

Abstract
Although bioturbation is commonly recognized in contourites, only a few studies have analyzed the ichnological content of these deposits in detail. These studies have mainly focused on meso-scale bigradational sequence (a coarsening upward followed by a fining-upward sequence resulting from variations in current velocity). Here we present data from gravitational cores collected along the NW Iberian Margin showing systematic variation in ichnological content across proximal to distal depocenters within a large-scale elongated contourite drift. Data demonstrate that tracemakers’ behavior varies depending on the distance relative to the bottom current core. Trace fossils are already known to be a useful tool for studying of contouritic deposits and are even used as criterion for differentiating associated facies (e.g., turbidites, debrites), though not without controversy. We propose a mechanism by which the distance to the bottom current core exerts tangible influence on specific macro-benthic tracemaker communities in contourite deposits. This parameter itself reflects other bottom current features, such as hydrodynamic energy, grain size, nutrient transport, etc. Ichnological analysis can thus resolve cryptic features of contourite drift depositional settings.
Subject terms: Palaeontology, Sedimentology, Palaeoceanography
Introduction
The role of bottom currents in shaping deep-sea deposits (i.e., contourites) is currently a matter of debate in the scientific community1,2. Due to their implications for reconstruction of depositional conditions, contourite deposits have become a critical topic of investigation within the sub-disciplines of paleoceanography, slope-stability, and petroleum exploration. Due to their relative inaccessibility, however, contourites remain somewhat enigmatic. Only few studies have managed to investigate bioturbation and ichnofabrics in contour current settings3–6. Despite ongoing controversies7,8, trace fossil content is considered both a criterion for characterizing contouritic deposits and also a proxy for paleoenvironmental conditions1,5,7. These records are typically overprinted by bottom current activity9. In recent years, detailed ichnological studies conducted on contourite deposits in outcrops and core material have provided new insights into depositional processes, environmental conditions and the influence of bottom currents on tracemakers5,6,10,11. Due to lack of detailed records, the ichnological paradigm for contourites remains somewhat tentative. Here we describe unequivocal trace fossils in contouritic deposits from deep-sea gravity cores. Comparison of features reveals distinctive lateral variation with relative to bottom current cores.
The present study investigated core material collected from about 3,000 m water depth12 during the ForSaGal 09 research cruise around the Galicia Interior Basin (GIB; Fig. 1). The location is known to be affected by northward bottom currents that interact with bathymetry to generate a contouritic drift along the basin. This setting provides a detailed record of Quaternary contourite deposits13–16. Contouritic facies appear as massive to coarsely laminated silt to very fine silty sand and, show varying degrees of bioturbation17.
Figure 1.
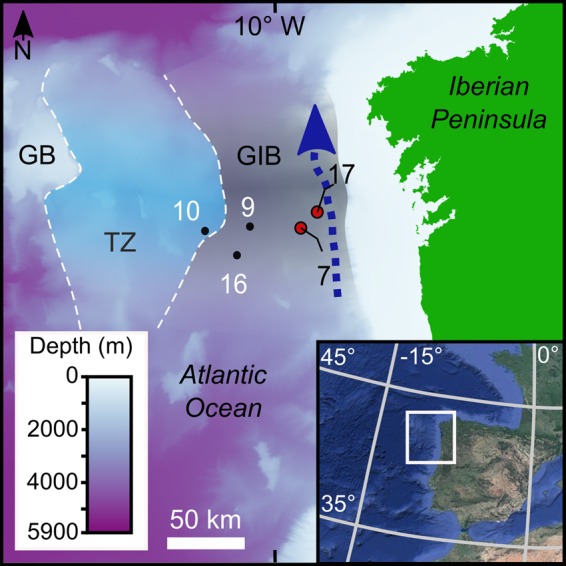
Location of cores analysed in this study. GB, Galicia Bank; GIB, Galicia Interior Basin; TZ, Transitional Zone. The arrow indicates the location of the bottom current (Mediterranean Outflow Water) during contourite deposition. The name of each gravity core sample begins with FSG09-. Inlay map from Google, Landsat/Copernicus. Globe image by Pixabay.
Ichnological content of contourites
Ichnological analysis of contouritic intervals from selected cores revealed an assemblage with relatively low diversity. In order of most to least dominant, contourite deposits contained Thalassinoides, Planolites, Palaeophycus, and Zoophycos (Fig. 2). Thalassinoides is defined as a 3D system of sub-horizontal burrows connected to the surface by sub-vertical shafts. Only limited sections of horizontal burrows appeared in cores analyzed by this study18,19. Burrows range from 4 to 18 mm in height and from 10 to 62 mm in length. Planolites are horizontal cylindrical tunnels, actively filled by the tracemaker20. These appear as sub-circular cross sections ranging from 3 to 15 mm in diameter. Palaeophycus are sub-horizontal cylindrical burrows characterized by passive filling and a lined wall20,21. These appear as lined sub-circular sections, 2–5 mm high and 4–19 mm long, filled by a darker sediment. Finally, Zoophycos is a complex helicoidal structure appearing as horizontal spreiten burrows in vertical sections of several cores22,23. The distribution of the ichnotaxa throughout the cores and their facies relationships (pelagic, hemipelagic, contourites and ice rafted debris or ‘IRD’) exhibit a clear pattern with contouritic intervals dominated by Thalassinoides and Planolites (Fig. 3). Abundant Palaeophycus or Zoophycos appear only occasionally (Fig. 3).
Figure 2.

Ichnotaxa identified within contourite intervals. Pa, Palaeophycus; Pl, Planolites; Th, Thalassinoides; Zo, Zoophycos. Scale bar 1 cm. Apparent lamination is an artefact produced during core slabbing.
Figure 3.

Sedimentological log and ichnologcial content of contourite deposits identified based on high-resolution images and CT data. BS: Bioturbated Surface.
Bioturbated surfaces vary between 0 and 32% within contouritic intervals with a mean value of 6.8%. This value corresponds to a low to moderate BI (0 to 3) (Fig. 3). Contouritic intervals exhibit only minor bioturbation but they show a variable percentage of bioturbated surface, depending on the site. Core material from site FSG09-17 showed less bioturbation (3.7% bioturbated surface average) than that collected from site FSG09-07 (10.8% average) (Fig. 3). Every analyzed contouritic interval showed noteworthy vertical differences in percentages of bioturbated surface and in ichnological composition from bottom to top. At site FSG09-07 both contouritic intervals show increased bioturbation, which exceeded 30% and consist mainly of Thalassinoides at the bottom and Zoophycos at the top. The contouritic intervals from site FSG09-17 likewise record an increase in the bioturbated surface due to abundant Palaeophycus (Fig. 3).
Ichnofabrics and paleoenvironmental conditions
The generally mottled background observed in contouritic core material reflects a complete reworking of the uppermost centimeters of the sediment. The degree of bioturbation in turn indicates relatively good environmental conditions for tracemakers working the unconsolidated substrate24,25. In this context, vertical changes in contourite ichnological features record significant variations in environmental parameters. Increasing percentages of bioturbated surface together with a shift in the dominant ichnotaxa reveal better conditions for particular tracemakers during the final stage of contourite deposition. Variation in Zoophycos in FSG09-07 and Palaeophycus in FSG09-17 indicates different ecological and depositional settings.
The organisms responsible for Palaeophycus constitute a eurybathic, facies-crossing ichnogenus, which occurs in a wide range of marine and non-marine settings. Palaeophycus itself represents a combined feeding/temporary dwelling burrow made by organisms with filter-, suspensive- and carnivore-feeding behaviors20,26. Their presence suggests high rates of organic matter transport to the sediment. Palaeophycus is a horizontal shallow burrow that is typically formed in shallow/middle tiers and remains open while the tracemakers live inside20. Dense occurrences of Palaeophycus tubularis, and the absence of other macroburrows, have been interpreted as evidence of ecologically stressful conditions during substrate colonization including salinity fluctuations, oxygen depletion, and turbidity. These features can also indicate a change in sedimentation rate or other conditions26.
Zoophycos is a deep tier structure with several ethological interpretations, but consensus favors the interpretation of cache behavior developed by vermiform animals27,28. A recent analysis describes relations between deep-marine Zoophycos, sedimentation rate, seasonal primary productivity, and oxygenation29. Accordingly, Zoophycos primarily appears in glacial periods with intensive seasonal productivity, reflecting high fluxes and intermediate sedimentation rates from 5 to 20 cm ka−1. Under these conditions, Zoophycos tracemakers collect nutrients at the sediment surface and transport them to deeper layers within the sediment to prevent oxidation29.
As outlined above, established interpretations of ichnological shifts attribute them to variations in sedimentation rate, organic matter availability, and oxygenation. Deposits from Galicia Interior Basin show clear spatiotemporal domains and associated sedimentation processes which themselves document the paleoenvironmental changes12. The westernmost FSG09-10 core was collected along the east flank of the Galicia Bank, in the so-called Transitional Zone30. This dome-like elevation is strongly influenced by bottom current activity that generates abraded surfaces31. Cores FSG09-09 and FSG09-16 were collected from the central part of the basin, between the Transitional Zone31 and lower slope12,14, which are dominated by pelagic and hemipelagic sedimentation12. Core FSG09-07, and especially the easternmost core FSG09-17, correspond to a contouritic and hemipelagic depositional setting developed on the lower continental slope14. Given their locations within the basin, sedimentation rate, organic matter availability and oxygenation vary with distance to the bottom current core.
Muddy contourites form due to the action of weak bottom currents that transport a substantial volume of organic matter particles5,32,33 which in turn feed benthic organisms34,35. Along the NW Iberian Margin for example, contour currents transport between 2–4 g m−3 of suspended material containing 40–100 mg m−3 of organic matter particles33. Distal zones to the core bottom current receive less sediment and organic matter than proximal settings, suggesting potential attendant variations in macrobenthic tracemaker communities. Site FSG09-17 (i.e., distal with respect to the bottom current core) experienced relatively low sedimentation rate and low organic matter flux. Organic matter at the sediment surface may be rapidly oxidized, preventing the development of shallow/middle tier structures. Under these conditions, only deep tier structures are produced, by organisms able to store organic matter deeper in the sediment as in the case of the Zoophycos tracemaker. Sediments from site FSG09-07 (i.e., the proximal site) indicate higher sedimentation rate and organic matter flux. Under these conditions, organic matter burial prevents oxidation and allows the development of shallow/middle tier dwelling structures, e.g. Palaeophycus (Fig. 4). The ichnological record, thus, systematically varies within the distal versus the proximal depositional zones,considering distance to core bottom current, of contourite drifts.
Figure 4.

Schematic diagrams of ichnological features that characterize distal and proximal, to core bottom current, settings within a muddy contourite drift. Pa, Palaeophycus; Pl, Planolites; Th, Thalassinoides; Zo, Zoophycos.
Conclusions
Sedimentation rate, oxygen conditions, and organic matter availability influence the macrobenthic tracemaker communities and resulting trace fossil assemblage in actively forming contourite drifts. This report provides novel evidence of systematic proximal, to core bottom current, versus distal variation in ichnological features from muddy contourites. Varying depositional conditions are interpreted to reflect distance from the bottom current core. Sedimentation rate and organic matter availability are higher in proximal areas where organic matter is rapidly buried. This prevents oxidation and makes organic matter available for shallow tier tracemakers (e.g. Palaeophycus producers). In distal settings, sedimentation rate and organic matter availability is lower. Organic matter is rapidly oxidized at the surface, favoring development of middle and deep tier tracemakers, which transport organic matter to deeper layers of the sediment (e.g., Zoophycos producers). Systematic lateral variation in ichnological content of contourite drifts demonstrates an impactful role for ichnological analysis in contourite research. Trace fossils not only could differentiate contourites from turbidites and associated deposits, they also record proximal to distal deposition within a contourite drift relative to core bottom current features.
Materials and Methods
Five gravity cores were collected during ForSaGal 09 in 2009 from the GIB, a narrow basin (around 100 km width; 2500–3000 m water depth) along the NW Iberian margin17. Sedimentary facies (i.e., Pelagic, Hemipelagic, Contouritic, Turbitidic and IRD layers) were previously characterized based on grain size, composition, sedimentary features and Computed Tomography (CT) scanning12,17. Contourite deposits, concretely muddy contourites, were identified in three cores. Only cores FSG09-07 (42.16°N, 9.84°W; 2393 mbsl) and FSG09-17 (42.26°N, 9.72°W; 2156 mbsl) (Fig. 1) contained a record long enough for detailed analysis. The present contribution used high resolution digital and CT images to perform ichnological analysis. Imaging steps used a CT scanner (HITACHI ECLOS 16 Multislice CT) at the Veterinary Teaching Hospital Rof Codina of Lugo (Galicia, Spain).
Ichnological analysis identified ichnotaxa based on ichnotaxabases (i.e., standard morphological features, wall, filling, etc.), tiering (i.e., vertical distribution of bioturbation structures within the sediment), crosscutting relations, and degree of bioturbation. The degree of bioturbation was determined by calculating the percentage of bioturbated surface using a quantification method based on high resolution digital images36, then calculated according to the Bioturbation Index scale37,38. These values reflect the spatial extent of discrete trace fossils identified over a common mottled background.
Age was calculated using the age model based on XRF data and AMS- 14C ages for the cores material analyzed12.
Acknowledgements
The contribution and research by JD was funded through the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 792314 (ICON-SE). The research of FJR-T was funded by project CGL2015-66835-P (Secretaría de Estado de Investigacion, Desarrollo e Innovacion, Spain), Research Group RNM-178 (Junta de Andalucía), and Scientific Excellence Unit UCE-2016-05 (Universidad de Granada). AM’s research is funded by the I2C program of the Xunta de Galicia Postdoctoral programme (ED481B 2016/029-0). The research was conducted as part of “The Drifters Research Group” (RHUL) and “Ichnology and Palaeoenvironment Research Group” (UGR) programs. We thank Prof. Grzegorz Pienkowski and Abdelouahed Lagnaoui for reviews and comments, Prof. Hernández-Molina for suggestions and Paul Kretchmer for editing the English.
Author contributions
J.D. and F.J.R.-T. designed the study and performed ichnological analysis and paleoenvironmental interpretations. A.M. and G.F. conducted sedimentary analysis and obtained high resolution and CT images. Finally, all authors discussed the results and contributed to the final version of the manuscript.
Data availability
All data analyzed during this study are summarized in this published article. The original datasets are not publicly available due to size restrictions but are available from the corresponding author by request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Rebesco M, Hernández-Molina FJ, Van Rooij D, Wåhlin A. Contourites and associated sediments controlled by deep-water circulation processes: State-of-the-art and future considerations. Mar. Geol. 2014;352:111–154. doi: 10.1016/j.margeo.2014.03.011. [DOI] [Google Scholar]
- 2.Shanmugam G. Contourites: Physical oceanography, process sedimentology, and petroleum geology. Petrol Explor Dev. 2007;44:183–216. doi: 10.1016/S1876-3804(17)30023-X. [DOI] [Google Scholar]
- 3.Fu S, Werner F. Distribution and Composition of Biogenic Structures on the Iceland-Faeroe Ridge: Relation to Different Environments. PALAIOS. 1994;9:92–101. doi: 10.2307/3515082. [DOI] [Google Scholar]
- 4.Baldwin CT, McCave IN. Bioturbation in an active deep-sea area; implications for models of trace fossil tiering. PALAIOS. 1999;14:375–388. doi: 10.2307/3515463. [DOI] [Google Scholar]
- 5.Wetzel, A., Werner, F. & Stow, D. A. V. Bioturbation and biogenic sedimentary structures in contourites. in Contourites (eds. Rebesco, M. & Camerlenghi, A.), 183–202 (2008).
- 6.Rodríguez-Tovar FJ, Hernández-Molina FJ, Hüneke H, Llave E, Stow D. Contourite facies model: Improving contourite characterization based on the ichnological analysis. Sediment. Geol. 2019;384:60–69. doi: 10.1016/j.sedgeo.2019.03.010. [DOI] [Google Scholar]
- 7.Rodríguez-Tovar FJ, Hernández-Molina FJ. Ichnological analysis of contourites: Past, present and future. Earth-Science Rev. 2018;182:28–41. doi: 10.1016/j.earscirev.2018.05.008. [DOI] [Google Scholar]
- 8.Shanmugam, G. Comment on “Ichnological analysis of contourites: Past, present and future” by Francisco J. Rodríguez-Tovar and F. Javier Hernández-Molina [Earth-Science Reviews, 182 (2018), 28–41]. Earth-Science Rev. 184, 46–49 (2018).
- 9.Bahr A, et al. Deciphering bottom current velocity and paleoclimate signals from contourite deposits in the Gulf of Cádiz during the last 140 kyr: An inorganic geochemical approach. Geochem Geophys. 2014;15:3145–3160. doi: 10.1002/2014GC005356. [DOI] [Google Scholar]
- 10.Alonso B, et al. Contourite vs gravity-flow deposits of the Pleistocene Faro Drift (Gulf of Cadiz): Sedimentological and mineralogical approaches. Mar. Geol. 2016;377:77–94. doi: 10.1016/j.margeo.2015.12.016. [DOI] [Google Scholar]
- 11.Reolid J, Betzler C. The ichnology of carbonate drifts. Sedimentology. 2018;66:1427–1448. doi: 10.1111/sed.12563. [DOI] [Google Scholar]
- 12.Mena A, et al. Evolution of the Galicia Interior Basin over the last 60 ka: sedimentary processes and palaeoceanographic implications. J. Quat. Sci. 2018;33:536–549. doi: 10.1002/jqs.3032. [DOI] [Google Scholar]
- 13.Hernández-Molina FJ, et al. Along-slope oceanographic processes and sedimentary products around the Iberian margin. Geo-Marine Lett. 2011;31:315–341. doi: 10.1007/s00367-011-0242-2. [DOI] [Google Scholar]
- 14.Bender VB, et al. Control of sediment supply, palaeoceanography and morphology on late Quaternary sediment dynamics at the Galician continental slope. Geo-Marine Lett. 2012;32:313–335. doi: 10.1007/s00367-012-0282-2. [DOI] [Google Scholar]
- 15.Hanebuth TJJ, Zhang W, Hofmann AL, Löwemark LA, Schwenk T. Oceanic density fronts steering bottom-current induced sedimentation deduced from a 50ka contourite-drift record and numerical modeling (off NW Spain) Quat. Sci. Rev. 2015;112:207–225. doi: 10.1016/j.quascirev.2015.01.027. [DOI] [Google Scholar]
- 16.Petrovic A, et al. Post-LGM upward shift of the Mediterranean Outflow Water recorded in a contourite drift off NW Spain. Mar. Geol. 2019;407:334–349. doi: 10.1016/j.margeo.2018.11.015. [DOI] [Google Scholar]
- 17.Mena A, et al. A novel sedimentological method based on CT-scanning: Use for tomographic characterization of the Galicia Interior Basin. Sediment. Geol. 2015;321:123–138. doi: 10.1016/j.sedgeo.2015.03.007. [DOI] [Google Scholar]
- 18.Frey RW, Curran A. h. & Pemberton, S. G. Tracemaking activities of crabs and their environmental significance: the ichnogenus Psilonichnus. J. Paleontol. 1984;58:511–528. [Google Scholar]
- 19.Bromley, R. G. Trace fossils. Biology, Taphonomy and Applications. (Chapman & Hall, 1996).
- 20.Pemberton SG, Frey RW. Trace fossil nomenclature and the Planolites-Palaeophycus dilemma. J. Paleontol. 1982;56:843–881. [Google Scholar]
- 21.Keighley DG, Pickerill RK. The ichnotaxa Palaeophycus and Planolites: historical perspectives and recommendations. Ichnos. 1995;3:301–309. doi: 10.1080/10420949509386400. [DOI] [Google Scholar]
- 22.Olivero, D. & Gaillard, C. A constructional model for Zoophycos. in Trace fossils: concepts, problems, prospects (ed. Miller, W. I.) 466–477 (Elsevier, 2007).
- 23.Zhang L, Fan R, Gong Y. Zoophycos macroevolution since 541 Ma. Sci. Rep. 2015;5:14954. doi: 10.1038/srep14954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ekdale, A. A., Bromley, R. G. & Pemberton, S. G. Ichnology: The Use of Trace Fossils in Sedimentology and Stratigraphy. Short Course. 15, (Society of Economics Paleontologists and Mineralogists, 1984).
- 25.Wetzel A, Uchman A. Hemipelagic and Pelagic Basin Plains. Trace Foss. as Indic. Sediment. Environ. Dev. Sedimentol. 2012;64:673–701. doi: 10.1016/B978-0-444-53813-0.00022-8. [DOI] [Google Scholar]
- 26.Villegas-Martín J, Netto RG. Permian macroburrows as microhabitats for meiofauna organisms: an ancient behaviour common in extant organisms. Lethaia. 2019;52:31–43. doi: 10.1111/let.12288. [DOI] [Google Scholar]
- 27.Wetzel A. Deep-sea ichnology: Observations in modern sediments to interpret fossil counterparts. Acta Geol. Pol. 2010;60:125–138. [Google Scholar]
- 28.Löwemark L. Testing ethological hypotheses of the trace fossil Zoophycos based on Quaternary material from the Greenland and Norwegian Seas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015;425:1–13. doi: 10.1016/j.palaeo.2015.02.025. [DOI] [Google Scholar]
- 29.Dorador J, Wetzel A, Rodríguez-Tovar FJ. Zoophycos in deep-sea sediments indicates high and seasonal primary productivity: Ichnology as a proxy in palaeoceanography during glacial–interglacial variations. Terra Nov. 2016;28:323–328. doi: 10.1111/ter.12224. [DOI] [Google Scholar]
- 30.Vázquez JT, et al. Cenozoic deformational structures on the Galicia Bank Region (NW Iberian continental margin) Mar. Geol. 2008;249:128–149. doi: 10.1016/j.margeo.2007.09.014. [DOI] [Google Scholar]
- 31.Ercilla G, et al. Imaging the recent sediment dynamics of the Galicia Bank region (Atlantic, NW Iberian Peninsula) Mar. Geophys. Res. 2011;32:99–126. doi: 10.1007/s11001-011-9129-x. [DOI] [Google Scholar]
- 32.McCave IN. Properties of suspended sediment over the HEBBLE area on the Nova Scotia rise. Mar. Geol. 1985;66:169–188. doi: 10.1016/0025-3227(85)90028-3. [DOI] [Google Scholar]
- 33.Thomsen L, Van Weering T, Gust G. Processes in the benthic boundary layer at the Iberian continental margin and their implication for carbon mineralization. Prog. Oceanogr. 2002;52:315–329. doi: 10.1016/S0079-6611(02)00013-7. [DOI] [Google Scholar]
- 34.Thistle D, Yingst JY, Fauchald K. A deep-sea benthic community exposed to strong nearbottom currents on the Scotian Rise (western. Atlantic). Mar. Geol. 1985;66:91–112. doi: 10.1016/0025-3227(85)90024-6. [DOI] [Google Scholar]
- 35.Lavaleye MSS, Duineveld GCA, Berghuis EM, Kok A, Witbaard R. A comparison between the megafauna communities on the N.W. Iberian and Celtic continental margins – effects of coastal upwelling? Prog. Oceanogr. 2002;52:459–476. doi: 10.1016/S0079-6611(02)00019-8. [DOI] [Google Scholar]
- 36.Dorador J, et al. Quantitative estimation of bioturbation based on digital image analysis. Mar. Geol. 2014;349:55–60. doi: 10.1016/j.margeo.2014.01.003. [DOI] [Google Scholar]
- 37.Reineck HE. Sedimentgefüge im Bereich der südliche Nordsee. Abhandlungen Senckenbergishen Naturforsche de Gesellschaft. 1963;505:1–138. [Google Scholar]
- 38.Taylor AM, Goldring R. Description and analysis of bioturbation and ichnofabric. J. Geol. Soc. London. 1993;150:141–148. doi: 10.1144/gsjgs.150.1.0141. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data analyzed during this study are summarized in this published article. The original datasets are not publicly available due to size restrictions but are available from the corresponding author by request.