra) or the upper (pyelocaliceal cavities and ureter) urinary tract. Bladder tumors account for 90-95% of UCs and are the most common urinary tract malignancy. Sixty percent of upper tract urothelial cancers are invasive at diagnosis compared with 15-25% of bladder tumors [98]. The high recurrence rate and potential of metastasis are two critical characteristics of bladder cancer [99, 100]. Environmental (smoking and exposure to chemical-occupational toxins) and genetic factors all play a role in the etiology of bladder cancer, as does gender since it is more frequent in men older than 65 years of age [101].
Several studies have highlighted the importance of abnormal redox and cellular signaling in the incidence of bladder cancer [102]. Various reports suggest that alterations in H2S synthesis pathways may increase the risk of bladder cancer [103, 104], suggesting that the modification of these pathways may lead to the development of novel diagnostic and therapeutic approaches for urological cancers [4].
H2S has been detected in bladder homogenates of trout, mice, pigs, rats, and humans [105–109]. In humans, H2S is involved in the control of bladder tone homeostasis [110], as it has previously been shown that exogenous H2S or its substrate, L-cysteine, could decrease the tone of human and rat bladder strips in a dose-dependent manner [107, 108]. All of the H2S-producing enzymes are also found in rat and human bladders, whereas in the mouse, only CSE could be detected [107–109]. The expression of these enzymes in human bladder cancer tissues and cell lines has been investigated. A recent study examined the expression of H2S-producing enzymes in human bladder cancer tissues and compared them to healthy ones. They compared 94 human bladder cancer at different stages/grades and 20 human healthy bladder tissues in term of H2S content as well as the H2S synthases while attempting to find a correlation between the expression of H2S-producing enzymes and the malignant progression of bladder cancer. They showed that H2S content, as well as the expression of CBS, CSE, and 3-MPST, was higher in bladder cancer than in healthy samples. More interestingly, the enzyme expression of all three enzymes was correlated to different stages of bladder cancer. They suggested that this correlation between the malignancy and the expression of H2S enzymes could lead to novel diagnosis and treatment applications [111]. Another recent study also showed, both in vitro and in vivo models, that apoptosis of bladder cancer cell lines or tissues with cisplatin was enhanced after the inhibition of H2S production by propargylglycine (PAG) [23] and was inhibited upon adding the exogenous H2S. These authors suggested the activation of the Erk1/2 signaling pathway and the blockage of mitochondrial apoptosis as the possible mechanisms behind their results [112].
Exogenous H2S administration has also been shown to affect bladder cancer cell lines. The in vitro treatment of the bladder cancer cell line EJ with NaHS enhances cell proliferation and the invasion ability of the cells [113]. Interestingly, these authors also found that the expression of matrix metalloproteinases (MMP) 2 and 9, which are essential for the digestion of collagen IV, was increased in a dose-dependent manner upon the treatment of bladder cancer cells with NaHS. These two enzymes are essential in hydrolyzing the extracellular matrix during the invasion; therefore, H2S might be necessary for the invasion of bladder cancer [113]. In addition, nicotinamide phosphoribosyltransferase (Nampt) is the rate-limiting step of nicotinamide adenine dinucleotide synthesis also increased in some cancers [114]. The signal transducer and activator of transcription 3 (Stat3) is one of the cell signaling molecules of the H2S, and its activation induces Nampt protein expression via a positive feedback loop. A recent study showed that UC is immunoreactive for the enzymatically active phosphor-Stat3 signal transduction pathway and increased the Nampt and CBS protein expression [115]. Overall, bladder cancer appears to present with higher H2S levels in cancer tissue homogenates and increased the expression of H2S-producing enzymes, which suggests that H2S may be essential for bladder cancer progression and growth, especially in the context of the induction of cell proliferation, inhibition of apoptosis, and facilitation of tissue invasion. Further research is needed to establish consistent expression patterns and other cellular mechanisms for potential diagnostic and therapeutic approaches.
3.2. Renal Cell Carcinoma
Renal cell carcinoma (RCC) represents 2-3% of all cancers with the highest incidence in Western countries. The incidence varies globally, with the highest rates in developed countries such as North America and Europe and the lowest rates in Asia and Africa [116]. Over 300,000 men and women are diagnosed with kidney cancer around the world each year, and approximately 150,000 patients will die of the disease [96].
Clear cell renal cell carcinoma (ccRCC), papillary carcinoma, and chromophobe are the common subtypes of RCC [117], although ccRCC accounts for 80% of all RCCs [118]. Because of the lack of early warning signs and the absence of screening tests for people with a higher risk of kidney cancer, more than 30% of patients are at the metastatic stage at the time of diagnosis [119]. Metastatic RCC is highly resistant to systemic chemotherapy and radiation therapy [120, 121].
Inactivation of the Von Hippel-Lindau (VHL) tumor suppressor, which is responsible for the degradation of hypoxia-inducible factor alpha subunits (HIF-1/2α) during normoxia, occurs in 90% of ccRCC cases [122, 123]. As a result, HIF-1/2α subunits are not degraded under normoxic conditions in RCC cells, and the cells become pseudohypoxic [118]. The Warburg effect, which refers to a shift from mitochondrial respiration to glycolysis and production of lactate [124], enhances tumor growth and metastasis in RCC [125]. Using live cell imaging, Sonke et al. have previously shown that VHL-deficient ccRCC cell lines (769-P and 786-O) have significantly higher H2S levels in comparison to ccRCC cells with wild-type VHL (Caki-1). They also showed that the inhibition of H2S-producing enzymes by hydroxylamine (HA), which is an inhibitor of CBS and CSE, and PAG, an inhibitor of CSE, significantly decreases the H2S levels in VHL-deficient ccRCC cell lines and subsequently inhibits their proliferation and metabolic activity. Moreover, this inhibition of H2S synthesis in VHL-deficient ccRCC cell lines results in a twofold reduction in cell survival rate in comparison to untreated cells. Another key finding from this work was that systemic inhibition of H2S enzymes by HA administration in xenografted ccRCC in chicken embryos inhibited their vascularization and the subsequent growth of xenografts, which supports the known angiogenic activity of H2S [79].
Two more recent studies have also evaluated the expression of H2S enzymes in ccRCC. Shackelford et al. compared the expression of CBS in between human benign and Fuhrman grade I-IV ccRCC tissues by using tissue microarray and immunohistochemistry. They showed that CBS expressed weakly in benign tissues and even weaker in Fuhrman grade I ccRCC; however, its expression increased with increasing Fuhrman grades, and CBS expression was the highest in Fuhrman grade IV ccRCC samples [95]. Moreover, the Nmpt expression was correlated with CBS in increasing grade of tumors. Therefore, H2S may play a contributory role in the progression of RCC [95]. Breza et al. also investigated the expression of H2S-producing enzymes in 21 human ccRCC tissues and compared it to the normal/healthy portion of the same kidney sample using microarray and immunohistochemistry. They found that 66% of ccRCC tissue samples exhibited stable expression of CBS, and the remaining samples showed downregulation. CSE was downregulated in all samples except in three where it was unchanged. The expression of 3-MPST was decreased by 70% of ccRCC samples and remained unchanged in 30% of ccRCC samples [121]. These data suggest that the expression of H2S enzymes is heterogeneously regulated in ccRCC. The contradiction between results might be attributed to Shackelford et al. not comparing benign/malignant tissues from the same patient. Breza et al. also showed that, upon induction of apoptosis, the expression of these enzymes was upregulated in the RCC4 cell line (human RCC cell line) and silencing of CBS and CSE expression made the cells resistant to apoptosis [121]. It is possible that endogenous H2S induces apoptosis in ccRCC as it was previously reported with exogenous administration [126–130]. The mechanisms behind RCC progression are not well understood, but it is suggested that knocking down of heat shock protein 60 (HSP60) increases the epithelial to mesenchymal transition and enhances invasion and also disturbs the respiratory complex 1 and triggers reactive oxygen molecules and then DNA methylation for further tumorigenesis [131–133]. Tang et al. results supported that suggestion and showed that HSP60 expression is lower in ccRCC tissues compared to pericancerous tissues [134]. The PI3K/AKT pathway is another important pathway in RCC progression, and it is reported that exogenous H2S inhibits this pathway, and therefore, exogenous H2S could be a novel targeted therapy of RCC [135, 136]. Overall, the expression of H2S enzymes could one day become a new tool for establishing prognosis in patients with RCC. However, further studies are necessary to elucidate the exact role of H2S in RCC and to explain the contradictions between different studies.
3.3. Prostate Cancer
Prostate cancer (PCa) is the second most common cancer in men, with an estimated 1.1 million new cases worldwide in 2012, accounting for 15% of all cancers diagnosed. The incidence of PCa varies widely between different geographical areas, highest in developed countries, mainly due to the use of prostate-specific antigen (PSA) testing and the aging of the population [96]. Surgery, radiotherapy, and androgen deprivation therapies are the primary treatment modalities that are effective, especially in the early stages of the disease [137]. Although a physical exam and the serum PSA test are commonly used to screen and detect for prostate cancer; their utility is ineffective in diagnosing early stages of prostate cancer.
The relationship between H2S activity and prostate cancer has been reviewed previously [138]. The expression of H2S-producing enzymes was compared between cancerous and healthy prostate tissues [108, 139]. Endogenous H2S and all three enzymes (CBS, CSE, and 3-MPST) have been demonstrated in healthy and prostate cancer. CSE has been shown to have a higher expression in the smooth muscle layer of the prostate cancer samples [94]. However, in another study, they could not detect the expression of 3-MPST in both normal and cancerous prostate tissues, but they showed that CSE was significantly downregulated in prostate cancer, whereas CBS was not changed in each sample. This study also showed that antiandrogen-resistant prostate cancer cells express less CSE and have lower H2S content in comparison to the parental cell line [140].
Moreover, new evidence suggests that H2S-releasing molecules could be effective in the treatment of chemotherapy-resistant prostate cancers [141]. The stromal part of the prostate tissue and the stromal cell line showed average to high CSE expression [139]. In addition, both CBS and CSE are present in mouse prostate cancers, unrelated to androgen dependency, and in vitro work showed that CSE is the main contributor to H2S production in prostate cancer cell lines (PC-3). The critical role of CSE was confirmed upon finding that aged CSE knockout mice have higher cell proliferation and significantly less H2S production in the prostate [142]. Interestingly, the androgen-dependent prostate cell line showed the highest expression of CBS and CSE, and their expression was downregulated upon dihydrotestosterone treatment [139]. These data suggest that CSE may be a potential therapeutic target and diagnostic tool for prostate cancer.
As mentioned earlier, thiosulfate is the stable breakdown product of H2S in the mitochondria that can be tracked in the urine. Therefore, the thiosulfate level in urine can be an indicator of exposure to H2S or disruption in the breakdown process. Chwatko et al. investigated urinary thiosulfate levels amongst the malignant in comparison to benign prostate hyperplasia (BPH) patients and healthy volunteers. They also found that the urine level of thiosulfate in malignant prostate cancer patients was 50 times higher than the healthy volunteers and five times higher than the BPH patients, and also, there was a positive correlation between the size of the prostate and the urine level of thiosulfate in comparison between the BPH and the control group [143]. In the nude mouse model of human prostate cancer, the plasma concentration of cysteine was significantly decreased after advanced tumor growth [144]. Contrary to these results, five years after prostatectomy, cysteine, homocysteine, and cystathionine were found to be higher in the urine of recurrent prostate cancer patients in comparison to recurrence-free patients [145]. Recent studies showed that methionine catabolism [146], and increased level of cystathionine [147] and sarcosine (N-methylglycine), a by-product of methionine catabolism [148], in urine correlated with prostate cancer stage. In addition, recent data suggest that neuroendocrine-like differentiation of prostate cancer (LNCaP) cells contributes to the androgen-independent growth [149, 150]. The expression and activity of CSE and CBS, in LNCaP cell, are much more than those in healthy prostatic epithelial cells [139]. The H2S donors, NaHS and Na2S, further enhance the upregulated calcium channels in the LNCaP cells [151]. Overall, it appears that cysteine, homocysteine, cystathionine, and sarcosine could all potentially be biomarkers for prostate cancer.
4. Conclusion
Despite significant research efforts in recent years, the role of H2S in the context of cancer pathophysiology remains controversial (Table 1). Several studies have partially elucidated the vital role of H2S activity, which plays a different role in urological malignancies (Figure 1). Interestingly, the expression patterns of H2S-producing enzymes appear to be contradictory, depending upon the subtype of cancer, which was evaluated and in fact, may be tissue dependent. However, these studies, as mentioned earlier, lay the groundwork for future work that may lead to the development of new diagnostic tools for detecting urinary cancers in earlier stages. Moreover, pharmacological modulation of H2S synthetic pathways and exogenous administration of donor molecules may one day provide us with additional therapeutic avenues in treating patients with urological malignancies.
Table 1.
Summary of H2S and its producing enzymes in three common urinary cancers.
| H2S highlights | |
|---|---|
| Urothelial carcinoma | (i) Expressions of H2S and its synthases are higher in cancer tissue [111]. (ii) H2S protects the bladder cancer against apoptosis [112]. (iii) H2S increases the cell proliferation, invasion, and metastasis of bladder cancer [113]. |
|
| |
| Renal cell carcinoma | (i) Enhanced expression of H2S in ccRCC due to VHL deficiency improves the survival, growth, and metastasis [79]. (ii) Controversial reports about the comparative expression of H2S enzymes [95, 121]. (iii) H2S contributes to the induction of apoptosis in RCC [121]. |
|
| |
| Prostate cancer | (i) H2S enzymes are expressed in the prostate [107]. (ii) CSE is the main H2S-producing enzyme in the prostate [107, 139, 142]. (iii) CSE is downregulated in prostate cancer [140]. |
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- 1.Fagone P., Mazzon E., Bramanti P., Bendtzen K., Nicoletti F. Gasotransmitters and the immune system: mode of action and novel therapeutic targets. European Journal of Pharmacology. 2018;834:92–102. doi: 10.1016/j.ejphar.2018.07.026. [DOI] [PubMed] [Google Scholar]
- 2.Wang R. Gasotransmitters: growing pains and joys. Trends in Biochemical Sciences. 2014;39(5):227–232. doi: 10.1016/j.tibs.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 3.Kimura H. Hydrogen sulfide: its production, release and functions. Amino Acids. 2011;41(1):113–121. doi: 10.1007/s00726-010-0510-x. [DOI] [PubMed] [Google Scholar]
- 4.Li L., Rose P., Moore P. K. Hydrogen sulfide and cell signaling. Annual Review of Pharmacology and Toxicology. 2011;51(1):169–187. doi: 10.1146/annurev-pharmtox-010510-100505. [DOI] [PubMed] [Google Scholar]
- 5.Ida T., Sawa T., Ihara H., et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proceedings of the National Academy of Sciences of the United States of America. 2014;111(21):7606–7611. doi: 10.1073/pnas.1321232111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yadav P. K., Martinov M., Vitvitsky V., et al. Biosynthesis and reactivity of cysteine persulfides in signaling. Journal of the American Chemical Society. 2016;138(1):289–299. doi: 10.1021/jacs.5b10494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kabil O., Banerjee R. Enzymology of H2S biogenesis, decay and signaling. Antioxidants & Redox Signaling. 2014;20(5):770–782. doi: 10.1089/ars.2013.5339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Olson K. R. H2S and polysulfide metabolism: conventional and unconventional pathways. Biochemical Pharmacology. 2018;149:77–90. doi: 10.1016/j.bcp.2017.12.010. [DOI] [PubMed] [Google Scholar]
- 9.Giuffrè A., Vicente J. B. Hydrogen sulfide biochemistry and interplay with other gaseous mediators in mammalian physiology. Oxidative Medicine and Cellular Longevity. 2018;2018:31. doi: 10.1155/2018/6290931.6290931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pandey S. Hydrogen sulfide: a new node in the abscisic acid-dependent guard cell signaling network? Plant Physiology. 2014;166(4):1680–1681. doi: 10.1104/pp.114.251686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hancock J. T., Whiteman M. Hydrogen sulfide and cell signaling: team player or referee? Plant Physiology and Biochemistry. 2014;78:37–42. doi: 10.1016/j.plaphy.2014.02.012. [DOI] [PubMed] [Google Scholar]
- 12.Wang R. Physiological implications of hydrogen sulfide: a whiff exploration that blossomed. Physiological Reviews. 2012;92(2):791–896. doi: 10.1152/physrev.00017.2011. [DOI] [PubMed] [Google Scholar]
- 13.Paul B. D., Snyder S. H. H2S signalling through protein sulfhydration and beyond. Nature Reviews Molecular Cell Biology. 2012;13(8):499–507. doi: 10.1038/nrm3391. [DOI] [PubMed] [Google Scholar]
- 14.Liu Y., Yang R., Liu X., et al. Hydrogen sulfide maintains mesenchymal stem cell function and bone homeostasis via regulation of Ca2+ channel sulfhydration. Cell Stem Cell. 2014;15(1):66–78. doi: 10.1016/j.stem.2014.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kabil O., Banerjee R. Redox biochemistry of hydrogen sulfide. The Journal of Biological Chemistry. 2010;285(29):21903–21907. doi: 10.1074/jbc.R110.128363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shen X., Carlstrom M., Borniquel S., Jadert C., Kevil C. G., Lundberg J. O. Microbial regulation of host hydrogen sulfide bioavailability and metabolism. Free Radical Biology & Medicine. 2013;60:195–200. doi: 10.1016/j.freeradbiomed.2013.02.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Whiteman M., Le Trionnaire S., Chopra M., Fox B., Whatmore J. Emerging role of hydrogen sulfide in health and disease: critical appraisal of biomarkers and pharmacological tools. Clinical Science. 2011;121(11):459–488. doi: 10.1042/CS20110267. [DOI] [PubMed] [Google Scholar]
- 18.Kuo M. M., Kim D. H., Jandu S., et al. MPST but not CSE is the primary regulator of hydrogen sulfide production and function in the coronary artery. American Journal of Physiology-Heart and Circulatory Physiology. 2016;310(1):H71–H79. doi: 10.1152/ajpheart.00574.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kashfi K., Kodela R., Nath N., Chattopadhyay M., Nesbitt D. E., Velázquez-Martínez C. A. Hydrogen sulfide-releasing naproxen suppresses colon cancer cell growth and inhibits NF-κB signaling. Drug Design, Development and Therapy. 2015;9, article 4873 doi: 10.2147/dddt.s91116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hellmich M. R., Szabo C. Hydrogen sulfide and cancer. Handbook of Experimental Pharmacology. 2015;230:233–241. doi: 10.1007/978-3-319-18144-8_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ahn H., Lee E., Kim K., Lee C. Effect of glutathione and its related enzymes on chemosensitivity of renal cell carcinoma and bladder carcinoma cell lines. The Journal of Urology. 1994;151(1):263–267. doi: 10.1016/S0022-5347(17)34929-7. [DOI] [PubMed] [Google Scholar]
- 22.Simic T., Savic-Radojevic A., Pljesa-Ercegovac M., Matic M., Mimic-Oka J. Glutathione S-transferases in kidney and urinary bladder tumors. Nature Reviews Urology. 2009;6(5):281–289. doi: 10.1038/nrurol.2009.49. [DOI] [PubMed] [Google Scholar]
- 23.Ianaro A., Cirino G. Hydrogen sulfide pathway and cancer. Brain Metastases from Primary Tumors, Volume 3. 2016;3:133–144. doi: 10.1016/b978-0-12-803508-5.00009-3. [DOI] [Google Scholar]
- 24.Coletta C., Papapetropoulos A., Erdelyi K., et al. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(23):9161–9166. doi: 10.1073/pnas.1202916109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pupo E., Fiorio Pla A., Avanzato D., et al. Hydrogen sulfide promotes calcium signals and migration in tumor-derived endothelial cells. Free Radical Biology and Medicine. 2011;51(9):1765–1773. doi: 10.1016/j.freeradbiomed.2011.08.007. [DOI] [PubMed] [Google Scholar]
- 26.Davis S. L., Fadel P. J., Cui J., Thomas G. D., Crandall C. G. Skin blood flow influences near-infrared spectroscopy-derived measurements of tissue oxygenation during heat stress. Journal of Applied Physiology. 2006;100(1):221–224. doi: 10.1152/japplphysiol.00867.2005. [DOI] [PubMed] [Google Scholar]
- 27.Cai W.-J., Wang M.-J., Ju L.-H., Wang C., Zhu Y.-C. Hydrogen sulfide induces human colon cancer cell proliferation: role of Akt, ERK and p21. Cell Biology International. 2010;34(6):565–572. doi: 10.1042/CBI20090368. [DOI] [PubMed] [Google Scholar]
- 28.Bellingham G. A., Smith R. S., Morley-Forster P., Murkin J. M. Use of near infrared spectroscopy to detect impaired tissue oxygen saturation in patients with complex regional pain syndrome type 1. Canadian Journal of Anaesthesia. 2014;61(6):563–570. doi: 10.1007/s12630-014-0140-y. [DOI] [PubMed] [Google Scholar]
- 29.Cao Q., Zhang L., Yang G., Xu C., Wang R. Butyrate-stimulated H2S production in colon cancer cells. Antioxidants & Redox Signaling. 2010;12(9):1101–1109. doi: 10.1089/ars.2009.2915. [DOI] [PubMed] [Google Scholar]
- 30.Kimura Y., Goto Y.-I., Kimura H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxidants & Redox Signaling. 2010;12(1):1–13. doi: 10.1089/ars.2008.2282. [DOI] [PubMed] [Google Scholar]
- 31.Yang G., Wu L., Jiang B., et al. H2S as a physiologic vasorelaxant: hypertension in mice with deletion of Cystathionine γ-Lyase. Science. 2008;322(5901):587–590. doi: 10.1126/science.1162667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ma D. K., Vozdek R., Bhatla N., Horvitz H. R. CYSL-1 Interacts with the O2-Sensing Hydroxylase EGL-9 to Promote H2S-Modulated Hypoxia-Induced Behavioral Plasticity in C. elegans. Neuron. 2012;73(5):925–940. doi: 10.1016/j.neuron.2011.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bhatia M. Hydrogen sulfide and substance P in inflammation. Antioxidants & Redox Signaling. 2010;12(10):1191–1202. doi: 10.1089/ars.2009.2927. [DOI] [PubMed] [Google Scholar]
- 34.Xue R., Hao D.-D., Sun J.-P., et al. Hydrogen sulfide treatment promotes glucose uptake by increasing insulin receptor sensitivity and ameliorates kidney lesions in type 2 diabetes. Antioxidants & Redox Signaling. 2013;19(1):5–23. doi: 10.1089/ars.2012.5024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yang C. T., Chen L., Xu S., Day J. J., Li X., Xian M. Recent development of hydrogen sulfide releasing/stimulating reagents and their potential applications in cancer and glycometabolic disorders. Frontiers in Pharmacology. 2017;8 doi: 10.3389/fphar.2017.00664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fu M., Zhang W., Wu L., Yang G., Li H., Wang R. Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(8):2943–2948. doi: 10.1073/pnas.1115634109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lobb I., Davison M., Carter D., et al. Hydrogen sulfide treatment mitigates renal allograft ischemia-reperfusion injury during cold storage and improves early transplant kidney function and survival following allogeneic renal transplantation. The Journal of Urology. 2015;194(6):1806–1815. doi: 10.1016/j.juro.2015.07.096. [DOI] [PubMed] [Google Scholar]
- 38.Yang G.-D., Wang R. H(2)S and cellular proliferation and apoptosis. Sheng Li Xue Bao. 2007;59(2):133–140. [PubMed] [Google Scholar]
- 39.Szabó C. Hydrogen sulphide and its therapeutic potential. Nature Reviews Drug Discovery. 2007;6(11):917–935. doi: 10.1038/nrd2425. [DOI] [PubMed] [Google Scholar]
- 40.Guo R., Lin J., Xu W., et al. Hydrogen sulfide attenuates doxorubicin-induced cardiotoxicity by inhibition of the p38 MAPK pathway in H9c2 cells. International Journal of Molecular Medicine. 2013;31(3):644–650. doi: 10.3892/ijmm.2013.1246. [DOI] [PubMed] [Google Scholar]
- 41.Du J., Huang Y., Yan H., et al. Hydrogen sulfide suppresses oxidized low-density lipoprotein (Ox-LDL)-stimulated monocyte chemoattractant protein 1 generation from macrophages via the nuclear factor κB (NF-κB) pathway. The Journal of Biological Chemistry. 2014;289(14):9741–9753. doi: 10.1074/jbc.M113.517995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Panza E., de Cicco P., Armogida C., et al. Role of the cystathionine γ lyase/hydrogen sulfide pathway in human melanoma progression. Pigment Cell & Melanoma Research. 2015;28(1):61–72. doi: 10.1111/pcmr.12312. [DOI] [PubMed] [Google Scholar]
- 43.Whitfield N. L., Kreimier E. L., Verdial F. C., Skovgaard N., Olson K. R. Reappraisal of H2S/sulfide concentration in vertebrate blood and its potential significance in ischemic preconditioning and vascular signaling. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2008;294(6):R1930–R1937. doi: 10.1152/ajpregu.00025.2008. [DOI] [PubMed] [Google Scholar]
- 44.Whiteman M., Moore P. K. Hydrogen sulfide and the vasculature: a novel vasculoprotective entity and regulator of nitric oxide bioavailability? Journal of Cellular and Molecular Medicine. 2009;13(3):488–507. doi: 10.1111/j.1582-4934.2009.00645.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ishigami M., Hiraki K., Umemura K., Ogasawara Y., Ishii K., Kimura H. A source of hydrogen sulfide and a mechanism of its release in the brain. Antioxidants & Redox Signaling. 2009;11(2):205–214. doi: 10.1089/ars.2008.2132. [DOI] [PubMed] [Google Scholar]
- 46.Levitt M. D., Abdel-Rehim M. S., Furne J. Free and acid-labile hydrogen sulfide concentrations in mouse tissues: anomalously high free hydrogen sulfide in aortic tissue. Antioxidants & Redox Signaling. 2011;15(2):373–378. doi: 10.1089/ars.2010.3525. [DOI] [PubMed] [Google Scholar]
- 47.Cooper C. E., Brown G. C. The inhibition of mitochondrial cytochrome oxidase by the gases carbon monoxide, nitric oxide, hydrogen cyanide and hydrogen sulfide: chemical mechanism and physiological significance. Journal of Bioenergetics and Biomembranes. 2008;40(5):533–539. doi: 10.1007/s10863-008-9166-6. [DOI] [PubMed] [Google Scholar]
- 48.Baskar R., Bian J. Hydrogen sulfide gas has cell growth regulatory role. European Journal of Pharmacology. 2011;656(1–3):5–9. doi: 10.1016/j.ejphar.2011.01.052. [DOI] [PubMed] [Google Scholar]
- 49.Szabo C., Coletta C., Chao C., et al. Tumor-derived hydrogen sulfide, produced by cystathionine-β-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(30):12474–12479. doi: 10.1073/pnas.1306241110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hildebrandt T. M., Grieshaber M. K. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. The FEBS Journal. 2008;275(13):3352–3361. doi: 10.1111/j.1742-4658.2008.06482.x. [DOI] [PubMed] [Google Scholar]
- 51.Kashfi K., Olson K. R. Biology and therapeutic potential of hydrogen sulfide and hydrogen sulfide-releasing chimeras. Biochemical Pharmacology. 2013;85(5):689–703. doi: 10.1016/j.bcp.2012.10.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jackson M. R., Melideo S. L., Jorns M. S. Human sulfide:quinone oxidoreductase catalyzes the first step in hydrogen sulfide metabolism and produces a sulfane sulfur metabolite. Biochemistry. 2012;51(34):6804–6815. doi: 10.1021/bi300778t. [DOI] [PubMed] [Google Scholar]
- 53.Lagoutte E., Mimoun S., Andriamihaja M., Chaumontet C., Blachier F., Bouillaud F. Oxidation of hydrogen sulfide remains a priority in mammalian cells and causes reverse electron transfer in colonocytes. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2010;1797(8):1500–1511. doi: 10.1016/j.bbabio.2010.04.004. [DOI] [PubMed] [Google Scholar]
- 54.Olson K. R. Hydrogen sulfide as an oxygen sensor. Antioxidants & Redox Signaling. 2015;22(5):377–397. doi: 10.1089/ars.2014.5930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mustafa A. K., Gadalla M. M., Sen N., et al. H2S signals through protein S-sulfhydration. Science Signaling. 2009;2(96, article ra72) doi: 10.1126/scisignal.2000464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sen N., Paul B. D., Gadalla M. M., et al. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Molecular Cell. 2012;45(1):13–24. doi: 10.1016/j.molcel.2011.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Szabó C., Papapetropoulos A. Hydrogen sulphide and angiogenesis: mechanisms and applications. British Journal of Pharmacology. 2011;164(3):853–865. doi: 10.1111/j.1476-5381.2010.01191.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Levitt M. D., Furne J., Springfield J., Suarez F., DeMaster E. Detoxification of hydrogen sulfide and methanethiol in the cecal mucosa. The Journal of Clinical Investigation. 1999;104(8):1107–1114. doi: 10.1172/JCI7712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Weisiger R. A., Pinkus L. M., Jakoby W. B. Thiol S-methyltransferase: suggested role in detoxication of intestinal hydrogen sulfide. Biochemical Pharmacology. 1980;29(20):2885–2887. doi: 10.1016/0006-2952(80)90029-5. [DOI] [PubMed] [Google Scholar]
- 60.Polhemus D. J., Lefer D. J. Emergence of hydrogen sulfide as an endogenous gaseous signaling molecule in cardiovascular disease. Circulation Research. 2014;114(4):730–737. doi: 10.1161/CIRCRESAHA.114.300505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kajimura M., Fukuda R., Bateman R. M., Yamamoto T., Suematsu M. Interactions of multiple gas-transducing systems: hallmarks and uncertainties of CO, NO, and H2S gas biology. Antioxidants & Redox Signaling. 2010;13(2):157–192. doi: 10.1089/ars.2009.2657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Vitvitsky V., Yadav P. K., An S., Seravalli J., Cho U.-S., Banerjee R. Structural and mechanistic insights into hemoglobin-catalyzed hydrogen sulfide oxidation and the fate of polysulfide products. The Journal of Biological Chemistry. 2017;292(13):5584–5592. doi: 10.1074/jbc.M117.774943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhang L., Qi Q., Yang J., et al. An anticancer role of hydrogen sulfide in human gastric cancer cells. Oxidative Medicine and Cellular Longevity. 2015;2015:8. doi: 10.1155/2015/636410.636410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lv M., Li Y., Ji M.-H., Zhuang M., Tang J.-H. Inhibition of invasion and epithelial-mesenchymal transition of human breast cancer cells by hydrogen sulfide through decreased phospho-p38 expression. Molecular Medicine Reports. 2014;10(1):341–346. doi: 10.3892/mmr.2014.2161. [DOI] [PubMed] [Google Scholar]
- 65.Hellmich M. R., Coletta C., Chao C., Szabo C. The therapeutic potential of cystathionine β-synthetase/hydrogen sulfide inhibition in cancer. Antioxidants & Redox Signaling. 2015;22(5):424–448. doi: 10.1089/ars.2014.5933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Cao X., Ding L., Xie Z., et al. A review of hydrogen sulfide synthesis, metabolism, and measurement: is modulation of hydrogen sulfide a novel therapeutic for cancer? Antioxidants & Redox Signaling. 2019;31(1):1–38. doi: 10.1089/ars.2017.7058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wu D., Si W., Wang M., Lv S., Ji A., Li Y. Hydrogen sulfide in cancer: friend or foe? Nitric Oxide. 2015;50:38–45. doi: 10.1016/j.niox.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 68.Kaium M. A., Liu Y., Zhu Q., et al. H2S donor, S-propargyl-cysteine, increases CSE in SGC-7901 and cancer-induced mice: evidence for a novel anti-cancer effect of endogenous H2S? PLoS One. 2011;6(6, article e20525) doi: 10.1371/journal.pone.0020525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ramasamy S., Singh S., Taniere P., Langman M. J. S., Eggo M. C. Sulfide-detoxifying enzymes in the human colon are decreased in cancer and upregulated in differentiation. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2006;291(2):G288–G296. doi: 10.1152/ajpgi.00324.2005. [DOI] [PubMed] [Google Scholar]
- 70.Chattopadhyay M., Kodela R., Nath N., Barsegian A., Boring D., Kashfi K. Hydrogen sulfide-releasing aspirin suppresses NF-κB signaling in estrogen receptor negative breast cancer cells in vitro and in vivo. Biochemical Pharmacology. 2012;83(6):723–732. doi: 10.1016/j.bcp.2011.12.019. [DOI] [PubMed] [Google Scholar]
- 71.Bhattacharyya S., Saha S., Giri K., et al. Cystathionine beta-synthase (CBS) contributes to advanced ovarian cancer progression and drug resistance. PLoS One. 2013;8(11, article e79167) doi: 10.1371/journal.pone.0079167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Módis K., Coletta C., Asimakopoulou A., et al. Effect of S-adenosyl-l-methionine (SAM), an allosteric activator of cystathionine-β-synthase (CBS) on colorectal cancer cell proliferation and bioenergetics in vitro. Nitric Oxide. 2014;41:146–156. doi: 10.1016/j.niox.2014.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Szczesny B., Marcatti M., Zatarain J. R., et al. Inhibition of hydrogen sulfide biosynthesis sensitizes lung adenocarcinoma to chemotherapeutic drugs by inhibiting mitochondrial DNA repair and suppressing cellular bioenergetics. Scientific Reports. 2016;6(1) doi: 10.1038/srep36125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Meram A. T., Chen J., Patel S., et al. Hydrogen sulfide is increased in oral squamous cell carcinoma compared to adjacent benign oral mucosae. Anticancer Research. 2018;38(7):3843–3852. doi: 10.21873/anticanres.12668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lee Z.-W., Teo X.-Y., Tay E. Y.-W., et al. Utilizing hydrogen sulfide as a novel anti‐cancer agent by targeting cancer glycolysis and pH imbalance. British Journal of Pharmacology. 2014;171(18):4322–4336. doi: 10.1111/bph.12773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ma Z., Bi Q., Wang Y. Hydrogen sulfide accelerates cell cycle progression in oral squamous cell carcinoma cell lines. Oral Diseases. 2015;21(2):156–162. doi: 10.1111/odi.12223. [DOI] [PubMed] [Google Scholar]
- 77.Szabo C., Hellmich M. R. Endogenously produced hydrogen sulfide supports tumor cell growth and proliferation. Cell Cycle. 2014;12(18):2915–2916. doi: 10.4161/cc.26064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Renga B. Hydrogen sulfide generation in mammals: the molecular biology of cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE) Inflammation & Allergy - Drug Targets. 2011;10(2):85–91. doi: 10.2174/187152811794776286. [DOI] [PubMed] [Google Scholar]
- 79.Sonke E., Verrydt M., Postenka C. O., et al. Inhibition of endogenous hydrogen sulfide production in clear-cell renal cell carcinoma cell lines and xenografts restricts their growth, survival and angiogenic potential. Nitric Oxide. 2015;49:26–39. doi: 10.1016/j.niox.2015.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Jurkowska H., Placha W., Nagahara N., Wróbel M. The expression and activity of cystathionine-γ-lyase and 3-mercaptopyruvate sulfurtransferase in human neoplastic cell lines. Amino Acids. 2011;41(1):151–158. doi: 10.1007/s00726-010-0606-3. [DOI] [PubMed] [Google Scholar]
- 81.Untereiner A. A., Pavlidou A., Druzhyna N., Papapetropoulos A., Hellmich M. R., Szabo C. Drug resistance induces the upregulation of H2S-producing enzymes in HCT116 colon cancer cells. Biochemical Pharmacology. 2018;149:174–185. doi: 10.1016/j.bcp.2017.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chattopadhyay M., Nath N., Kodela R., et al. Hydrogen sulfide-releasing aspirin inhibits the growth of leukemic Jurkat cells and modulates β-catenin expression. Leukemia Research. 2013;37(10):1302–1308. doi: 10.1016/j.leukres.2013.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Chattopadhyay M., Kodela R., Nath N., et al. Hydrogen sulfide-releasing NSAIDs inhibit the growth of human cancer cells: a general property and evidence of a tissue type-independent effect. Biochemical Pharmacology. 2012;83(6):715–722. doi: 10.1016/j.bcp.2011.12.018. [DOI] [PubMed] [Google Scholar]
- 84.Zhang C., Zhang Q.-Z., Zhang K., et al. Dual-biomarker-triggered fluorescence probes for differentiating cancer cells and revealing synergistic antioxidant effects under oxidative stress. Chemical Science. 2019;10(7):1945–1952. doi: 10.1039/C8SC03781G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zhang K., Zhang J., Xi Z., et al. A new H2S-specific near-infrared fluorescence-enhanced probe that can visualize the H2S level in colorectal cancer cells in mice. Chemical Science. 2017;8(4):2776–2781. doi: 10.1039/C6SC05646F. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Rose P. Hydrogen sulfide protects colon cancer cells from chemopreventative agent β-phenylethyl isothiocyanate induced apoptosis. World Journal of Gastroenterology. 2005;11(26):3990–3997. doi: 10.3748/wjg.v11.i26.3990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wu D., Li M., Tian W., et al. Hydrogen sulfide acts as a double-edged sword in human hepatocellular carcinoma cells through EGFR/ERK/MMP-2 and PTEN/AKT signaling pathways. Scientific Reports. 2017;7 doi: 10.1038/s41598-017-05457-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Muz B., de la Puente P., Azab F., Azab A. K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia. 2015;3:83–92. doi: 10.2147/hp.s93413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Takano N., Peng Y.-J., Kumar G. K., et al. Hypoxia-inducible factors regulate human and rat cystathionine β-synthase gene expression. The Biochemical Journal. 2014;458(2):203–211. doi: 10.1042/BJ20131350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Wang M., Guo Z., Wang S. Regulation of cystathionine γ-lyase in mammalian cells by hypoxia. Biochemical Genetics. 2014;52(1–2):29–37. doi: 10.1007/s10528-013-9624-7. [DOI] [PubMed] [Google Scholar]
- 91.Wu B., Teng H., Zhang L., et al. Interaction of hydrogen sulfide with oxygen sensing under hypoxia. Oxidative Medicine and Cellular Longevity. 2015;2015:9. doi: 10.1155/2015/758678.758678 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Malagrinò F., Zuhra K., Mascolo L., et al. Hydrogen sulfide oxidation: adaptive changes in mitochondria of SW480 colorectal cancer cells upon exposure to hypoxia. Oxidative Medicine and Cellular Longevity. 2019;2019:11. doi: 10.1155/2019/8102936.8102936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Vitvitsky V., Kabil O., Banerjee R. High turnover rates for hydrogen sulfide allow for rapid regulation of its tissue concentrations. Antioxidants & Redox Signaling. 2012;17(1):22–31. doi: 10.1089/ars.2011.4310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sen S., Kawahara B., Gupta D., et al. Role of cystathionine β-synthase in human breast cancer. Free Radical Biology and Medicine. 2015;86:228–238. doi: 10.1016/j.freeradbiomed.2015.05.024. [DOI] [PubMed] [Google Scholar]
- 95.Shackelford R. E., Abdulsattar J., Wei E. X., Cotelingam J., Coppola D., Herrera G. A. Increased nicotinamide phosphoribosyltransferase and Cystathionine-β-Synthase in renal oncocytomas, renal urothelial carcinoma, and renal clear cell carcinoma. Anticancer Research. 2017;37(7):3423–3427. doi: 10.21873/anticanres.11709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ferlay J., Soerjomataram I., Dikshit R., et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. International Journal of Cancer. 2015;136(5):E359–E386. doi: 10.1002/ijc.29210. [DOI] [PubMed] [Google Scholar]
- 97.Siegel R. L., Miller K. D., Jemal A. Cancer statistics, 2019. CA: A Cancer Journal for Clinicians. 2018;69(1):7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 98.Margulis V., Shariat S. F., Matin S. F., et al. Outcomes of radical nephroureterectomy: a series from the Upper Tract Urothelial Carcinoma Collaboration. Cancer. 2009;115(6):1224–1233. doi: 10.1002/cncr.24135. [DOI] [PubMed] [Google Scholar]
- 99.Krishna S. R., Konety B. R. Current concepts in the management of muscle invasive bladder cancer. Indian Journal of Surgical Oncology. 2017;8(1):74–81. doi: 10.1007/s13193-016-0586-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Zhang X., Han C., He J. Recent advances in the diagnosis and management of bladder cancer. Cell Biochemistry and Biophysics. 2015;73(1):11–15. doi: 10.1007/s12013-015-0632-6. [DOI] [PubMed] [Google Scholar]
- 101.Althunayan A., Kassouf W. Asymptomatic microscopic hematuria: clinical significance and evaluation. Current Medical Literature. 2011;17 [Google Scholar]
- 102.Wallerand H., Reiter R. R., Ravaud A. Molecular targeting in the treatment of either advanced or metastatic bladder cancer or both according to the signalling pathways. Current Opinion in Urology. 2008;18(5):524–532. doi: 10.1097/MOU.0b013e3283097889. [DOI] [PubMed] [Google Scholar]
- 103.Chung C.-J., Pu Y.-S., Su C.-T., et al. Polymorphisms in one-carbon metabolism pathway genes, urinary arsenic profile, and urothelial carcinoma. Cancer Causes & Control. 2010;21(10):1605–1613. doi: 10.1007/s10552-010-9589-3. [DOI] [PubMed] [Google Scholar]
- 104.Moore L. E., Malats N., Rothman N., et al. Polymorphisms in one‐carbon metabolism and trans‐sulfuration pathway genes and susceptibility to bladder cancer. International Journal of Cancer. 2007;120(11):2452–2458. doi: 10.1002/ijc.22565. [DOI] [PubMed] [Google Scholar]
- 105.Dombkowski R. A. Hydrogen sulfide mediates hypoxia-induced relaxation of trout urinary bladder smooth muscle. The Journal of Experimental Biology. 2006;209(16):3234–3240. doi: 10.1242/jeb.02376. [DOI] [PubMed] [Google Scholar]
- 106.Fernandes V. S., Ribeiro A. S. F., Barahona M. V., et al. Hydrogen sulfide mediated inhibitory neurotransmission to the pig bladder neck: role of KATP channels, sensory nerves and calcium signaling. The Journal of Urology. 2013;190(2):746–756. doi: 10.1016/j.juro.2013.02.103. [DOI] [PubMed] [Google Scholar]
- 107.Fusco F., d’Emmanuele di Villa Bianca R., Mitidieri E., Cirino G., Sorrentino R., Mirone V. Sildenafil Effect on the human bladder involves the L-cysteine/hydrogen sulfide pathway: a novel mechanism of action of phosphodiesterase type 5 inhibitors. European Urology. 2012;62(6):1174–1180. doi: 10.1016/j.eururo.2012.07.025. [DOI] [PubMed] [Google Scholar]
- 108.Gai J.-W., Wahafu W., Guo H., et al. Further evidence of endogenous hydrogen sulphide as a mediator of relaxation in human and rat bladder. Asian Journal of Andrology. 2013;15(5):692–696. doi: 10.1038/aja.2013.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Matsunami M., Miki T., Nishiura K., et al. Involvement of the endogenous hydrogen sulfide/Cav3.2 T‐type Ca2+ channel pathway in cystitis‐related bladder pain in mice. British Journal of Pharmacology. 2012;167(4):917–928. doi: 10.1111/j.1476-5381.2012.02060.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.de Groat W. C., Yoshimura N. Pharmacology of the lower urinary tract. Annual Review of Pharmacology and Toxicology. 2001;41(1):691–721. doi: 10.1146/annurev.pharmtox.41.1.691. [DOI] [PubMed] [Google Scholar]
- 111.Gai J.-W., Qin W., Liu M., et al. Expression profile of hydrogen sulfide and its synthases correlates with tumor stage and grade in urothelial cell carcinoma of bladder. Urologic Oncology: Seminars and Original Investigations. 2016;34(4):166.e15–166.e20. doi: 10.1016/j.urolonc.2015.06.020. [DOI] [PubMed] [Google Scholar]
- 112.Wahafu W., Gai J., Song L., et al. Increased H2S and its synthases in urothelial cell carcinoma of the bladder, and enhanced cisplatin-induced apoptosis following H2S inhibition in EJ cells. Oncology Letters. 2018;15 doi: 10.3892/ol.2018.8373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Liu H., Chang J., Zhao Z., Li Y., Hou J. Effects of exogenous hydrogen sulfide on the proliferation and invasion of human bladder cancer cells. Journal of Cancer Research and Therapeutics. 2017;13(5):829–832. doi: 10.4103/jcrt.JCRT_423_17. [DOI] [PubMed] [Google Scholar]
- 114.Shackelford R. E., Mayhall K., Maxwell N. M., Kandil E., Coppola D. Nicotinamide phosphoribosyltransferase in malignancy: a review. Genes & Cancer. 2013;4(11–12):447–456. doi: 10.1177/1947601913507576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Sanokawa-Akakura R., Ostrakhovitch E. A., Akakura S., Goodwin S., Tabibzadeh S. A H2S-Nampt dependent energetic circuit is critical to survival and cytoprotection from damage in cancer cells. PLoS One. 2014;9(9, article e108537) doi: 10.1371/journal.pone.0108537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Capitanio U., Bensalah K., Bex A., et al. Epidemiology of renal cell carcinoma. European Urology. 2019;75(1):74–84. doi: 10.1016/j.eururo.2018.08.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kovacs G., Akhtar M., Beckwith B. J., et al. The Heidelberg classification of renal cell tumours. The Journal of Pathology. 1997;183(2):131–133. doi: 10.1002/(SICI)1096-9896(199710)183:2<131::AID-PATH931>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 118.van Houwelingen K. P., van Dijk B. A. C., Hulsbergen-van de Kaa C. A., et al. Prevalence of von Hippel-Lindau gene mutations in sporadic renal cell carcinoma: results from the Netherlands cohort study. BMC Cancer. 2005;5(1) doi: 10.1186/1471-2407-5-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rini B. I., Campbell S. C., Escudier B. Renal cell carcinoma. The Lancet. 2009;373(9669):1119–1132. doi: 10.1016/S0140-6736(09)60229-4. [DOI] [PubMed] [Google Scholar]
- 120.Motzer R. J., Russo P. Systemic therapy for renal cell carcinoma. The Journal of Urology. 2000;163(2):408–417. doi: 10.1016/S0022-5347(05)67889-5. [DOI] [PubMed] [Google Scholar]
- 121.Breza J., Soltysova A., Hudecova S., et al. Endogenous H2S producing enzymes are involved in apoptosis induction in clear cell renal cell carcinoma. BMC Cancer. 2018;18 doi: 10.1186/s12885-018-4508-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Audenet F., Yates D. R., Cancel-Tassin G., Cussenot O., Rouprêt M. Genetic pathways involved in carcinogenesis of clear cell renal cell carcinoma: genomics towards personalized medicine. BJU International. 2012;109(12):1864–1870. doi: 10.1111/j.1464-410X.2011.10661.x. [DOI] [PubMed] [Google Scholar]
- 123.Maxwell P. H., Wiesener M. S., Chang G.-W., et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399(6733):271–275. doi: 10.1038/20459. [DOI] [PubMed] [Google Scholar]
- 124.Schulze A., Harris A. L. How cancer metabolism is tuned for proliferation and vulnerable to disruption. Nature. 2012;491(7424):364–373. doi: 10.1038/nature11706. [DOI] [PubMed] [Google Scholar]
- 125.Chen X., Qian Y., Wu S. The Warburg effect: evolving interpretations of an established concept. Free Radical Biology & Medicine. 2015;79:253–263. doi: 10.1016/j.freeradbiomed.2014.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.De Cicco P., Panza E., Ercolano G., et al. ATB-346, a novel hydrogen sulfide-releasing anti-inflammatory drug, induces apoptosis of human melanoma cells and inhibits melanoma development in vivo. Pharmacological Research. 2016;114:67–73. doi: 10.1016/j.phrs.2016.10.019. [DOI] [PubMed] [Google Scholar]
- 127.Lencesova L., Vlcek M., Krizanova O., Hudecova S. Hypoxic conditions increases H2S-induced ER stress in A2870 cells. Molecular and Cellular Biochemistry. 2016;414(1–2):67–76. doi: 10.1007/s11010-016-2659-4. [DOI] [PubMed] [Google Scholar]
- 128.Lu S., Chen L., Huang Q., et al. Decomposition of ammonia and hydrogen sulfide in simulated sludge drying waste gas by a novel non-thermal plasma. Chemosphere. 2014;117:781–785. doi: 10.1016/j.chemosphere.2014.10.036. [DOI] [PubMed] [Google Scholar]
- 129.Markova J., Hudecova S., Soltysova A., et al. Sodium/calcium exchanger is upregulated by sulfide signaling, forms complex with the β1 and β3 but not β2 adrenergic receptors, and induces apoptosis. Pflügers Archiv - European Journal of Physiology. 2014;466(7):1329–1342. doi: 10.1007/s00424-013-1366-1. [DOI] [PubMed] [Google Scholar]
- 130.Misak A., Grman M., Bacova Z., et al. Polysulfides and products of H2S/S-nitrosoglutathione in comparison to H2S, glutathione and antioxidant Trolox are potent scavengers of superoxide anion radical and produce hydroxyl radical by decomposition of H2O2. Nitric Oxide. 2018;76:136–151. doi: 10.1016/j.niox.2017.09.006. [DOI] [PubMed] [Google Scholar]
- 131.Cappello F., Conway de Macario E., Marasà L., Zummo G., Macario A. J. L. Hsp60 expression, new locations, functions, and perspectives for cancer diagnosis and therapy. Cancer Biology & Therapy. 2008;7(6):801–809. doi: 10.4161/cbt.7.6.6281. [DOI] [PubMed] [Google Scholar]
- 132.Ghosh J. C., Dohi T., Kang B. H., Altieri D. C. Hsp60 regulation of tumor cell apoptosis. The Journal of Biological Chemistry. 2008;283(8):5188–5194. doi: 10.1074/jbc.M705904200. [DOI] [PubMed] [Google Scholar]
- 133.Tsai Y.-P., Yang M.-H., Huang C.-H., et al. Interaction between HSP60 and β-catenin promotes metastasis. Carcinogenesis. 2009;30(6):1049–1057. doi: 10.1093/carcin/bgp087. [DOI] [PubMed] [Google Scholar]
- 134.Tang H., Chen Y., Liu X., et al. Downregulation of HSP60 disrupts mitochondrial proteostasis to promote tumorigenesis and progression in clear cell renal cell carcinoma. Oncotarget. 2016;7(25) doi: 10.18632/oncotarget.9615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Dong Q., Yang B., Han J.-G., et al. A novel hydrogen sulfide-releasing donor, HA-ADT, suppresses the growth of human breast cancer cells through inhibiting the PI3K/AKT/mTOR and Ras/Raf/MEK/ERK signaling pathways. Cancer Letters. 2019;455:60–72. doi: 10.1016/j.canlet.2019.04.031. [DOI] [PubMed] [Google Scholar]
- 136.Guo H., German P., Bai S., et al. The PI3K/AKT pathway and renal cell carcinoma. Journal of Genetics and Genomics. 2015;42(7):343–353. doi: 10.1016/j.jgg.2015.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Valenca L. B., Sweeney C. J., Pomerantz M. M. Sequencing current therapies in the treatment of metastatic prostate cancer. Cancer Treatment Reviews. 2015;41(4):332–340. doi: 10.1016/j.ctrv.2015.02.010. [DOI] [PubMed] [Google Scholar]
- 138.Liu M., Wu L., Montaut S., Yang G. Hydrogen sulfide signaling axis as a target for prostate cancer therapeutics. Prostate Cancer. 2016;2016:9. doi: 10.1155/2016/8108549.8108549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Guo H., Gai J.-W., Wang Y., Jin H.-F., Du J.-B., Jin J. Characterization of hydrogen sulfide and its synthases, cystathionine β-synthase and cystathionine γ-lyase, in human prostatic tissue and cells. Urology. 2012;79(2):483.e1–483.e5. doi: 10.1016/j.urology.2011.10.013. [DOI] [PubMed] [Google Scholar]
- 140.Zhao K., Li S., Wu L., Lai C., Yang G. Hydrogen sulfide represses androgen receptor transactivation by targeting at the second zinc finger module. The Journal of Biological Chemistry. 2014;289(30):20824–20835. doi: 10.1074/jbc.M114.559518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bigagli E., Luceri C., de Angioletti M., et al. New NO- and H2S-releasing doxorubicins as targeted therapy against chemoresistance in castration-resistant prostate cancer: in vitro and in vivo evaluations. Investigational New Drugs. 2018;36(6):985–998. doi: 10.1007/s10637-018-0590-0. [DOI] [PubMed] [Google Scholar]
- 142.Pei Y., Wu B., Cao Q., Wu L., Yang G. Hydrogen sulfide mediates the anti-survival effect of sulforaphane on human prostate cancer cells. Toxicology and Applied Pharmacology. 2011;257(3):420–428. doi: 10.1016/j.taap.2011.09.026. [DOI] [PubMed] [Google Scholar]
- 143.Chwatko G., Forma E., Wilkosz J., et al. Thiosulfate in urine as a facilitator in the diagnosis of prostate cancer for patients with prostate-specific antigen less or equal 10 ng/mL. Clinical Chemistry and Laboratory Medicine. 2013;51(9):1825–1831. doi: 10.1515/cclm-2013-0069. [DOI] [PubMed] [Google Scholar]
- 144.Al-Awadi F., Yang M., Tan Y., Han Q., Li S., Hoffman R. M. Human tumor growth in nude mice is associated with decreased plasma cysteine and homocysteine. Anticancer Research. 2008;28(5A):2541–2544. [PubMed] [Google Scholar]
- 145.Zhang W., Braun A., Bauman Z., Olteanu H., Madzelan P., Banerjee R. Expression profiling of homocysteine junction enzymes in the NCI60 panel of human cancer cell lines. Cancer Research. 2005;65(4):1554–1560. doi: 10.1158/0008-5472.CAN-04-1554. [DOI] [PubMed] [Google Scholar]
- 146.Kimura F., Franke K. H., Steinhoff C., et al. Methyl group metabolism gene polymorphisms and susceptibility to prostatic carcinoma. The Prostate. 2000;45(3):225–231. doi: 10.1002/1097-0045(20001101)45:3<225::AID-PROS4>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 147.Stabler S., Koyama T., Zhao Z., et al. Serum methionine metabolites are risk factors for metastatic prostate cancer progression. PLoS One. 2011;6(8, article e22486) doi: 10.1371/journal.pone.0022486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Stephan C., Jung K., Miller K., Ralla B. New biomarkers in serum and urine for detection of prostate cancer. Aktuelle Urologie. 2015;46(2):129–143. doi: 10.1055/s-0034-1398544. [DOI] [PubMed] [Google Scholar]
- 149.Deeble P. D., Cox M. E., Frierson H. F., Jr., et al. Androgen-independent growth and tumorigenesis of prostate cancer cells are enhanced by the presence of PKA-differentiated neuroendocrine cells. Cancer Research. 2007;67(8):3663–3672. doi: 10.1158/0008-5472.CAN-06-2616. [DOI] [PubMed] [Google Scholar]
- 150.Komiya A., Suzuki H., Imamoto T., et al. Neuroendocrine differentiation in the progression of prostate cancer. International Journal of Urology. 2009;16(1):37–44. doi: 10.1111/j.1442-2042.2008.02175.x. [DOI] [PubMed] [Google Scholar]
- 151.Heo J. H., Seo H. N., Choe Y. J., et al. T-type Ca2+ channel blockers suppress the growth of human cancer cells. Bioorganic & Medicinal Chemistry Letters. 2008;18(14):3899–3901. doi: 10.1016/j.bmcl.2008.06.034. [DOI] [PubMed] [Google Scholar]