

Abstract
Deep vein thrombosis (DVT) is one of the most common cardiovascular diseases. The apoptosis of vascular endothelial cells is the most important cause of venous thrombosis. MicroRNAs (miRNAs) play important roles in the regulation of cell apoptosis. miRNA (miR)-195 is upregulated in the blood of patients with DVT, and it was predicted that Bcl-2 is a potential target of miR-195-5p. Therefore, it was hypothesized that miR-195-5p may play an important role in the development of DVT by targeting Bcl-2. The present study aimed to investigate the expression of miR-195-5p in DVT patients, and to explore whether miR-195-5p is involved in the development of DVT by regulating the apoptosis of vascular endothelial cells. The level of miR-195-5p was detected using reverse transcription-quantitative PCR. Dual luciferase reporter assays were used to determine the relationship between Bcl-2 and miR-195-5p. Cell viability was detected using MTT assays, and cell apoptosis was analyzed by flow cytometry. Protein levels of Bcl-2 and Bax were measured by western blotting. The results indicated that miR-195-5p was significantly upregulated in the blood of DVT patients. It was also revealed that Bcl-2 was a direct target of miR-195-5p, and that Bcl-2 was downregulated in the blood of patients with DVT. miR-195-5p downregulation promoted cell viability and inhibited the apoptosis of human umbilical vein endothelial cells (HUVECs). miR-195-5p upregulation inhibited cell viability and increased the apoptosis of HUVECs. All of the observed effects of miR-195-5p upregulation on HUVECs were reversed by raised Bcl-2 expression. In conclusion, miR-195-5p was significantly upregulated in patients with DVT, and it may be involved in the development of DVT by regulating the apoptosis of vascular endothelial cells. Therefore, miR-195-5p may be a potential target for predicting and treating DVT.
Keywords: deep venous thrombosis, vascular endothelial cells, apoptosis, microRNA-195-5p, Bcl-2
Introduction
Venous thromboembolism (VTE) is a clinically common vascular disease, including deep vein thrombosis (DVT) and pulmonary embolism (1,2). DVT refers to thrombosis in deep veins, including, but not limited to, femoral veins, iliac veins and intramuscular veins. According to the site of occurrence, DVT can be divided into upper limb DVT and lower limb DVT, with lower limb DVT being more common (3). The typical clinical features of patients with DVT are lower limb muscle soreness, swelling and tenderness, but there is a lack of specific symptoms (4). On average, ~50% of DVT patients have no typical clinical manifestation, which may lead to missed diagnosis and misdiagnosis, and therefore increase the difficulty of treating DVT (2,5). Additionally, post-thrombotic syndrome (PTS) occurs in 20–50% of patients with DVT, even after the appropriate treatment (6). PTS is the most common long-term complication in patients with DVT. The development of PTS is also associated with an increased risk of the recurrence of VTE, severely affecting the quality of life of patients and increasing the economic burden of families and society (7–9). Therefore, timely and accurate diagnosis of DVT, as well as effective interventions, are of importance for alleviating the suffering of patients and even saving lives.
Currently, the mechanism of action behind the pathogenesis of DVT remains unclear, hindering its prevention and treatment (10). It is now widely considered that vascular endothelial cells, the coagulation/anticoagulation system, the fibrinolysis/antifibrin system, platelets, changes in blood rheology, inflammatory factors and other factors are all involved in the pathophysiology behind DVT (11). At present, vascular wall injury, changes in blood flow and abnormal blood components are considered to be the three most notable factors behind thrombosis (12). Among them, blood vessel wall injury mainly constitutes damage to vascular endothelial cells. Vascular endothelial cell damage includes apoptosis, which is the most important cause of venous thrombosis (13,14). Previous studies have confirmed that damage to vascular endothelial cells are closely associated with the development of DVT, and the apoptosis of vascular endothelial cells reduces the levels of active substances and impairs their multiple defense functions in blood vessels, as well as reducing the stability of the anticoagulation-fibrinolytic system, thus increasing the risk of thrombus formation (14–17).
MicroRNAs (miRNAs) are endogenous non-coding single-stranded small-molecule RNAs, found in eukaryotic cells, that are ~22 nucleotides in length (18,19). miRNAs regulate gene expression at the post-transcriptional level mainly by binding to the 3′-untranslated region (3′-UTR) of target mRNAs, inhibiting translation or degradation (18). miRNAs are widely involved in the regulation of physiological and pathological processes in various cells and tissues, and are involved with cell differentiation, proliferation, apoptosis, and the development of tissues and organs (20–22). In recent years, increasing evidence has indicated that miRNAs play an important role in the development of DVT (23–27). miRNA (miR)-195-5p has been studied in several cancer types including breast cancer (28), non-small cell lung cancer (29), cervical carcinoma (30) and human endometrial carcinoma (31). miR-195-5p has also been found to regulate hair follicle inductivity of dermal papilla cells by suppressing the activation of the Wnt/β-Catenin signaling pathway (32). Furthermore, it has been suggested that both peripheral blood and urinary miR-195-5p may be a potential biomarker for membranous nephropathy (33). All of these observations indicated that miR-195-5p is expressed differently under different pathophysiological conditions and that miR-195-5p plays a very important role in regulating cell growth. Additionally, miR-195-5p has been proven to promote pulmonary arterial smooth muscle cell proliferation and migration in pulmonary arterial hypertension (34). Additionally, higher expression of miR-195-5p inhibits angiogenesis in preeclampsia (35). A recent study reported that miR-195 is upregulated in the blood of DVT patients (36); however, to the best of our knowledge, the expression and role of miR-195-5p in DVT remain unclear. Bcl-2, the founding member of the Bcl-2 protein family, has anti-apoptotic activity, and the anti-apoptotic function of Bcl-2 is mediated by its effects on intracellular Ca2+ homeostasis and dynamics (37). It was found by bioinformatics analysis that miR-195-5p binds to the 3′-UTR of Bcl-2, suggesting a direct interaction between miR-195-5p and Bcl-2. Therefore, it was hypothesized that miR-195-5p may be involved in DVT development by regulating vascular endothelial cell apoptosis.
Therefore, the aim of the present study was to investigate the expression of miR-195-5p in DVT patients, and to explore whether miR-195-5p is involved in the development of DVT by regulating the apoptosis of vascular endothelial cells.
Materials and methods
Clinical samples
This present study was approved by the ethics review committee of the Gansu Provincial Hospital of TCM, and all patients provided their written informed consent. Peripheral blood (5 ml/subject) was collected from 15 patients with DVT and from 15 healthy volunteers. All DVT patients were confirmed by color Doppler ultrasonography (38). The Doppler ultrasonographic criteria for DVT were as follows: No flow signal; direct clot visualization; the absence of spontaneous flow; and the absence of respiration-modulated phasicity of the evaluated veins. The criterion for diagnosis of DVT was a filling defect of the venous lumen in more than two observations, following the American College of Chest Physicians Evidence-Based Clinical Practice Guidelines of diagnosis for DVT (39). The exclusion criteria for the study were as follows: Ongoing anticoagulation treatment for >3 months; pregnancy; duration of symptoms for more than 1 year; and previous thrombosis within the last year.
Cell culture
HUVECs were obtained from the American Type Culture Collection. Cells were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.) and maintained at 37°C with 5% CO2.
Dual luciferase reporter assay
The binding sites between miR-195-5p and Bcl-2 were predicted using TargetScan bioinformatics software (version 7.1; www.targetscan.org/vert_71). To confirm predictions, dual luciferase reporter assays were performed. The wild-type (WT)-Bcl-2 and mutant (MUT)-Bcl-2 3′-UTRs of Bcl-2 were cloned into a pmiR-RB-Report™ dual luciferase reporter gene plasmid vector (Guangzhou RiboBio Co., Ltd.) according to the manufacturer's instructions. HUVECs were co-transfected with WT-Bcl-2 or MUT-Bcl-2 and miR-195-5p mimic or mimic control by using Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 48 h. A total of 48 h after cell transfection, the luciferase activity was analyzed using the dual-luciferase assay system (Promega Corporation). Luciferase activity was normalized to the Renilla luciferase activity in the current study.
Cell transfection
Inhibitor control (chemically modified RNA single strand), miR-195-5p inhibitor (chemically modified RNA single strand), mimic control and miR-195-5p mimic were purchased from Guangzhou RiboBio Co., Ltd. HUVECs were plated into 6-well plates at a density of 1×106 cells/well and cultured at 37°C with 5% CO2 for 24 h. Then, 100 nM miR-195-5p inhibitor (5′-GCCAAUAUUUCUGUGCUGCUA-3′), 100 nM inhibitor control (5′-CAGUACUUUUGUGUAGUACAA-3′), 50 nM miR-195-5p mimic (5′-UAGCAGCACAGAAAUAUUGGC-3′), 50 nM mimic control (5′-UUCUCCGAACGUGUCACGUTT-3′), 1 µg Bcl-2 CRISPR Activation Plasmid (Bcl-2 plasmid; cat no. sc400025-ACT; Santa Cruz Biotechnology, Inc.), 1 µg control CRISPR Activation Plasmid (control plasmid; cat no. sc-437275; Santa Cruz Biotechnology, Inc.), or 50 nM miR-195-5p mimic + 1 µg Bcl-2 plasmid was transfected into HUVECs by using Lipofectamine 2000, according to the manufacturer's protocols. A total of 48 h after cell transfection, reverse transcription-quantitative PCR (RT-qPCR) was performed to assess the transfection efficiency.
MTT assay
MTT assays were performed to detect the cell viability. Briefly, 48 h after cell transfection, HUVECs were seeded into a 96-well plate (2×104 cells/well). Then, 10 µl MTT reagent (Beyotime Institute of Biotechnology) was added to each well and the cells were further incubated for 4 h at 37°C, following which 150 µl DMSO was used to dissolve the purple formazan crystals. Finally, absorbance at a wavelength of 490 nm was measured using an automatic enzyme-linked immune detector. Cell viability was calculated using the following formula: Cell viability=optical density (OD) of treated cells/OD (control) ×100%. Tests were repeated three times.
Cell apoptosis assay
After transfection for 48 h, the apoptosis of HUVECs was analyzed by using the Annexin V-FITC/propidium iodide apoptosis detection kit [cat no. 70-AP101-100; Hangzhou Multi Sciences (Lianke) Biotech Co., Ltd.] in line with the manufacturer's instructions. BD FACSCalibur™ flow cytometer with Cell Quest software (version 5.1; BD Biosciences), was used to detect the cell apoptosis rate. Each experiment was repeated three times.
RT-qPCR
To extract the total RNA from blood samples and cells, TRIzol® reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used according to the manufacturer's instructions. Total RNA was reverse transcribed into cDNAs using the miScript Reverse Transcription kit (Qiagen GmbH). The temperature protocol for the reverse transcription reaction was as follows: 25°C for 5 min, 42°C for 60 min and 80°C for 2 min. For qPCR analysis, the QuantiFast SYBR Green PCR kit (Qiagen GmbH) was used. Amplification conditions were as follows: 95°C for 10 min; and 35 cycles of 95°C for 15 sec and 55°C for 40 sec. GAPDH was used as the internal control for Bcl-2 and Bax mRNA expression, and U6 was used as the internal control for miR-195-5p expression. Primer sequences for PCR were as follows: GAPDH, forward 5-′CTTTGGTATCGTGGAAGGACTC-3′, reverse 5-′GTAGAGGCAGGGATGATGTTCT-3′; U6, forward 5-′GCTTCGGCAGCACATATACTAAAAT-3′, reverse 5-′CGCTTCACGAATTTGCGTGTCAT-3′; miR-195-5p forward 5-′GGGGTAGCAGCACAGAAAT-3′, reverse 5-′TCCAGTGCGTGTCGTGGA-3′; Bcl2 forward 5′-GCCCTGTGGATGACTGAGTA-3′, reverse, 5′-GGCCGTACAGTTCCACAAAG-3′; and Bax, forward 5′-TGGCAGCTGACATGTTTTCTGAC-3′, reverse 5′-TCACCCAACCACCCTGGTCTT-3′. Relative gene expression was calculated by using the 2−ΔΔCq method (40).
Western blotting
Proteins from cells or blood samples were extracted using the RIPA lysis buffer (Beyotime Institute of Biotechnology). Protein concentration was quantified using Bicinchoninic Acid Protein Assay kit. Proteins (30 µg/lane) were separated by using SDS-PAGE on a 12% gel, and then transferred onto the PVDF membranes. Membranes were then blocked with 5% skim milk at room temperature for 1.5 h and incubated with primary antibodies overnight at 4°C: Bcl-2 (cat no. 4223; 1:1,000; Cell Signaling Technology, Inc.), Bax (cat no. 5023; 1:1,000; Cell Signaling Technology, Inc.), and β-actin (cat no. 4970; 1:1,000; Cell Signaling Technology, Inc.). The membranes were finally incubated with the horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody (cat no. 7074; dilution ratio: 1:5,000; Cell Signaling Technology, Inc.) at room temperature for 2 h. To visualize the protein bands, the ECL detection system (Thermo Fisher Scientific, Inc.) was used. Protein bands were quantified using. β-actin was used as a reference protein for each experiment and each experiment was repeated three times. Densitometric analyzes were performed using ImageJ software (version 1.38X; National Institutes of Health).
Statistical analysis
Data were analyzed using SPSS 18.0 software (SPSS, Inc.), and are presented as the mean ± SD. Comparisons between groups were made by Students t-test or one-way ANOVA with Tukey's post hoc tests. P<0.05 was considered to indicate a statistically significant difference.
Results
miR-195-5p is upregulated in patients with DVT
Levels of miR-195-5p in the peripheral blood were first analyzed in samples from 15 DVT patients and in 15 volunteers, using RT-qPCR. Levels of miR-195-5p in the peripheral blood from DVT patients was significantly higher than those in the peripheral blood from the healthy volunteers (Fig. 1). These data indicated that miR-195-5p is involved in DVT progression.
Figure 1.
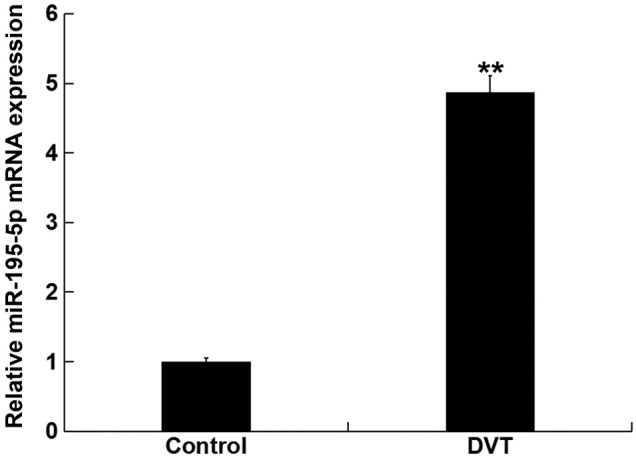
miR-195-5p is upregulated in DVT patients. Levels of miR-195-5p in the peripheral blood from 15 healthy volunteers (Control) and in the peripheral blood from 15 DVT patients were detected using reverse transcription-quantitative PCR. Data are presented as the mean ± SD. **P<0.01 vs. Control. DVT, deep vein thrombosis; miR, microRNA.
Bcl-2 is a direct target of miR-195-5p
Results from TargetScan bioinformatics software indicated that miR-195-5p has hundreds of potential target genes, including Bcl-2 (Fig. 2A). Bcl-2, the founding member of the Bcl-2 protein family, is a widely known anti-apoptotic gene (37). Apoptosis plays a crucial role in the pathophysiology of venous thrombosis (13,14), and Bcl-2 has been identified to participate in the development of thrombosis (41). Therefore, it was hypothesized that miR-195-5p may play an important role in the development of DVT by affecting the apoptosis of vascular endothelial cells by regulating the expression of Bcl-2. Thus, Bcl-2 was chosen for further study, and the dual luciferase reporter assay supported the prediction (Fig. 2B). The results showed that Bcl-2 was a direct target of miR-195-5p.
Figure 2.

Bcl-2 is a direct target gene of miR-195-5p. (A) TargetScan software was used to predict a binding site for miR-195-5p in the 3′UTR of Bcl-2. (B) Luciferase activity of a dual-luciferase reporter vector containing wild-type 3′UTR-Bcl-2 or a mutant 3′UTR-Bcl-2. Data are presented as the mean ± SD of three independent experiments. (C) The mRNA expression levels of Bcl-2 in the peripheral blood from 15 healthy volunteers (Control) and in the peripheral blood from 15 DVT patients using reverse transcription-quantitative PCR. (D) Representative western blotting of Bcl-2 in the peripheral blood from 2 healthy volunteers (Control) and in the peripheral blood from 2 DVT patients. (E) The ratio of Bcl-2/β-actin was calculated in the different groups. WT, wild-type; MUT, mutant-type; WT-Bcl-2, HUVECs co-transfected with WT 3′UTR-Bcl-2 and either mimic control or miR-195-5p; MUT-Bcl-2, HUVECs transfected with MUT 3′UTR-Bcl-2. Data are presented as the mean ± SD. **P<0.01 vs. mimic control; ##P<0.01 vs. control. DVT, deep vein thrombosis; HUVECs, human umbilical vein endothelial cells; UTR, untranslated region; miR, microRNA.
Furthermore, the mRNA levels of Bcl-2 in the peripheral blood from DVT patients was significantly lower than that in the peripheral blood from the healthy volunteers (Fig. 2C). Randomly selected blood samples from 2 DVT patients and 2 healthy volunteers were tested for Bcl-2 protein expression. The protein level of Bcl-2 in the peripheral blood from DVT patients was lower than that in the peripheral blood from the healthy volunteers (Fig. 2D and E). As Bcl-2 is a well-known anti-apoptotic gene, it was hypothesized that miR-195-5p may be involved in the development and progression of DVT by regulating the apoptosis of vascular endothelial cells.
miR-195-5p downregulation promotes cell viability and inhibits apoptosis in HUVECs
To investigate the effect of miR-195-5p downregulation on the proliferation and apoptosis of HUVECs, control inhibitor or miR-195-5p inhibitor was transfected into HUVECs for 48 h. Compared with the inhibitor control group, miR-195-5p inhibitor significantly inhibited miR-195-5p expression in HUVECs (Fig. 3A). miR-195-5p inhibitor significantly enhanced the cell viability and inhibited the apoptosis of HUVECs (Fig. 3B and C). Additonally, compared with the inhibitor control group, the mRNA level of Bcl-2 was significantly enhanced, while Bax mRNA expression was reduced (Fig. 3D and E). Compared with the inhibitor control group, Bcl-2 protein levels appeared raised, while Bax protein expression levels appeared reduced in HUVECs transfected with miR-195-5p inhibitor (Fig. 3F).
Figure 3.

Effect of miR-195-5p downregulation on HUVECs. (A) After transfection with inhibitor control or miR-195-5p inhibitor for 48 h, the level of miR-195-5p in HUVECs was detected using RT-qPCR. (B) After transfection with inhibitor control or miR-195-5p inhibitor for 48 h, the cell viability of HUVECs was detected using MTT assay. (C) After transfection with inhibitor control or miR-195-5p inhibitor for 48 h, cell apoptosis of HUVECs was detected using flow cytometry, and the cell apoptosis rate (Q2 + Q4) was calculated and presented. After transfection with inhibitor control or miR-195-5p inhibitor for 48 h, the mRNA level of (D) Bcl-2 and (E) Bax in HUVECs was detected using RT-qPCR. (F) After transfection with inhibitor control or miR-195-5p inhibitor for 48 h, the protein level of Bcl-2 and Bax in HUVECs was detected using western blotting. Control, HUVECs without any treatment; inhibitor control, HUVECs transfected with inhibitor control for 48 h; inhibitor, HUVECs were transfected with miR-195-5p inhibitor for 48 h. Data are presented as the mean ± SD. *P<0.05, **P<0.01 vs. inhibitor control. HUVECs, human umbilical vein endothelial cells; miR, microRNA; PI, propidium iodide; Q, quadrant; RT-qPCR, reverse transcription-quantitative PCR.
miR-195-5p upregulation inhibits cell viability and induces apoptosis in HUVECs
To investigate the effect of miR-195-5p upregulation on the proliferation and apoptosis of HUVECs, HUVECs were transfected with miR-195-5p mimic, mimic control, Bcl-2-plasmid, control-plasmid, or miR-195-5p mimic + Bcl-2-plasmid, for 48 h. Compared with the mimic control group, miR-195-5p mimic significantly raised miR-195-5p expression in HUVECs (Fig. 4A). Transfection with the Bcl-2-plasmid significantly increased the mRNA expression of Bcl-2 in HUVECs (Fig. 4B). Similarly, the Bcl-2-plasmid appeared to markedly raise the protein expression of Bcl-2 (Fig. 4C). miR-195-5p mimic significantly inhibited the cell viability and induced the apoptosis of HUVECs, although these effects were reversed by Bcl-2-plasmid (Fig. 4D and E). Additionally, compared with the mimic control group, miR-195-5p mimic appeared to greatly reduce the protein levels of Bcl-2, while also appearing to increase Bax protein expression levels (Fig. 4F). These changes appeared to reverse when HUVECs were also transfected with the Bcl-2-plasmid. Compared with the mimic control group, miR-195-5p mimic significantly reduced the mRNA levels of Bcl-2, while Bax mRNA expression increased (Fig. 4G and H), and these changes were reversed when the Bcl-2-plasmid was co-transfected with miR-195-5p mimic.
Figure 4.

Effect of miR-195-5p upregulation on HUVECs. (A) After transfection with mimic control or miR-195-5p mimic for 48 h, the level of miR-195-5p in HUVECs was detected using RT-qPCR. After transfection with control-plasmid or Bcl-2-plasmid for 48 h, the mRNA and protein level of Bcl-2 in HUVECs was detected using (B) RT-qPCR and (C) western blotting. (D) After transfection with mimic control, miR-195-5p mimic or miR-195-5p mimic + Bcl-2-plasmid for 48 h, the cell viability of HUVECs was detected using MTT assay. (E) After transfection with mimic control, miR-195-5p mimic or miR-195-5p mimic + Bcl-2-plasmid for 48 h, the cell apoptosis of HUVECs was detected using flow cytometry, and the cell apoptosis rate (Q2 + Q4) was calculated and presented. (F) After transfection with mimic control, miR-195-5p mimic or miR-195-5p mimic + Bcl-2-plasmid for 48 h, the protein level of Bcl-2 and Bax in HUVECs was detected using western blotting. After transfection with mimic control, miR-195-5p mimic or miR-195-5p mimic+Bcl-2-plasmid for 48 h, the mRNA level of (G) Bcl-2 and (H) Bax in HUVECs was detected using RT-qPCR. Control, HUVECs without any treatment; mimic control, HUVECs transfected with mimic control for 48 h; miR-195-5p mimic, HUVECs transfected with miR-195-5p mimic for 48 h; control-plasmid, HUVECs transfected with control-plasmid for 48 h; Bcl-2-plasmid, HUVECs transfected with Bcl-2-plasmid for 48 h; miR-195-5p mimic+Bcl-2-plasmid, HUVECs co-transfected with miR-195-5p mimic + Bcl-2-plasmid for 48 h. Data are presented as the mean ± SD. **P<0.01 vs. mimic control or control-plasmid; ##P<0.01 vs. miR-195-5p mimic. HUVECs, human umbilical vein endothelial cells; miR, microRNA; PI, propidium iodide; Q, quadrant; RT-qPCR, reverse transcription-quantitative PCR.
Discussion
DVT is a clinically common peripheral vascular lesion (1,2). Current clinical treatments for DVT include anti-coagulation, thrombolysis and surgical thrombectomy. However, all of the aforementioned methods have the disadvantages of PTS, low long-term patency rate, and easy recurrence. Therefore, developing more effective treatments for DVT is an important area of research for vascular surgeons. There is increasing evidence indicating that miRNAs, as micro-regulators, play an important role in regulating angiogenic signaling pathways (36,42).
miR-195-5p has been discovered to play important roles in various cancer types, including cervical carcinoma, human endometrial carcinoma, colorectal cancer, melanoma and osteosarcoma by regulating cell proliferation and apoptosis (30,31,43–45). A recent study also reported the inhibitory role of miR-195 in angiogenesis in human prostate cancer (46). Furthermore, Sandrim et al (35) revealed an anti-angiogenic role for miR-195-5p in endothelial cells. miR-195 is upregulated in the blood of DVT patients (36). However, the expression and role of miR-195-5p in DVT remain unclear. This present study investigated the expression of miR-195-5p in DVT patients, and explored whether miR-195-5p was involved in the pathophysiology of DVT by regulating the apoptosis of vascular endothelial cells.
Firstly, miR-195-5p levels were detected in the blood of DVT patients and healthy controls using RT-qPCR. The results indicated that compared with the healthy controls, miR-195-5p was significantly upregulated in the peripheral blood of DVT patients, indicating that miR-195-5p has a role in the development of DVT. Bcl-2, a well-known anti-apoptotic gene, was identified as a direct target of miR-195-5p, which also appeared to be downregulated in the peripheral blood of DVT patients. Thus, it was hypothesized that miR-195-5p may be involved in the development and progression of DVT by regulating apoptosis in vascular endothelial cells. It has to be noted, however, that a limitation of this present study is that only two DVT samples were analyzed.
Subsequently, the role of miR-195-5p in the regulation of cell viability and apoptosis of vascular endothelial cells was investigated. The findings suggested that miR-195-5p inhibition significantly promoted the cell viability and inhibited the apoptosis of HUVECs, while miR-195-5p upregulation significantly inhibited cell viability and increased the apoptosis of HUVECs. Additionally, miR-195-5p upregulation significantly inhibited Bcl-2 expression and induced Bax expression in HUVECs, while miR-195-5p inhibition demonstrated the opposite effects. It is worth mentioning that all the effects of miR-195-5p mimic on HUVECs were significantly reduced by Bcl-2 overexpression.
Taken together, the present study indicated that miR-195-5p was significantly upregulated in DVT patients, and that it may be involved in the development of DVT by regulating the apoptosis of vascular endothelial cells. It was further identified that Bcl-2 was a direct target of miR-195-5p, and that Bcl-2 was downregulated in the peripheral blood of DVT patients. Thus, it is thought that the downregulation of Bcl-2 in the peripheral blood of DVT patients was associated with the upregulation of miR-195-5p. Moreover, the data of this present study indicated that downregulation of miR-195-5p significantly promoted the cell viability and inhibited the apoptosis of HUVECs. Raised miR-195-5p expression inhibited cell viability and induced apoptosis in HUVECs, changes that were reversed by raised Bcl-2 expression. These data indicated that miR-195-5p was involved in the development of DVT by regulating the apoptosis of vascular endothelial cells. Therefore, miR-195-5p may be a potential DVT biomarker and therapeutic target. However, this study is only a preliminary study of miR-195-5p in DVT and as such, contains limitations. The sample size of this study was based on previous studies (21–23). A power analysis should be performed for future studies to determine the effective sample volume. Furthermore, a ‘mimic + control plasmid group’ was not set up in this study. Finally, in vivo experiments are also required to further this research.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
JLJ contributed to study design, data collection, statistical analysis, data interpretation and manuscript preparation. CXW, YJOY and DDZ contributed to data collection and statistical analysis.
Ethics approval and consent to participate
This study was approved by the Ethics Review Committee of the Gansu Provincial Hospital of TCM, and all patients have provided their written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Sakuma M, Nakamura M, Yamada N, Ota S, Shirato K, Nakano T, Ito M, Kobayashi T. Venous thromboembolism: Deep vein thrombosis with pulmonary embolism, deep vein thrombosis alone and pulmonary embolism alone. Circ J. 2009;73:305–309. doi: 10.1253/circj.CJ-08-0372. [DOI] [PubMed] [Google Scholar]
- 2.Wang KL, Yap ES, Goto S, Zhang S, Siu CW, Chiang CE. The diagnosis and treatment of venous thromboembolism in asian patients. Thromb J. 2018;16:4. doi: 10.1186/s12959-017-0155-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hattab Y, Küng S, Fasanya A, Ma K, Singh AC, DuMont T. Deep venous thrombosis of the upper and lower extremity. Crit Care Nurs Q. 2017;40:230–236. doi: 10.1097/CNQ.0000000000000165. [DOI] [PubMed] [Google Scholar]
- 4.Hollenhorst MA, Battinelli EM. Thrombosis, Hypercoagulable states and anticoagulants. Prim Care. 2016;43:619–635. doi: 10.1016/j.pop.2016.07.001. [DOI] [PubMed] [Google Scholar]
- 5.Zhang Y, Xia H, Wang Y, Chen L, Li S, Hussein IA, Wu Y, Shang Y, Yao S, Du R. The rate of missed diagnosis of lower-limb DVT by ultrasound amounts to 50% or so in patients without symptoms of DVT: A meta-analysis. Medicine (Baltimore) 2019;98:e17103. doi: 10.1097/MD.0000000000017103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rabinovich A, Kahn SR. The postthrombotic syndrome: Current evidence and future challenges. J Thromb Haemost. 2017;15:230–241. doi: 10.1111/jth.13569. [DOI] [PubMed] [Google Scholar]
- 7.Heit JA, Spencer FA, White RH. The epidemiology of venous thromboembolism. J Thromb Thrombolysis. 2016;41:3–14. doi: 10.1007/s11239-015-1311-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kahn SR. The post-thrombotic syndrome. Hematology Am Soc Hematol Educ Program. 2016;2016:413–418. doi: 10.1182/asheducation-2016.1.413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stain M, Schonauer V, Minar E, Bialonczyk C, Hirschl M, Weltermann A, Kyrle PA, Eichinger S. The post-thrombotic syndrome: Risk factors and impact on the course of thrombotic disease. J Thromb Haemost. 2005;3:2671–2676. doi: 10.1111/j.1538-7836.2005.01648.x. [DOI] [PubMed] [Google Scholar]
- 10.Robert-Ebadi H, Righini M. Management of distal deep vein thrombosis. Thromb Res. 2017;149:48–55. doi: 10.1016/j.thromres.2016.11.009. [DOI] [PubMed] [Google Scholar]
- 11.Olaf M, Cooney R. Deep venous thrombosis. Emerg Med Clin North Am. 2017;35:743–770. doi: 10.1016/j.emc.2017.06.003. [DOI] [PubMed] [Google Scholar]
- 12.Mammen EF. Pathogenesis of venous thrombosis. Chest. 1992;102(Suppl):640S–644S. doi: 10.1378/chest.102.6.640S. [DOI] [PubMed] [Google Scholar]
- 13.Kirwan CC, McCollum CN, McDowell G, Byrne GJ. Investigation of proposed mechanisms of chemotherapy-induced venous thromboembolism: Endothelial cell activation and procoagulant release due to apoptosis. Clin Appl Thromb Hemost. 2015;21:420–427. doi: 10.1177/1076029615575071. [DOI] [PubMed] [Google Scholar]
- 14.Chen L, Wang J, Wang B, Yang J, Gong Z, Zhao X, Zhang C, Du K. MiR-126 inhibits vascular endothelial cell apoptosis through targeting PI3K/Akt signaling. Ann Hematol. 2016;95:365–374. doi: 10.1007/s00277-015-2567-9. [DOI] [PubMed] [Google Scholar]
- 15.Shyy JY, Chien S. Role of integrins in endothelial mechanosensing of shear stress. Circ Res. 2002;91:769–775. doi: 10.1161/01.RES.0000038487.19924.18. [DOI] [PubMed] [Google Scholar]
- 16.Bombeli T, Karsan A, Tait JF, Harlan JM. Apoptotic vascular endothelial cells become procoagulant. Blood. 1997;89:2429–2442. doi: 10.1182/blood.V89.7.2429. [DOI] [PubMed] [Google Scholar]
- 17.Mo J, Huang H, He F, et al. Influence of apoptosis signal pathway in traumatic deep vein thrombosis. J Kunming Med Univ. 2007;28:5–7. [Google Scholar]
- 18.Bartel DP. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 19.Hammond SM. An overview of microRNAs. Adv Drug Deliv Rev. 2015;87:3–14. doi: 10.1016/j.addr.2015.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Soifer HS, Rossi JJ, Saetrom P. MicroRNAs in disease and potential therapeutic applications. Mol Ther. 2017;15:2070–2079. doi: 10.1038/sj.mt.6300311. [DOI] [PubMed] [Google Scholar]
- 21.Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11:597–610. doi: 10.1038/nrg2843. [DOI] [PubMed] [Google Scholar]
- 22.O'Connell RM, Rao DS, Chaudhuri AA, Baltimore D. Physiological and pathological roles for microRNAs in the immune system. Nat Rev Immunol. 2010;10:111–122. doi: 10.1038/nri2708. [DOI] [PubMed] [Google Scholar]
- 23.Ten Cate H. MicroRNA and venous thrombosis. Thromb Haemost. 2016;116:205. doi: 10.1160/TH16-06-0473. [DOI] [PubMed] [Google Scholar]
- 24.Wang X, Sundquist K, Elf JL, Strandberg K, Svensson PJ, Hedelius A, Palmer K, Memon AA, Sundquist J, Zöller B. Diagnostic potential of plasma microRNA signatures in patients with deep-vein thrombosis. Thromb Haemost. 2016;116:328–336. doi: 10.1160/TH16-01-0071. [DOI] [PubMed] [Google Scholar]
- 25.Wang W, Zhu X, Du X, Xu A, Yuan X, Zhan Y, Liu M, Wang S. MiR-150 promotes angiogensis and proliferation of endothelial progenitor cells in deep venous thrombosis by targeting SRCIN1. Microvasc Res. 2019;123:35–41. doi: 10.1016/j.mvr.2018.10.003. [DOI] [PubMed] [Google Scholar]
- 26.Kong L, Hu N, Du X, Wang W, Chen H, Li W, Wei S, Zhuang H, Li X, Li C. Upregulation of miR-483-3p contributes to endothelial progenitor cells dysfunction in deepvein thrombosis patients via SRF. J Transl Med. 2016;14:23. doi: 10.1186/s12967-016-0775-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Qin J, Liang H, Shi D, Dai J, Xu Z, Chen D, Chen X, Jiang Q. A panel of microRNAs as a new biomarkers for the detection of deep vein thrombosis. J Thromb Thrombolysis. 2015;39:215–221. doi: 10.1007/s11239-014-1131-0. [DOI] [PubMed] [Google Scholar]
- 28.Yang R, Xing L, Zheng X, Sun Y, Wang X, Chen J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol Cancer. 2019;18:4. doi: 10.1186/s12943-018-0933-7. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 29.Zheng J, Xu T, Chen F, Zhang Y. MiRNA-195-5p functions as a tumor suppressor and a predictive of poor prognosis in non-small cell lung cancer by directly targeting CIAPIN1. Pathol Oncol Res. 2019;25:1181–1190. doi: 10.1007/s12253-018-0552-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li M, Ren CX, Zhang JM, Xin XY, Hua T, Wang HB, Wang HB. The Effects of miR-195-5p/MMP14 on proliferation and invasion of cervical carcinoma cells through TNF signaling pathway based on bioinformatics analysis of microarray profiling. Cell Physiol Biochem. 2018;50:1398–1413. doi: 10.1159/000494602. [DOI] [PubMed] [Google Scholar]
- 31.Kong F, Ma J, Yang H, Yang D, Wang C, Ma X. Long non-coding RNA PVT1 promotes malignancy in human endometrial carcinoma cells through negative regulation of miR-195-5p. Biochim Biophys Acta Mol Cell Res. 2018 Jul 19; doi: 10.1016/j.bbamcr.2018.07.008. doi: 10.1016/j.bbamcr.2018.07.008 (Epub ahead of print) [DOI] [PubMed] [Google Scholar]
- 32.Zhu N, Huang K, Liu Y, Zhang H, Lin E, Zeng Y, Li H, Xu Y, Cai B, Yuan Y, et al. miR-195-5p regulates hair follicle inductivity of dermal papilla cells by suppressing Wnt/β-catenin activation. Biomed Res Int. 2018;2018:4924356. doi: 10.1155/2018/4924356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhou G, Zhang X, Wang W, Zhang W, Wang H, Xin G. Both peripheral blood and urinary miR-195-5p, miR-192-3p, miR-328-5p and their target genes PPM1A, RAB1A and BRSK1 may be potential biomarkers for membranous nephropathy. Med Sci Monit. 2019;25:1903–1916. doi: 10.12659/MSM.913057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zeng Z, Yao J, Li Y, Xue Y, Zou Y, Shu Z, Jiao Z. Anti-apoptosis endothelial cell-secreted microRNA-195-5p promotes pulmonary arterial smooth muscle cell proliferation and migration in pulmonary arterial hypertension. J Cell Biochem. 2018;119:2144–2155. doi: 10.1002/jcb.26376. [DOI] [PubMed] [Google Scholar]
- 35.Sandrim VC, Dias MC, Bovolato AL, Tanus-Santos JE, Deffune E, Cavalli RC. Plasma from pre-eclamptic patients induces the expression of the anti-angiogenic miR-195-5p in endothelial cells. J Cell Mol Med. 2016;20:1198–1200. doi: 10.1111/jcmm.12767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Latronico MV, Catalucci D, Condorelli G. Emerging role of microRNAs in cardiovascular biology. Circ Res. 2007;101:1225–1236. doi: 10.1161/CIRCRESAHA.107.163147. [DOI] [PubMed] [Google Scholar]
- 37.Hirata H, Lopes GS, Jurkiewicz A, Garcez-do-Carmo L, Smaili SS. Bcl-2 modulates endoplasmic reticulum and mitochondrial calcium stores in PC12 cells. Neurochem Res. 2012;37:238–243. doi: 10.1007/s11064-011-0600-5. [DOI] [PubMed] [Google Scholar]
- 38.Naz R, Naz S, Mehboob M, Achakzai A, Khalid GH. Diagnostic yield of color Doppler ultrasonography in deep vein thrombosis. J Coll Physicians Surg Pak. 2005;15:276–279. [PubMed] [Google Scholar]
- 39.Bates SM, Jaeschke R, Stevens SM, Goodacre S, Wells PS, Stevenson MD, Kearon C, Schunemann HJ, Crowther M, Pauker SG, et al. Diagnosis of DVT: Antithrombotic therapy and prevention of thrombosis, 9th ed: American college of chest physicians evidence-based clinical practice guidelines. Chest. 2012;141(2 Suppl):e351S–e418S. doi: 10.1378/chest.11-2299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 41.Yang H, Meng Z, Zhang C, Zhang P, Wang Q. Establishing a new rat model of central venous sinus thrombosis and analyzing its pathophysiological and apoptotic changes. J Neurosci Methods. 2012;203:130–135. doi: 10.1016/j.jneumeth.2011.09.028. [DOI] [PubMed] [Google Scholar]
- 42.Cordes KR, Srivastava D. MicroRNA regulation of cardiovascular development. Circ Res. 2009;104:724–732. doi: 10.1161/CIRCRESAHA.108.192872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jin Y, Wang M, Hu H, Huang Q, Chen Y, Wang G. Overcoming stemness and chemoresistance in colorectal cancer through miR-195-5p-modulated inhibition of notch signaling. Int J Biol Macromol. 2018;117:445–453. doi: 10.1016/j.ijbiomac.2018.05.151. [DOI] [PubMed] [Google Scholar]
- 44.Chai L, Kang XJ, Sun ZZ, Zeng MF, Yu SR, Ding Y, Liang JQ, Li TT, Zhao J. MiR-497-5p, miR-195-5p and miR-455-3p function as tumor suppressors by targeting hTERT in melanoma A375 cells. Cancer Manag Res. 2018;10:989–1003. doi: 10.2147/CMAR.S163335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yang C, Wu K, Wang S, Wei G. Long non-coding RNA XIST promotes osteosarcoma progression by targeting YAP via miR-195-5p. J Cell Biochem. 2018;119:5646–5656. doi: 10.1002/jcb.26743. [DOI] [PubMed] [Google Scholar]
- 46.Cai C, He H, Duan X, Wu W, Mai Z, Zhang T, Fan J, Deng T, Zhong W, Liu Y, et al. miR-195 inhibits cell proliferation and angiogenesis in human prostate cancer by downregulating PRR11 expression. Oncol Rep. 2018;39:1658–1670. doi: 10.3892/or.2018.6240. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.