

Abstract
Oocyte meiotic maturation is a vital and final process in oogenesis. Unlike somatic cells, the oocyte needs to undergo two continuous meiotic divisions (meiosis I and meiosis II) to become a haploid gamete. Notably, oocyte meiotic progression includes two rounds of unique meiotic arrest and resumption. The first arrest occurs at the G2 (germinal vesicle) stage and meiosis resumption is stimulated by a gonadotropin surge; the second arrest takes place at the metaphase II stage, the stage from which it is released when fertilization takes place. The maturation-promoting factor, which consists of cyclin B1 (CCNB1) and cyclin-dependent kinase 1 (CDK1), is responsible for regulating meiotic resumption and progression, while CDK1 is the unique CDK that acts as the catalytic subunit of maturation-promoting factor. Recent studies showed that except for cyclin B1, multiple cyclins interact with CDK1 to form complexes, which are involved in the regulation of meiotic progression at different stages. Here, we review and discuss the control of oocyte meiotic progression by cyclins A1, A2, B1, B2, B3, and O.
Keywords: cyclin, CDK1, meiosis, oocyte
Introduction
Cyclin/cyclin-dependent kinase (CDK) combinations drive and regulate cell cycle events at the G1, G1/S, S, and G2/M phases. A variety of cyclins (A1, A2, B1, B2, B3, C, D1, D2, D3, E1, E2, F, O, and others) and CDKs (CDK1-8 and others) have been identified in eukaryotic cells. CDK activity requires the combination with specific cyclins, and one CDK can bind with several cyclins. It has been suggested that CDK1 is essential for mitotic cell cycle progression in mammals given that deletion of only CDK1 leads to early embryonic death in mice [1, 2].
In oocyte meiosis, CDK1 is solely responsible for maturation-promoting factor (MPF) activation to promote meiotic progression because conditional knockout of CDK1 using growth differentiation factor 9 (Gdf-9) promoter-mediated Cre recombinase results in female infertility due to failure of oocyte meiotic resumption [3]. Cyclin A (A1, A2), cyclin B (B1-3), and cyclin O are shown to function in meiosis regulation (Table 1). Cyclin B1 has long been believed to be the major partner of CDK1; in addition, cyclin B2 (CCNB2) and cyclins A1 (CCNA1) and A2 (CCNA2) can also combine with CDK1 [4]. Both Ccna1 and Ccnb2 knockout mice are viable and develop normally [5, 6]. In contrast, deletion of either Ccnb1 or Ccna2 results in embryonic lethality [6, 7], making it impossible to study their functions in meiosis. Recently, conditional knockout (cKO) mice for Ccnb1 and Ccna2 were generated [8–10] and both cKO female mice were infertile. Surprisingly, cyclin B1-null oocytes resumed and finished meiosis I (MI) but were then arrested at meiosis interphase. Similarly, cyclin A2-null oocytes also showed no apparent delay on meiosis I progression but had disrupted the meiosis II (MII) spindle. Cyclin B3 is a newly discovered cyclin that potentially forms active kinase complexes with CDK1; its knockdown and knockout arrested the oocytes at metaphase I [11–13]. Cyclin O (CCNO, also called UNG2 or UDG2) is a novel cyclin family protein expressed in oocytes whose knockdown inhibited CDK1 (Tyr15) dephosphorylation and arrested oocytes at the germinal vesicle (GV) stage [14]; however, Ccno-deficient male and female mice are partly fertile [15]. In this review, we will summarize and discuss the functions of various cyclins mentioned above in line with time course of meiotic maturation.
Table 1.
Consequences of deletions of cyclins in mouse oocytes.
| Cyclins deleted | Female meiotic phenotype | References |
|---|---|---|
| Cyclin A1 | Fertile without abnormalities | [5] |
| Cyclin A2 | Infertile, abnormal meiosis II spindle formation, and kinetochore–microtubule attachment | [10] |
| Cyclin B1 | Infertile, meiosis II arrest failure and interphase entry | [9] |
| Cyclin B2 | Fertile, but with severe delay of meiotic resumption and progression | [6, 9] |
| Cyclin B3 | Infertile, failure of anaphase I onset | [9, 12] |
| Cyclin O | Female mice that were not severely affected by hydrocephalus were fertile at maturity age | [15] |
Ethic statement
This study was approved by the Institutional Animal Care Committee of the Institute of Zoology at the University of Chinese Academy of Sciences.
Cyclins in regulation of meiotic resumption
Cyclin B1 and cyclin B2
Meiosis resumption, marked by germinal vesicle breakdown (GVBD), is the most critical step during oocyte maturation, which is dependent on the activity of MPF. MPF is a heterodimer comprised of the regulatory subunit cyclin B and the catalytic subunit CDK1. For a long time, it has been believed that cyclin B1 synthesis/accumulation and its interaction with CDK1 is a prerequisite for MPF activation in oocytes. This view was mostly based on the embryonic lethal phenotype of Ccnb1 knockout in mice [6], because no homozygous Ccnb1−/− embryos were recovered at embryonic day 10. A recent study showed that Ccnb1−/− embryos were arrested at the G2 phase after just two rounds of cell divisions, indicating that cyclin B1 is essential for mitosis in mouse embryos [16]. However, the essentiality of cyclin B1 in meiotic resumption remains unclear. Our latest study on mouse oocytes using conditional knockout technology demonstrated that Ccnb1−/− oocytes readily underwent GVBD (equivalent to G2/M) because of the presence of cyclin B2 [9] (Figure 1), indicating that MPF was activated by cyclin B2 in the absence of cyclin B1. Notably, the translocation of cyclin B1 from the cytoplasm to the nucleus is an important step to trigger GVBD, and we also observed the nuclear translocation of cyclin B2 before GVBD. It reveals a compensatory mechanism between cyclin B2 and cyclin B1 in MPF activation, updating our understanding of MPF. In contrast, Ccnb2−/− oocytes showed a significant decrease in undergoing GVBD [9], which is in accordance with the in vitro experimental results [17]. It raises an interesting question on why cyclin B1 cannot compensate for the loss of cyclin B2 during meiotic resumption. We took it for granted before that cyclin B1 covered all functions of cyclin B2 in reproduction given the knockout phenotype; however, cyclin B2 may play a special role in meiosis. It is possible that cyclin B2 combines with different substrates from cyclin B1, facilitating MPF activation. In vitro studies showed that overexpression of cyclin B2 induced GVBD more efficiently than cyclin B1 in the presence of dbcAMP [18], also illustrating the significance of cyclin B2 in meiotic resumption. Taken together, it will be fascinating to reveal the regulatory mechanism of meiotic resumption by cyclin B2.
Figure 1.
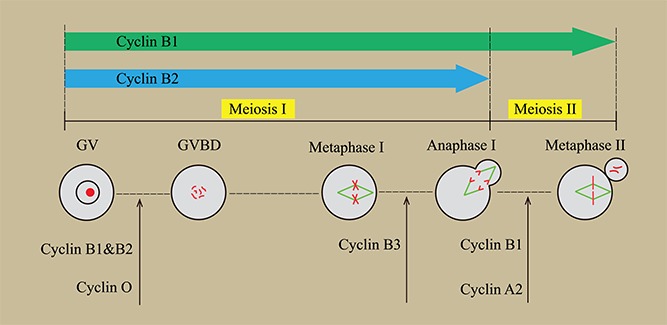
Diagram of cyclin function in oocyte meiotic maturation.
Cyclin A1 and cyclin A2
Both cyclin A1 and cyclin A2 can bind with CDK1 [4], but they seem to be dispensable for oocyte meiotic resumption because deletion of each has no effect on meiosis I progression [5, 10]. Taken the compensatory effect between the cyclin family members, the consequence is unknown when deleting cyclin A1 and cyclin A2 together in oocytes. In mitosis, cyclin A/CDK1 assists the activation of MPF by inhibiting Weel kinase, which depends on cyclin B; therefore, cyclin B may cover the loss of cyclin A in MPF activation. However, in oocytes, we found that overexpression of cyclin A can induce GVBD in the absence of cyclin B1 and cyclin B2 (unpublished data), indicating that cyclin A is able to activate MPF directly independent of cyclin B in meiosis. Hence, the physiological functions of cyclin A1 and cyclin A2 in MPF activation in oocytes remain to be further studied.
Cyclin O
Cyclin O is a cyclin-like DNA glycosylase that removes the cytosine deamination or misincorporated uracil on DNA. Cyclin O is highly expressed in oocytes and localizes in the nucleus of GV-stage oocytes. We found that cyclin O was essential for oocyte meiotic resumption because Ccno knockdown inhibited CDK1 (Tyr15) dephosphorylation, which was required for MPF activation [14]. Ccno knockout mice have been generated, and 57% mutant mice developed a prominent hydrocephalus, leading to abnormal head morphology and growth retardation; remaining male and female mice that were not severely affected by hydrocephalus were fertile at maturity age [15]. Ccno knockdown also destroyed the formation of microtubule-organizing centers at the perinuclear region in oocytes before meiotic resumption, similar to the defect of centriole amplification in Ccno-deficient mouse multiciliated cells. To confirm the function of cyclin O in MPF regulation, its overexpression in GV-stage oocytes still needs to be performed.
Cyclins regulating metaphase I-anaphase I transition
Cyclin B1 and cyclin B2
After the oocyte enters meiosis I, cyclin B1 accumulation and consequent MPF activation promote M-phase progression [19]. Cyclin B1 continues to accumulate and reaches its peak at metaphase I, and then cyclin B1 is degraded dramatically by a highly conserved anaphase-promoting complex (APC/C)-dependent process. Cyclin B1 destruction and the resulting MPF inactivation is the premise of metaphase I-anaphase I transition because high MPF activity prevents chromosome segregation by inhibiting separase activity. Overexpression or stable expression of Ccnb1 arrests the oocytes at metaphase I [18, 20]. A recent study showed that non-CDK1-bound cyclin B1 is gradually destroyed before CDK1-bound cyclin B1; the excess of cyclin B1 is important to preserve CDK1 activity and the prolonged CDK1 activity assists the spindle checkpoint and prevents aneuploidy [21]. Similarly, cyclin B2 keeps accumulating until metaphase I and then declines precipitously [17], indicating that cyclin B2 is also involved in MPF activation and inactivation during metaphase I-anaphase I transition. Moreover, cyclin B2 perfectly regulates MPF activity for meiosis I progression in the cyclin B1-null oocytes [9]. Different from prior work, we found that a portion of Ccnb2-overexpressing oocytes remained arrested in metaphase I [9]; besides, nondegradable Ccnb2-expressing oocytes were unable to progress beyond metaphase I (unpublished data). Therefore, it is very likely that cyclin B1 and cyclin B2 cooperate in the control of MPF activity in oocytes.
Cyclin B3
Cyclin B3 is a relatively new member of the cyclin family, distinct from other cyclins but it contains structural motifs characteristic of both A- and B-type cyclins [22, 23]. We found that cyclin B3 controls anaphase onset in meiotic oocytes, which is independent of the spindle assembly checkpoint [11]. Consistently, two mutant mice were generated recently, and the female mice were sterile due to the failure of metaphase I-anaphase I transition [12, 13]. Cyclin B3 may disturb the APC/C-dependent degradation of cyclin B1 and securin, resulting in high MPF activity and inactivation of separase activity. It was also found that cyclin B3 is a potential cyclin to form complexes with CDK1, which is different from the previous view that cyclin B3 combines with CDK2 [24]. In any case, it remains to be determined how cyclin B3/CDK adjusts APC/C activity through targeting its substrates. Additionally, it will be interesting to investigate the physiological role of cyclin B3/CDK1 in meiotic resumption.
Cyclins in MI-MII transition
Cyclin B1
After the first polar body extrusion (PBE), oocytes progress into metaphase II soon without a defined interphase. MPF reactivation by cyclin B1 reaccumulation is required for the MI-MII transition because cyclin B1-null oocytes enter interphase arrest marked by nucleus reformation after PBE [9], accompanied by spindle disassembly and chromatin decondensation. Actually, cyclin B1 is not destroyed completely during the MI-MII transition [25], suggesting that residual cyclin B1 is important to facilitate and guarantee the rapid reactivation of MPF. Nevertheless, the interphase arrest of cyclin B1-null oocytes can be rescued by exogenous cyclin B2 supplement, implying that cyclin B2 may be too low to reactivate MPF and the APC/C activity is relatively higher in the absence of cyclin B1.
Cyclin A2
Currently, Ccna2 conditional knockout mice have also been generated [10]. It was shown that cyclin A2 was necessary for normal MII spindle formation even though it was not required for MI progression. In the cyclin A2-null oocytes, achievement of the bipolar MII spindle was delayed markedly after formation of the first polar body; furthermore, after establishment of bipolarity, the chromosomes did not align on the spindle midzone properly, but migrated from the midzone and were distributed over the developing spindle. Taken together, cyclin A2 appears to modulate the kinetochore–microtubule attachment in meiosis II, but how it mediates and assures the rapid formation of the MII spindle remains unknown.
Conclusion
Undoubtedly, cyclins play critical roles during oocyte meiotic maturation. The new findings on cyclins reported in recent years enriched the regulatory network and most of them are relevant to MPF, especially the cyclin B1’s functional compensation by cyclin B2 in MPF activation for meiotic resumption. Apart from cyclin O, the other cyclins discussed here can bind with CDK1 directly and different cyclin/CDK1 complexes probably have their specific substrates. Interestingly, some cyclins seem to function in a stage-specific way, such as cyclin B3 that acts in meiosis I and cyclin A2 that acts in meiosis II (Figure 1). In the future, identification of the potential distinct substrates for different cyclins in meiosis is expected. Furthermore, the results we discussed here are all from knockout mice; whether these results apply to all mammals and whether there are species differences in cyclin or CDK function remain unknown.
Conflict of interest
The authors have declared that no conflict of interest exists.
Acknowledgment
The authors would like to express thanks to Prof. Heide Schatten at the University of Missouri-Columbia for her critical reading of the manuscript.
References
- 1. Santamaria D, Barriere C, Cerqueira A, Hunt S, Tardy C, Newton K, Caceres JF, Dubus P, Malumbres M, Barbacid M. Cdk1 is sufficient to drive the mammalian cell cycle. Nature 2007; 448:811–U818. [DOI] [PubMed] [Google Scholar]
- 2. Diril MK, Ratnacaram CK, Padmakumar VC, Du TH, Wasser M, Coppola V, Tessarollo L, Kaldis P. Cyclin-dependent kinase 1 (Cdk1) is essential for cell division and suppression of DNA re-replication but not for liver regeneration. Proc Natl Acad Sci USA 2012; 109:3826–3831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Adhikari D, Zheng W, Shen Y, Gorre N, Ning Y, Halet G, Kaldis P, Liu K. Cdk1, but not Cdk2, is the sole Cdk that is essential and sufficient to drive resumption of meiosis in mouse oocytes. Hum Mol Genet 2012; 21:2476–2484. [DOI] [PubMed] [Google Scholar]
- 4. Satyanarayana A, Kaldis P. Mammalian cell-cycle regulation: several Cdks, numerous cyclins and diverse compensatory mechanisms. Oncogene 2009; 28:2925–2939. [DOI] [PubMed] [Google Scholar]
- 5. Liu D, Matzuk MM, Sung WK, Guo QX, Wang P, Wolgemuth DJ. Cyclin A1 is required for meiosis in the male mouse. Nat Genet 1998; 20:377–380. [DOI] [PubMed] [Google Scholar]
- 6. Brandeis M, Rosewell I, Carrington M, Crompton T, Jacobs MA, Kirk J, Gannon J, Hunt T. Cyclin B2-null mice develop normally and are fertile whereas cyclin B1-null mice die in utero. Proc Natl Acad Sci USA 1998; 95:4344–4349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Murphy M, Stinnakre MG, SenamaudBeaufort C, Winston NJ, Sweeney C, Kubelka M, Carrington M, Brechot C, SobczakThepot J. Delayed early embryonic lethality following disruption of the murine cyclin A2 gene. Nat Genet 1997; 15:83–86. [DOI] [PubMed] [Google Scholar]
- 8. Tang JX, Li J, Cheng JM, Hu B, Sun TC, Li XY, Batool A, Wang ZP, Wang XX, Deng SL, Zhang Y, Chen SR et al. . Requirement for CCNB1 in mouse spermatogenesis. Cell Death Dis 2017; 8:e3142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Li J, Tang JX, Cheng JM, Hu B, Wang YQ, Aalia B, Li XY, Jin C, Wang XX, Deng SL, Zhang Y, Chen SR et al. . Cyclin B2 can compensate for Cyclin B1 in oocyte meiosis I. J Cell Biol 2018; 217:3901–3911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Zhang QH, Yuen WS, Adhikari D, Flegg JA, FitzHarris G, Conti M, Sicinski P, Nabti I, Marangos P, Carroll J. Cyclin A2 modulates kinetochore-microtubule attachment in meiosis II. J Cell Biol 2017; 216:3133–3143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Zhang T, Qi ST, Huang L, Ma XS, Ouyang YC, Hou Y, Shen W, Schatten H, Sun QY. Cyclin B3 controls anaphase onset independent of spindle assembly checkpoint in meiotic oocytes. Cell Cycle 2015; 14:2648–2654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Karasu ME, Bouftas N, Keeney S, Wassmann K. Cyclin B3 promotes anaphase I onset in oocyte meiosis. J Cell Biol 2019; 218:1265–1281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Li Y, Wang L, Zhang L, He Z, Feng G, Sun H, Wang J, Li Z, Liu C, Han J, Mao J, Li P et al. . Cyclin B3 is required for metaphase to anaphase transition in oocyte meiosis I. J Cell Biol 2019; 218:1553–1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ma JY, Ou-Yang YC, Luo YB, Wang ZB, Hou Y, Han ZM, Liu ZH, Schatten H, Sun QY, Cyclin O. Regulates germinal vesicle breakdown in mouse oocytes. Biol Reprod 2013; 88:110. [DOI] [PubMed] [Google Scholar]
- 15. Funk MC, Bera AN, Menchen T, Kuales G, Thriene K, Lienkamp SS, Dengjel J, Omran H, Frank M, Arnold SJ. Cyclin O (Ccno) functions during deuterosome-mediated centriole amplification of multiciliated cells. EMBO J 2015; 34:1078–1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Strauss B, Harrison A, Coelho PA, Yata K, Zernicka-Goetz M, Pines J. Cyclin B1 is essential for mitosis in mouse embryos, and its nuclear export sets the time for mitosis. J Cell Biol 2018; 217:179–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Gui L, Homer H. Hec1-dependent cyclin B2 stabilization regulates the G2-M transition and early prometaphase in mouse oocytes. Dev Cell 2013; 25:43–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ledan E, Polanski Z, Terret ME, Maro B. Meiotic maturation of the mouse oocyte requires an equilibrium between cyclin B synthesis and degradation. Dev Biol 2001; 232:400–413. [DOI] [PubMed] [Google Scholar]
- 19. Polanski Z, Homer H, Kubiak JZ. Cyclin B in mouse oocytes and embryos: importance for human reproduction and aneuploidy. Results Probl Cell Differ 2012; 55:69–91. [DOI] [PubMed] [Google Scholar]
- 20. Herbert M, Levasseur M, Homer H, Yallop K, Murdoch A, McDougall A. Homologue disjunction in mouse oocytes requires proteolysis of securin and cyclin B1. Nat Cell Biol 2003; 5:1023–1025. [DOI] [PubMed] [Google Scholar]
- 21. Levasseur MD, Thomas C, Davies OR, Higgins JMG, Madgwick S. Aneuploidy in oocytes is prevented by sustained CDK1 activity through degron masking in cyclin B1. Dev Cell 2019; 48:672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Nieduszynski CA, Murray J, Carrington M. Whole-genome analysis of animal A- and B-type cyclins. Genome Biol 2002; 3:research0070.1–research0070.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Gunbin KV, Suslov VV, Turnaev II, Afonnikov DA, Kolchanov NA. Molecular evolution of cyclin proteins in animals and fungi. BMC Evo Biol 2011; 11:224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Nguyen TB, Manova K, Capodieci P, Lindon C, Bottega S, Wang XY, Refik-Rogers J, Pines J, Wolgemuth DJ, Koff A. Characterization and expression of mammalian cyclin b3, a prepachytene meiotic cyclin. J Biol Chem 2002; 277:41960–41969. [DOI] [PubMed] [Google Scholar]
- 25. Huo LJ, Yu LZ, Liang CG, Fan HY, Chen DY, Sun QY. Cell-cycle-dependent subcellular localization of cyclin B1, phosphorylated cyclin B1 and p34cdc2 during oocyte meiotic maturation and fertilization in mouse. Zygote 2005; 13:45–53. [DOI] [PubMed] [Google Scholar]